
Abstract
The analysis of glycosylation from native biological sources is often frustrated by the low abundances of available material. Here, ion mobility combined with electrospray ionization mass spectrometry have been used to extract the spectra of N-glycans released with PNGase F from a serial titration of recombinantly expressed envelope glycoprotein, gp120, from the human immunodeficiency virus (HIV). Analysis was also performed on gp120 expressed in the α-mannosidase inhibitor, and in a matched mammalian cell line deficient in GlcNAc transferase I. Without ion mobility separation, ESI spectra frequently contained no observable ions from the glycans whereas ions from other compounds such as detergents and residual buffer salts were abundant. After ion mobility separation on a Waters T-wave ion mobility mass spectrometer, the N-glycans fell into a unique region of the ion mobility/m/z plot allowing their profiles to be extracted with good signal:noise ratios. This method allowed N-glycan profiles to be extracted from crude incubation mixtures with no clean-up even in the presence of surfactants such as NP40. Furthermore, this technique allowed clear profiles to be obtained from sub-microgram amounts of glycoprotein. Glycan profiles were similar to those generated by MALDI-TOF MS although they were more susceptible to double charging and fragmentation. Structural analysis could be accomplished by MS/MS experiments in either positive or negative ion mode but negative ion mode gave the most informative spectra and provided a reliable approach to the analysis of glycans from small amounts of glycoprotein.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Although mass spectrometry, particularly with matrix-assisted laser desorption/ionization (MALDI) and electrospray (ESI) ionization, has made much impact on the analysis of carbohydrates in recent years, many samples presented for analysis fail to give signals due to the low amount of carbohydrate and to the presence of contaminants that are difficult to remove by conventional techniques. These difficulties are evidenced by the range of techniques developed for removing contaminants including, for example, the use of various types of resin and other adsorbing materials [1–4], porous graphitized carbon [5–7], various membranes [8, 9], lectins [10, 11], immobilized boronic acids [10, 12], and drop dialysis [13]. In our laboratory, Nafion 117 [8] and other membranes are routinely used but, even though these sometimes give spectacular improvements in signals [14], on other occasions they make little or no difference to the detection of glycans. Recently, we have been evaluating ion mobility mass spectrometry as an analytical system for carbohydrates [15], particularly N-linked glycans, and have found that this technique is excellent for removing signals from contaminating compounds allowing ions from the glycans to be clearly seen even when no evidence of their presence is visible in the parent spectrum.
Several investigators have explored various aspects of ion mobility mass spectrometry for studies of carbohydrates. For example, Fenn and McLean [16–18] have shown that compounds of different structural types such as lipids, peptides and carbohydrates can be separated and, recently, this group has applied the technique to examination of N-linked glycans released from ribonuclease B [18]. Ion mobility separation coupled with principal component analysis of N-linked glycans from serum glycoproteins has shown promise for detection of cancer biomarkers [19] and ion mobility separations have also been used to examine N-linked glycan heterogeniety from monoclonal antibodies [20, 21] although, in these cases, glycopeptides rather than the released glycans were examined. Isomer separation of small carbohydrates has been demonstrated [22–27] but, with current technology, the resolution of the mobility cells is not sufficient to resolve any but the smaller N-glycans [15, 28]. For a review on instrumentation see [29].
Previous work from this laboratory with IgG and dextran oligomers (manuscript in preparation) has shown good ion mobility separation of singly charged N-linked glycans from peptides, buffer salts and multiply-charged ions. Some separation was also found between the N-glycans and other carbohydrate types such as dextran oligomers. In the present work, the utility of the ion mobility technique to extract glycan profiles from contaminated samples and directly from incubation mixtures obtained from glycan release experiments has been explored in more detail. This work focuses on N-linked glycans obtained from human immunodeficiency virus (HIV) gp120 expressed from 293T and 293S mutant cell lines or in the presence of glycosylation inhibitors. Modified glycoforms of viral glycoprotein are currently the subject of intense research by many groups, including our own, in the context of antiviral therapies [30–32], vaccine design [33–36] and structural biology [37–41]. In the present study, glycoproteins were expressed in human embryonic kidney (HEK) 293T cells and, for the production of glycoproteins with modified glycans, in the presence of the glycosidase inhibitor, N-butyldeoxynojirimycin (NBDNJ), or produced in HEK 293S cells that are deficient in GlcNAc transferase I [42] which is involved in the biosynthesis of the N-glycans. We have previously used a panel of gp120 glycoforms with native and modified glycans to resolve the specificity of anti-carbohydrate antibodies following immunization with yeasts mannans [34].
In the analysis of glycoprotein glycosylation, such as in the above experiments, sample size can be limited and, in such cases, it is frequently difficult to obtain satisfactory mass spectra without extensive chemical clean-up with its concomitant losses which tend to intensify the problem. We [15, 43] and others [18] have already explored the use of ion mobility mass spectrometry to separate glycans from other compounds and we have now investigated the ability of this technique to separate glycan from background ions with the object of reducing the clean-up steps necessary for the production of spectra containing complete glycan profiles from very small amounts of starting glycoprotein. The development of methods for the analysis of low abundance viral glycoproteins, such as those from gp120 from HIV discussed here, will enable the analysis of native viral glycoproteins released directly from infectious virions where the low sample size is of critical importance.
2 Materials and Methods
2.1 Materials
The recombinant ectodomain of gp120BaL glycoprotein, from amino acid residue 1 to 507 (numbering based on alignment with the HxB2 reference strain), was cloned into the pHLsec vector [44] and transiently expressed in HEK 293T cells with and without the presence of 2 mM of the glycosidase inhibitor NBDNJ. The glycoprotein was also expressed in a mutant HEK 293S cell line deficient in GlcNAc-transferase I [42]. NBDNJ was added at the time of transient transfections, which were performed using a mixture of polyethylenimine and 2 mg DNA per liter of cell culture. Recombinant gp120 was initially isolated by immobilized metal affinity chromatography then further purified by size exclusion chromatography using a Superdex 200 Prep Grade column (GE Health Care, Amersham, UK). Solvents and other chemicals were from Sigma (Poole, UK). NBDNJ was a gift from Dr. T. Butters (University of Oxford), 2,5-dihydroxybenzoic acid (DHB, the MALDI matrix) was obtained from Aldrich Chemicals (Poole, UK) and protein N-glycosidase F (PNGase F) was from New England Biolabs (Ipswich, MA, USA).
2.2 Glycan Release
2.2.1 In-Gel Release
N-Glycans were released from glycoproteins with PNGase F essentially by a modification of the in-gel method first described by Küster et al. [45]. Briefly, the glycoproteins (20, 0.5, 0.1. 0.05, 0.01 μg) were separated on sodium dodecyl sulfate (SDS) polyacrylamide electrophoresis (PAGE) gels (NuPage 4%–12% Bis-Tris SDS-PAGE gel (Invitrogen, Paisley, UK) using MOPS running buffer (Invitrogen), stained with Coomassie blue dye and the areas containing the glycoproteins were excised and destained. PNGase F was infused in water and the gel pieces were incubated overnight (about 16 hours) at 37 °C. The released glycans were extracted with several additions of water and the resulting solution was filtered with a 0.45 μm hydrophobic protein binding Immobilon membrane held in a 96-well MultiScreen sterile plate (Millipore, Billerica, MA, USA). Samples (1 μL from approx 500 μL) were analysed following further clean-up with a Nafion 117 membrane as described earlier by Börnsen et al. [8] or, if the original enzymatic-release samples contained only a low amount of glycan, they were first concentrated to about 10 μL.
2.2.2 Solution Release
The glycoprotein was incubated overnight at 37 °C with PNGase F both with and without the denaturant nonylphenoxylpolyethoxylethanol (NP40) and dithiothreitol (DTT). Samples were examined either directly from the incubation solution or following clean-up with a Nafion 117 membrane.
2.3 Instrumentation
2.3.1 Ion Mobility
Ion mobility data were acquired with Waters Synapt (first generation, referred to as the G1 instrument) and G2 mass spectrometers (Waters MS-Technologies, Manchester, UK) [46, 47] fitted either with an Advion Nanomate (Advion Biosystems, Ithaca, New York, USA) sample introduction system (G1 instrument) or an inlet device for holding Proxeon (Proxeon Biosystems, Odense, Denmark) borosilicate nano-capillaries (G1 and G2 instruments). Results from the two instruments were similar except for the resolution (both MS and ion mobility), which was higher on the G2 instrument. Samples were dissolved in water:methanol (1:1, vol:vol) containing a trace of sodium chloride (positive ion) or either ammonium nitrate or ammonium phosphate (negative ion) and spun briefly (13,000 rpm) to sediment any solid material. Data were recorded with a T-wave velocity of 450 m/s and a wave height of 14 V (positive ion) and 16 V (negative ion) with the G1 instrument and wave velocity 600 m/s (positive ion) and 450 V (negative ion), both with a wave height of 40 V on the G2 instrument. Fragmentation was performed in the transfer collision cell (following ion mobility) with argon as the collision gas.
2.3.2 MALDI-TOF
MALDI-TOF data were acquired with a Shimadzu-Kratos TOF2 mass spectrometer (Kratos Analytical, Manchester, UK). Samples, in water (1 μL), were deposited on the stainless steel target, mixed with 0.5 μL of the matrix solution [DHB, 10 mg in 1 mL water:MeCN (1:1 by volume)] and allowed to dry under ambient conditions. The dried sample spot was then recrystallised from the minimum amount of ethanol (about 0.2 μL). The instrument was operated in reflectron, delayed extraction mode with focusing conditions optimized for DHB and the delayed extraction set for a mass of 1700 Da.
2.3.3 ESI-MS/MS
Electrospray mass spectrometry was performed with a Waters quadrupole-time-of-flight (Q-TOF) Ultima Global instrument in negative ion mode. Samples in 1:1 (vol:vol) methanol:water containing 0.5 mM ammonium phosphate were infused through Proxeon nanospray capillaries (Proxeon Biosystems, Odense, Denmark). The ion source conditions were: temperature, 120 °C; nitrogen flow 50 L/h; infusion needle potential, 1.2 kV; cone voltage 100 V; RF-1 voltage 150 V. Spectra (2 s scans) were acquired with a digitization rate of 4 GHz and accumulated until a satisfactory signal:noise ratio had been obtained. For MS/MS data acquisition (collision-induced decomposition, CID), the parent ion was selected at low resolution (about 5 m/z mass window) to allow transmission of isotope peaks and fragmented with argon at a pressure (recorded on the instrument’s pressure gauge) of 0.5 mBar. The voltage on the collision cell was adjusted with mass and charge to give an even distribution of fragment ions across the mass scale. Typical values were 80–120 V. Other voltages were as recommended by the manufacturer. Instrument control, data acquisition and processing were performed with MassLynx software ver. 4.1.
3 Results
3.1 Direct Analysis of PNGase F-Released Glycans from gp120 Expressed in HEK 293T Cells
Figure 1b shows the positive ion electrospray spectrum of a sample of N-glycans released in-gel with PNGase F from the HIV glycoprotein gp120 expressed in HEK 293T cells (10 μg) and cleaned with a protein binding Immobilon-P membrane followed by dialysis on a Nafion membrane. Five ions due to N-glycans were visible but the signal:noise ratio was poor. The sample was then examined by ion mobility MS (Synapt G1 instrument with Nanomate sample introduction) and the separation shown in the Driftscope display (m/z:drift time) at the top of the figure (Figure 1a) was obtained. The region enclosed by the oval contains singly charged ions from the N-glycans. The spectrum from this region was selected and displayed with the Waters MassLynx software (Figure 1c). Ions produced by the N-glycans were now clearly visible with a good signal:noise ratio and the spectrum was similar to a MALDI-TOF spectrum (from 20 μg glycoprotein) shown in Figure 1d. Structures were assigned, in this case, from parallel experiments using negative ion ESI MS/MS with a Waters Q-TOF Ultima Global instrument [48–51] and are listed in Table 1. A rough outline of the N-glycan processing pathway is shown in Scheme 1. Although qualitatively the MALDI-TOF and ESI-IM-Q-TOF spectra are similar, there appears to be a bias against the larger glycans in the ESI spectrum. The band of ions immediately above that from the N-glycans in Figure 1a consisted of unidentified (non-glycan) singly charged ions whereas the other bands were mainly from doubly charged ions of unknown origin.

(a) Driftscope display from the G1 instrument of positive ions from N-glycans released with PNGase F from the glycoprotein gp120 expressed in HEK 293 T cells. The region enclosed by the oval contains the ions from the N-glycans, which gave the spectrum shown in (c). (b) The ESI spectrum of the crude sample before mobility separation. (c) Extracted spectrum of the N-glycans. (d) MALDI-TOF spectrum. The ions at m/z 2012 and 2174 did not appear in the ESI spectrum

Outline scheme for the biosynthesis of N-glycans. Key to symbols used to draw the N-glycan structures: ■ = GlcNAc, ○ = mannose,  = fucose, ♢ = galactose, □ = glucose. The angle of the lines connecting the symbols shows linkage with full lines indicating β-bonds and broken lines showing α-bonds. For further details see reference [56]. The first stage in the biosynthesis of these compounds is the attachment of the tetradecyl glycan (I) to the developing protein followed by removal of the glucose residues by two enzymes, glucosidases I and II give the “high-mannose” glycan known as “Man-9”, II. Following this step, mannosidases trim the outer four mannose residues to give Man-5 (VI) via III, IV, and V. The drug nBDNJ blocks this process so that the mannosidases can only remove mannose residues from the 6-antenna to give X via IX. However, an “escape” pathway involving an endomannosidase can cleave the 3-antenna of I to give XI, which is then a substrate for the exo-mannosidases giving Man-5 (VI). Man-5 is a substrate for the enzyme GlcNAc-transferase I, which then adds a GlcNAc residue to the 3-antenna to give VII, which is then a substrate for another exomannosidase which trims the remaining two outer mannose residues from the 6-antenna to give VIII. This glycan is a substrate for GlcNAc-transferase II, which adds a second GlcNAc residue to the 2-position of the remaining mannose of the 6-antenna. This compound is the route for biosynthesis of the “complex” glycans of which only a few are shown in Scheme 1. The scheme shows addition of two galactose residues to give the “biantennary” complex glycan, XIII, and addition of a third antenna (in several stages) to give the “triantennary” glycan XIV. Fucose can be added at the 6-position of the reducing-terminal GlcNAc to give the compounds XV and XVI. Antennae are frequently terminated with sialic acid. Further biosynthesis (not shown) can proceed in a similar manner to build up much larger structures. Additional information can be found in standard glycobiology text books, e.g., [57]
= fucose, ♢ = galactose, □ = glucose. The angle of the lines connecting the symbols shows linkage with full lines indicating β-bonds and broken lines showing α-bonds. For further details see reference [56]. The first stage in the biosynthesis of these compounds is the attachment of the tetradecyl glycan (I) to the developing protein followed by removal of the glucose residues by two enzymes, glucosidases I and II give the “high-mannose” glycan known as “Man-9”, II. Following this step, mannosidases trim the outer four mannose residues to give Man-5 (VI) via III, IV, and V. The drug nBDNJ blocks this process so that the mannosidases can only remove mannose residues from the 6-antenna to give X via IX. However, an “escape” pathway involving an endomannosidase can cleave the 3-antenna of I to give XI, which is then a substrate for the exo-mannosidases giving Man-5 (VI). Man-5 is a substrate for the enzyme GlcNAc-transferase I, which then adds a GlcNAc residue to the 3-antenna to give VII, which is then a substrate for another exomannosidase which trims the remaining two outer mannose residues from the 6-antenna to give VIII. This glycan is a substrate for GlcNAc-transferase II, which adds a second GlcNAc residue to the 2-position of the remaining mannose of the 6-antenna. This compound is the route for biosynthesis of the “complex” glycans of which only a few are shown in Scheme 1. The scheme shows addition of two galactose residues to give the “biantennary” complex glycan, XIII, and addition of a third antenna (in several stages) to give the “triantennary” glycan XIV. Fucose can be added at the 6-position of the reducing-terminal GlcNAc to give the compounds XV and XVI. Antennae are frequently terminated with sialic acid. Further biosynthesis (not shown) can proceed in a similar manner to build up much larger structures. Additional information can be found in standard glycobiology text books, e.g., [57]
Without Nafion clean-up, it was difficult to achieve a stable spray with the Nanomate but, after treatment, all samples gave spectra. In other experiments that employed Proxeon capillaries that have a wider orifice, spectra could often be obtained without Nafion clean-up although pretreatment was usually an advantage.
In order to assess usefulness of ion mobility to extract glycan spectra from incubation mixtures with minimum cleanup, the glycoprotein gp120 (10 μg) was incubated directly in solution with PNGase F and the mixture was examined by diluting 10% of the sample (2 μL of the incubation mixture) with 5 μL of ESI solvent and infusing into the instrument with a Proxeon capillary. The positive ion ESI spectrum (Figure 2a) showed no glycan ions above the noise level but when separated by ion mobility, as above, the spectrum shown in Figure 2b was obtained. Another sample of the incubation mixture was allowed to stand on a Nafion membrane for three hours before being examined by ESI as above but, again, glycan ions were not apparent in the ESI spectrum. Clean-up by ion mobility again produced a clean spectrum which was identical to that obtained without Nafion treatment confirming no fractionation of the glycans by this clean-up method. A comparison of signal strength of the background and N-glycan ions before and after Nafion treatment showed that the signal:noise ratio had improved by about a factor of three. However, the Nafion treatment alone did not allow the glycan ions to be seen.

(a) Positive ion electrospray mass spectrum (G1 instrument) of N-glycans released in-solution from 10 μg gp120. No glycans are visible. (b) Spectrum of the N-glycans extracted by ion mobility. Symbols for the glycan structures are as defined in the legend to Scheme 1
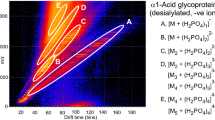
Release of N-glycans by PNGase F from native, non-denatured glycoproteins is not considered to be quantitative because of steric effects and the normal practice is to denature the glycoprotein before release or to cleave it with trypsin or another protease as was done by Fenn and McLean [18] in a study with ribonuclease B glycans. It would obviously be an advantage if the proteolysis step could be dispensed with and, consequently, the glycans were released from 10 μg of the glycoprotein in the presence of the denaturants, NP-40 and DTT. Glycan release from the resulting denatured glycoprotein gave a considerably increased number and abundance of the background ions but also of the abundance of ions from the glycans (Figure 3). After a 3 hour treatment with Nafion, the ESI spectrum showed evidence of the presence of several glycans (Figure 3b) but the mass region containing the ions from these compounds also contained many other ions from unidentified singly and doubly charged oligomers (see Figure 3c inset). The NP-40 detergent, being of lower molecular weight, gave most ions below the N-glycan region. After ion mobility separation (Figure 3a), a clean spectrum of the N-glycans was obtained (Figure 3e). The profile was different to that obtained without the denaturant, particularly in the relative abundance of the high-mannose glycans Man5-9GlcNAc2. These compounds are known to be closely packed on the surface of the glycoprotein [33, 52], presumably inhibiting release by the endoglycosidase PNGase F in the absence of denaturant. Although falling in the same mass region as the N-glycans, the singly and doubly charged ions from the NP-40 oligomers exhibited different mobilities to the N-glycans and were easily separated (see regions 1 and 3, Figure 3a). Also separated were a series of [M + Na2]2+ ions from the N-glycans (Region 4, Figure 3g). The quantitative profile of these ions was different from that of the singly charged ions, as observed before [53]; the larger compounds gave relatively stronger signals, probably accounting for the bias against the higher mass singly charged ions noted above. The relative abundance of the singly and doubly charged glycan ions could be varied by alteration of the ion source focusing conditions but was not investigated thoroughly in these experiments. The graph in Figure 3b shows the ion mobility:m/z plot for the major compounds separated as in Figure 3a.

(a) DriftScope display (positive ion, G2 instrument) of ions from the solution release by PNGase F of N-glycans from gp-120 in the presence of NP-40 and DTT. Circled regions are: 1, singly charged unidentified oligomers; 2, singly charged N-glycans ([M + Na]+ ions); 3, doubly charged unidentified oligomers; 4, doubly charged ([M + Na2]2+ ions); 5, NP-40. (b) Mobility:m/z plot of the major compounds from the mobility separation shown in 3a. The numbers in the legend refer to the regions in panel (a). (c) Total electrospray spectrum. The insert shows the N-glycan region. (d) Extracted mass spectrum of the singly charged unidentified oligomers (Region 1). (e) Extracted mass spectrum of the N-glycans (Region 2). (f) Extracted mass spectrum of the doubly charged unidentified oligomers (Region 3) (g) Extracted spectrum of the doubly charged N-glycan ions and unidentified oligomers (Region 4). (h) Extracted mass spectrum of the NP-40 ions (Region 5). Symbols for the glycan structures are as defined in the legend to Scheme 1
3.2 Determination of Lowest Glycoprotein Amount from Which Usable Glycan Profiles Could Be Obtained
N-glycans were released from known amounts of gp120 glycoprotein (20.0, 0.5, 0.1, 0.05 and 0.01 μg) and examined by both MALDI-TOF MS and ESI ion mobility-TOF MS (Synapt G2 instrument). A strong MALDI-TOF spectrum was observed from the most concentrated (20 μg) sample but not from the other four. The positive ion ESI-ion mobility spectra, on the other hand produced glycan spectra (Figure 4) from all samples although only the major ions were observed from the least concentrated sample (0.01 μg, Figure 4d). 20% of each sample was taken for analysis; thus the samples analysed were effectively from 4.0, 0.1, 0.02, 0.01, and 0.002 μg of glycoprotein. However, it should be noted that gp120 contains 26 occupied glycosylation sites and that relatively more starting material would be required for analysis of a glycoprotein with fewer occupied sites. A log/log plot of sample amount against signal strength for four of the most abundant ions in the spectra was roughly linear (Figure 4b). Use of Nafion for clean-up prior to ion mobility separation made little difference to the appearance of the spectra.

(a) Driftscope display (G2 instrument) of positive ions from gp120 released from 20 μg of gp120 glycoprotein. The N-glycan region is contained within the oval. (b) Double log plot of the major N-glycan ions released from each of five samples of gp120 glycoprotein at different concentrations (20, 0.5, 0.1. 0.05, and 0.01 μg). Ions are identified on the graph. (c) Extracted ESI spectrum of the N-glycans from the 20 μg sample. (d) Extracted ESI spectrum of the N-glycans from the 0.01 μg sample. Symbols for the glycan structures are as defined in the legend to Scheme 1
3.3 Fragmentation
Following ion mobility separation, the glycans could be fragmented in the transfer collision cell to obtain structural information. The ion mobility separation was generally insufficient to resolve isomers or conformers but, to avoid any possible problems, fragmentation spectra were acquired by averaging all spectra taken across the drift time peak. Figure 5a shows the positive ion CID spectra of the high-mannose glycan Man5GlcNAc2 from gp120. The fragmentation pattern was similar to that obtained earlier by CID with a Q-TOF instrument [53]. The spectra were dominated by glycosidic cleavage fragments mainly resulting in losses of mannose residues from the [M + Na]+ and B4 (from Man5,6GlcNAc2) and B5 from Man7-9GlcNAc2) fragments (the Domon and Costello nomenclature [54] is used to describe the fragment ions). This spectrum was not as informative as the negative ion spectrum [48–51] shown in Figure 5b. In this spectrum, the 2,4A5, B4, and 2,4A4 ions at m/z 1072, 1012, and 869, respectively, define the di-N-acetylchitobiose core and the D, [D-18]– and O,3A3 ions at m/z 647, 629, and 575, respectively, define the composition of the 6-antenna. The mannose residues capping the antennae produce the C1 ion at m/z 179.

(a) Positive ion CID spectrum of Man5GlcNAc2. (b) Negative ion CID spectrum of Man5GlcNAc2
3.4 HIV gp120 Expressed in HEK-293 Cells Deficient in N-Acetylglucosyltransferase I
The Golgi-resident glycosyltransferase, N-acetylglucosyltransferase I (GnT) adds a GlcNAc residue to the glycan Man5GlcNAc2 (VI, see a description of the biosynthetic pathway in the legend to Scheme 1) to give the glycan VII, the precursor to complex glycan formation (Compounds XIII–XVI). Consequently, the glycan profile from gp120 expressed in the cell line deficient in this enzyme would be expected to contain only the high-mannose glycans II–VI. The positive ion ESI spectrum (Figure 6a) contained a hint of Man5GlcNAc2 (Figure 6a inset) but the ion was not sufficiently distinguishable from the noise for a positive confirmation. No signals were seen from other expected glycans. After ion mobility separation, the five expected glycans (II–VI) were well separated from the background noise Figure 6b). Because this cell line is unable to add GlcNAc to the Man5GlcNAc2 glycan (VI), no further processing occurs except for a trace amount of fucosylated Man5GlcNAc2 which arises due to the GnT I-independent fucosylation pathway [55].

(a) Positive ion ESI mass spectrum (G1 instrument) of N-glycans from gp120 Expressed in HEK 293S cells deficient in N-acetylglucosyltransferase I. The inset shows an expanded region of m/z 1250–1265 with an arrow indicating the [M + Na]+ ion from Man5GlcNAc2. (b) The N-glycan region from the sample shown in (a) extracted from the mobility:m/z plot (the same mass scale is used). (c) The corresponding negative ion mobility-extracted spectrum. (d) Extracted ESI spectrum of N-glycans released with PNGase F from the glycoprotein gp120 expressed in the presence of NBDNJ. Symbols for the glycan structures are as defined in the legend to Scheme 1
The glycan isolation also worked equally well in negative ion mode. The negative ion ESI spectrum again did not show evidence of any glycans until after ion mobility separation when all five high-mannose glycans were detected (Figure 6c, [M + H2PO4]– adducts). The spectrum was not as clean as the positive ion spectrum because of the presence of fragment ions; predominantly 2,4A6 cleavages of the reducing terminal GlcNAc residue. These ions were not separated from the glycan molecular ions by ion mobility. However, none of these fragment ions gave reasonable compositions for native glycans allowing the true glycans to be identified. The advantage of the use of negative ions, as discussed above, is that their fragmentation leads to much more diagnostic spectra than does fragmentation of [M + Na]+ ions [48–51].
3.5 N-Glycans from gp120 Obtained in the Presence of the Glycosidase Inhibitor NBDNJ
NBDNJ is a glucosidase inhibitor and is expected to prevent the initial removal of the glucose residues from the 3-antenna of Glc3Man9GlcNAc2 (I), the glycan that is initially added to the protein during biosynthesis. N-glycans were released from gp120 that had been expressed in HEK-293 cells in the presence of NBDNJ and, again, no glycans were observed in the ESI spectrum. After selecting the glycan region from the positive ion mobility spectrum, glycans were observed with a good signal:noise ratio. NBDNJ was observed to inhibit glucose removal and to maintain the integrity of the 3-antenna. The ions at m/z 2229 and 2067 in Figure 6d are the result of α-mannosidases removing the two mannose residues from the 6-antenna (Compounds IX and X) as shown by negative ion fragmentation. Although NBDNJ blocks the classical degradation pathway of the 3-antenna (sequential removal of glucose and mannose residues) leading to glycans VI and VII, complex glycans were still observed in the profile. This can be accounted for by the action of an endogenous endomannosidase that is able to cleave the 3-antenna between the second and third mannose residues to give glycan XI directly from glycan I, thus releasing the glucose residues and enabling glycan processing to proceed normally (XI to VI and then to VII, VIII, and so forth).
4 Discussion
A major problem with obtaining spectra of released glycans and other compounds, when sample sizes are limited, is signal suppression or masking from other compounds present in the analytical samples. This problem is particularly evident in biological samples where compounds such as buffer salts, anti-oxidants, and detergents are frequently present. Many of these compounds are difficult to remove and clean-up steps inevitably cause loss of material, thus exacebating the problem. The work presented here uses ion mobility to separate the ions from these different compounds, enabling only the region of interest to be examined. Many of the contaminating compounds more easily form multiply charged ions than the compounds of interest, in this case N-glycans released from glycoproteins, allowing considerable separations in mobility to be achieved. Thus, many contaminating compounds occupy regions of the mobility/m/z map remote from the glycans by exhibiting higher mobilities. A few compounds were observed to have lower mobilities than the N-glycans (singly charged DTT oligomers in this work) but even these were well separated. Even different carbohydrates, such as dextran oligomers, could be separated (not shown). These compounds, being linear, had lower mobilities than the branched N-glycans. The term “signal suppression” is often applied to samples that fail to show the ions of interest but, in this work, at least, much of the problem with observation of the required ions appears to be simple masking of the ions of interest by the ions from the contamination compounds. As ion mobility shows, the required ions are often still present and can be revealed by techniques that are able to perform such compound-specific separations.
Electrospray ionization is possibly not the ideal ionization technique for examination of these compounds because of the tendency of many of them to form multiply charged ions and to undergo in-source fragmentation. [M + H]+ and [M – H]– ions are particularly prone to fragmentation but, fortunately, glycans usually ionize as [M + Na]+ ions in positive ion mode, particularly in the presence of small amounts of added sodium salts, as was the situation in this work. In negative ion mode, glycans can be stabilized as anionic adducts with, for example, Cl–, NO –3 , or H2PO –4 . In the spectra shown above (Figures 1c, 2b, 4c, and 6d), the high-mannose glycans appeared to ionize without fragmentation but the presence, in low abundance, of ions such as m/z 1444 (positive ion spectra) corresponding to a composition of Hex4HexNAc3dHex1 that were absent, or of low relative abundance in the MALDI-TOF spectra, suggest the presence of a Y4 cleavage product from the fucosylated biantennary glycan, Gal2G1cNAc4Man4Fuc1 (Compound XV) present in several of the samples. These fragment ions probably arose from [M + H]+ precursors. In this respect, MALDI might be a better option for ionizing these compounds but has not yet been investigated (MALDI was not available on the instruments used for this work). On the other hand, MALDI tends not to produce multiple charging, meaning that the contaminating compounds may not separate as well in the mobility map.
5 Conclusions
Ion mobility ESI-MS offers a robust method for the analysis of N-glycans from low abundance biological materials such as viral glycoproteins. The ion mobility MS approaches outlined here allow for the analysis of sub-microgram quantities of glycoprotein and, when combined with negative ion MS/MS, provide a reliable approach to the structural analysis of glycosylation.
References
Viëtor, R., Loutelier-Bourhis, C., Fitchette, A.-C., Margerie, P., Gonneau, M., Faye, L., Lerouge, P.: Protein N-glycosylation is similar in the Moss Physcomitrella patens and in higher plants. Planta 218, 269–275 (2003)
Yu, Y.Q., Gilar, M., Kaska, J., Gebler, J.C.: A rapid sample preparation method for mass spectrometric characterization of N-linked glycans. Rapid Commun. Mass Spectrom. 19, 2331–2336 (2005)
Parry, S., Hanisch, F.G., Leir, S.-H., Sutton-Smith, M., Morris, H.R., Dell, A., Harris, A.: N-Glycosylation of the MUC1 mucin in epithelial cells and secretions. Glycobiology 16, 623–634 (2006)
Higai, K., Aoki, Y., Azuma, Y., Matsumoto, K.: Glycosylation of site-α-acid glycoprotein and alterations in acute and chronic inflammation. Biochim. Biophys. Acta 1725, 128–135 (2005)
An, H.J., Franz, A.H., Lebrilla, C.B.: Improved capillary electrophoretic separation and mass spectrometric detection of oligosaccharides. J. Chromatogr. A 1004, 121–129 (2003)
Mills, P.B., Mills, K., Mian, N., Winchester, B.G., Clayton, P.T.: Mass spectrometric analysis of glycans in elucidating the pathogenesis of CDG type IIx. J. Inherit. Metab. Dis. 26, 119–134 (2003)
Forno, G., Fogolin, M.B., Oggero, M., Kratje, R., Etcheverrigaray, M., Conradt, H.S., Nimtz, M.: N- and O-linked carbohydrates and glycosylation site occupancy in recombinant human granulocyte-macrophage colony-stimulating factor secreted by a Chinese hamster ovary cell line. Eur. J. Biochem. 271, 907–919 (2004)
Börnsen, K.O., Mohr, M.D., Widmer, H.M.: Ion exchange and purification of carbohydrates on a Nafion(R) membrane as a new sample pretreatment for matrix-assisted laser desorption-ionization mass spectrometry. Rapid Commun. Mass Spectrom. 9, 1031–1034 (1995)
Mechref, Y., Muzikar, J., Novotny, M.V.: Comprehensive assessment of N-glycans derived from a murine monoclonal antibody: a case for multimethodological approach. Electrophoresis 26, 2034–2046 (2005)
Sparbier, K., Wenzel, T., Kostrzewa, M.: Exploring the binding profiles of ConA, boronic acid and WGA by MALDI-TOF/TOF MS and magnetic particles. J. Chromatogr. B 840, 29–36 (2006)
Vosseller, K., Trinidad, J.C., Chalkley, R.J., Specht, C.G., Thalhammer, A., Lynn, A.J., Snedecor, J.O., Guan, S., Medzihradszky, K.F., Maltby, D.A., Schoepfer, R., Burlingame, A.L.: O-Linked N-acetylglucosamine proteomics of postsynaptic density preparations using lectin weak affinity chromatography and mass spectrometry. Mol. Cell. Proteom. 5, 923–934 (2006)
Lee, J.H., Kim, Y., Ha, M.Y., Lee, E.K., Choo, J.: Immobilization of aminophenylboronic acid on magnetic beads for the direct determination of glycoproteins by matrix assisted laser desorption ionization mass spectrometry. J. Am. Soc. Mass Spectrom. 16, 1456–1460 (2005)
Luke, N.R., Allen, S., Gibson, B.W., Campagnari, A.A.: Identification of a 3-deoxy-D-manno-octulosonic acid biosynthetic operon in Moraxella catarrhalis and analysis of a KdsA-deficient isogenic mutant. Infect. Immun. 71, 6426–6434 (2003)
Harvey, D.J., Küster, B., Wheeler, S.F., Hunter, A.P., Bateman, R.H., Dwek, R.A.: Matrix-assisted laser desorption/ionization mass spectrometry of N-linked carbohydrates and related compounds. In: Burlingame, A.L., Carr, S.A., Baldwin, M.A. (eds.) Mass spectrometry in biology and medicine, pp. 407–437. Humana Press, Totowa (2000)
Williams, J.P., Grabenauer, M., Carpenter, C.J., Holland, R.J., Wormald, M.R., Giles, K., Harvey, D.J., Bateman, R.H., Scrivens, J.H., Bowers, M.T.: Characterization of simple isomeric oligosaccharides and the rapid separation of glycan mixtures by ion mobility mass spectrometry. Int. J. Mass Spectrom. 298, 119–127 (2010)
Fenn, L.S., McLean, J.A.: Biomolecular structural separations by ion mobility–mass spectrometry. Anal. Bioanal. Chem. 391, 905–909 (2008)
Fenn, L.S., Kliman, M., Mahsut, A., Zhao, S.R., McLean, J.A.: Characterizing ion mobility-mass spectrometry conformation space for the analysis of complex biological samples. Anal. Bioanal. Chem. 394, 235–244 (2009)
Fenn, L.S., McLean, J.A.: Simultaneous glycoproteomics on the basis of structure using ion mobility-mass spectrometry. Mol. Biosyst. 5, 1298–1232 (2009)
Isailovic, D., Kurulugama, R.T., Plasencia, M.D., Stokes, S.T., Kyselova, Z., Goldman, R., Mechref, Y., Novotny, M.V., Clemmer, D.E.: Profiling of human serum glycans associated with liver cancer and cirrhosis by IMS-MS. J. Proteome Res. 7, 1109–1117 (2008)
Damen, C.W.N., Chen, W., Chakraborty, A.B., van Oosterhout, M., Mazzeo, J.R., Gebler, J.C., Schellens, J.H.M., Rosing, H., Beijnen, J.H.: Electrospray ionization quadrupole ion-mobility time-of-flight mass spectrometry as a tool to distinguish the lot-to-lot heterogeneity in N-glycosylation profile of the therapeutic monoclonal antibody Trastuzumab. J. Am. Soc. Mass Spectrom. 20, 2021–2033 (2009)
Olivova, P., Chen, W., Chakraborty, A.B., Gebler, J.C.: Determination of N-glycosylation sites and site heterogeneity in a monoclonal antibody by electrospray quadrupole ion-mobility time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 22, 29–40 (2008)
Clowers, B.H., Dwivedi, P., Steiner, W.E., Hill, H.H.J., Bendiak, B.: Separation of sodiated isobaric disaccharides and trisaccharides using electrospray ionization-atmospheric pressure ion mobility-time of flight mass spectrometry. J. Am. Soc. Mass Spectrom. 16, 660–669 (2005)
Dwivedi, P., Bendiak, B., Clowers, B.H., Hill, H.H.J.: Rapid resolution of carbohydrate isomers by electrospray ionization ambient pressure ion mobility spectrometry-time-of-flight mass spectrometry (ESI-APIMS-TOFMS). J. Am. Soc. Mass Spectrom. 18, 1163–1175 (2007)
Gabryelski, W., Froese, K.L.: Rapid and sensitive differentiation of anomers, linkage, and position isomers of disaccharides using high-field asymmetric waveform ion mobility spectrometry (FAIMS). J. Am. Soc. Mass Spectrom. 14, 265–277 (2003)
Yamagaki, T., Sato, A.: Peak width-mass correlation in CID MS/MS of isomeric oligosaccharides using traveling-wave ion mobility mass spectrometry. J. Mass Spectrom. 44, 1509–1517 (2009)
Yamagaki, T., Sato, A.: Isomeric oligosaccharides analyses using negative-ion electrospray ionization ion mobility spectrometry combined with collision-induced dissociation MS/MS. Anal. Sci. 25, 985–988 (2009)
Zhu, M., Bendiak, B., Clowers, B., Hill, H.H.: Ion mobility-mass spectrometry analysis of isomeric carbohydrate precursor ions. Anal. Bioanal. Chem. 394, 1853–1867 (2009)
Plasencia, M.D., Isailovic, D., Merenbloom, S.I., Mechref, Y., Clemmer, D.E.: Resolving and assigning N-linked glycan structural isomers from ovalbumin by IMS-MS. J. Am. Soc. Mass Spectrom. 19, 1706–1715 (2008)
Kanu, A.B., Dwivedi, P., Tam, M., Matz, L., Hill, H.H.J.: Ion mobility-mass spectrometry. J. Mass Spectrom. 43, 1–22 (2008)
Fischer, P.B., Collin, M., Karlsson, G.B., James, W., Butters, T.D., Davis, S.J., Gordon, S., Dwek, R.A., Platt, F.M.: The alpha-glucosidase inhibitor N-butyldeoxynojirimycin inhibits human immunodeficiency virus entry at the level of post-CD4 binding. J. Virol. 69, 5791–5797 (1995)
Pollicita, M., Schols, D., Aquaro, S., Peumans, W.J., Van Damme, E.J., Perno, C.F., Balzarini, J.: Carbohydrate-binding agents (CBAs) inhibit HIV-1 infection in human primary monocyte-derived macrophages (MDMs) and efficiently prevent MDM-directed viral capture and subsequent transmission to CD4+ T lymphocytes. J. Virol. 370, 382–391 (2008)
Auwerx, J., François, K.O., Vanstreels, E., Van Laethem, K., Daelemans, D., Schols, D., Balzarini, J.: Capture and transmission of HIV-1 by the C-type lectin L-SIGN (DC-SIGNR) is inhibited by carbohydrate-binding agents and polyanions. Antivir. Res. 83, 61–70 (2009)
Scanlan, C.N., Offer, J., Zitzmann, N., Dwek, R.A.: Exploiting the defensive sugars of HIV-1 for drug and vaccine design. Nature 446, 1038–1045 (2007)
Dunlop, D.C., Bonomelli, C., Mansab, F., Vasiljevic, S., Doores, K.J., Wormald, M.R., de Sa Palma, A., Feizi, T., Harvey, D.J., Dwek, R.A., Crispin, M., Scanlan, C.N.: Polysaccharide mimicry of the epitope of the broadly neutralising anti-HIV antibody, 2G12, induces enhanced antibody responses to self oligomannose glycans. Glycobiology. 20, 812–823 (2010)
Scanlan, C.N., Ritchie, G.E., Baruah, K., Crispin, M., Harvey, D.J., Singer, B.B., Lucka, L., Wormald, M.R., Wentworth, P.J., Zitzmann, N., Rudd, P.M., Burton, D.R., Dwek, R.A.: Inhibition of mammalian glycan biosynthesis produces non-self antigens for a broadly neutralising, HIV-1 specific antibody. J. Mol. Biol. 372, 16–22 (2007)
Doores, K.J., Bonomelli, C., Harvey, D.J., Vasiljevic, S., Dwek, R.A., Burton, D.R., Crispin, M., Scanlan, C.N.: Envelope glycans of immunodeficiency virions are almost entirely oligomannose antigens. Proc. Natl Acad. Sci. USA 107, 13800–13805 (2010)
Chang, V.T., Crispin, M., Aricescu, A.R., Harvey, D.J., Nettleship, J.E., Fennelly, J.A., Yu, C., Boles, K.S., Evans, E.J., Stuart, D.I., Dwek, R.A., Jones, E.Y., Owens, R.J., Davis, S.J.: Glycoprotein structural genomics: solving the glycosylation problem. Structure 15, 267–273 (2007)
Bowden, T.A., Crispin, M., Graham, S.C., Harvey, D.J., Grimes, J.M., Jones, E.Y., Stuart, D.I.: Unusual molecular architecture of the Machupo virus attachment glycoprotein. J. Virol. 83, 8259–8265 (2009)
Bowden, T.A., Crispin, M., Harvey, D.J., Aricescu, A.R., Grimes, J.M., Jones, E.Y., Stuart, D.I.: Crystal structure and carbohydrate analysis of Nipah virus attachment glycoprotein: a template for antiviral and vaccine design. J. Virol. 82, 11628–11636 (2008)
Bowden, T.A., Crispin, M., Harvey, D.J., Jones, E.Y., Stuart, D.I.: Dimeric architecture of the Hendra virus attachment glycoprotein: evidence for a conserved mode of assembly. J. Virol. 84, 6208–6217 (2010)
Crispin, M., Bowden, T.A., Coles, C.H., Harlos, K., Aricescu, A.R., Harvey, D.J., Stuart, D.I., Jones, E.Y.: Carbohydrate and domain architecture of an immature antibody glycoform exhibiting enhanced effector functions. J. Mol. Biol. 387, 1061–1066 (2009)
Reeves, P.J., Callewaert, N., Contreras, R., Khorana, H.G.: Structure and function in rhodopsin: high-level expression of rhodopsin with restricted and homogeneous N-glycosylation by a tetracycline-inducible N-acetylglucosaminyltransferase I-negative HEK293S stable mammalian cell line. Proc. Natl Acad. Sci. USA 99, 13419–13424 (2002)
Harvey, D.J., Scrivens, J., Holland, R., Williams, J.P., Wormald, M.R.: Ion-mobility separation coupled with negative ion fragmentation of N-linked carbohydrates. Proceedings of the 56th ASMS Conference on Mass Spectrometry, Denver, CO, USA (2008), Proceedings CD, MOG 09.10 am
Aricescu, A.R., Lu, W., Jones, E.Y.: A time- and cost-efficient system for high-level protein production in mammalian cells. Acta Crystallogr. D Biol. Crystallogr. 62, 1243–1250 (2006)
Küster, B., Wheeler, S.F., Hunter, A.P., Dwek, R.A., Harvey, D.J.: Sequencing of N-linked oligosaccharides directly from protein gels: in-gel deglycosylation followed by matrix-assisted laser desorption/ionization mass spectrometry and normal-phase high performance liquid chromatography. Anal. Biochem. 250, 82–101 (1997)
Giles, K., Pringle, S.D., Worthington, K.R., Little, D., Wildgoose, J.L., Bateman, R.H.: Applications of a travelling wave-based radio-frequency-only stacked ring ion guide. Rapid Commun. Mass Spectrom. 18, 2401–2414 (2004)
Pringle, S.D., Giles, K., Wildgoose, J.L., Williams, J.P., Slade, S.E., Thalassinos, K., Bateman, R.H., Bowers, M.T., Scrivens, J.H.: An investigation of the mobility separation of some peptide and protein ions using a new hybrid quadrupole/travelling wave IMS/oa-TOF instrument. Int. J. Mass Spectrom. 261, 1–12 (2007)
Harvey, D.J.: Fragmentation of negative ions from carbohydrates: part 2. Fragmentation of high-mannose N-linked glycans. J. Am. Soc. Mass Spectrom. 16, 631–646 (2005)
Harvey, D.J.: Fragmentation of negative ions from carbohydrates: part 1; Use of nitrate and other anionic adducts for the production of negative ion electrospray spectra from N-linked carbohydrates. J. Am. Soc. Mass Spectrom. 16, 622–630 (2005)
Harvey, D.J.: Fragmentation of negative ions from carbohydrates: part 3, Fragmentation of hybrid and complex N-linked glycans. J. Am. Soc. Mass Spectrom. 16, 647–659 (2005)
Harvey, D.J., Royle, L., Radcliffe, C.M., Rudd, P.M., Dwek, R.A.: Structural and quantitative analysis of N-linked glycans by MALDI and negative ion nanospray mass spectrometry. Anal. Biochem. 376, 44–60 (2008)
Zhu, X., Borchers, C., Bienstock, R.J., Tomer, K.B.: Mass spectrometric characterization of the glycosylation pattern of HIV-gp120 expressed in CHO cells. Biochemistry 39, 11194–11204 (2000)
Harvey, D.J.: Collision-induced fragmentation of underivatised N-linked carbohydrates ionized by electrospray. J. Mass Spectrom. 35, 1178–1190 (2000)
Domon, B., Costello, C.E.: A systematic nomenclature for carbohydrate fragmentations in FAB-MS/MS spectra of glycoconjugates. Glycoconj. J. 5, 397–409 (1988)
Crispin, M., Harvey, D.J., Chang, V.T., Yu, C., Aricescu, A.R., Jones, E.Y., Davis, S.J., Dwek, R.A., Rudd, P.M.: Inhibition of hybrid- and complex-type glycosylation reveals the presence of the GlcNAc transferase I-independent fucosylation pathway. Glycobiology 16, 748–756 (2006)
Harvey, D.J., Merry, A.H., Royle, L., Campbell, M.P., Dwek, R.A., Rudd, P.M.: Proposal for a standard system for drawing structural diagrams of N- and O-linked carbohydrates and related compounds. Proteomics 9, 3796–3801 (2009)
Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E.: 2008. Essentials of Glycobiology, 2nd ed. Cold Spring Harbor Laboratory Press: 2008
Acknowledgments
The authors thank Professor Raymond Dwek, Director of the Oxford Glycobiology Institute, for his help and encouragement and Dr. James McCullagh (Department of Chemistry, Oxford) for access to the Synapt G1 mass spectrometer. They also thank the International AIDS Vaccine Initiative (IAVI) and the Oxford Glycobiology Institute endowment for financial support. They are particularly grateful to IAVI for an equipment grant to purchase a MALDI mass spectrometer.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Harvey, D.J., Sobott, F., Crispin, M. et al. Ion Mobility Mass Spectrometry for Extracting Spectra of N-Glycans Directly from Incubation Mixtures Following Glycan Release: Application to Glycans from Engineered Glycoforms of Intact, Folded HIV gp120. J. Am. Soc. Mass Spectrom. 22, 568–581 (2011). https://doi.org/10.1007/s13361-010-0053-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13361-010-0053-0