
Abstract
Morocco is a very rich country in terms of wetlands, including bays, estuaries, lagoons, lakes, and rivers, among others. However, many of them need to be well managed and better exploited, thus requiring baseline descriptive studies to assess faunal patterns and trends. Oualidia lagoon is not an exception. In fact, the studies conducted to date in the lagoon mainly focused on its oceanographic features and aquaculture capacities. The present paper presents the first study on the whole macrofaunal assemblages of Oualidia lagoon, having as main objectives analyze the composition and structure of the assemblages, as well as their relationships with the main environmental variables explaining their spatial distribution variability in the enclosed ecosystem of the Oualidia coastal lagoon. The benthic macroinvertebrates of Oualidia lagoon were studied in summer 2013 based on 43 stations spread over the whole lagoon by using a Van Veen grab. Two replicate samples were collected for macrobenthos and one additional replicate for sediment analyses at each station. Also, single water salinity and temperature measurements were recorded at each station. Fifty-one species from seven different taxa were recorded, among which mollusks (37%), polychaetes (25%) and crustaceans (25%) dominated in terms of species number. Peringia ulvae, Abra alba, Sphaeroma serratum, Cerastoderma edule and Corophium sp. accounted for 88% of total abundance. The benthos was structured along a downstream–upstream gradient, showing three assemblages, named according to the IndVal index: (1) Diopatra cf. marocensis, (2) Peringia ulvae, and (3) Cerastoderma edule, while the main factors driving their distribution were granulometry and salinity. The Biotic and Environmental analysis showed that the combination of granulometry and salinity was the major factors controlling the spatial distribution of the macrozoobenthos in the Oualidia Lagoon.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Coastal lagoons are highly productive ecosystems acting as feeding, nursery, reproduction, and protection spots for invertebrates, fishes, and birds, as well as rich fishery grounds (Newton et al. 2018). However, they are fragile ecosystems, highly susceptible to both natural and anthropogenic disturbances (Blanchet et al. 2008; Rossi et al. 2006; Affian et al. 2009).
Many interactions occurring in coastal lagoons are mediated through macrobenthic organisms which: (1) contribute to mineralize, promote, and mix the oxygen flux into the sediment, (2) recycle the organic matter and nutrients, (3) decompose detrital matter, (4) stabilize sediments, and (5) contribute to the trophic web by linking producers to higher consumers such as fishes, birds and epibenthic crustaceans (Pearson and Rosenberg 1978; Nybakken 1993; Heilskov and Holmer 2001; Newton et al. 2018). Such a relevant role, together with their sensitivity to human disturbance, leads these organisms to be often employed as indicators of coastal ecosystems health, and coastal lagoons are not an exception (Dauer 1993; Borja et al. 2000; Salas et al. 2004). The relatively limited capacity of dispersion and sedentary habits of adult benthic organisms does not allow them to easily avoid unfavorable environmental conditions. Consequently, they are considered as sensitive indicators of both naturally and anthropogenically driven changes in coastal ecosystems (Salas et al. 2004; Borja et al. 2000). Moreover, their relatively long lifespan allows them to integrate water and sediment quality conditions through time, as well as to show responses to temporal and chronic disturbances. Particularly, both stress tolerant and sensitive benthic species may show changes in diversity, biomass and abundance, while their assemblages may show changes in trophic and functional structure (Pearson and Rosenberg 1978; Reiss and Kröncke 2005).
The above-mentioned relationships between benthic organisms and environmental conditions in coastal lagoons have been seldom analyzed along Moroccan coasts (El Asri et al. 2018, 2021). Nevertheless, the country has a rich network of lagoons, particularly in the Atlantic littoral where Oualidia lagoon is located. Oualidia is the most important wintering area for migratory birds in Morocco (El Hamoumi et al. 2003) and a Wetland of International Importance (under the RAMSAR Convention). It also provides a valuable environment for a rich variety of plants, fish, and other wildlife (El Hamoumi et al. 2003) and is currently considered a Natural Park. Moreover, the lagoon watershed supports intensive agricultural, cattle rearing, shell-fishing and tourism activities being thus an on-going economical pole (Maanan et al. 2017). Despite its intrinsic interest, most previous studies in Oualidia focused on the effects of pollution (e.g., Zourarah et al. 2007; Hassou et al. 2014; Maanan et al. 2014), while only one described its hydrographic functioning (Hilmi et al. 2005) and four analyzed specific macrobenthic taxa (Chbicheb 1996; El Asri et al. 2015, 2018, 2021).
The present paper presents the first study on the whole macrofaunal assemblages of Oualidia lagoon, having as main objectives: (1) to describe the species composition and abundance, (2) to analyze the assemblage structure and distribution, and (3) to identify the main environmental variables driving the assemblage structure and distribution.
Materials and methods
Study area
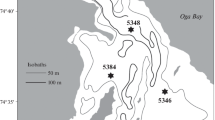
Oualidia lagoon, located in the Moroccan Atlantic Ocean coast (32°44′42″ N, 9°02′50″ W, Fig. 1), measures 7 km long and about 1 km wide. Characteristically, it shows numerous side channels connected to a meandering main channel with 2 m of average depth and 5 m of maximum depth during flood tides (Bidet and Carruesco 1982). Flood tides cover more than 75% (2.25 km2) of the lagoon surface, bringing salt water to its upstream reaches and into a saline marsh beyond its second dam (Maanan et al. 2014; Hilmi et al. 2017). Salinity ranges from 30 to 36% in the inner areas with very low mixing and very weak stratification of the water column (Makaoui et al. 2018). The rainfall reached c.a. 250 mm per year, and the prevailing winds are W–NW and E, almost parallel to the major lagoon axis (Zourarah et al. 2007).

Map showing the location of the study area and the sampling sites in Oualidia lagoon
Sampling and data analysis
Forty-three stations were sampled in July 2013 (Fig. 1) using a Van Veen grab (0.125 m2 surface area). Two replicate samples were collected for macrobenthos and one additional replicate for sediment analyses at each station. Also, single water salinity and temperature measurements were recorded at each station with the help of a thermo-salinometer. The macrobenthos replicates were sieved in situ through a 1 mm pore-sized mesh, and the retained materials were transferred to plastic containers and fixed in a 10% formalin solution. The sediment replicates were transferred to plastic contained and kept cold until analyzed.
In the laboratory, the macrofauna was sorted under a binocular microscope, identified at the lowest taxonomic level possible, counted and classified into trophic groups (e.g., Fauchald and Jumars 1979; Hily and Bouteille 1999; Pranovi et al. 2000; Afli et al. 2008; Khedhri et al. 2015) as follows: (1) herbivores (H), algae-feeding organisms (e.g., some echinoids), (2) scavengers, feeding on carrions deposited on the bottom (essentially gastropods and decapods), (3) detritus feeders, feeding on particulate organic matter, essentially vegetable detritus (mainly amphipods and tanaids), (4) carnivores, predators like errant polychaetes or sea-anemones, (5) suspension feeders, feeding on food suspended in the water column (e.g., most bivalves and some polychaetes), (6) surface deposit feeders, feeding on organic particles settling on the sediment (most sedentary polychaetes), and (7) subsurface deposit feeders, burrowers that ingest the sediment from which they extract their food.
Sediment replicates were used to measure: (1) chlorophyll ‘a’contents (mg/m2, Lorenzen method, Holm-Hansen et al. 1965), (2) percentages of the total organic matter (TOM, % of weight loss on ignition 4 h at 450 °C), and (3) grain size (µm, laser granulometer Malvern, Mastersizer).
Macrobenthic spatial distribution and biodiversity were described by the species richness (S), abundance (A), Shannon diversity (H′, as log2) (Shannon 1948) and evenness (J′) (Pielou 1966), as estimated with the Paleontological Statistics program (PAST v2.14) (Hammer et al. 2001). The structure of the assemblages was analyzed by a Hierarchical Ascending Classification (HAC) on a density [transformed to log10 (x + 1) to limit the influence of the most dominant taxa] per station matrix using the Bray–Curtis distance and the Ward’s method. Each identified assemblage was classified according to the IndVal index (Dufrêne and Legendre 1997). Visually observed patterns and correlations of species density with abiotic parameters were quantitatively explored with the BIOENV (Biotic and Environmental linking) routine, based on Euclidean distances (for the abiotic dataset) and Spearman’s rank correlations. All multivariate analyses were executed on an R environment (www.r-project.org). The results were plotted on maps using ARCGIS 10.0 (Figs. 2, 3).

Temperature (a) and salinity (b) trends in Oualidia lagoon

Spatial distribution of organic matter (a), chlorophyll ‘a’ (b), and granulometry (c) in Oualidia lagoon
Results
Macrofaunal descriptors
A total of 51 species of macrobenthic invertebrates were identified (Table 1), of with mollusks were the most diverse group (19 species, 37%), followed by polychaetes and crustaceans (13, 25.5% each) and echinoderms (three, 5.9%). Cnidarians, insects and tunicates (one species each) constituted a 6.0%. Mollusks clearly dominate in abundance (77% of all specimens recorded), followed by far by crustaceans (15%), polychaetes (5%), and echinoderms, cnidarians, insects and tunicates (3% altogether).
Density ranged from 0 to 6336 individuals per m2 (1274 ± 1469 ind. m−2, mean ± SD). The stations showing the highest densities were roughly situated in the center of the lagoon (Fig. 4a). The number of species per station ranged from 0 (station 7) to 18 (station 40), with 7.4 ± 3.7 species (mean ± SD). The stations showing the highest richness were roughly situated close to the inlet of the lagoon (Fig. 4b). Diversity (0–3.8 bits, 1.6 ± 0.7 bits, mean ± SD) and evenness (0.9, 0.6 ± 0.2, mean ± SD) showed both a heterogeneous distribution (Fig. 5a and b).

Spatial distribution of macrofauna abundance (a) and species richness (b) in Oualidia lagoon

Spatial distribution of Shannon (a) and evenness (b) indexes in Oualidia lagoon
The “best combination” of abiotic parameters explaining the observed density patterns was obtained by combining granulometry and salinity (BIOENV, R = 0.49) (Table 2).
Assemblage structure
Three groups of benthic organisms were identified in the lagoon (HAC, Figs. 6, 7). Group 1 included six stations from the outer part, close to the inlet. It was characterized by sediments with median-coarser sand with low organic matter content, which were subject to strong tidal currents. The total number of species was 43, mainly distributed among mollusks (37%), crustaceans (28%) and polychaetes (28%) (Fig. 8a). The average density was low (529 ± 475 ind.m−2) comparing to other groups, and the dominant taxa were mollusks (58%) and crustaceans (25%) (Fig. 8b), while the average evenness was 0.7 ± 0.2 (Table 3). The assemblage was dominated in density by both surface deposit and suspension feeders (76.1%) (Fig. 9a), while carnivores (22%), detritus feeders (20%), surface deposit feeders (18%) and suspension feeders (18%) dominate in species richness (Fig. 9b). The most dominant species were the gastropod Peringia ulvae (31.7%), the crustacean Corophium sp. (24%), followed by far by the bivalve Abra alba (18%).

Dendrogram showing the groups of stations derived from the cluster analysis based on macrofaunal abundances in Oualidia lagoon. I, group 1; II, group II; III, group III

Spatial distribution of the macrofaunal communities in Oualidia lagoon

Faunal composition structure of the macrofauna community expressed in relative number of species (a) and individuals (b)

Trophic structure of the macrofauna community expressed in relative number of species (a) and individuals (b). Surface deposit feeders (SDF), Subsurface deposit feeders (SSDF), Detritus feeders (Dt), Suspension feeders (S), Carnivores (C), Scavengers (N), Herbivores (H)
Group 2 included 21 stations mainly located inwards, along the middle area of the lagoon, and characterized by having mixed silty-sand and sandy-silt sediments. The average density was 1712 ± 1644 ind.m-2, and mollusks were largely the most dominant, with more than 90% of the total (Fig. 8b). There were 25 species, most of which belonging to crustaceans (11.44%) and mollusks (9. 36%) (Fig. 8a) and the evenness was low (0.5 ± 0.2) (Table 3). The assemblage was dominated in density by surface deposit feeders (ca. 80% of the total density), while other trophic groups were present with very low densities (Fig. 9b). Surface deposit feeders also dominate in number of species (24%), but were closely followed by detritus feeders (22%) and scavengers (18%) (Fig. 9a). The most dominant species were the gastropod P. ulvae (66%), followed by far by the bivalve A. alba (22%).
Group 3 included 16 stations mainly located in the most inner area of the lagoon and characterized by variably silty sediment with high organic matter content (in shallower bottoms) and muddy sediments (in deeper bottoms). The total number of species was 25, which were distributed mainly among crustaceans, mollusks and polychaetes (Fig. 8a). The mean density was 1,643 ± 1,641 ind.m−2, with the mollusks being the most dominant (62%), followed by crustaceans (31%). All remaining groups just represented a ≤ 2% (Fig. 8b). The evenness was 0.7 ± 0.1 (Table 3). The assemblage was dominated in density by surface deposit feeders (44.7%), followed by detritus feeders (26.3%) and suspension feeders (25.8%) and the other groups (≤ 3%) (Fig. 9b). Conversely, the number of species detritus feeders, suspension feeders and surface deposit feeders is 24% each (Fig. 9a). The most dominant species were A. alba (24.2%), Sphaeroma serratum (21.3%), P. ulvae (21%), Cerastoderma edule (16.9%) and Corophium sp. (4%).
Discussion
The present study represents a reliable baseline for future ecological research by providing a detail description of the observed spatial variability in summer conditions, as well as the relationships between the environmental factors, the biotic descriptors and the assemblage structure. The water temperature increased from outer to inner parts of the lagoon due to the influence of the cold ocean waters entering through the inlet (Fig. 2a). The highest salinities are observed mainly at the stations located downstream of the lagoon, directly subject to marine influences. The low values recorded upstream of the lagoon are due to the resurgences of fresh water which are responsible for the slight desalination of the lagoon water upstream. The low value observed at station 43 is explained by the dilution by fresh water from nearby water sources. Both granulometry (Fig. 3c) and organic matter (Fig. 3a) patterns mirrored that of temperature, revealing the relevant influence of the hydrodynamic functioning of the lagoon, with the inner area tending to be relatively calm, thus to having more fine sediments and organic matter (Hilmi et al. 2005; Maanan et al. 2014). Conversely, salinity decreased from outer to inner parts (Fig. 2b), where there were the main freshwater arrivals (Hilmi et al. 2005; Damsiri et al. 2014; Hassou et al. 2014). Moreover, also, a minimum of salinity downstream was detected (but in a single station), which was probably caused by the numerous underwater freshwater outbreaks located in this area of the lagoon (Rharbi et al. 2001; Hilmi et al. 2005).
This environmental framework characterizes Oualidia as a typical lagoon environment, where the benthic macrofauna is dominated by gastropods, bivalves, amphipods, isopods and polychaetes (Mistri 2002; Bazaïri et al. 2003). However, the presence of cnidarians, tunicates and insects was also recorded. The former two occurred likely in connection with the seaweed coverage, while the later was only present in the most enclosed parts of the lagoon, where the salinity was low due to the numerous underwater freshwater outbreaks.
The biodiversity in Oualidia largely doubled the previous record in the area by Chbicheb (1996), 51 versus 24, and was also higher than those reported in nearby lagoons (Mergaoui et al. 2003; Marchini et al.2004; Chaouti and Bayed 2005). This relatively higher number of species may be related to the combined influence of the open sea at the entrance, the underground freshwater seepage in the central area and the main freshwater inputs in the most enclosed area (Carruesco 1989; Hilmi et al. 2005; El Asri et al. 2015, 2018). This particular structure seemed to favor a mixed presence of marine, brackish and freshwater species in a lagoon environment (Zabi and Leloeuf 1993).
Our results based on the whole macrofauna in summer agree with the previously reported structuring based on mollusk (El Asri et al. 2015) and polychaete assemblages in summer and winter in Oualidia (El Asri et al. 2018) when were studied independently and, to some extent, with those of the whole macrofauna in winter (El Asri et al. 2021). In all cases, there were three assemblages clearly organized along the outer–inner axis of the lagoon. As reported for other lagoons, this type of assemblage structuring resulted from combining the particular habitat preferences of all macrobenthic species (Bazaïri et al. 2003; Lakhdar Idrissi et al. 2004; Chaouti and Bayed 2008; Rodrigues et al. 2011; Lefrere et al. 2015). However, lagoon environments generally show marked seasonal variability (e.g., Bertrán et al. 2016), and there are no reasons allowing us to suspect that Oualidia could be an exception. Therefore, to assess the possible seasonal differences in the macrofaunal assemblages of Oualidia, there is an on-going research attempting to compare the summer situation analyzed in the present work with the results of a previous study carried out in winter conditions (El Asri et al. 2021).
In Oualidia, the benthic assemblages conforming the three groups identified in the HAC were named according to the IndVal index (Table 3). The Diopatra cf. marocensis assemblage (Group 1) was mainly distributed in the outer part of the lagoon and mollusks, crustaceans and polychaetes were almost equally dominant. The species richness was the highest found in the lagoon, due to the presence of both lagoon and marine species, the later entering the lagoon environment thanks to the proximity to the inlet (Cherkaoui 2006; Koaudio et al. 2008). Such a proximity also influenced the assemblage density, with the low numbers being likely caused by the high hydrodynamics favoring sedimentary instability (El Asri et al. 2015). Nevertheless, the assemblage revealed to be well-balanced, as indicated by a relatively high evenness.
The Peringia ulvae assemblage (Group 2) was the largest in number of stations and occurred mainly in the middle area of the lagoon. The hydrodynamic regime was moderated and the environment showed a relatively high nutrient enrichment (Rharbi et al. 2001; Hilmi et al. 2005). These conditions favored some of the established species, which led this assemblage to show the maximum average density combines with a poor structuring (as indicated by the very low evenness). Likely, this also explained the extreme dominance (i.e., ca. 90% of the total density) of two mollusk species, P. ulvae that gave the name to the assemblage, and the bivalve A. alba.
The Cerastoderma edule assemblage (Group 3) was mainly distributed in the inner part of the lagoon (Fig. 6) and harbored the same number of species as the P. ulvae assemblage (i.e., 25). The average density was also relatively high, with ca. 90% being represented by mollusks and crustaceans. However, contrary to the P. ulvae assemblage, the structure was well balanced (as indicated by the relatively high evenness), likely in connection with the relatively high level of nutrient enrichment characterizing this zone (Rharbi et al. 2001).
It can be seen that there is a direct relationship between the nature of the sediment and the dominance of certain trophic groups. In fact, surface deposit feeders and subsurface deposit feeders are better represented on silted substrates than on sandy substrates, deposit feeders are favored by a low water current speed unlike suspensivores which require a stronger current (McLusky and McIntyre 1988). This further explains the very pronounced numerical presence of suspensivores at the level of downstream stations where the speed of the tidal currents is higher, these results were found in several coastal marine ecosystems (e.g., Dauvin 1988; Grall and Glémarec 1997; Bazaïri et al. 2003).
The overall structure of the macrobenthic population of the lagoon of Oualidia is similar to that of the lagoon of Smir (Chaouti 2003), but also to that of the lagoon of Valle di Gorino in Italy, for example (Mistri et al. 2002). It is characterized by a small number of abundant and constant species and by many rare species. This type of structure is found mainly in environments under environmental stress (Blondel 1979).
The dominant species (Peringia ulvae, Abra alba, Sphaeroma serratum, Corophium sp., Cerastoderma edule and Hediste diversicolor) are characteristic of the lagoon littoral environments and correspond to the paralic biocoenosis within the meaning of Guélorget and Perthuisot (1983). Macrofauna seems to be well represented in this type of environment. Quantitative data show that variations in overall density are mainly due to numerically dominant species.
The structure of the macrobenthic community of the Oualidia lagoon was approached through the calculation of several indices. The analysis of the structure of the populations of the Oualidia lagoon reveals a low specific diversity compensated by an increase in the density of species and by the widening of their spatial niche. According to Blondel (1979), this type of stand is found particularly in environments subject to restrictive conditions. Nevertheless, the values of the H 'index of less than 3 bits / Ind, encountered in this ecosystem, are characteristic of paucispecific populations present in pecilohaline lagoon environments such as the Prévost lagoon (Amanieu et al. 1977, 1978) and the Grau du Piemanson pond (Camargue) (Grimonet 1979).
The values of the structure indices showed a great spatial variation testifying to a great heterogeneity in terms of community structure. The maximum values of these indices are observed at the downstream part of the lagoon which is most influenced by the ocean. The marine influence manifests itself in species enrichment but also in the maintenance of a relatively stable environment allowing the establishment and organization of a more structured community. The central zone, on the other hand, has the lowest values of these structure indices. It is a transition zone toward the interior of the lagoon where the environment is relatively unstable. The macrobenthic community at this level is unstructured and strongly dominated by only two species (Peringia ulvae and Abra alba) which are described as surface deposits. The upstream zone has the characteristics of a confined zone where the number of species is reduced but with appreciable diversity and above all fairness. The reduction in the specific richness in this area is the consequence of a selection of species by this particular environment resulting in a stable and characteristic community of the area. Similar results have been obtained in other lagoons such as the Ria d'Aveiro lagoon in Portugal (Garnerot et al. 2004), the Sacca di Goro lagoon in northeastern Italy (Mistri et al. 2001), or even in Dypsø fjord in Denmark (Garnerot et al. 2004). At the level of the Bizerte lagoon in Tunisia, Belkhodja-Mahjoud et al. (2007) distinguished two different zones, one well structured with high diversity and regularity and another less structured where these indices are low reflecting a pronounced imbalance benthic populations. The work of Chaouti and Bayed (2005) showed weak structure indices over the whole of the Smir lagoon in Morocco, testifying to a weak structure of the benthic fauna throughout this lagoon.
The weak structuring of benthic communities observed in general in paralic environments is the direct cause of the importance of variations in abiotic factors such as salinity (Llanso et al. 2002; Teske and Wooldridge 2003; Giménez et al. 2005; Pasquaud 2006), temperature and hydrodynamics (Stora et al. 1995; Blanchet et al. 2005; Tlig-Zouari et al. 2008). Added to this is the sediment factor which plays an important role in the structuring of endogenous benthic communities (Gray 1981; Mannino and Montagna 1997; Teske and Wooldridje 2001; Ysebaert and Herman 2002). Other biotics such as the trophic environment can also intervene in the structural organization of benthic macrofauna (Solidoro et al. 2004).
The results suggest that the spatial patterns of the macrobenthic assemblages of Oualidia were mainly influenced by the salinity gradient and the sediment composition, with hydrodynamics also playing a relevant role. This agrees with previous results found in different lagoons and enclosed bays, such as the Epe Lagoon (Uwadiae 2013), the Guanabara Bay (Mendes et al. 2007), or the Paranagua Bay (Boehs et al. 2004). However, the environmental factors controlling species distribution in different paralic environments may vary. In the Smir lagoon (Chaouti and Bayed 2008), the vegetation and grain size were the major factors influencing the structure of species. In the Sacca di Goro (Mistri et al. 2001), the main control was attributed to dissolved oxygen, temperature and salinity. In the Monolimni lagoon (Kevrekidis 2004), the most influencing factors were temperature and depth. In the Ria de Aveiro (Rodrigues et al. 2011), the spatial distribution of the fauna was found to be mainly related to the hydrodynamic regime and the salinity gradient, while the sediment grain size was much less important. Conversely, in the Arcachon Bay (Do et al. 2011), the structure of benthic communities was largely influenced by the combination of sediment grain size distribution and seagrass presence. Concerning the vegetation found at the level of the lagoon, it is the halophytic phanerogams characteristic of brackish water "the eelgrass", which predominate the intertidal zone and at the level of the schorres of the lagoon of Oualidia, as well as the green algae "the seaweeds". ulves.” The phytoplankton is abundant and represented by five large groups (Bennouna 1999); indeed, surely, the vegetation has an effect on the distribution of the benthic macrofauna; however, it has not been studied and its impact pending to be determined.
All these paralic environments differ considerably from one another in morphology and water depth, which certainly gave rise to differences in hydrodynamic regime, temperature, salinity, granulometry, vegetal coverage, etc. This may certainly explain the reported changes in the environmental factors controlling species distribution, but also the differences in the composition and abundance of the established macrobenthos (Hernandez-Guevara et al. 2008). Moreover, the existing interactions between the macrobenthic fauna and other members of the ecosystem, including different components of the benthic compartment, but also all organisms inhabiting the lagoon (e.g., fish, birds, etc.) are certainly affecting the reported differences, particularly through predation, competition, etc. (Gambi et al. 1998; Nicolaidou et al. 2006; Schückel et al. 2015; Khedhri et al. 2017). Finally, one cannot discard the anthropogenic influences, which tend to be a major force in driving the functioning of most paralic environments all around the world (e.g., Vezzone et al. 2020; Fang et al. 2020) and, certainly, Oualidia is not an exception (Jayed et al. 2015; Lakhlalki el al. 2017).
Conclusion
This study on the analysis of the structural characteristics and spatial organization of macrobenthic fauna in Oualidia lagoon has contributed to 54,407 macrozoobenthic organisms from 51 species in Oualidia, with the gastropod Peringia ulvae, the bivalves Abra alba and Cerastoderma edule, the isopod Sphaeroma serratum and the amphipod Corophium sp. being the most abundant species. Overall, the macrobenthos was structured in three assemblages distributed along an inner–outer axis, with the observed patterns in assemblage composition and structure being mainly driven by granulometry and salinity, with the hydrodynamics also playing a relevant role. This study must be considered as a baseline for future monitoring programs on the coastal management in the Oualidia lagoon. Therefore, further investigations are needed to see the whole picture of biodiversity status of macrofauna in the Oualidia lagoon, and to monitor the ecosystem health, it would be very interesting to study the seasonal variability of the benthic community in the lagoon and its linkage to the fluctuating physical–chemical parameters. To maintain high biodiversity, it is necessary to develop a specific management plan that will take into account the importance of the shallow water habitats in maintaining the ecological functions of the lagoon including that of nursery for juvenile fishes.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Affian K, Robin M, Maanan M, Digbehi B, Djagoua EV, Kouamé F (2009) Heavy metal and polycyclic aromatic hydrocarbons in Ebrié lagoon sediments, Côte d’Ivoire. J Environ Monit 159:531–541. https://doi.org/10.1007/s10661-008-0649-z
Afli A, Ayari R, Brahim M (2008) Trophic organization of the macrozoobenthic assemblages within coastal areas subjected to anthropogenic activities. J Mar Biol Assoc UK 88:663–674. https://doi.org/10.1017/S0025315408001318
Amanieu M, Guelorget O, Michel P (1977) Richesse et diversité de la macrofaune benthique d’une lagune littorale méditerranéenne. Vie Milieu 27(85):109
Amanieu M, Do Chi T, Galzin R, Lasserre G (1978) Structure et importance des populations de crabe vert Carcinus mediterraneus dans l’étang de Prévost (Languedoc, France). Bull Ecol 9:329–341. https://doi.org/10.1080/19425120.2017.1310155
Bazaïri H, Bayed A, Glémarec M, Hily C (2003) Spatial organisation of macrozoobenthic communities in response to environmental factors in a coastal lagoon of the NW African coast (Merja Zerga, Morocco). Oceanol Acta 26:457–471. https://doi.org/10.1016/S0399-1784(03)00041-0
Belkhodja Mahjoub H, Rhomdane MS, Missaoui H (2007) Répartition spatiale de la faune malacologique de la lagune de Bizerte : analyses écologiques. Rapports De La Commission Internationale Pour L’exploration De La Mer Méditerranée 38:430
Bennouna A (1999) Etude du phytoplancton nuisible et de son environnement dans la lagune de Oualidia et Sidi Moussa (Maroc). Université Chouiab Doukkali, El Jadida (Maroc), Thèse de Doctorat, p 153
Bertrán C, Fierro P, Encalada E, Peña-Cortés F, Tapia J, Hauenstein E, Vargas-Chacoff L (2016) Macrobenthos of the coastal Budi Lagoon, southern Chile: changes associated with seasonal environmental variation. Braz J Oceanogr 64(3):239–248. https://doi.org/10.1590/S1679-87592016113306403
Bidet JC, Carruesco C (1982) Étude sédimentologique de la lagune Oualidia (Maroc). Oceanol Acta. pp 29–37
Blanchet H, De Montaudouin X, Chardy P, Bachelet G (2005) Structuring factors and recent changes in subtidal macrozoobenthic communities of a coastal lagoon, Arcachon Bay (France). Estuar Coast Shelf Sci 64(4):561–576. https://doi.org/10.1016/j.ecss.2005.03.016
Blanchet H, Lavesque N, Ruellet T, Dauvin JC, Sauriau PG, Desroy N, Desclaux C, Leconte M, Bachelet G, Janson A-L, Bessineton C, Duhamel S, Jourde J, Mayot S, Simon S, De MX (2008) Use of biotic indices in semi-enclosed coastal ecosystems and transitional waters habitats - Implications for the implementation of the European Water Framework Directive. Ecol Indic 8:360–372. https://doi.org/10.1016/j.ecolind.2007.04.003
Blondel J (1979) Biogéographie et écologie. Masson ed., Paris. p 221
Boehs G, Absher T, Da Cruz-Kaled A (2004) Composition and distribution of benthic molluscs on intertidal flats of Paranaguá Bay (Paraná, Brazil). Sci Mar 68(4):537–543. https://doi.org/10.3989/scimar.2004.68n4537
Borja A, Franco J, Perez V (2000) A marine biotic index to stablish the ecological quality of soft bottom benthos within European estuarine and coastal environments. Mar Pollut Bull 40:1100–1114. https://doi.org/10.1016/S0025-326X(00)00061-8
Carruesco C (1989) Genèse et évolution de trois lagunes du littoral atlantique depuis l’holocène : Oualidia, Moulay Bou Salham (Maroc) et Arcachon (France). Université de Bordeaux I, Bordeaux (France), Thèse de Doctorat, p 485
Chaouti A (2003) Biotypologie du macrozoobenthos de la lagune méditerranéenne de Smir: analyses spatiale et spatio-temporelle. Univ Mohammed V, Rabat, Thèse Doctorat, p 302
Chaouti A, Bayed A (2005) Diversité taxonomique et structure de la macrofaune benthique des substrats meubles de la lagune de Smir. Trav L’institut Sci Rabat Sér Gén 4:33–42
Chaouti A, Bayed A (2008) Spatial patterns of soft-bottom macro-invertebrates and relationships with environmental conditions in a north African coastal lagoon (Smir lagoon, Morocco). Vie Milieu 58:25–35
Chbicheb A (1996) Organisation biogéologique d’un bassin paralique : la lagune d’Oualidia (côte atlantique marocaine). Université de Nantes, Nantes (France), Thèse de Doctorat, p 167
Cherkaoui E (2006) Structure et organisation des peuplements macrozoobenthiques de l’estuaire du Bou Regreg après la construction du barrage. Université Mohammed V, Faculté des Sciences, Rabat (Maroc), Thèse de Doctorat, p 258p
Damsiri Z, Natij L, Khalil K, Loudiki M, Rabouille C, Ettahiri O (2014) Spatio-temporal nutrients variability in the Oualidia lagoon (Atlantic Moroccan coast). Int J Adv Res 2(8):609–618
Dauer DM (1993) Biological criteria, environmental health and estuarine macrobenthic community structure. Mar Pollut Bull 26:249–257. https://doi.org/10.1016/0025-326X(93)90063-P
Dauvin J-C (1988) Structure et organisation trophique du peuplement des sables grossiers à Amphioxus lanceolatus – Venus fasciata de la baie de Morlaix (Manche occidentale). Cah Biol Mar 29:163–185
Do VT, de Montaudouin X, Lavesque N, Hugues B, Guyard H (2011) Seagrass colonization: knock-on effects on zoobenthic community, populations and individual health. Estuar Coast Shelf Sci 95:458–469. https://doi.org/10.1016/j.ecss.2011.10.022
Dufrêne M, Legèndre P (1997) Species assemblages and indicator species : the need for a flexible asymmetrical approach. Ecol Monogr 67:345–366
El Asri F, Zidane H, Maanan M, Tamsouri M, Errhif A (2015) Taxonomic diversity and structure of the molluscan fauna in Oualidia lagoon (Moroccan Atlantic coast). Environ Monit Assess 187(8):545. https://doi.org/10.1007/s10661-015-4752-7
El Asri F, Zidane H, Errhif A, Tamsouri MN, Maanan M, Idrissi MM, Martin D (2018) Polychaete diversity and assemblage structure in the Oualidia Lagoon, Moroccan Atlantic coast. J Mar Biol Assoc UK 98(6):1337–1346. https://doi.org/10.1017/S0025315417000388
El Asri F, Errhif A, Martin D, Tamsouri MN, Maanan M, Zidane H (2021) Community structure and spatial patterns of soft-bottom macrozoobenthos in Oualidia lagoon, Moroccan Atlantic. Thalassas Int J Mar Sci 37:119–129. https://doi.org/10.1007/s41208-020-00263-3
Fang X, Li X, Xiang Y, Hao C, Zhao Y, Zhang Y (2020) Cumulative impact of anthropogenic nutrient inputs on lagoon ecosystems—A case study of Xincun Lagoon, Hainan. China Reg Stud Mar Sci 35:101213. https://doi.org/10.1016/j.rsma.2020.101213
Fauchald K, Jumars P-A (1979) The diet of worms: a study of polychaete feeding guilds. Oceanogr Mar Biol Ann Rev 17:193–284. https://doi.org/10.1146/annurev-marine-010814-020007
Gambi MC, Conti G, Bremec CS, Zoologica S, Dohrn A (1998) Polychaete distribution, diversity and seasonality related to seagrass cover in shallow soft bottoms of the Tyrrhenian Sea (Italy). Sci Mar 62:1–17. https://doi.org/10.3989/scimar.1998.62n1-21
Garnerot F, Bouchereau J-L, Rebelo JE, Guelorget O (2004) L’ichtyofaune dans l’organisation biologique d’un système paralique de type lagunaire, La Ria d’Aveiro (Portugal), en 1987–1988 et 1999–2000. Cybium 28:63–75
Giménez L, Borthagaray AI, Rodríguez M, Brazeiro A, Dimitriadis C (2005) Scale–dependent patterns of macrofaunal distribution in soft–bottom sediment intertidal habitats along a large–scale estuarine gradient. Helgol Mar Res 59:224–236. https://doi.org/10.1007/s10152-005-0223-9
Grall J, Glémarec M (1997) Using biotic indices to estimate macrobenthic community perturbations in the Bay of Brest. Est Coast Shelf Sci 44:43–53. https://doi.org/10.1016/S0272-7714(97)80006-6
Gray JS (1981) The ecology of marine sediments. Cambridge University Press
Grimonet J (1979) Introduction à l’étude démographique de la macrofaune benthique de l’étang du Grau de Piémanson (Camargue). Mém. Ingén., Laboratoire d'Hydrobiologie Université des sciences et technique du Languedoc, Montpellier : p 101
Guélorget O, Penhuisot J-P (1983) Le domaine paralique: Expressions géologiques biologiques et économiques du confinement. Trav Lab Geol ENS 16:136
Hammer Ø, Harper D, Ryan P (2001) PAST: Paleontological statistics software package for education and data analysis. Palaeontol Electron 4:1–9
El Hamoumi R, Dakki M, Rguibi Idrissi H, Radi M (2003) Fiche descriptive sur les zones humides Ramsar (FDR) : Complexe de Sidi Moussa-Walidia (Maroc)
Hassou N, Maanan M, Hennani M, Zourarah B, Assobhei O (2014) Original research article spatial and temporal variation of faecal pollution indicators (Escherichia coli and faecal streptococci ) and physico-chemical parameters at the Oualidia lagoon and its watershed (Morocco). Int J Curr Microbiol Appl Sci 3:675–694
Heilskov AC, Holmer M (2001) Effects of benthic fauna on organic matter mineralization in fish-farm sediments: Importance of size and abundance. ICES J Mar Sci 58:427–434. https://doi.org/10.1006/jmsc.2000.1026
Hernandez-Guevara NA, Pech D, Ardisson P-L (2008) Temporal trends in benthic macrofauna composition in response to seasonal variation in a tropical coastal lagoon, Celestun. Gulf of Mexico Mar Freshwater Res 59(9):772–779
Hilmi K, Koutitonsky V, Orbi A, Lakhdar J, Chagdali M (2005) Oualidia lagoon, Morocco: an estuary without a river. African J Aquat Sci 30:1–10. https://doi.org/10.2989/16085910509503828
Hilmi K, Orbi A, Makaoui A, Bouksim H, Idrissi M, Ettahiri O, Benyounes A, Ait Chattou M (2017) Circulation marine de la baie de Dakhla (Sud du Maroc) par modèle hydrodynamique 2D. Eur Sci J 13:69–82
Hily C, Bouteille M (1999) Modifications of the specific diversity and feeding guilds in an intertidal sediment colonized by an eelgrass meadow (Zostera marina) (Brittany, France). C r Acad Sci Paris 322:1121–1131
Holm-Hansen O, Lorenzen CJ, Holmes RW, Strickland JDH (1965) Fluorometric determination of chlorophyll. ICES J Mar Sci 30:3–15. https://doi.org/10.1093/icesjms/30.1.3
Jayed M, Benbrahim S, Bakkas S, Ramdani M, Flower R (2015) Accumulation of organochlorines in the European clam (Ruditapes decussatus) and sediment of the Oualidia lagoon (Morocco). Bull Environ Contam Toxicol 94:614–621. https://doi.org/10.1007/s00128-015-1517-5
Kevrekidis T (2004) Seasonal variation of the macrozoobenthic community structure at low salinities in a Mediterranean lagoon (Monolimni Lagoon, Northern Aegean). Int Rev Hydrobiol 89(4):07–425. https://doi.org/10.1002/iroh.200310703
Khedhri I, Djabou H, Afli A (2015) Trophic and functional organization of the benthic macrofauna in the lagoon of Boughrara – Tunisia (SW Mediterranean Sea). J Mar Biolog Assoc 4:647–659. https://doi.org/10.1017/S0025315414001544
Khedhri I, Afli A, Aleya L (2017) Structuring factors of the spatio-temporal variability of macrozoobenthos assemblages in a southern Mediterranean lagoon: How useful for bioindication is a multi-biotic indices approach? Mar Pollut Bull 114:515–527. https://doi.org/10.1016/j.marpolbul.2016.10.023
Koaudio KN, Diomandé D, Ouattara A, Koné YJM, Gourène G (2008) Taxonomic diversity and structure of benthic macroinvertebrates in Aby Lagoon (Ivory Coast, West Africa). Pak J Biol Sci 11:2224–2230. https://doi.org/10.3923/pjbs.2008.2224.2230
Lakhdar Idrissi J, Orbi A, Zidane F, Hilmi K, Sarf F, Massik Z, Makaoui A (2004) Organisation et fonctionnement d’un écosystème côtier du Maroc: la lagune de Khnifiss. Rev Sci Eau 174:447–462. https://doi.org/10.7202/705542ar
Lakhlalki H, Jayed M, Benbrahim S, Rharbi N, Bouthir F, Ali Benhra A, Brahim Moutaki B (2017) Accumulation of organochlorine pesticides and polychlorinated biphenyls in the sediments of Oualidia lagoon (Morocco). Bull Soc Zool Fr 142(3):101–118
Lefrere L, Ouassas M, Guillois B, Gillet P, Moukrim A (2015) Macrobenthic community structure of soft-bottom sediments in the Khnifiss lagoon, South of Morocco. J Mater Environ Sci 6:3226–3235
Llanso RJ, Scott LC, Dauer DM, Hyland JL, Russell DE (2002) An estuarine benthic index of biotic integrity for the mid- Atlantic region of the United States. I. Classification of assemblages and habitat definition. Estuaries 25:1219–1230. https://doi.org/10.1007/BF02692219
Maanan M, Ruiz-Fernández AC, Maanan M, Fattal P, Zourarah B, Sahabi M (2014) A long-term record of land use change impacts on sediments in Oualidia lagoon, Morocco. Int J Sediment Res 29:1–10. https://doi.org/10.1016/S1001-6279(14)60017-2
Maanan M, El Barjy M, Hassou N, Zidane H, Zourarah B, Maanan M (2017) Origin and potential ecological risk assessment of trace elements in the watershed topsoil and coastal sediment of the Oualidia lagoon. Morocco Hum Ecol Risk Assess 24(3):602–614. https://doi.org/10.1080/10807039.2017.1394176
Makaoui A, Idrissi M, Agouzouk A, Larissi J, Baibai T, Ouehabi ZE, Laamel MA, Bessa I, Ettahiri O, Hilmi K (2018) Oceanographic state of the Oualidia Lagoon, Morocco (2011–2012). Eur Sci J 14:93
Mannino A, Montagna PA (1997) Small-scale spatial variation in macrobenthic community structure. Estuaries 20:159–173
Marchini A, Gauzer K, Occhipinti-Ambrogi A (2004) Spatial and temporal variability of hard-bottom macrofauna in a disturbed coastal lagoon (Sacca di Goro, Po River Delta, Northwestern Adriatic Sea). Mar Poll Bull 48:1084–1095. https://doi.org/10.1016/j.marpolbul.2003.12.015
Mclusky DS, Mcintyre AD (1988) Characteristics of the fauna. In: Postma H, Zijlstra JJ (eds) Continental shelves ecosystems of the world. Elsevier
Mendes CLT, Tavares M, Soares-Gomes A (2007) Taxonomic sufficiency for soft-bottom sublittoral mollusks assemblages in a tropical estuary, Guanabara Bay, Southeast Brazil. Mar Poll Bull 54:377–384. https://doi.org/10.1016/j.marpolbul.2006.08.026
Mergaoui L, Fekhaoui M, Bouya D, Gheït A, Stambouli A (2003) Qualité des eaux et macrofaune benthique d’un milieu estuarien du Maroc : cas de l’estuaire de Sebou. Bull De L’inst Sci Section Sci De La Vie 25(67–7):5
Mistri M, Rossi R, Fano EA (2001) Structure and secondary production of a soft bottom macrobenthic community in a brackish lagoon (Sacca di Goro, north- eastern Italy). Estuar Coast Shelf Sci 52:605–616. https://doi.org/10.1006/ecss.2001.0757
Mistri M, Ghion F, Modugno S, Rossi R (2002) Response of macrobenthic communities to an hydraulic intervention in an enclosed lagoon (Valle di Gorino, northern Italy). J Marine Biol Ass United Kingdom 82:771–778. https://doi.org/10.1017/S0025315402006136
Newton A, Brito AC, Icely JD, Derolez V, Clara I, Angus S, Schernewski G, Inácio M, Lillebø AI, Sousa AI, Béjaoui B, Solidoro C, Tosic M, Cañedo-Argüelles M, Yamamuro M, Reizopoulou S, Tseng H-C, Donata C, Roselli L, Maanan M, Cristina S, Ruiz-Fernández AC, Pastres R, Snoussi M, Turpie J, Tuchkovenko Y, Dyack B, Brookes J, Povilanskas R, Khokhlov V (2018) Assessing quantifying and valuing the ecosystem services of coastal lagoons. J Nat Conserv 44:50–67. https://doi.org/10.1016/j.jnc.2018.02.009
Nicolaidou A, Petrou K, Kormas KA, Reizopoulou S (2006) Inter-annual variability of soft bottom macrofaunal communities in two Ionian Sea lagoons. Hydrobiologia 555:89–98. https://doi.org/10.1007/s10750-005-1108-2
Nybakken JW (1993) Marine biology: an ecological approach. Harper Collins College, New York, p 445
Pasquaud S (2006) Les relations trophiques : éléments de structuration des peuplements ichtyologiques en milieu estuarien Application à l’estuaire de la Gironde. Dissertation, Université de Bordeaux I
Pearson TH, Rosenberg R (1978) Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanogr Mar Biol Ann Rev 16:229–311
Pielou EC (1966) Shannon’s formula as a measure of specific diversity: Its use and measure. Am Nat 100:463–465
Pranovi F, Curiel D, Rismondo A, Marzocchi M, Scattolin M (2000) Determination of food sources for benthic invertebrates in a salt marsh (Aiguillon Bay, France) by carbon and nitrogen stable isotopes: importance of locally produced sources. Sci Mar 64(3):303–310. https://doi.org/10.3989/scimar.2000.64n3303
Reiss H, Kröncke I (2005) Seasonal variability of benthic indices: an approach to test the applicability of different indices for ecosystem quality assessment. Mar Poll Bull 50:1490–1499. https://doi.org/10.1016/j.marpolbul.2005.06.017
Rharbi N, Ramdani M, Berraho A, Idrissi JL (2001) Caractéristiques hydrologiques et écologiques de la lagune d’Oualidia : milieu paralique de la côte atlantique marocaine. Mar Life 11:3–9
Rodrigues AM, Quintino V, Sampaio L, Freitas R, Neves R (2011) Benthic biodiversity patterns in Ria de Aveiro, Western Portugl: environmental-biological relationships. Est Coast Shelf Sci 95:338–348. https://doi.org/10.1016/j.ecss.2011.05.019
Rossi F, Castelli A, Lardicci C (2006) Distribution of macrobenthic assemblages along a marine gradient in Mediterranean eutrophic coastal lagoons. Mar Ecol 27:66–75. https://doi.org/10.1111/j.1439-0485.2006.00078.x
Salas F, Neto JM, Borja A, Marques JC (2004) Evaluation of the applicability of a marine biotic index to characterize the status of estuarine ecosystems: the case of Mondego estuary (Portugal). Ecol Indic 4:215–225. https://doi.org/10.1016/j.ecolind.2004.04.003
Schückel U, Beck M, Kröncke I (2015) Macrofauna communities of tidal channels in Jade Bay (German Wadden Sea): spatial patterns, relationships with environmental characteristics, and comparative aspects. Mar Biodiv 45:841–855. https://doi.org/10.1007/s12526-014-0308-2
Shannon CE (1948) A mathematical theory of communication. Bell Syst Tech J 27:379–423. https://doi.org/10.1145/584091.584093
Solidoro C, Pastres R, Cossarini G, Ciavatta S (2004) Seasonal and spatial variability of water quality parameters in the lagoon of Venice. J Mar Syst 51:7–18. https://doi.org/10.1016/j.jmarsys.2004.05.024
Stora G, Arnoux A, Galas M (1995) Time and spatial dynamics of Mediterranean lagoon macrobenthos during an exceptionally prolonged interruption of freshwater inputs. Hydrobiologia 300:123–132. https://doi.org/10.1007/BF00024454
Teske P, Wooldridge T (2001) A comparison of the macrobenthic faunas of permanently open and temporarily open/closed South African estuaries. Hydrobiologia 464:227–243. https://doi.org/10.1023/A:1013995302300
Teske PR, Wooldridge TH (2003) what limits the distribution of subtidal macrobenthos in permanently open and temporarily open/closed South African estuaries? Salinity vs sediment particle size. Estuar Coast Shelf Sci 57:225–238. https://doi.org/10.1016/S0272-7714(02)00347-5
Tlig-zouari S, Maamouri-mokhtar F (2008) Macrozoobenthic species composition distribution in the Northern lagoon of Tunis. Transit Waters Bull 2:1–15. https://doi.org/10.1285/i18252273v2n2p1
Uwadiae RE (2013) Spatial patterns in diversity and distribution of benthic molluscs in a weak tidal tropical lagoon. Int J Environ Sci 3:111–120. https://doi.org/10.5376/ijms.2013.03.0014
Vezzone M, Dos Anjos RM, Cesar RG, Muniz M, Cardoso R, Felizardo JP, Vasconcelos D, Polivanov H (2020) Using stable isotopes to discriminate anthropogenic impacts of the sedimentary organic matter pollution in the Rodrigo de Freitas Lagoon (RJ, Brazil). Environ Sci Poll Res 28:4515–4530. https://doi.org/10.1007/s11356-020-10835-8
Ysebaert T, Herman PMJ (2002) Spatial and temporal variation in benthic macrofauna and relationships with environmental variables in an estuarine, intertidal soft-sediment environment. Mar Ecol Prog Ser 244:105–124. https://doi.org/10.3354/meps244105
Zabi SG, Leloeuf P (1993) Review of knowledge on the benthic fauna of marginal and littorals environment of West Africa 22nd part Settlements biotopes. Rev Hydrobiol Trop 26:19–51
Zourarah B, Maanan M, Carruesco C, Aajjane A, Mehdi K, Conceição Freitas M (2007) Fifty-year sedimentary record of heavy metal pollution in the lagoon of Oualidia (Moroccan Atlantic coast). Est Coast Shelf Sci 72:359–369. https://doi.org/10.1016/j.ecss.2006.11.007
Acknowledgements
We are deeply grateful to Mr Brahim Moutaki (INRH Oualidia) and Mr Kaddioui (PhD student) for their help during sampling and to the LETG (UMR 6554, University of Nantes) for kindly allowing us to carry out the granulometric analyses.
Funding
FE, AE, MT, MM, HZ were partly funded by a research program on coastal resources of the LERL/DRH-Casablanca (National Fisheries Research Institute). DM received funding from the Consolidated Research Group on Marine Benthic Ecology, Generalitat de Catalunya (http://agaur.gencat.cat), grant 2017SGR378, and the Agencia Estatal de Investigación (AEI) (http://www.ciencia.gob.es/portal/site/MICINN/aei), European Regional Development Funds (FEDER) (https://ec.europa.eu/regional_policy/en/funding/erdf/), grant CTM2017–88080 / PopCOmics. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All applicable international, national, and/or institutional guidelines for animal testing, animal care and use of animals were followed by the authors.
Sampling and field studies
All necessary permits for sampling and observational field studies have been obtained by the authors from the competent authorities and are mentioned in the acknowledgements, if applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
El Asri, F., Errhif, A., Tamsouri, MN. et al. Analysis of the structural characteristics and spatial organization of macrobenthic fauna in Oualidia lagoon, Morocco. Appl Water Sci 12, 96 (2022). https://doi.org/10.1007/s13201-022-01629-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13201-022-01629-z