
Abstract
Earlier work has demonstrated that attention is indirectly cognitively malleable by processes of self-association – processes by which agents explicitly associate an item with the self. We extend this work by considering the manipulation of attention to both salient and non-salient objects. We demonstrate that self-association impacts attentional processing not only of non-salient objects (i.e., shapes), but also regarding salient items known to command attention (i.e., images of food). This result indicates the flexibility and susceptibility of attentional processing to cognitive manipulation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
1 Introduction
A wide range of work in social and cognitive psychology demonstrates that processes of self-association can enhance the attentional processing of items, leading to effects on preference (Locksley et al. 1980), effects within perception (Sui et al. 2012), and within memory (Symons and Johnson 1997). In this paper we explore the scope of self-association’s impact by investigating whether automatic attentional biases towards salient items can be modulated via association of those items with the self.
In doing so, we seek to extend recent work on the cognitive manipulation of attentional processing stems from work by Sui et al. (2012). Sui et al. (2012) asked participants to form associations between labels and shapes. The labels either invoked a concept of self (i.e., ‘you’) or not (i.e., ‘friend’ or ‘stranger’). After associating the labels with shapes, participants were shown label-shape pairings and were asked to decide whether these were correct or had been changed. Participants were quicker and more accurate in the case of the self-labelled pairs. This is consistent with an attentional bias in favour of items that have simply been associated with the self. A follow-up series of studies demonstrated that the self-association benefit is not a function of the frequency with which self-label pairs appeared (Sui et al. 2015). Not only does the self-association benefit occur when self-label pairs are infrequent (and thus unexpected), if self-label pairs are presented along with other highly frequent pairings, the self-association benefit will override the benefit associated with the frequency of other pairings.
Work on the neural underpinnings of the impact of self-association on attention suggest that the self-association impacts attention in a way analogous to the impact of what is sometimes called ‘perceptual salience,’ or simply salience. Sui et al. (2013) had participants form associations between self, friend, and stranger and various shapes. After forming the associations, participants had to discriminate between the shapes. Although friend- and stranger-associated shapes did not interfere with each other, self-associated shapes interfered with friend and stranger. Moreover, Sui et al. found activation in left intraparietal sulcus (IPS) when participants had to ignore a self-associated shape to select a different shape. This is consistent with a hypothesized function of the left IPS, namely, the function of suppressing salient environmental cues (Mevorach et al. 2010).
The above studies involved the manipulation of attention processing regarding items that are, in themselves, of little salience to agents. The study we report below seeks to extend the scope of this work by applying it to the case of salient and potentially tempting cues – in particular, to images of food.
One might think the manipulation of attention towards such items would prove more difficult. For agents who have developed strong attractions to certain behaviors or outcomes seem to also develop strong attentional biases (Field and Cox 2008; Garavan and Hester 2007; Hou et al. 2011). One explanation of this is that cues related to highly rewarding outcomes are coded as highly salient and rewarding, thereby facilitating increased processing of these cues along with downstream motivational effects (Robinson and Berridge 2001). So, for example, Yokum et al. (2011) have demonstrated that people with higher body-mass index (BMI) respond more quickly to food cues than non-food cues, that BMI correlates with higher activations in brain regions associated with attentional processing and the encoding of reward, and that activations in orbitofrontal cortex during reallocation of attention to unappealing food cues were predictive of increased BMI. And Meule and Platte (2016) have recently demonstrated that while neither automatic attentional biases to high-calorie food nor higher degrees of reported behavioural impulsivity predicted weight gain alone, the combination of the two was predictive of weight gain.
We ask whether it is possible to alter automatic attentional biases towards food items by self-related learning. To this aim, we first selected three types of food items, natural food (food items that have not been cooked), transformed food (food items that have been cooked), and rotten food. The first category includes the types of food that according to most current recommendations should be eaten more frequently as part of a healthy diet. The second category includes the types of food that according to most current recommendations should be eaten less frequently as part of a healthy diet. The third category included food types that were unlikely to be found palatable and which are unambiguously unhealthy to consume. Then we asked participants in the healthy-weight range to learn associations between the three types of food items and person-related labels, i.e. ‘you’, ‘friend’, ‘stranger’, prior to carrying out a food type-label matching task. The stranger pairing condition served as the baseline condition, so that the comparison between the self-related condition and the stranger-related condition will reveal the self-association effects. The friend pairing condition served to control for the effects of familiarity in associative matching. This comparison will allow us to clarify whether the effects associated to self-pairing can be merely explained by the familiar nature of self-information. We measured the effect of self-prioritisation on accuracy and reaction times in food type-label matching.
In line with previous studies on self-association learning (Sui et al. 2012, 2013; Sui and Humphreys 2015a), we hypothesized that pairing food items with the self would lead to a substantial advantage of matching self-related pairs over the pairings of other stimuli. This advantage would be underpinned by greater accuracy and shorter reaction times in the self-related pairing relative to the other related pairing. Furthermore, we predict the magnitude of the self-pairing effect to be greater in the food items of positive value, i.e. natural and transformed food, than in rotten food, since naturally it is less likely to relate oneself with items of negative than positive value (Mather and Carstensen 2005). Moreover, if the benefit from an attentional bias is related to pairing the food items with a familiar stimulus, we should observe a similar advantage when pairing the food items with oneself and with the friend. Furthermore, we might expect differences between the three food categories in the stranger condition (baseline condition), since negative, high-salience items, capture attention to a greater extent than other stimuli (Eastwood et al. 2003).
2 Study
2.1 Participants
Participants were recruited through the online participant recruitment scheme of the [University redacted]. Eligibility requirements meant that participants were aged 18–40, had no history of neurological illness, and no medical history of eating disorders. A total of thirty-eight participants took part in the experiment. Eight participants were excluded from the sample because they scored below-chance in the pairing task (i.e. errors on >50% of trials in at least one of the experimental conditions). One participant had a BMI greater than 30 (obese individual), and was also excluded from the sample given known attentional biases associated with high BMI (Castellanos et al. 2009; Nijs et al. 2010; Yokum et al. 2011). The final sample was 29 participants (13 men) aged 19–35 years (mean = 23.21, SD = 4.126). The sample characteristics are described in Table 1. Participants gave their informed consent prior to the study, with approval by the Ethics Committee, Department of [Redacted].
2.2 Stimulus Material
A set of 60 food pictures was selected from The Foodcast Research Image Database (FRIDa) (Foroni et al. 2013) that included standardised ratings on a number of dimensions such as valence, perceived calories, caloric value per 100 g, familiarity, and discriminability. The food stimuli (530 × 530 pixels) comprised 20 pictures of natural food, 20 pictures of transformed food, and 20 pictures of rotten-food (Fig. 1). These food categories significantly differed in valence (defined in the FRIDa database as the degree to which subjects found the food items pleasant), ranging from ‘very positive’ to ‘very negative’ (natural food, M = 67.09, SD = 11.29; transformed food, M = 64.89, SD = 8.22; rotten food, M = 6.29, SD = 4.90; F(2,38) = 353.486, p < .001), perceived calories (natural food, M = 22.71, SD = 10.91; transformed food, M = 67.32, SD = 12.27; rotten food, M = 25.81, SD = 14.85; F(2,38) = 99.64, p < .001), caloric value per 100 g (natural food, M = 97.35, SD = 105.92; transformed food, M = 298.35, SD = 170.82; rotten food, M = 102.55, SD = 113.18; F(2,38) = 99.64, p < .001), familiarity (natural food, M = 54.71, SD = 19.52; transformed food, M = 51.48, SD = 14.13; rotten food, M = 11.27, SD = 4.89; F(2,38) = 58.210, p < .001), and discriminability (natural food, M = 10.60, SD = 9.49; transformed food, M = 14.73, SD = 10.20; rotten food, M = 42.60, SD = 27.10; F(2,38) = 18.619, p < .001).

Examples of food items
Natural and transformed food had comparable valences, and they both significantly differed from rotten food (all t > 22.19, df = 19, all p < .01). Transformed food items had significantly greater perceived calories and caloric value per 100 g than natural and rotten food items (all t > 12.33, df = 19, all ps < .01). Natural and transformed food were comparable in familiarity and discriminability, but both differed from rotten food (all t > 4.30, df = 19, all ps < .01), which by nature is less available in food contexts (i.e. less familiar) and often harder to discriminate than non-rotten food. All food stimuli were comparable for the following variables: size, spatial frequency, brightness, and arousal (all t < 2.44, df = 19, all ps > .05).
Moreover, the stimuli comprised three words “You”, “Friend”, and “Stranger” (font size 18-point, font type Courier New) in black font. Both the labels and the food items were presented centrally on a white background at a distance of 70 cm from the participants’ eyes (monitor resolution 1024 × 786 pixels). E-prime software (Psychology Software Tools) was used to present the stimuli and record behavioural responses.
2.3 Experimental Procedure
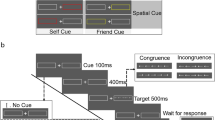
The experimental session was divided in three runs. Each of the three runs started with the pretest phase, whereby participants were instructed to associate the food items (20 natural, 20 transformed, 20 rotten food items) with one of three things: their self, their best friend, or an unfamiliar person (pretest phase). For example, a participant was told “You are natural food, Tom (the named best friend of the participant) is transformed food and a stranger is represented by rotten food”. Note the meaning of the terms natural, transformed, and rotten food (described above) had been explained to the participants. The food items themselves were not presented at this stage. The pairing food type-label was counterbalanced and pseudo-randomized within participants across runs, and the label ‘you’ was associated in each of the three runs with either natural, transformed, or rotten food.
After this, the matching phase of the experiment was performed. As shown in Fig. 2, each trial started with the presentation of a fixation cross (500 ms), followed by the label (80 ms), and subsequently the food item (duration until response). The food type-label pairing could conform to the instructions given in the pretest phase, or it could be a recombination of a label with a different food item. The pairing of the food item and the label was equally distributed, such that each food item was equally likely to appear with each label, and the ratio between match and mismatch trials was 1:1.

Timeline of the experimental procedure
Participants’ task was to judge whether the food item was correctly assigned to the relevant person by pressing one of the two response buttons as quickly and accurately as possible. Response buttons were counterbalanced across participants. Feedback (correct or incorrect) was presented on the screen for 500 ms after participants responded in every trial. Participants were also informed of their overall accuracy at the end of each run.
The overall experiment consisted of 1080 randomized trials presented in 3 runs (360 trials per run) following some practice trials, where self, friend, stranger, and re-paired stimuli occurred equally often in a random order. Thus, there were 60 trials per condition (self-matched natural/transformed/rotten food, self-nonmatched natural/transformed/rotten food, friend-matched natural/transformed/rotten food, friend-nonmatched natural/transformed /rotten food, stranger-matched natural/transformed/rotten food, stranger-nonmatched natural/transformed/rotten food).
2.4 Self-Report Measures
After the matching task, to ensure that the participants included in sample were comparable in the these measures that may influence food perception, we measured Body Mass Index (BMI) based on self-reported height and weight, and feelings of hunger by using a 10 cm visual analogical scale (VAS) ranging from 0 (“I am not hungry at all”) to 100 (“I am starving”). In addition, we measured participant’s eating behaviour with the Three-Factor Eating Questionnaire-R21 (TFEQ), which includes three domains of eating behaviour: cognitive restraint (6 items), uncontrolled eating (9 items), and emotional eating (6 items). Most items were rated on a four-point Likert scale ranging from a minimum of 0 to a maximum of 4. One item was rated on an eight-point numerical scale (minimum = 0, maximum = 8) (Cappelleri et al. 2009).
To assess the familiarity effect, we asked participants to report the length of the friendship (in years), level of familiarity on a scale from 1 (not familiar at all) to 7 (highly familiar), the frequency in which they meet on a scale from 1 (not frequently at all) to 7 (very frequently), and whether they meet daily, weekly, monthly, or otherwise.
The results of the self-report measures are reported in the Supplementary material.
2.5 Data Analysis
Data was analysed using SPSS Statistics 22 (IBM). Analysis were centred on the matching trials, because we were principally interested in testing whether matching a food item with oneself modulates attentional biases towards the food item. Differences in accuracy and reaction times between conditions were analysed using repeated measures ANOVAs with within-subject factors food type (natural, transformed, rotten) and label (you, friend, stranger). To control for the influence of carrying over pairing effects from one block to the next, the repeated measures ANOVAs also included the variable block order (first, second, third, fourth, fifth, sixth) as between-subject factor. Additionally, paired t-tests were performed. Mauchly’s W was computed to check for violations of the sphericity assumption and Greenhouse–Geisser adjustments to the degrees of freedom were applied when needed. The p values were corrected for multiple comparisons using stepwise Bonferroni-Holm correction.
2.6 Results
Below we report results on three different measures: participant accuracy, participant response time, and participant responses to our self-report questions.
2.6.1 Participant Accuracy
Table 2 represents mean accuracy for each condition. A three-way repeated-measures ANOVA of accuracy with factors food type (natural, transformed, rotten), label (self, friend, stranger) and block order (first, second, third, fourth, fifth, sixth) revealed the following. First, we found a significant difference between self, friend and stranger as indicated by a main effect of label (F(2,46) = 8.836, p = .002, η2 = .278). The results also showed that the self versus stranger differences were influenced by the order of presentation, as indexed by an interaction between label and block order (F(10,46) = 2.846, p = .014, η2 = .382). Second, we found significant differences between the three food types (i.e. main effect of food type, F(2,46) = 11.815, p < .001, η2 = .339). Importantly, the ANOVA results showed that the food type-label pairing effect was different for the self, the friend and the stranger condition, as indicated by interactions between food type and label (F(4,92) = 5.923, p < .001, η2 = .205) as well as between label, food type, and block order (F(4,92) = 1.985, p = .015, η2 = .301).
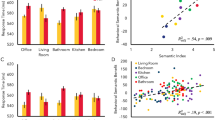
To directly test for the pairing effects on the matching processes of the three food types we performed three separate ANOVAs for natural, transformed and rotten food with the within-subject factor label (self, friend, stranger), and the block order as a between-subject factor. For natural food, the results of the ANOVA showed significant differences between self, friend and stranger (F(2,46) = 16.428, p < .001, η2 = .417). Crucially, this effect did not interact with the factor block order showing that the pairing effects on natural food items are independent from the pairing order across blocks. Follow-up t-tests revealed that there was a significant advantage for the process of pairing natural food items with the self over the pairing with the stranger (t(28) = 5.467, p < .000, d = 1.03) and with the friend (t(28) = 2.886, p = .007, r = 0.229). There was also an advantage for pairing natural food with the friend as compared to the stranger (t(28) = 3.119, p = .004, d =. 5894; Fig. 3, see Table 2 for accuracy scores). Furthermore, the significant advantage of the self over the friend label indicates that the pairing effects observed in the self-condition do not reflect familiarity alone.

Accuracy scores
In contrast, the ANOVA performed for transformed food did not reveal significant differences between labels (F(2,46) = 3.222, p = .049, η2 = .123). For rotten food, the results of the ANOVA showed no main differences between self, friend and stranger (F(2,26) = 2.940; p = .063; η2 = .113), but the order of presentation lead to differences in the association of rotten food with self, friend and stranger (i.e. interaction between label and block order (F(2,46) = 4.642, p < .001, η2 = .502). The follow-up tests showed differences when pairing rotten food with the friend versus the stranger (F(1,23) = 8.844, p = .000, η2 = .658). Although this interaction can be interesting to explain the friend and stranger pairing effects, it expands beyond the aim of the study, i.e. to investigate whether self-association can modulate food attentional bias. Therefore, this interaction will not be further tested. The analysis did not reveal any other main significant effects or in interaction (all Fs < 2.927, df = 1,23, all ps > 0.035).
2.6.2 Participant Response Times
Mirroring the analysis performed for the accuracy measure, we tested differences in reaction times by performing a three-way repeated measures ANOVA with factors food type (natural, transformed, rotten), label (self, friend, stranger), and block order (first, second, third, fourth, fifth, sixth). This analysis revealed significant differences between self, friend and stranger (F(2,46) = 29.013, p < .001, η2 = .558), significant differences between the food types (F(2,46) = 14.579, p < .001, η2 = .388), and significant differences when pairing the three labels – self, friend, stranger, with the three food items – natural, transformed and rotten (F(4,92) = 12.886, p < .001, η2 = .359). In view of the differential effects of associating the three labels with the three food items, we then performed further ANOVAs to directly test for the pairing effects on the three types of food items (natural, transformed, rotten food) separately. The ANOVAs with within-subject factor label (self, friend, stranger) showed a main effect of label for natural food (F(2,56) = 40.445, p < .001, η2 = .591), for transformed food (F(2,56) = 7.021, p = .004, η2 = .200) and for rotten food (F(2,56) = 20.188, p < .001, η2 = .419). Follow-up t-tests revealed faster reaction times when matching natural food with the self versus stranger (t(28) = −7.037, p < .001, d = 1.32), the self versus the friend (t(28) = −6.305, p < .001, d = 1.191), and when matching natural food with the friend versus the stranger (t(28) = −4.706, p < .001, d =. 889). For transformed food items, there was also a significant advantage for self-matching versus to the stranger-matching condition (t(28) = −3.497, p = .002, d =. 661). For rotten food items, results revealed faster reaction times for the self-pairing condition versus the friend-matching condition (t(28) = −5.657, p < .001, d = 1.0691), as well as an advantage for the friend versus the stranger-matching condition (t(28) = 3.843, p = .001, d =. 7263).
Importantly, these results revealed a significant advantage in reaction times when pairing the food items with the self as opposed to the friend or the stranger (Fig. 4). This demonstrates that the self-pairing manipulation was effective across all types of food items. Moreover, the differences between the self and the friend labels suggest that the advantage that we observe when associating the food items with the self are not related to familiarity. Furthermore, the lack of interaction with the factor block order suggests that it is unlikely that the pairing effects in the self-related condition are related to the presentation order of the blocks in the experimental session.

Reaction time scores (ms)
Overall, then, the results suggests that when food items were paired to the self label, relative to when they were matched to the stranger or the friend label, there was a substantial advantage for matching natural food items as indicated by greater accuracy and shorter reaction times for self-related pairing relative to the other related pairing. In addition, pairing food items with the label self led to faster responses to all food items.
Furthermore, to further control that the results were not influenced by the relatively large number of excluded participants (N = 9), we re-ran the analysis of accuracy of responses and reaction times on the full sample (N = 38). These analyses show a very similar pattern of result as that of the analysis performed on the final sample. This is, associating natural food with oneself results in greater accuracy and shorter reaction times, in comparison to associating the natural food with a friend or a stranger (see Supplemental material for details of the analysis).
3 Discussion
Our main aim in this study was to explore the scope of a well-documented self-association effect upon attentional processing. In particular, we aimed to investigate whether we can modulate automatic attentional biases towards salient food images by making self-related associations to the food items. Participants were asked to form associations between three labels referring to themselves, a friend, or a stranger, and three types of food items: natural, transformed, and rotten food items, and then to immediately judge if subsequent label-food item pairings matched. The results of the study showed that when different food types were matched to a stranger then all the food types were identified equally by subjects. Despite the expected tendency to rapidly respond to negative, highly-salient, rotten food items, subjects did not make more accurate match responses to natural, transformed, or rotten food types (green bars on right hand sides of Fig. 3). In other words, the food types had a similar attentional salience when they were associated with the stranger.
However, when food items were paired with the self, the results were dramatically different (blue bars on left hand side of Figs. 3 and 4). There was a substantial advantage for matching natural food items to the self-label, relative to when the natural food items were associated with the other. This advantage was underpinned by both a greater accuracy and shorter reaction times for the self-related pairing relative to the other (whether friend or stranger) related pairing. We thus demonstrated that associating salient food items with the self has an important impact. It seems that associating oneself with items in the world is a robust way to manipulate the way that attentional processing responds to those items.
Converging evidence from associative learning studies suggests that self-advantages on perceptual matching arise independently from the advantages observed in stimulus associated with reward, suggesting that self- and reward-advantage have different origins (Northoff and Hayes 2011; Sui and Humphreys 2015a, b). This evidence fits with the hypothesis that the self-related advantage for natural food could be linked to competing processes between the self-related network and the attentional control network. Specifically, the greater magnitude of the self-related effects in natural food items relative to transformed (i.e. high-calorie) food items observed in the current study might be linked to the interaction of two parallel attentional processes, one primed by the self-label, one primed by the high-calorie, high-reward, food items. In contrast, because natural food items are typically less salient than high-calorie food items, the activation of the attentional system driven by self-representations might have overcome parallel activations driven by the natural food item. Further research is needed to elucidate the relationship between the self- and the reward-related attentional mechanisms in the context of food processing.
The current study has a series of limitations. First, the three food categories employed in the study differed in a series of parameters such as valence and familiarity. For example, natural food items were rated as most pleasant relative to transformed or rotten food items. Besides, rotten food items were rated as less familiar than the other two types of food items. These differences could have potentially influenced the results. Future studies should include additional control food categories matched for such a parameters. Second, due to limitations of the experimental design, the learning associating patterns were presented in blocks through the experiment (i.e. self-related block, friend-related block, stranger-related block). Although supplementary analysis indicate that the block order did not affect the self-pairing advantage of natural food items, future studies should aim for counterbalance design where the association pattern regularly changes. Moreover, the present study did not measure whether it is possible to influence behaviour – i.e., food consumption – by manipulating attention via the self-association procedure. It thus remains unclear how effective this particular manipulation of attention may be when agents are subject to temptation.
In spite of these limitations, however, this study represents evidence low-level attentional processes can be directly manipulated in line with previous evidence suggesting that otherwise neutral stimuli benefit from an attentional bias if they are associated with the self (Sui et al. 2012, 2013; Sui and Humphreys 2015a). Thus, we suggest that self-association learning could be potentially used to enhance automatic perceptual bias towards natural (healthy) food items, which could be especially helpful to those individuals that exhibit low-levels of self-control towards food, that are at risk of developing overweight and obesity. Future studies should investigate whether self-association learning could be used to make effective changes on food choices and weight loss.
4 Conclusion
We have demonstrated that processes of self-association impact the processing of salient items – in this case, images of food. This result further elucidates the structure of attentional processing and demonstrates the flexibility of this cognitive manipulation. Simply by associating the self with a type of food item, attentional processing towards the item is enhanced.
References
Cappelleri, J.C., A.G. Bushmakin, R.A. Geber, N.K. Leidy, C.C. Sexton, M.R. Lowe, and J. Karlsson. 2009. Psychometric analysis of the three-factor eating questionnaire-R21: Results from a large diverse sample of obese and non-obese participants. International Journal of Obesity 33 (6): 611–620.
Castellanos, E.H., E. Charboneau, M.S. Dietrich, S. Park, B.P. Bradley, K. Mogg, and R.L. Cowan. 2009. Obese adults have visual attention bias for food cue images: Evidence for altered reward system function. International Journal of Obesity 33: 1063–1073.
Eastwood, J.D., D. Smilek, and P.M. Merikle. 2003. Negative facial expression captures attention and disrupts performance. Perception & Psychophysics 65 (3): 352–358.
Field, M., and W.M. Cox. 2008. Attentional bias in addictive behaviors: A review of its development, causes, and consequences. Drug Alcohol Depend. 97 (1–2): 1–20.
Foroni, F., G. Pergola, G. Argiris, and R.I. Rumiati. 2013. The FoodCast research image database (FRIDa). Frontiers in Human Neuroscience 51 (7): 1–19.
Garavan, H., and R. Hester. 2007. The role of cognitive control in cocaine dependence. Neuropsychology Review 17 (3): 337–345.
Hou, R., K. Mogg, B.P. Bradley, R. Moss-Morris, R. Peveler, and A. Roefs. 2011. External eating, impulsivity and attentional bias to food cues. Appetite 56 (2): 424–427.
Locksley, A., V. Ortiz, and C. Hepburn. 1980. Social categorization and discriminatory behavior: Extinguishing the minimal intergroup discrimination effect. Journal of Personality and Social Psychology 39 (5): 773–783.
Mather, M., and L.L. Carstensen. 2005. Aging and motivated cognition: The positivity effect in attention and memory. Trends in Cognitive Sciences 9 (10): 496–502.
Meule, A., and P. Platte. 2016. Attentional bias toward high-calorie food-cues and trait motor impulsivity interactively predict weight gain. Health Psychology Open 3 (1): 205510291664958.
Mevorach, C., J. Hodsoll, H. Allen, L. Shalev, and G. Humphreys. 2010. Ignoring the elephant in the room: A neural circuit to downregulate salience. Journal of Neuroscience 30 (17): 6072–6079.
Nijs, I.M., P. Muris, A.S. Euser, and I.H. Franken. 2010. Differences in attention to food and food intake between overweight/obese and normal-weight females under conditions of hunger and satiety. Appetite 54: 243–254.
Northoff, G., and D.J. Hayes. 2011. Is our self nothing but reward? Biological Psychiatry 69 (11): 1019–1025.
Robinson, T.E., and K.C. Berridge. 2001. Incentive-sensitization and addiction. Addiction 96 (1): 103–114.
Sui, J., and G.W. Humphreys. 2015a. The interaction between self-bias and reward: Evidence for common and distinct processes. The Quarterly Journal of Experimental Psychology 68 (10): 1952–1964.
Sui, J., and G.W. Humphreys. 2015b. More of me! Distinguishing self and reward bias using redundancy gains. Attention and Perception Psychophysics 77 (8): 2549–2561.
Sui, J., X. He, and G.W. Humphreys. 2012. Perceptual effects of social salience: Evidence from self-prioritization effects on perceptual matching. Journal of Experimental Psychology: Human Perception and Performance 38 (5): 1105–1117.
Sui, J., P. Rotshtein, and G.W. Humphreys. 2013. Coupling social attention to the self forms a network for personal significance. Proceedings of the National Academy of Sciences of the United States of America 110 (9): 7607–7612.
Sui, J., F. Enock, J. Ralph, and G.W. Humphreys. 2015. Dissociating hyper- and hypo-self biases to a core self-representation. Cortex 70: 202–212.
Symons, C.S., and B.T. Johnson. 1997. The self-reference effect in memory: A meta-analysis. Psychological Bulletin 121 (3): 371–394.
Yokum, S., J. Ng, and E. Stice. 2011. Attentional Bias to food images associated with elevated weight and future weight gain: An fMRI study. Obesity 19 (9): 1775–1783.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOCX 12.5 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Sel, A., Sui, J., Shepherd, J. et al. Self-Association and Attentional Processing Regarding Perceptually Salient Items. Rev.Phil.Psych. 10, 735–746 (2019). https://doi.org/10.1007/s13164-018-0430-3
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13164-018-0430-3