
Abstract
The MDM4 protein (also known as MDMX or HDMX) is a negative regulator of p53, not only by direct interaction but also through its interaction with MDM2. Further, MDM4 overexpression and amplification have been observed in several cancer forms. Recently, a single nucleotide polymorphism (SNP) in the 3’ untranslated region of the MDM4 gene, SNP34091A > C (rs4245739) was reported to alter MDM4 messenger RNA (mRNA) stability by modulating a microRNA binding site, thereby leading to decreased MDM4 levels. In this case-control study, we aimed to evaluate the possible association between MDM4 SNP34091 status and cancer risk by comparing the genotype frequencies in large hospital-based cohorts of endometrial- (n = 1404) and ovarian (n = 1385) cancer patients with healthy female controls (n = 1870). Genotype frequencies were compared by odds ratio (OR) estimates and Fisher exact tests. We found that individuals harboring the MDM4 SNP34091AC/CC genotypes had a significantly elevated risk for serous ovarian cancer (SOC) in general and high-grade serous ovarian cancer (HGSOC) in particular (SOC: OR = 1.18., 95 % CI = 1.01–1.39; HGSOC: OR = 1.25, CI = 1.02–1.53). No association between SNP34091 genotypes and endometrial cancer risk was observed. Our data indicate the MDM4 SNP34091AC/CC genotypes to be associated with an elevated risk for SOC and in particular the HGSOC type.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Maintaining the correct levels of p53 is imperative to cell survival and normal tissue homeostasis, and thus, the p53 protein plays a pivotal role in cancer biology [1]. The protein product of the murine double minute 2 gene, MDM2, and its homologue MDM4 (also referred to as MDMX or HDMX) are known to be the major negative regulators of p53 [2]. While both MDM2 and MDM4 inhibits p53 by direct binding and masking of its transactivation domain [3–6], only MDM2 possesses E3 ubiquitin ligase activity and may downregulate p53 by targeting it for ubiquitin-proteasome-dependent degradation [7–9]. However, heterodimerization with MDM4 enhances MDM2’s E3 ligase activity towards p53 [10, 11]. Taken together, these data suggest that elevated levels of MDM4 can prevent p53-mediated tumor suppression. In line with this, the MDM4 gene has been found amplified and overexpressed in several cancer forms (reviewed in [12]), and studies in transgenic mice have shown that overexpression of Mdm4 induced spontaneous tumor formation and accelerated tumorigenesis [13].
In the last decades, several single nucleotide polymorphisms in the MDM2 [14, 15] as well as the MDM4 [16–18] loci have been associated with elevated or reduced cancer risk, although data are at variance. Recently, a single nucleotide polymorphism (SNP) in the MDM4 3’ untranslated region, MDM4 SNP34091A > C (rs4245739) was reported to affect MDM4 messenger RNA (mRNA) stability and protein levels [19, 20]. The SNP34091C variant creates a functional target site for hsa-miR-191, and both ovarian and prostate cancer cells harboring the C- allele displayed reduced MDM4 mRNA and protein levels [19, 20].
Conflicting evidence has linked rs4245739 genotypes to breast cancer risk. Thus, while case-control studies suggested the SNP34091C-allele to be associated with a reduced risk for breast cancer in general [17, 21] and among individuals carrying a MDM2 SNP309GG genotype in particular [17], recent genome-wide association studies (GWAS) have reported the C-allele to be associated with an increased risk for estrogen receptor (ER) negative and, in particular, triple-negative breast cancer [22–24]. Regarding other malignancies, the C-allele has been associated with a reduced risk of non-Hodgkin lymphoma [25], esophageal squamous cell carcinoma [26], and prostate cancer [27] but not with risk of cancer of the lung or colon [17].
In this study, we assessed the impact of MDM4 SNP34091 status on the risk of ovarian and endometrial cancer in large hospital-based sample sets.
Materials and methods
Study population
In this study, we successfully genotyped the MDM4 SNP34091A > C (rs4245739) status in endometrial cancer samples (n = 1404) obtained from patients treated at Haukeland University Hospital during 2001–2009 [28] and from patients included in the MoMaTEC (Molecular Markers in Treatment of Endometrial Cancer) study between 2007–2010 [29]. Further, we genotyped ovarian cancer samples (n = 1385) from patients treated at the Oslo University Hospital Radium Hospital in the period between 1993 and 2011 [15]. Notably, patients with known mutations related to hereditary ovarian cancer (BRCA1 or BRCA2) were excluded from the analysis.
For the endometrial cancer samples we had histological and FIGO status for 1321 and 1238 patients, respectively. As for the ovarian cancer, the grade of differentiation (high-grade serous ovarian cancer (HGSOC), low-grade serous ovarian cancer (LGSOC), clear cell ovarian cancer, endometrioid ovarian cancer, and mucinous ovarian cancer) was available for 1071 patients.
In order to evaluate OR for ovarian and endometrial cancer related to MDM4 SNP34091 status, we compared our findings to the SNP status among 1870 healthy controls. These were the female fraction of a sample set of 3747 healthy individuals, previously genotyped [17], and they were all obtained from the population-based Cohort of Norway (CONOR) study [30].
MDM4 SNP34091 genotyping
All samples were genotyped for MDM4 SNP34091 status using a custom LightSNiP assay (TIB MOLBIOL Syntheselabor GmbH, Berlin, Germany) on a LightCycler 480 II instrument (Roche, Basel, Switzerland) as previously described in detail [17].
Statistical analysis
Potential deviations from Hardy-Weinberg equilibrium were assessed by calculating the expected genotype distribution based on the observed allele frequencies and comparing the output with the observed genotype distribution using chi-square tests.
Potential associations between MDM4 SNP34091 and risk of ovarian and endometrial cancer as well as cancer risk within different subgroups were estimated by calculating odds ratios (OR) with 95 % confidence intervals (CI) and by Fisher’s exact tests.
All statistical analyses were performed using the IBM SPSS 22 software (IBM Corp, Armonk, NY, USA). p values are given as two-sided and p values from Fisher’s exact tests are given as cumulative.
Results
Distribution of MDM4 SNP34091
Among the 1870 healthy female controls previously genotyped [17], we recorded a minor allele frequency (MAF) of 0.27. Regarding the present analyses, MDM4 SNP34091 status was successfully genotyped in 1385 ovarian and 1404 endometrial cancers cases.
The genotype frequencies were found to be in Hardy-Weinberg equilibrium (p > 0.8 for all comparisons). A comprehensive overview of the MDM4 SNP34091 distribution in the healthy controls as well as the two cancer types analyzed is given in Table 1.
MDM4 SNP34091 status and cancer risk in ovarian cancer
In order to estimate the potential impact of MDM4 SNP34091 status on ovarian cancer risk, we compared the frequency of the MDM4 SNP34091 genotypes among ovarian cancer patients (n = 1385) to healthy female controls (n = 1870). Although we observed no significant association between MDM4 SNP34091 status and ovarian cancer risk, applying the dominant model (SNP34091CC + AC vs. AA) we observed a non-significant association with increased risk for ovarian cancer (OR = 1.12; 95 % CI = 0.98–1.29; Table 1).
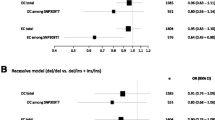
Following the observation of a potential association between the SNP34091CC/AC genotypes and increased risk of ovarian cancer in general, we performed separate analyses for the different subgroups of the disease with respect to histology class and grade of differentiation. By doing so, we observed a significant association between carriers of the SNP34091C allele (dominant model) and increased risk of serous ovarian cancer (OR = 1.18; 95 % CI = 1.01–1.39), but not any increased risk for clear cell, endometrioid, or mucinous ovarian cancers (Table 2, Fig. 1). Further, by stratifying serous ovarian carcinoma into the high-grade and low-grade type, we found that the increased risk conferred by the SNP34091C allele was highest in HGSOC (OR = 1.25; 95 % CI = 1.02–1.53; Table 2, Fig. 1).

Impact of MDM4 SNP34091 on ovarian cancer risk. Forest plot showing the effect of SNP34091 in the different histological ovarian cancer types as compared to healthy female controls. LGSOC low-grade serous ovarian cancer, HGSOC high-grade serous ovarian cancer
MDM4 SNP34091 status and cancer risk in endometrial cancer
Comparing the genotypes of 1404 endometrial cancer patients to the 1870 healthy controls, no association between MDM4 SNP34091 status and endometrial cancer were observed, either by applying the dominant or the recessive model (Table 1).
We further stratified the endometrial cancer patients according to histological and FIGO status; however, no association between SNP34091 genotypes and endometrial cancer risk was observed in any of the subgroups (data not shown).
MDM4 SNP34091 and interaction with MDM2 SNP309
In the same sample set as analyzed in the present study, we have previously reported the genotypes for the two most widely studied functional SNPs in the MDM2 gene (MDM2 SNP285; rs117039649 and SNP309; rs2279744) [15, 28]. Since MDM4 and MDM2 act together in inhibiting the tumor suppressor function of p53, we investigated potential interactions/synergistic effects between MDM4 SNP34091 and MDM2 SNPs with respect to cancer risk.
Assessing ovarian cancer in general, we found a moderate synergistic effect of SNPs in the two genes. However, when restricting the analyses to HGSOC, we found particularly high risk of disease among individuals with the MDM4 SNP34091C-allele and MDM2 SNP309TT genotype (OR = 1.41; 95 % CI = 1.02–1.94; Supplementary Table S1).
In contrast, we found no synergistic effects between MDM4 SNP34091 and MDM2 SNPs with respect to endometrial cancer risk (data not shown).
MDM4 expression levels in ovarian and endometrial cell lines
Since we found an effect of MDM4 SNP34901 in ovarian—but not endometrial cancer, we mined the publically available data set from the Broad-Novartis Cancer Cell Line Encyclopedia (CCLE–Broad Institute; www. broadinstitute.org/ccle/home). Comparing the available data, we found the average MDM4 expression level among ovarian cancer cells (n = 51) to be significantly lower than endometrial cancer cells (n = 27; p = 0.003), indicating that ovarian cells may be more sensitive to subtle changes in MDM4 levels than endometrial cancer cells.
Discussion
In this study, we examined the association between the MDM4 SNP34091 status and ovarian- and endometrial cancer risk applying a case-control design. This is, to the best of our knowledge, the first case-control study estimating the effect of MDM4 SNP34091 on risk for endometrial cancer and, although a small study found an increased risk for relapse and early onset of ovarian cancer among individuals carrying the SNP34091A allele [20], no study has evaluated MDM4 SNP34091 status as a potential risk factor with respect to ovarian cancer.
In our overall analyses, we observed no association between MDM4 SNP34091 status and the risk for endometrial cancer, but a non-significant association between the SNP34091C allele and an increased risk of ovarian cancer. Notably, individuals carrying the SNP34091C allele had significantly increased risk for developing serous ovarian cancer, and in particular tumors of the HGSOC type, compared to individuals harboring the SNP34091AA genotype. The risk was particularly high among individuals carrying the MDM2 SNP309TT genotype. Although risk assessments are not directly comparable to survival analysis, our findings may seem somewhat contradictory to the report of the SNP34091A allele as a risk factor for recurrence and tumor-related death in ovarian cancer patients [20]. While the SNP34091A allele has been reported to confer higher MDM4 levels in ovarian [20] and prostate cancer cells [19], and the common assumption is that the oncogenic effect of high MDM4 levels is through the p53 pathway [31], it has been reported that over 90 % of all HGSOC have mutations in the TP53 gene [32]. Thus, it may be that the effect of MDM4 SNP34091 on ovarian cancer risk is mediated via additional pathways, other than p53.
While evidence linking SNP34091 status to breast cancer risk has been at variance [21–24], we recently found the MDM4 SNP34091AC genotype to be associated with a reduced risk of breast cancer among individuals carrying the MDM2 SNP309GG genotype [17], a genotype, in general, associated with elevated cancer risk [33, 34]. In the same study, no association between SNP34091 status and risk for cancer of the lung, colon, or prostate was observed [17]. Our findings in the present study are, however, in line with previous studies reporting SNP34091C to be associated with increased risk for triple-negative breast cancer [23, 24], a subclass of breast cancers sharing some mutational features with HGSOC [35]. The tissue-specific effects observed are also in line with the previously observed effect of the MDM2 SNP285G > C; where the C-allele is proposed to reduce the risk for ovarian, endometrial, and breast cancer, but not cancer of the prostate, lung, or colon [15, 28, 36, 37]. Notably, among cell lines registered in Cancer Cell Line Encyclopedia (CCLE–Broad Institute), we found a lower average MDM4 expression level among ovarian—than endometrial cancer cells. One may, therefore, speculate that ovarian cells in general are more sensitive than endometrial cells to subtle changes in the MDM4 levels, such as those induced by the different SNP34901 genotypes.
Previous candidate gene case-control studies assessing the effect of the SNP34091 on cancer susceptibility has been performed mainly in populations of Chinese ethnicity [21, 25, 26], and they have reported the SNP34091C-allele to be associated with a reduced risk of cancer. Notably, there is a substantial difference in the distribution of this SNP between Europeans and Asians with a MAF of 0.26 and 0.05, respectively [38]. This is also the case for the MDM2 promoter SNPs, SNP309, and SNP285: while the SNP309G allele is associated with an increased cancer risk, predominantly, among individuals of Asian ancestry [33, 34], the SNP285C-allele, which is associated with reduced cancer risk, [15, 28, 36], is absent in Asians and may therefore have a confounding effect on SNP309 risk estimates performed in Caucasian populations [39]. Thus, the somewhat variable results regarding MDM4 SNP34091 and cancer risk may also be explained by yet unknown functional SNP (s) that are in linkage disequilibrium (LD) with SNP34091.
A few years ago, Atwal and colleagues reported the MDM4 haplotype diversity across ethnic populations. They found a much greater diversity among individuals of African American and Ashkenazi Jewish ancestry than Caucasians ancestry. Further, they reported the SNP rs1563828T allele, which is in LD with the SNP34091A allele among Caucasians, to be associated with early onset of both familial and sporadic ovarian cancer [16]. To the best of our knowledge, the biological effects of SNP rs1563828 have not been elucidated; thus, the possibility exist that it may be SNP34091, which is known to have a biological effect [19, 20], that contributes to the observed effect of SNP rs1563828 previously reported by Atwal and colleagues.
In conclusion, we find the MDM4 SNP34091 to be associated with increased risk of SOC, in particular the HGSOC type, and in particular among individuals carrying the MDM2 SNP309TT genotype. In contrast, no effect on endometrial cancer risk was recorded. Although the observed ORs are too low to argue for any clinical use of MDM4 SNP status, further studies are warranted in order to reveal whether it could be a useful marker in any subgroup of cancers.
References
Lane DP. p53, guardian of the genome. Nature. 1992;358:15–6.
Wade M, Li YC, Wahl GM. MDM2, MDMX and p53 in oncogenesis and cancer therapy. Nat Rev Cancer. 2013;13:83–96.
Oliner JD, Pietenpol JA, Thiagalingam S, Gyuris J, Kinzler KW, Vogelstein B. Oncoprotein MDM2 conceals the activation domain of tumour suppressor p53. Nature. 1993;362:857–60.
Shvarts A, Bazuine M, Dekker P, Ramos YF, Steegenga WT, Merckx G, et al. Isolation and identification of the human homolog of a new p53-binding protein. Mdmx Genomics. 1997;43:34–42.
Shvarts A, Steegenga WT, Riteco N, van Laar T, Dekker P, Bazuine M, et al. MDMX: a novel p53-binding protein with some functional properties of MDM2. EMBO J. 1996;15:5349–57.
Thut CJ, Goodrich JA, Tjian R. Repression of p53-mediated transcription by MDM2: a dual mechanism. Genes Dev. 1997;11:1974–86.
Haupt Y, Maya R, Kazaz A, Oren M. Mdm2 promotes the rapid degradation of p53. Nature. 1997;387:296–9.
Honda R, Tanaka H, Yasuda H. Oncoprotein MDM2 is a ubiquitin ligase E3 for tumor suppressor p53. FEBS Lett. 1997;420:25–7.
Kubbutat MHG, Jones SN, Vousden KH. Regulation of p53 stability by Mdm2. Nature. 1997;387:299–303.
Kawai H, Lopez-Pajares V, Kim MM, Wiederschain D, Yuan ZM. RING domain-mediated interaction is a requirement for MDM2’s E3 ligase activity. Cancer Res. 2007;67:6026–30.
Linares LK, Hengstermann A, Ciechanover A, Muller S, Scheffner M. HdmX stimulates Hdm2-mediated ubiquitination and degradation of p53. Proc Natl Acad Sci U S A. 2003;100:12009–14.
Toledo F, Wahl GM. Regulating the p53 pathway: in vitro hypotheses, in vivo veritas. Nat Rev Cancer. 2006;6:909–23.
Xiong S, Pant V, Suh YA, Van Pelt CS, Wang Y, Valentin-Vega YA, et al. Spontaneous tumorigenesis in mice overexpressing the p53-negative regulator Mdm4. Cancer Res. 2010;70:7148–54.
Bond GL, Hu W, Bond EE, Robins H, Lutzker SG, Arva NC, et al. A single nucleotide polymorphism in the MDM2 promoter attenuates the p53 tumor suppressor pathway and accelerates tumor formation in humans. Cell. 2004;119:591–602.
Knappskog S, Bjornslett M, Myklebust LM, Huijts PE, Vreeswijk MP, Edvardsen H, et al. The MDM2 promoter SNP285C/309G haplotype diminishes Sp1 transcription factor binding and reduces risk for breast and ovarian cancer in Caucasians. Cancer Cell. 2011;19:273–82.
Atwal GS, Kirchhoff T, Bond EE, Montagna M, Menin C, Bertorelle R, et al. Altered tumor formation and evolutionary selection of genetic variants in the human MDM4 oncogene. Proc Natl Acad Sci U S A. 2009;106:10236–41.
Gansmo LB, Romundstad P, Birkeland E, Hveem K, Vatten L, Knappskog S, et al. MDM4 SNP34091 (rs4245739) and its effect on breast-, colon-, lung-, and prostate cancer risk. Cancer Med. 2015;4:1901–7.
Kulkarni DA, Vazquez A, Haffty BG, Bandera EV, Hu W, Sun YY, et al. A polymorphic variant in human MDM4 associates with accelerated age of onset of estrogen receptor negative breast cancer. Carcinogenesis. 2009;30:1910–5.
Stegeman S, Moya L, Selth LA, Spurdle AB, Clements JA, Batra J. A genetic variant of MDM4 influences regulation by multiple microRNAs in prostate cancer. Endocr Relat Cancer. 2015;22:265–76.
Wynendaele J, Bohnke A, Leucci E, Nielsen SJ, Lambertz I, Hammer S, et al. An illegitimate microRNA target site within the 3′ UTR of MDM4 affects ovarian cancer progression and chemosensitivity. Cancer Res. 2010;70:9641–9.
Liu J, Tang X, Li M, Lu C, Shi J, Zhou L, et al. Functional MDM4 rs4245739 genetic variant, alone and in combination with P53 Arg72Pro polymorphism, contributes to breast cancer susceptibility. Breast Cancer Res Treat. 2013;140:151–7.
Garcia-Closas M, Couch FJ, Lindstrom S, Michailidou K, Schmidt MK, Brook MN, et al. Genome-wide association studies identify four ER negative-specific breast cancer risk loci. Nat Genet. 2013;45:392–8. 8e1-2.
Purrington KS, Slager S, Eccles D, Yannoukakos D, Fasching PA, Miron P, et al. Genome-wide association study identifies 25 known breast cancer susceptibility loci as risk factors for triple-negative breast cancer. Carcinogenesis. 2014;35:1012–9.
Stevens KN, Vachon CM, Couch FJ. Genetic susceptibility to triple-negative breast cancer. Cancer Res. 2013;73:2025–30.
Fan C, Wei J, Yuan C, Wang X, Jiang C, Zhou C, et al. The functional TP53 rs1042522 and MDM4 rs4245739 genetic variants contribute to non-Hodgkin lymphoma risk. PLoS One. 2014;9, e107047.
Zhou L, Zhang X, Li Z, Zhou C, Li M, Tang X, et al. Association of a genetic variation in a miR-191 binding site in MDM4 with risk of esophageal squamous cell carcinoma. PLoS One. 2013;8, e64331.
Eeles RA, Olama AA, Benlloch S, Saunders EJ, Leongamornlert DA, Tymrakiewicz M, et al. Identification of 23 new prostate cancer susceptibility loci using the iCOGS custom genotyping array. Nat Genet. 2013;45:385–91. 91e1-2.
Knappskog S, Trovik J, Marcickiewicz J, Tingulstad S, Staff AC. MoMa TECsg et al. SNP285C modulates oestrogen receptor/Sp1 binding to the MDM2 promoter and reduces the risk of endometrial but not prostatic cancer. Eur J Cancer. 2012;48:1988–96.
Trovik J, Wik E, Stefansson IM, Marcickiewicz J, Tingulstad S, Staff AC, et al. Stathmin overexpression identifies high-risk patients and lymph node metastasis in endometrial cancer. Clin Cancer Res. 2011;17:3368–77.
Naess O, Sogaard AJ, Arnesen E, Beckstrom AC, Bjertness E, Engeland A, et al. Cohort profile: cohort of Norway (CONOR). Int J Epidemiol. 2008;37:481–5.
Danovi D, Meulmeester E, Pasini D, Migliorini D, Capra M, Frenk R, et al. Amplification of Mdmx (or Mdm4) directly contributes to tumor formation by inhibiting p53 tumor suppressor activity. Mol Cell Biol. 2004;24:5835–43.
Integrated genomic analyses of ovarian carcinoma. Nature. 2011;474:609-15.
Economopoulos KP, Sergentanis TN. Differential effects of MDM2 SNP309 polymorphism on breast cancer risk along with race: a meta-analysis. Breast Cancer Res Treat. 2010;120:211–6.
Hu Z, Jin G, Wang L, Chen F, Wang X, Shen H. MDM2 promoter polymorphism SNP309 contributes to tumor susceptibility: evidence from 21 case-control studies. Cancer Epidemiol Biomarkers Prev. 2007;16:2717–23.
TCGA. Comprehensive molecular portraits of human breast tumours. Nature. 2012;490:61-70.
Gansmo LB, Knappskog S, Romundstad P, Hveem K, Vatten L, Lonning PE. Influence of MDM2 SNP309 and SNP285 status on the risk of cancer in the breast, prostate, lung and colon. Int J Cancer. 2015;137:96–103.
Ryan BM, Calhoun KM, Pine SR, Bowman ED, Robles AI, Ambs S, et al. MDM2 SNP285 does not antagonize the effect of SNP309 in lung cancer. Int J Cancer. 2012;131:2710–6.
Genomes Project C, Abecasis GR, Auton A, Brooks LD, DePristo MA, Durbin RM, et al. An integrated map of genetic variation from 1,092 human genomes. Nature. 2012;491:56–65.
Knappskog S, Gansmo LB, Dibirova K, Metspalu A, Cybulski C, Peterlongo P, et al. Population distribution and ancestry of the cancer protective MDM2 SNP285 (rs117039649). Oncotarget. 2014;5:8223–34.
Acknowledgments
Most of this work was performed in the Mohn Cancer Research Laboratory, Haukeland University Hospital/University of Bergen. This study was supported by grants from the Bergen Research Foundation, the Norwegian Cancer Society’s Pink Ribbon Campaign, the Norwegian Health Region West, and the Norwegian Research Council. The authors thank Beryl Leirvaag for her skilled technical assistance.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical Approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards. The study was approved by the Regional Committee for Ethics in Medical Research.
Informed Consent
Informed consent was obtained from all individual participants included in the study.
Conflicts of interest
None
Additional information
Helga B. Salvesen deceased.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Table S1
(DOCX 15 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Gansmo, L.B., Bjørnslett, M., Halle, M.K. et al. The MDM4 SNP34091 (rs4245739) C-allele is associated with increased risk of ovarian—but not endometrial cancer. Tumor Biol. 37, 10697–10702 (2016). https://doi.org/10.1007/s13277-016-4940-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13277-016-4940-2