
Abstract
Isolated patches resulting from habitat fragmentation can be surrounded by matrices with different permeabilities that can restrict dispersal and affect space use patterns. In this study, we examined the consequences of being isolated by matrices with different permeabilities on the space use patterns of the stone marten (Martes foina). We radio-tracked 41 martens at two study sites: a highly isolated site (HIS) in villages inside Białowieża Primeval Forest and a low-isolated site (LIS) in villages within a heterogenous landscape comprising a mosaic of agriculture and forest patches. Observations since 1991 documented the population as having a high proportion of males, which significantly declined after 2011. At both sites, stone martens used larger home ranges in spring-summer than in autumn-winter, and males had two to five times larger home ranges than females. Martens adopted two strategies of home range use—stationary or roaming. Roamer individuals only occurred at the HIS, had a 7-fold larger home range size, and moved farther between independent locations than stationary individuals. Roamers often switched between strategies and were in worse condition than stationary martens. Stationary martens used some of the smallest home ranges in Europe, with sizes similar to martens living in cities, while roamers had some of the largest, similar to stone martens inhabiting forests. Seasonal home ranges of stationary individuals did not differ between the study sites, but at the HIS, fidelity to home ranges was lower. Analyses of genetic relationship between individuals showed that dispersal distances from natal areas were shorter at the HIS. The study showed that stone martens exhibit great plasticity in space use.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Habitat fragmentation is a key issue in both population and conservation biology (Harrison and Bruna 1999; Chapman et al. 2007). The process that leads to habitat fragmentation has three other effects: an increase in the number of patches, a decrease in patch size, and an increase in the isolation of patches (Fahrig 2003). The classical theory of landscape fragmentation derives from concepts of landscapes in which suitable habitats (habitat islands/patches) are separated by uninhabitable matrices. Such conceptual frameworks generally suggest that the size, shape, and distribution of suitable habitat within a landscape (e.g., distance between patches, or their configuration) affect only the populations inhabiting particular patches (Forman 1981). Island biogeography and metapopulation theories have attached less attention to matrix type. The occupancy of patches also depends on the degree of their isolation and connectivity (Bollmann et al. 2011), but these factors are strongly related to the matrix separating patches (Kupfer et al. 2006). Recently, it has been recognized that the matrix has an important influence on biodiversity in human-modified landscapes (Franklin and Lindenmayer 2009). Therefore, understanding the ecological processes that affect populations inhabiting fragmented landscapes will not only require knowledge of the properties of habitat patches themselves, but also of the influence of the matrix between them. The type of matrix can potentially have positive or negative impacts on animal behavior and ecology (Kupfer et al. 2006). For example, the ecotone zone between patches and the matrix can increase biodiversity by increasing food abundance or shelter availability (e.g., Pozo-Montuy et al. 2011). On the other hand, low matrix permeability can decrease connectivity between patches, reduce gene flow, and create physical barriers that are never crossed by organisms (Cortes-Delgado and Perez-Torres 2011). Therefore, at the landscape level, permeability (or resistance) is one of the most important parameters of the matrix, and the isolation level of patches may vary in relation to the matrix type.
At the patch level, it is predicted that the first response of individuals to increases in patch isolation due to decreased matrix permeability is to change their behavior, including emigration attempts and altered space use (Grant and Hawley 1996). With increasing isolation, immigration and emigration is expected to decrease, as the probability of an individual successfully arriving at a patch decreases, and as individuals making dispersal forays return after failing to locate new patches (Kozakiewicz 1985). Therefore, in more isolated populations offspring should settle closer to their parents. In areas with low levels of emigration, higher densities force individuals to occupy smaller home ranges or territories and cause home ranges to overlap more often. Higher isolation and densities lead to more frequent contacts between individuals. This can lead to higher levels of aggression that can increase mortality in the population due to interference interactions (Linnell and Strand 2000). On the other hand, increased isolation and decreased dispersal between subpopulations increases the relatedness between individuals inhabiting a patch. In accordance with the kin-selection hypothesis (Hamilton 1963; Smith and Wynneedwards 1964), increased relatedness should increase tolerance between related individuals, which can lead to an increase in home range overlap (Ratnayeke et al. 2002; Hauver et al. 2010) and promote female natal philopatry that affects spatial organization (Rogers 1987; Ratnayeke et al. 2002; Zeyl et al. 2009).
In solitary carnivores, female home range size is related to food abundance, while male home range size and distribution is related to female spatial distribution; this is because males adjust their behavior to increase their encounter probability with females and enhance their reproduction success (Sandell 1989; Kovach and Powell 2003). Males have various strategies for achieving higher life reproductive output, depending on their age, body size, or social status, or the density of females (Sandell 1989; Kovach and Powell 2003; Zalewski 2012). First, males can increase their home range sizes by roaming over more extensive areas (roaming behavior) (e.g., Sandell 1989). A roaming strategy is often used by dominant males, which are usually larger and/or older. Large males encounter larger numbers of breeding females, thus paternity may be skewed towards a small number of the largest males (Kovach and Powell 2003). In contrast, subadult males may establish smaller home ranges than adult males, which greatly overlap with those of unrelated females to increase their chances of reproducing with at least one female (Zalewski 2012). These various strategies cause great variation in spacing patterns and home range sizes that are related to the conditions at various locations (Zalewski and Jędrzejewski 2006; Herr et al. 2009). For example, the home range sizes of Martes species have been found to vary from 0.5 to 30 km2 across their geographic ranges (e.g., Pulliainen 1984; Zalewski and Jędrzejewski 2006), and significant variations have also been observed in home range sizes within locations, suggesting that individuals can use different spacing pattern strategies (Lachat Feller 1993). On the other hand, martens have been found to have high fidelity to their home ranges, which suggests that spacing patterns are highly stable (O’Doherty et al. 1997; Zalewski and Jędrzejewski 2006). In relatively long-lived Martes, compared to smaller mustelids (e.g., weasels), a stable life home range and living in a known site (rather than roaming across unknown habitats) may positively affect survival and thus overall life reproduction output in males. The selection of this strategy may also be related to the degree to which a population is isolated by its habitat (especially as a result of matrix type), as isolation reduces opportunities for dispersal and increases relatedness between individuals.
The stone marten (Martes foina) is a medium-sized solitary carnivore (Herr 2008). It occurs across a large part of Europe except for the British Isles, Norway, Sweden, Finland, and northern Russia (Broekhuizen 1999; Proulx et al. 2004). In central and eastern Europe, the stone marten is primarily associated with human habitation, such as villages, small and medium towns, and even the centers of large cities (Bissonette and Broekhuizen 1995; Goszczyński et al. 2007). It willingly moves through open and ecotone areas, and small forest patches in agricultural landscapes, but avoids larger forest complexes (Pedrini et al. 1995; Goszczyński et al. 2007; Lanszki et al. 2009; Wereszczuk and Zalewski 2015; Wereszczuk et al. 2017). Habitat selection analyses have shown that stone marten in north-eastern Poland use only villages and cities (which create suitable habitats) and avoid other types of habitat (which create matrices separating suitable habitat; Wereszczuk and Zalewski 2015). However, gene flow between utilized sites varies due to the varying permeabilities of natural habitats (Wereszczuk et al. 2017). Dispersal and gene flow at the landscape scale is related to geographic distance, which is the only factor that influences the genetic structure of stone marten in Poland, as the isolation due to the matrix in populations inhabiting villages in a mosaic of agricultural habitat and small forest patches is low (Wereszczuk et al. 2017). In sites with high density of villages and high forest fragmentation, stone marten inhabits also small forest patches which often use artificial elements (e.g., rubble, dumps of rubbish, compost heaps, or culverts; Goszczyński et al. 2007). In contrast, large forest complexes, like Białowieża Forest, create barriers that decrease dispersal, and the populations inhabiting villages (patches) inside forest complexes are highly isolated (Wereszczuk et al. 2017). The different levels of isolation of villages surrounded by these two types of matrices with different permeability can affect stone marten population demography parameters, such as spacing and movement patterns, mortality, and offspring dispersal.
In this study, we compared two populations of stone marten inhabiting fragmented landscapes with different types of matrices between suitable patches. We expected that these matrices with different permeabilities influence the space use patterns and dispersal distances of stone martens. The aims of our study were to compare the following: (1) demography parameters (sex ratio, body size, and condition) between the two stone marten populations, (2) spacing patterns (home range size and overlap) between study sites with different matrix permeabilities, (3) stability of spacing patterns in time (home range fidelity), and (4) influence of matrix permeability on dispersal distance (distance between related and unrelated individuals). We expect that home range sizes will be smaller, fidelities higher, and dispersal distances shorter in the population highly isolated by the forest complex than in the less isolated population surrounded by a matrix with higher permeability, namely, a mosaic of agricultural and small forest patches.
Materials and methods
Study area
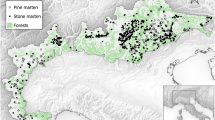
Data were collected at two study sites in north-eastern Poland: a highly isolated site (HIS) and a less isolated site (LIS) (Fig. 1). The HIS was located in villages within Białowieża Primeval Forest (BPF; 52.689722 N, 23.867222 E) and a small town outside. Here, the matrix separating suitable patches (villages) is a large forest complex (covering an area of 661 km2). BPF is a large primeval woodland whose main forest types are mixed conifer and ash-alder. Inside the forest are five villages of various areas (from the 24 ha built-up area of Podolany to the 218 ha of Białowieża), surrounded by meadows or the open valleys of small rivers and a large forest complex. The average distance between villages inside BPF is 4.9 km (range 1.0–8.9 km). The average distance from these villages to the dense forest is 0.6 km (range 0.2–1.3 km). Hajnówka is a town located outside BPF; it covers an area of 21 km2 and has 21,185 inhabitants, with a population density of 995 people per km2. The distance from the nearest village inside BPF to Hajnówka is 8 km, and the distance from Białowieża (the main study site) is 17 km.

Map of the study sites: low-isolated site (LIS) and highly isolated site (HIS) in north-eastern Poland
The LIS site (53.001388 N, 23.626388 E) was located in a heterogenous landscape comprising a mosaic of agricultural and forest patches with human settlements (area of 491 km2). It has four villages and a small town, as well as scattered farms and houses between the villages. The matrix separating the habitat suitable for martens (in this landscape villages and towns; Wereszczuk and Zalewski 2015) is a mosaic of fields, meadows, pastures, and small woodland patches. The average distance between villages in the LIS was almost the same as in the HIS—4.8 km (range 2.0–6.6 km). Also, at 0.7 km (range 0.3–1.5 km), the average distance between the villages and the forest was similar to that in the HIS. The small town, Michałowo, is located at the edge of the site; it covers an area of 2 km2 and has 3107 inhabitants, with a population density of 1445 people per km2.
Body size, body condition, and survival rate
All statistical analyses were performed in R versions 3.3.2 and 3.4.1 (R Development Core Team 2016). In most analyses, to obtain sets of models, we used the “dredge” function of the R package “MuMIn” (Bartoń 2012) and computed model support using Akaike’s information criterion with a correction for small sample size (AICc). We selected the model with the smallest ∆AICc as the best among all compared models; however, models within an ∆AICc of 2.00 were considered equally supported (Burnham and Anderson 2004).
To analyze demographic parameters (sex ratio, body size, and condition), we trapped stone martens (for the trapping procedure see below) and collected carcasses of stone martens killed in road traffic collisions and shot by hunters from 1991 to 2016. Both trapped martens and those found dead were sexed, aged, weighed, and measured (body length without the tail). A marten’s age was determined according to tooth wear, and two classes were distinguished: “juveniles” and “adults”, depending on whether individuals were younger or older than 1 year of age, respectively. Marten body condition was estimated as individual residuals from the linear regression model of body mass as predicted by body length, calculated separately for females and males. To analyze marten weight and condition, only data from captured individuals was used as the animals found dead could have affected these relations. To avoid biasing female body weight variation due to pregnancy, females sampled in March and April were excluded from the analyses. We fitted a linear model (package “stats”) to determine the effects of sex, study site, and space use strategy on stone marten body size and condition.
To evaluate overall survival rates for radio-tracked martens (tracking methods described below), we used a generalized linear modeling framework in the nest survival module of the program MARK (White and Burnham 1999) via R using the package “RMark” (Laake 2013). The nest survival model supports telemetry data, where individuals are tracked at irregular intervals, and frequency of monitoring varies throughout the year (Hupp et al. 2008; Blomberg et al. 2014). For each individual, we formatted the survival data into weekly encounter histories where we included the following: (1) the first week of observation as 1, (2) the last week the marten was known to be alive, (3) the last week the marten was checked, and (4) the fate of the marten. For martens that disappeared without evidence of mortality, the last week the marten was known to be alive was the same as the last week the marten was successfully checked. We included a global model that incorporated variation in survival between sexes and study sites.
Spatial pattern analyses
From September 1991 to October 2000, and from May 2011 to March 2016, 77 stone martens (43 females and 34 males) were captured in live traps baited with egg, chicken meat, or honey. Captured martens were anesthetized using 15 mg/kg ketamine and then sexed, aged, weighed, and measured (body length). Fifty-five individuals were fitted with radio collars (AVM, Lotek or ATS) that weighed 12–25 g, which was less than 2% of an individual’s weight. The life spans of the transmitters were 5–12 months. Martens were released at the site of capture after they recovered. All marten capture and handling procedures were approved by the Ministry of Environment and Local Ethics Committee for Animal Experiments at the University of Białystok (no: DL.gł-756/16/98; DL.gł-6713-21/35088/11/abr; DL.gł-6713-14/18806/11/abr; 2011/9).
The locations of radio-collared martens were determined by an observer on foot, one or two times per day (once during the day and/or once at night) at least four times per week. Bearings were obtained from a distance of < 500 m, usually about 200–300 m, and plotted on a 1:10,000 map. The coordinates were then transferred to ArcGIS 9.3 (Environmental Systems Research Institute, Redlands, California). In most cases, hand plotting on a map was more precise than triangulation, especially in the villages, where there were many easily distinguishable elements (e.g., houses, barns, roads). We used only one location within a minimum 2-h interval to reduce temporal autocorrelation among locations.
Spatial pattern analysis was based on a total 6247 locations (4895 from the HIS area and 1352 from the LIS area). Home range size was estimated using a minimum convex polygon with all the collected locations (MCP; Mohr 1947) and the kernel-density estimation method with 95% isopleths (KDE; Seaman and Powell 1996). We decided to use the fixed kernel method with the “cross-validated h” calculated from a least-square cross-validation approach as the bandwidth selection method. This is appropriate for a distribution type composed entirely of tight clumps of points, especially when animals spend most of their time visiting small patches (Gitzen et al. 2006). Home range analyses were performed with the “rhr” package in R (Signer and Balkenhol 2015). Home range size stabilized at 40–50 independent radiolocations per marten (Zalewski and Jędrzejewski 2006). We analyzed data in two seasons: spring-summer (SS) from 15 February to 14 September and autumn-winter (AW) from 15 September to 14 February. We combined data of home range size obtained from two periods (1991–2000 and 2011–2016) at HIS as the home range size in both periods was not significantly different.
We fitted generalized linear mixed-effects models (package “lme4”) to determine the effects of study site (HIS and LIS), season (SS and AW), sex, and the two-way interactions between them on life and seasonal home range sizes calculated by the MCP and KDE. In both sets of models, we used the family Gamma (link = identity). We used the number of fixes as a random effect to account for the non-independence of the number of fixes collected during the whole period of tracking for each individual (life home ranges). To account for non-independence between individuals in seasonal home ranges (e.g., an individual having a few seasonal home ranges), we incorporated the ID of individuals as a random effect in the model. We fitted all possible combinations of explanatory variables (5 candidate models for life home ranges and 17 for seasonal home ranges) and used the methods of model selection described above. We also measured the average distances between all consecutive independent locations for all tracked individuals and fitted a negative binomial generalized linear mixed-effects model (function “glmer.nb”, package “lme4”) due to the many zeros. In this analysis, we determined the effects of season and strategy with the ID of individuals as a random effect.
To assess stone marten home range fidelity, we used individuals that had been tracked for at least two seasons (HIS—15 individuals; LIS—4 individuals), and calculated the percentage overlap between two seasonal home ranges and the distances between the centers of these home ranges. The area of overlap was calculated in ArcMap 10.2.1 using the intersect function (Environmental Systems Research Institute, Redlands, California) on home ranges calculated by MCP. The estimated percentage overlap of an individual’s home range over two seasons was calculated in relation to the home range in the first season (White and Garrott 1990; O’Doherty et al. 1997). We also estimated the percentage overlap of home ranges between individuals at each study site. For this, we defined neighboring martens as those whose outermost range outlines overlapped. We estimated two percentage overlaps for each dyad.
To compare variation of home ranges in our study area with other locations in Europe, we collected the available literature on home range sizes of stone martens. We chose MCP 100% home ranges, usually counted for all locations (life home ranges). We divided study sites into three categories of habitat type: (1) forest, where the proportion of this habitat dominated across a study landscape; (2) rural, mainly comprising villages, farmlands, arable areas, and pastures, with only small forest patches; and (3) town or large city. We fitted a generalized linear model to determine the effects of sex, habitat type, longitude, and latitude on home range size across Europe. For this analysis, we used average life MCP home ranges of stationary individuals from both study sites.
Kin relationship
To compare dispersal distance between related (mother-offspring, father-offspring, and full siblings) and unrelated individuals at both sites, we collected tissue samples for DNA analysis from all trapped individuals and from carcasses of martens that died in car accidents or were shot by hunters from 2011 to 2016 (the period during which martens were studied at both sites). Tissue samples, a piece of skin or muscle, were kept frozen at − 20 °C until DNA extraction. We extracted DNA from tissue samples using a DNeasy Blood and Tissue Kit (Qiagen) according to the manufacturer’s instructions. Twenty-two microsatellite loci developed for martens were used to genotype all individuals; the details of the extraction and amplification procedures are described in Wereszczuk et al. (2017). We assessed the genetic relationship between individuals using the maximum-likelihood method implemented in COLONY v 2.0.6.4 (Jones and Wang 2010). We performed the analysis with the typing error rate set at 0.05. We assumed that both sexes were polygamous and that there was no inbreeding and used the full likelihood model with medium precision and information about sex but without information about the age of individuals. The distance between related and unrelated individuals was calculated as the Euclidean distance (in meters) between home range centroids (calculated as the mean of x and y coordinates) for radio-tracked individuals or the distance between the location of catching or finding for non-radio-tracked individuals. We tested whether geographic distances between pairs of individuals varied between related and unrelated pairs at both study sites (including interactions between both factors) using a generalized linear model (package "lme4") with family Gamma (log link) because of the positively skewed distribution resulting from the large number of unrelated individuals. We used the function “dredge” (package “MuMIn”) and AICc to identify the most parsimonious model in the candidate set (see the description of model selection above).
Results
Demography of population
From 1991 to 2016, we trapped or found dead 112 stone martens: 85 martens at the HIS and 27 at the LIS. Sex was not determined for 9 individuals. At the HIS, in the years 1991–2000, males constituted 64% (9 of 14 martens) of the population, in the years 2001–2010, 77% (10 of 13 martens), whereas in the years 2011–2016, only 37% (20 of 54 martens). At the LIS in the years 2011–2016, males also only comprised 31% (7 of 22) of stone martens capture or found dead. The mean weight of martens at the HIS was 1.32 ± 0.02 kg and at the LIS 1.29 ± 0.04 kg, and there were no significance differences between the study sites (β = 2.681, p = 0.959). The mean body mass of males (1.45 kg ± 0.03 SE, range: 1.12–1.93 kg) was greater than that of females (1.22 kg ± 0.01 SE, range 1.07–1.42 kg; β = 227.125, p < 0.001).
Thirteen radio-tracked martens were found dead (8 martens at the HIS and 5 at the LIS): five of them were killed by poachers, three were killed by a car, one was found in a stork (Ciconia ciconia) nest, and four were dead for unknown reasons. The top model of marten weekly survival rate included only the intercept, and the ΔAICc of the next competitive model was greater than 2. The survival of males and females at both sites was similar (weekly survival = 0.991, CI 0.98507–0.99457).
Home range size variation
For 41 individuals (28 from the HIS and 13 from the LIS area), we collected at least 40 locations and determined life home ranges. For 39 individuals (27 from the HIS and 12 from the LIS), we obtained 60 seasonal home ranges (44 from the HIS and 16 from the LIS) with at least 40 locations for each season (Table 1 and Appendix 1). The maximal life home ranges estimated using the MCP varied from 3.4 to 895.9 ha, and seasonal home ranges varied from 1.2 to 894.1 ha; the corresponding ranges estimated with KDE varied from 2.5 to 106.1 ha and from 0.5 to 93.7 ha, respectively (Table 1). The top model explaining variation in life MCP home range size (AICc weight = 0.860) included two variables (study site and sex), whereas that explaining variation in life KDE included only sex as a variable (AICc weight = 0.996; Table 2). Male home ranges were almost five times larger than those of females for MCP (182.0 ± 37.7 vs 45.9 ± 12.8 ha, respectively) and more than two times larger for KDE (25.9 ± 3.4 vs 11.4 ± 1.3 ha, respectively). Life home ranges were five times larger in the HIS than in the LIS for MCP (152.1 ± 29.7 vs 28.9 ± 8.2 ha, respectively), whereas for KDE, there were no differences between the study sites.
The top model for seasonal MCP home range size (AICc weight = 0.294) contained study site, season, sex, and a two-way interaction between study site and sex; the top model for seasonal KDE home ranges (AICc weight = 0.420) also contained study site, season, sex, and an interaction between study site and season (Table 2). Males had three (for MCP) or two (for KDE) times larger seasonal home ranges than females (MCP for males 109.3 ± 29.7 vs females 33.7 ± 11.6 ha; KDE for males 22.2 ± 3.7 vs females 8.1 ± 1.4 ha). Moreover, according to MCP calculations, both female and male seasonal home ranges at the HIS were twice the size of those at the LIS (females 43.3 ± 14.5 vs 14.4 ± 5.2 ha, males 127.7 ± 29.3 vs 40.8 ± 13.1 ha, respectively). At both sites, KDE home ranges were larger in spring-summer than in autumn-winter (Table 1). Inter and intrasexual overlap of seasonal home ranges varied between 2.4 and 22.9% (Table 3). At the HIS, intrasexual overlap was similar to intersexual overlap; however, at the LIS overlap between females was nine times smaller than between females and males (Table 3).
The fidelity of stone martens to their home ranges was estimated using two indexes: (1) the overlap between seasonal home ranges and (2) the distance between the arithmetic centers of seasonal home ranges. The average overlap between home ranges over consecutive seasons was 54.1% (± 5.8 SE; range 1.6–96.6). The best model explaining the overlap between seasonal home ranges included only the intercept (AICc weight = 0.61; Appendix S2). The top model analyzing variation in the distances between the centers of seasonal home ranges for each individual included study site and sex (AICc weight = 0.51; Appendix S2). The average distance between the arithmetic centers of seasonal home ranges was 184.6 ± 43.0 m (range 13.1–1051.0). The average shift between the centers of seasonal home ranges was more than three times larger at the HIS than at the LIS (216.0 ± 49.3 vs 58.8 ± 13.9 m) and was greater for males than females (251.2 ± 62.8 vs 84.6 ± 22.0 ha).
Strategies of space use
Based on seasonal home range size, we distinguished two groups of individuals with different home range utilization strategies: (1) stationary martens using KDE < 30 ha and (2) roamer martens using KDE > 30 ha (Fig. 2). Among all tracked martens, five males and one female from the HIS were classified as roamers, and roamer home ranges constituted 22.2% of all estimated seasonal home ranges: 27.3 and 30.0% of home ranges during spring-summer and autumn-winter, respectively. Stationary individuals showed high fidelity to their home range utilization strategy: 90% (N = 39) of individuals for which we estimated at least two seasonal home ranges did not change their home range use strategy in the subsequent season. However, among roamers (those for which we have at least two seasonal home ranges), all individuals changed their strategy from stationary to roamer or vice-versa (Table 4).

Stone marten (Martes foina) seasonal home range sizes (kernel-density estimation method with 95% isopleths) at the two study sites. We combined some of the final classes of home range size due to the large amount of scattering
Home range size variation was re-analyzed in the context of the two space use strategies. The seasonal home range sizes of roamers were seven times larger than those of stationary individuals (Table 5). Seasonal MCP home range sizes of roamers varied from 82.4 to 894.0 ha (average 441.1 ha), whereas for stationary individuals, they varied from 1.2 to 88.6 ha (overall average 26.7 ha; average for females 20.0 ha and males 33.7 ha). Life MCP home ranges of stationary individuals varied from 3.4 to 478.0 ha (average 46.2 ha; average for females 28.7 ha and for males 68.1 ha; Table 5). Distances between consecutive locations differed only between strategies, but not between seasons (AICc weight = 0.68; Appendix S3). In the HIS, the mean distance between consecutive independent locations was almost three times larger for roamers than stationary martens (454 ± 22 vs 160 ± 3 m). The top models analyzing home range fidelity, calculated as the distance between the centers of seasonal home ranges, included strategy, study site, and sex (AICc weight = 0.36), and the seasonal home range overlap model included only the intercept (AICc weight = 0.38; Appendix S2). Roamer individuals had lower home range fidelity than stationary martens; the shift between the centers of their seasonal home ranges was greater (349.1 ± 101.0 vs 143.4 ± 44.5 m) and the average seasonal home range overlap was lower (30.6% ± 16.4 vs 57.8% ± 5.7). Stationary individuals from the LIS had the same seasonal home range overlap as those from the HIS (53.9% ± 19.04 vs 59.1% ± 6.55), but shorter distances between the centers of their seasonal home ranges (58.8 m ± 20.8 vs 171.6 ± 58.5, respectively).
The body mass of males using either of the two strategies did not differ (Table 5). The top model explaining variation in the body condition index (AICc weight = 0.252; Appendix S4) included only the intercept, but the second competitive model was only 0.021 AICc units worse than the top model (AICc weight = 0.231; Appendix S4) and included home range use strategy. The condition index was lower in roamer compared to stationary martens (Table 5; mean condition index was − 64.0 ± 37.7 and 8.0 ± 11.7, respectively).
Home range size variation in Europe
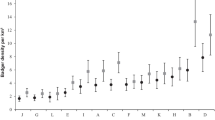
We summarized the available home range sizes of stone martens from 16 locations in Europe (Table 6). Home range sizes differed among sexes and habitat types across Europe and were not related to latitude or longitude (AICc weight = 0.495; Appendix S5). Male home ranges (averaged 170 ha; CI 120–239 ha) were two times larger than those of females (average 86 ha; CI 60–123 ha; Figs. 3). The largest home ranges were in forests (averaged 300 ha; CI 200–451 ha); medium in mosaics of farmlands with villages, arable lands, and small forest patches (averaged 110 ha; CI 73–165 ha); and smallest in towns (averaged 39 ha; CI 23–63 ha; Fig. 3).

Stone marten home range sizes of the two sexes in different habitat types averaged for the available data from Europe. Arithmetic means of life 100% minimum convex polygon home ranges of stationary individuals from the HIS and LIS sites are represented by dark stars, and roamer individuals from the HIS are represented by light stars

Estimated effect of the distances between pairs of stone marten individuals with different relationships, from the highly isolated (HIS) and low-isolated (LIS) sites in north-eastern Poland
Genetic relatedness
Twenty-one loci were genotyped for all martens, but three (Mer43, Gg454, Mvi057) were removed from further analyses due to low genetic variation (only 1–2 alleles per locus per site). The remaining 18 loci were polymorphic at both sites with a total number of alleles per locus ranging from 3 to 7. From 60 pairwise estimates of kinship, the following had probabilities above 0.7: 24 mother-offspring pairs (14 offspring of 6 mothers in the HIS and 10 offspring of 2 mothers in the LIS), 27 father-offspring pairs (23 offspring of 5 fathers in the HIS and 2 offspring of 1 father in the LIS), and 9 full siblings pairs (4 pairs in the HIS and 5 pairs in the LIS). Two additional father-offspring pairs were excluded from the analysis due to incorrect assignment that was verified on the basis of the age of individuals, year of their capture, and their fate. In terms of mother-offspring pairs at the HIS, two offspring each were assigned to four females and three offspring each to two females, while in the LIS, three and seven offspring were assigned to two females. In terms of father-offspring pairs at the HIS, 3 offspring were assigned to radio-tracked roamer male 19, 6 offspring to male 25 (which had stationary seasonal home ranges, but which shifted them over large distances), and 9 offspring to a male found dead due to a car accident.
Variation in the distances between the centroids of home ranges or places of capture/found individuals was explained by an interaction between genetic relationship and study site (AICc weight = 0.999; Appendix S6). The distances between mother-offspring and father-offspring were shorter at the HIS than at the LIS (0.9 vs 4.5 km and 1.5 vs 3.0 km, respectively) and at the HIS were shorter than for unrelated individuals, whereas at the LIS, they were similar to unrelated martens as the CI largely overlapped between these groups (Fig. 4). The distances between full siblings were larger at the HIS than at the LIS (0.3 vs 4.7 km); however, the small sample size and large variation produced a wide CI at the HIS (average 4.7 km; CI 1.4–16.0 km), which suggests that there was very high variation in dispersal distances among siblings (Fig. 4). The longest recorded distance between full siblings was 18 km, whereas between mother-offspring and father-offspring, it was 27 and 18 km, respectively.
Discussion
Our results demonstrated that higher levels of isolation due to an unfavorable matrix influence the space use patterns of stone marten. Our study sites were close together (22 km separate the northern border of the HIS from the southern border of the LIS), and the village structures were very similar: both sites had similar numbers of houses, stables, and gardens with fruit trees per unit area. Therefore, we should expect that stone martens inhabiting these sites have similar morphometric parameters and spacing patterns. Indeed, the body mass, survival, and proportion of males were similar at both sites. However, at the highly isolated site inside Białowieża Primeval Forest, individuals adopted two different home range utilization strategies. There were stationary individuals with small home ranges and roamers with much larger home ranges, and males using both strategies participated in reproduction. Stationary stone martens used similarly sized home ranges at both sites. The spacing pattern was highly unstable at the highly isolated site, probably due to the relatively frequent switching between the two strategies. The dispersal distance from the natal site was smaller at the highly isolated site than at the low-isolated site.
Stone marten home range sizes vary between sexes, seasons, and habitat types (Skirnisson 1986; Herrmann 1994; Powell 1994; Genovesi et al. 1997; Herr et al. 2009; Duduś 2014). In our extensive study, males utilized larger home ranges than females and larger home ranges in spring-summer (mating season) than in autumn-winter (non-mating season). Similar variations in home range sizes of males and females or between seasons have also been described in other stone marten populations in Europe (Skirnisson 1986; Herrmann 1994; Powell 1994; Genovesi et al. 1997; Herr et al. 2009; Duduś 2014). In Europe, stone marten have the largest home ranges at sites with mosaic habitat with higher proportions of forest, medium home ranges at sites with a mosaic of arable land and small forest patches, and the smallest in towns and cities (Table 6). As home range size is often related to food abundances (e.g., Krekorian 1976; Thompson and Colgan 1987), the variation in home range size among habitat types suggests that food availability is highest in urban areas and lowest in forests. Our study also showed that home range sizes (especially life) were different at both study sites; they were larger at the HIS than at the LIS, where the proportion of forest was higher. However, in our study, stone marten almost exclusively used developed areas (villages) and strongly avoided forest habitats (Wereszczuk and Zalewski 2015); therefore, the proportion of forest should not have affected the home range size of stone marten at either site.
Mean home ranges differed between the two sites due to the presence of individuals using the roamer strategy at the HIS, which had between 5 and 15 times larger home ranges (depending on the estimation method) than those of stationary individuals. The proportion of roamer home ranges was relatively high at the HIS (above 20%), as was the case in both seasons. The presence of roamer individuals at the HIS was most probably related to high isolation caused by the large forest complex. In other regions of Europe, stone marten have not shown such large variation in home range sizes at a single site and have probably not exhibited different space use strategies (e.g., Herrmann 1994; Herr et al. 2009; Duduś 2014). One exception is the Swiss Jura farmland where some stone marten individuals (three of eight) used much larger home ranges (range: 97–193 ha) than others (range: 20–57 ha) (Lachat Feller 1993). These individuals probably had a roaming strategy similar to that described in our study.
The roamer mating strategy has also been observed in other solitary carnivores, e.g., the stoat (Mustela ermine), mongoose (Herpestes ichneumon), and black bear (Ursus americanus), where dominant and/or larger males displayed roaming behavior and visited many females (Erlinge and Sandell 1986; Palomares 1993; Kovach and Powell 2003). In polygynous/promiscuous species, males increase in size with age, and younger males are generally outcompeted by older (larger) animals (Sandell 1986; Palomares 1993). However, roamer males in this study did not differ in body weight from stationary males, but they were in worse condition, suggesting that this strategy is energetically costly. In the black bear, encounter rates with breeding females were associated with body size but not home range size (Kovach and Powell 2003). In the badger, a social carnivore, 9% of males were found to use a roaming strategy with extremely large home ranges (Gaughran et al. 2018). This strategy could have been a mating tactic of dominant males to either gain access to many females, or a tactic for subordinate males, which are excluded from their natal range by dominant males; however, it was not possible to distinguish between these two possibilities (Gaughran et al. 2018). In our study, at least some of the males using the roamer strategy participated in reproduction and had at least one offspring, and five (of six) roamers were adults. Furthermore, martens using the roamer strategy adopted it throughout the year (regardless of whether it was mating season or not) and remained in a previously utilized area, as there was a 22.7% overlap between their seasonal home ranges. Interestingly, we observed one roamer female, who at first used a large area (KDE 45 ha) in spring-summer, and then in autumn-winter reduced her home range to only 2 ha.
The seasonal home ranges of stationary individuals did not differ between sites, despite the difference in isolation level. Furthermore, a comparison of home range size of stationary marten from our study (29 ha for females and 68 ha for males; life MCP) with those other studies showed that stone marten in NE Poland use very small home ranges that are comparable to home range sizes in towns across Europe (Table 6). The relatively small home ranges at our study sites suggest there is a rich and stable year-round food base available in the villages, where compost and garden fruits provide a food source comparable to human waste and garbage in towns. In contrast to what we expected, this may suggest that at our study sites, isolation does not lead to martens using smaller home ranges, because they already use very small areas. This has led to some individuals (especially males) adopting another strategy and utilizing large areas. Roamers had home range sizes (averaging 441 ha) similar to those found in poorer food base habitats (like forests or forest/rural areas).
Stone marten home range fidelity was relatively low (54.1%) compared with, for example, pine or American martens, for whom fidelity can reach up to 90% (Phillips et al. 1998; Zalewski and Jędrzejewski 2006). High fidelity to home ranges ensures stable access to resources, such as mates, food bases, and resting sites, and reduces predation risk. At the HIS, the seasonal home ranges of stationary individuals shifted much more than at the LIS, showing an unstable spacing pattern. The instability of the spacing pattern may be related to high mortality, as when an adjacent stationary individual dies, martens usually shift/enlarge their home ranges (Erlinge and Sandell 1986; Genovesi and Boitani 1995; Zalewski 2012; Gaughran et al. 2018). However, at both sites, the mortality rate of martens was similar, and thus this cannot explain the differences in home range shift between the sites. Our data may have been biased, as we tracked marten for a relatively short period relative to their lifespan (maximal 7 years). If intraspecific competition is one of the main primary selective agents driving the evolution of migration (Cox 1968), it is likely that a decrease in dispersal rate, as a result of isolation, can increase intraspecific competition and influence the stability of spacing patterns. Higher levels of aggressive interactions with neighboring martens may affect home range shift (Powell 1994) and may have led to the higher instability of spacing patterns at the HIS. The unstable home ranges of stationary individuals at the HIS may also have been due to the presence of roamers, which may modify home range utilization by stationary individuals. All roamers with at least two seasonal home ranges were observed switching strategies: two of four changed from stationary to roaming and the remaining two from roaming to stationary. This may also have contributed to the lower fidelity of stationary individuals by changing the arrangement of neighboring martens.
The large forest complex of BPF, an unfavorable matrix, constitutes a barrier for stone martens that reduces dispersal. The population inside BPF is strongly genetically different from both nearby and distant sites in Poland (Wereszczuk et al. 2017). However, in this study, we have shown that single individuals can successfully migrate from the villages inside the forest through BPF up to 18 km away. In the other type of matrix—a mosaic of fields, pastures, and small forests patches—stone marten migrated as far as 27 km. However, on average at the highly isolated site, offspring remained closer to their mothers than at the less isolated site. Similarly, short distance dispersal of subadult males away from natal home ranges has also been observed in pine martens (Zalewski 2012). Natal philopatry, which results in the spatial association of related individuals, is known to occur in many solitary species (Waser and Jones 1983). Higher natal philopatry at the highly isolated site may be the result of greater tolerance between related individuals.
The socio-spatial organization of stone martens is based on intrasexual territoriality (Herr et al. 2009; Newman et al. 2011); individuals of the same sex use separate areas but male home ranges overlap with those of females. Intraspecific tolerance of solitary carnivores can increase with increased food resources and/or relationships between individuals (Elbroch et al. 2016). In a solitary marmot, the rate of agonistic interactions decreased while the rate of amicable interactions increased with increasing relatedness, especially between mother-offspring and littermate sibling dyads (Maher 2009). However, the influence of increasing relatedness on the frequency of social interactions in solitary species is poorly documented (McEachern et al. 2007; Hirsch et al. 2013). For stone martens, highly abundant and stable food resources in villages may lead to higher intraspecific tolerance, especially with regard to related individuals. However, the number of simultaneously tracked individuals with overlapping home ranges was too low to analyze home range overlap in our study population, although we did observe a large tendency at the HIS for individuals of the same sex to have overlapping home ranges. The home range overlap between female-female and male-male dyads was equal to that of intersexual pairs. At the HIS, we also observed pairs of individuals foraging together in autumn-winter (Wereszczuk, unpublished data). The potential lack of non-occupied home ranges at the HIS may lead to a prolonged tolerance of offspring within the mother’s home range, and both male and female offspring have been found to stay close to the mother’s range (Zalewski 2012). Thus, social interactions are likely affected by strong philopatry rather than kin-based tolerance (Bartolommei et al. 2016).
Interestingly, the sex ratio changed over time, as in 1991–2000 males predominated in the HIS population (70%), whereas in 2011–2016 at both sites, they only constituted up to 37%. The unequal sex ratio, starting with an excess of males, may be related to the recent expansion of stone martens into north-eastern Poland (Wereszczuk et al. 2017). Białowieża Primeval Forest was probably colonized by stone martens in the early 1980s. A predominance of males is typical for declining populations (Erlinge 1983); however, it can also be indicative of an early phase of colonization, with low densities at the expansion front (Austerlitz et al. 1997; Excoffier et al. 2009). In addition, male martens travel further than females (Marchesi 1989; Zalewski et al. 2004), which could explain the higher proportion of males in areas located at the front of the expansion wave. Around 20–30 years after colonization, the proportion of males decreased to 31–37%. Similarly, low proportions of males have been observed in Germany and Luxemburg—which were 37 and 23% male, respectively (Herrmann 1994; Herr et al. 2009). At other locations at lower longitudes, the proportions of trapped males were higher, from 43 to 45% in Switzerland, up to 67% in Italy (Marchesi 1989; Lachat Feller 1993; Genovesi and Boitani 1995). The very low proportion of males in this study is unusual compared to other mustelids, in which the proportions of males among trapped individuals have been found to vary between 45 and 75% (Buskirk and Lindstedt 1989). A skewed sex ratio towards males is most probably a complex phenomenon related to sexual dimorphism in body size (Buskirk and Lindstedt 1989), but in our study, males were larger than females, and despite this males were trapped at lower proportions.
In conclusion, this study has shown that stone marten spacing patterns are highly plastic. Our findings suggest that the type of habitat forming the matrix and the level of isolation of a population can influence the space use of a solitary carnivore in a fragmented landscape. High levels of isolation force stone martens to adopt two strategies of home range use—stationary and roaming. Isolation by an impermeable matrix also results in shorter dispersal distances from natal home ranges and may promote greater tolerance of related individuals. The two space use strategies and low home range fidelity led to low stability of the spacing pattern in the highly isolated population. The low stability in socio-spatial organization, among other factors like inbreeding, may contribute to the higher risk of extinction in highly isolated, small populations of carnivores.
References
Austerlitz F, Jung-Muller B, Godelle B, Gouyon PH (1997) Evolution of coalescence times, genetic diversity and structure during colonization. Theor Popul Biol 51:148–164. https://doi.org/10.1006/tpbi.1997.1302
Bartolommei P, Gasperini S, Manzo E, Natali C, Ciofi C, Cozzolino R (2016) Genetic relatedness affects socio-spatial organization in a solitary carnivore, the European pine marten. Hystrix 27:2. https://doi.org/10.4404/hystrix-27.2-11876
Bartoń K (2012) MuMIn: multi-model inference. R package version 1.7.2, http://CRAN.R-project.org/package=MuMIn
Bissonette JA, Broekhuizen S (1995) Martes populations as indicators of habitat spatial patterns: the need for a multiscale approach. In: Lidicker WZ (ed) Landscape approaches in mammalian ecology and conservation. University of Minnesota Press. Minneapolis, London, pp 95–121
Blomberg EJ, Sedinger JS, Gibson D, Coates PS, Casazza ML (2014) Carryover effects and climatic conditions influence the postfledging survival of greater sage-grouse. Ecol Evol 4:4488–4499. https://doi.org/10.1002/ece3.1139
Bollmann K, Graf RF, Suter W (2011) Quantitative predictions for patch occupancy of capercaillie in fragmented habitats. Ecography 34:276–286. https://doi.org/10.1111/j.1600-0587.2010.06314.x
Broekhuizen S (1983) Habitat use of beech marten (Martes foina) in relation to landscape elements in a Dutch agricultural area. Proceedings from XVI Congress of the International Union of Game Biologist, ĆSSR, pp 614–624
Broekhuizen S (1999) Martes foina (Erxleben, 1777). In: Mittchell-Jones AJ, Amori G, Bogdanowicz W, Krystufek B, Reijnders PJH, Spitzenberger F, Stubbe M, Thissen JBM, Vohralik V, Zima J (eds) The atlas of European mammals. Academic Press, London, pp 342–343
Burnham KP, Anderson DR (2004) Multimodel inference—understanding AIC and BIC in model selection. Sociol Method Res 33:261–304. https://doi.org/10.1177/0049124104268644
Buskirk SW, Lindstedt SL (1989) Sex biases in trapped samples of Mustelidae. J Mammal 70:88–97. https://doi.org/10.2307/1381672
Chapman CA, Naughton-Treves L, Lawes MJ, Wasserman MD, Gillespie TR (2007) Population declines of colobus in Western Uganda and conservation value of forest fragments. Int J Primatol 28:513–528. https://doi.org/10.1007/s10764-007-9142-8
Cortes-Delgado N, Perez-Torres J (2011) Habitat edge context and the distribution of phyllostomid bats in the Andean forest and anthropogenic matrix in the Central Andes of Colombia. Biodivers Conserv 20:987–999. https://doi.org/10.1007/s10531-011-0008-1
Cox GW (1968) Role of competition in evolution of migration. Evolution 22:180–192. https://doi.org/10.2307/2406662
R Development Core Team (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/
Duduś L (2014) Biologia kuny domowej (Martes foina Erxleben, 1777) we Wrocławiu. PhD thesis. Instytutu Ochrony Przyrody PAN, Wrocław, Poland
Elbroch LM, Lendrum PE, Quigley H, Caragiulo A (2016) Spatial overlap in a solitary carnivore: support for the land tenure, kinship or resource dispersion hypotheses? J Anim Ecol 85:487–496. https://doi.org/10.1111/1365-2656.12447
Erlinge S (1983) Demography and dynamics of a stoat Mustela erminea population in a diverse community of vertebrates. J Anim Ecol 52:705–726. https://doi.org/10.2307/4449
Erlinge S, Sandell M (1986) Seasonal changes in the social organization of male stoats, Mustela erminea: an effect of shifts between two decisive resources. Oikos 47:57–62. https://doi.org/10.2307/3565919
Excoffier L, Foll M, Petit RJ (2009) Genetic consequences of range expansions. Annu Rev Ecol Evol S 40:481–501. https://doi.org/10.1146/annurev.ecolsys.39.110707.173414
Fahrig L (2003) Effects of habitat fragmentation on biodiversity. Annu Rev Ecol Evol S 34:487–515. https://doi.org/10.1146/annurev.ecolsys.34.011802.132419
Forman P (1981) Genesis of relativity—Einstein in context - Swenson, LS. Ann Sci 38:370–371
Franklin JF, Lindenmayer DB (2009) Importance of matrix habitats in maintaining biological diversity. P Natl Acad Sci USA 106:349–350. https://doi.org/10.1073/pnas.0812016105
Gaughran A, Kelly DJ, MacWhite T, Mullen E, Maher P, Good M, Marples NM (2018) Super-ranging. A new ranging strategy in European badgers. PLoS One 13:16. https://doi.org/10.1371/journal.pone.0191818
Genovesi P, Boitani L (1995) Preliminary data on the social ecology of the stone marten (Martes foina Erxleben 1777) in Tuscany (Central Italy). Hystrix 7:159–163
Genovesi P, Sinibaldi I, Boitani L (1997) Spacing patterns and territoriality of the stone marten. Can J Zool 75:1966–1971. https://doi.org/10.1139/z97-828
Gitzen RA, Millspaugh JJ, Kernohan BJ (2006) Bandwidth selection for fixed-kernel analysis of animal utilization distributions. J Wildlife Manage 70:1334–1344. https://doi.org/10.2193/0022-541x(2006)70[1334:bsffao]2.0.co;2
Goszczyński J, Posłuszny M, Pilot M, Gralak B (2007) Patterns of winter locomotion and foraging in two sympatric marten species: Martes martes and Martes foina. Can J Zool 85:239–249. https://doi.org/10.1139/z06-212
Grant J, Hawley A (1996) Some observations on the mating behaviour of captive American pine martens Martes americana. Acta Theriol 41:439–442. https://doi.org/10.4098/AT.arch.96-45
Hamilton WD (1963) Evolution of altruistic behavior. Am Nat 97:354–356. https://doi.org/10.1086/497114
Harrison S, Bruna E (1999) Habitat fragmentation and large-scale conservation: what do we know for sure? Ecography 22:225–232. https://doi.org/10.1111/j.1600-0587.1999.tb00496.x
Hauver SA, Gehrt SD, Prange S, Dubach J (2010) Behavioral and genetic aspects of the raccoon mating system. J Mammal 91:749–757. https://doi.org/10.1644/09-mamm-a-067.1
Herr J (2008) Ecology and behaviour of urban stone martens (Martes foina) in Luxembourg. PhD thesis, University of Sussex, Sussex, UK
Herr J, Schley L, Roper TJ (2009) Socio-spatial organization of urban stone martens. J Zool 277:54–62. https://doi.org/10.1111/j.1469-7998.2008.00510.x
Herrmann M (1994) Habitat use and spatial organization by the stone marten. In: Buskirk SW, Harestad AS, Raphael MG, Powell RA (eds) Martens, sables, and fishers: biology and conservation. Cornell University Press, Ithaca, pp 122–136
Hirsch BT, Prange S, Hauver SA, Gehrt SD (2013) Genetic relatedness does not predict racoon social network structure. Anim Behav 85:463–470. https://doi.org/10.1016/j.anbehav.2012.12.011
Hupp JW, Schmutz JA, Ely CR (2008) Seasonal survival of radiomarked emperor geese in western Alaska. J Wildlife Manage 72:1584–1595. https://doi.org/10.2193/2007-358
Jones OR, Wang JL (2010) COLONY: a program for parentage and sibship inference from multilocus genotype data. Mol Ecol Resour 10:551–555. https://doi.org/10.1111/j.1755-0998.2009.02787.x
Kovach AI, Powell RA (2003) Effects of body size on male mating tactics and paternity in black bears, Ursus americanus. Can J Zool 81:1257–1268. https://doi.org/10.1139/z03-111
Kozakiewicz M (1985) The role of habitat isolation in formation of structure and dynamics of the bank vole population. Acta Theriol 30:193–209. https://doi.org/10.4098/AT.arch.85-12
Krekorian OC (1976) Home range size and overlap and their relationship to food abundance in the desert iguana, Dipsosaurus dorsalis. Herpetologica 32:405–412
Kruger H-H (1990) Home-range and patterns of distribution of stone and pine martens. Trans 19th IUGB Congress, Trondheim 1989, Norway, pp 348–349
Kupfer JA, Malanson GP, Franklin SB (2006) Not seeing the ocean for the islands: the mediating influence of matrix-based processes on forest fragmentation effects. Glob Ecol Biogeogr 15:8–20. https://doi.org/10.1111/j.1466-822x.2006.00204.x
Laake JL (2013) RMark: an R interface for analysis of capture-recapture data with MARK. AFSC Processed Rep 2013–01. Alaska Fisheries Science Center, National Marine Fisheries Service, US Department of Commerce, Seattle, USA
Lachat Feller N (1993) Eco- éthologie de la fouine (Martes foina Erxleben, 1777) dans le Jura suisse. PhD thesis, Université de Neuchâtel, Neuchâtel, Switzerland
Lanszki J, Sardi B, Szeles GL (2009) Feeding habits of the stone marten (Martes foina) in villages and farms in Hungary. Natura Somogyiensis 15:231–246
Linnell JDC, Strand O (2000) Interference interactions, co-existence and conservation of mammalian carnivores. Divers Distrib 6:169–176. https://doi.org/10.1046/j.1472-4642.2000.00069.x
Lopez-Martin JM, Ruiz-Olmo J, Cahill S (1992) Autumn home range and activity of a stone marten (Martes foina Erxleben, 1777) in northeastern Spain. Miscellania Zoologica (Barcelona) 16:258–260
Maher CR (2009) Effects of relatedness on social interaction rates in a solitary marmot. Anim Behav 78:925–933. https://doi.org/10.1016/j.anbehav.2009.06.027
Marchesi P (1989) Ecologie et comportament de la Martre (Martes martes L.) dans le Jura suisse. PhD thesis, Université de Neuchâtel, Neuchâtel, Switzerland
McEachern MB, Eadie JM, Van Vuren DH, Ecol Graduate G (2007) Local genetic structure and relatedness in a solitary mammal, Neotoma fuscipes. Behav Ecol Sociobiol 61:1459–1469. https://doi.org/10.1007/s00265-007-0378-2
Mohr CO (1947) Table of equivalent populations of north American small mammals. Am Midl Nat 37:223–249. https://doi.org/10.2307/2421652
Müskens GJDM, Broekhuizen S (2005) De steenmarter (Martes foina) in Borgharen: aantal, overlast en schade. Report no. 1259, Wageningen, Alterra, The Netherland
Newman C, Zhou YB, Buesching CD, Kaneko Y, Macdonald DW (2011) Contrasting sociality in two widespread, generalist, mustelid genera, Meles and Martes. Mamm Study 36:169–188. https://doi.org/10.3106/041.036.0401
O'Doherty EC, Ruggiero LF, Henry SE (1997) Home-range size and fidelity of American martens in the Rocky Mountains of southern Wyoming. In: Proulx G, Bryant HN, Woodard PM (eds) Martens: taxonomy, ecology, techniques and management. The Provincial Museum of Alberta, pp 123–134
Palomares F (1993) Individual variations of male mating tactics in Egyptian mongooses (Herpestes ichneumon)—can body mass explain the differences. Mammalia 57:317–324. https://doi.org/10.1515/mamm.1993.57.3.317
Pedrini P, Prigioni C, Volcan G (1995) Distribution of mustelids in Adamello-Brenta Park and surrounding areas (central Italian Alps). Hystrix 7:39–44
Phillips DM, Harrison DJ, Payer DC (1998) Seasonal changes in home-range area and fidelity of martens. J Mammal 79:180–190. https://doi.org/10.2307/1382853
Powell RA (1994) Structure and spacing of Martes populations. In: Buskirk S, Harestad AS, Raphael MG, Powell RA (eds) Martens, sables and fishers—biology and conservation. Cornell University Press, Ithaca
Pozo-Montuy G, Serio-Silva JC, Bonilla-Sanchez YM (2011) Influence of the landscape matrix on the abundance of arboreal primates in fragmented landscapes. Primates 52:139–147. https://doi.org/10.1007/s10329-010-0231-5
Proulx G, Aubry K, Birks J, Buskirk S, Fortin C, Frost H, Krohn W, Mayo L, Monakhov V, Payer D, Saeki M, Santos-Reis M, Weir R, Zielinski W (2004) World distribution and status of the genus Martes in 2000. In: Harrison DJ, Fuller AK, Proulx G (eds) Martens and fishers (Martes) in human-altered environments: an international perspective. Springer, New York, pp 21–76
Pulliainen E (1984) Use of the home range by pine martens (Martes martes). Acta Zool Fenn 171:271–274
Ratnayeke S, Tuskan GA, Pelton MR (2002) Genetic relatedness and female spatial organization in a solitary carnivore, the raccoon, Procyon lotor. Mol Ecol 11:1115–1124. https://doi.org/10.1046/j.1365-294X.2002.01505.x
Rogers LL (1987) Effects of food supply and kinship on social behavior, movements, and population growth of black bears in northeastern Minnesota. Wildlife Monogr 97:1–72
Rondinini C, Boitani L (2002) Habitat use by beech martens in a fragmented landscape. Ecography 25:257–264. https://doi.org/10.1034/j.1600-0587.2002.250301.x
Sandell M (1986) Movement patterns of male stoats Mustela erminea during the mating season—differences in relation to social status. Oikos 47:63–70. https://doi.org/10.2307/3565920
Sandell M (1989) The mating tactics and spacing patterns of solitary carnivores. In: Gittleman JL (ed) Carnivore behavior, ecology, and evolution. Chapman & Hall, London & Cornell University Press, New York, pp 164–182
Santos-Reis M, Santos MJ, Lourenco S, Marques JT, Pereira I, Pinto B (2004) Relationships between stone martens, genets and cork oak woodlands in Portugal. In: Harrison DJ, Fuller AK, Proulx G (eds) Martens and fishers (Martes) in human-altered environments: an international perspective. Springer, New York, pp 147–172
Seaman DE, Powell RA (1996) An evaluation of the accuracy of kernel density estimators for home range analysis. Ecology 77:2075–2085. https://doi.org/10.2307/2265701
Signer J, Balkenhol N (2015) Reproducible home ranges (rhr): a new, user-friendly R package for analyses of wildlife telemetry data. Wildl Soc Bull 39:358–363. https://doi.org/10.1002/wsb.539
Simon O, Lang J (2007) Mit Hühnerei auf Marderfang—Methode und Fangerfolge in den Wäldern von Frankfurt. Nat Mus 137:1–11
Skirnisson K (1986) Untersuchungen zum Raum-Zeit-System freilebender Steinmarder (Martes foina Erxleben, 1777). Beitr Wildbiol 6:1–200
Smith JM, Wynneedwards VC (1964) Group selection and kin selection. Nature 201:1145–1147. https://doi.org/10.1038/2011145a0
Thompson ID, Colgan PW (1987) Numerical responses of martens to a food shortage in northcentral Ontario. J Wildlife Manage 51:824–835. https://doi.org/10.2307/3801748
Waser PM, Jones WT (1983) Natal philopatry among solitary mammals. Q Rev Biol 58:355–390. https://doi.org/10.1086/413385
Wereszczuk A, Zalewski A (2015) Spatial niche segregation of sympatric stone marten and pine marten—avoidance of competition or selection of optimal habitat? PLoS One 10:e0139852. https://doi.org/10.1371/journal.pone.0139852
Wereszczuk A, Leblois R, Zalewski A (2017) Genetic diversity and structure related to expansion history and habitat isolation: stone marten populating rural-urban habitats. BMC Ecol 17:16. https://doi.org/10.1186/s12898-017-0156-6
White GC, Burnham KP (1999) Program MARK: survival estimation from populations of marked animals. Bird Study 46:120–139
White GC, Garrott RA (1990) Analysis of wildlife radio-tracking data. Academic Press
Wierzbowska IA, Lesiak M, Zalewski A, Gajda A, Widera E, Okarma H (2017) Urban carnivores: a case study of sympatric stone marten (Martes foina) and red fox (Vulpes vulpes) in Krakow, southern Poland. In: Zalewski A, Wierzbowska IA, Aubry KB, Birks JDS, O’Mahony DT, Proulx G (eds) The Martes Complex in the 21st century: ecology and conservation. Mammal Research Institute Polish Academy of Sciences, Białowieża, pp 161–178
Zalewski A (2012) Alternative strategies in the acquisition of home ranges by male pine martens in a high-density population. Acta Theriol 57:371–375. https://doi.org/10.1007/s13364-012-0086-9
Zalewski A, Jędrzejewski W (2006) Spatial organisation and dynamics of the pine marten Martes martes population in Białowieża Forest (E Poland) compared with other European woodlands. Ecography 29:31–43. https://doi.org/10.1111/j.2005.0906-7590.04313.x
Zalewski A, Jędrzejewski W, Jędrzejewska B (2004) Mobility and home range use by pine martens (Martes martes) in a Polish primeval forest. Ecoscience 11:113–122. https://doi.org/10.1080/11956860.2004.11682815
Zeyl E, Aars J, Ehrich D, Wiig O (2009) Families in space: relatedness in the Barents Sea population of polar bears (Ursus maritimus). Mol Ecol 18:735–749. https://doi.org/10.1111/j.1365-294X.2008.04049.x
Acknowledgements
We are grateful to E. Bujko for help in the catching and tracking of stone martens, as well as to J. Metseelar and other students, J. Bullard, G. Butrykowska, C. Dalglish, S. Flack, T. Green, J. Matuszczyk, A. Morrice, M. Needle, C. O’Brien-Moran, K. O’Mahony, J. Połocka, T. Pritchard, D. Williams, and K. Żurek for their help with field work. We thank J. Chilecki, T. Kamiński, M. Kołodziej-Sobocińska, R. Kowalczyk, T. Tumiel, and M. Wereszczuk for their help in collecting samples for genetic analysis. We are especially grateful to H. Zalewska for her laboratory work with genetic samples and to T. Borowik for statistical support. We wish to sincerely thank T. Diserens and A. Michalak for correcting the English in the manuscript.
Funding
Research was funded by National Science Centre Poland, (grant number UMO 2011/01/NZ/NZ8/04525).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Karol Zub
Electronic Supplementary Material
ESM 1
(DOCX 22 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Wereszczuk, A., Zalewski, A. Does the matrix matter? Home range sizes and space use strategies in stone marten at sites with differing degrees of isolation. Mamm Res 64, 71–85 (2019). https://doi.org/10.1007/s13364-018-0397-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-018-0397-6