
Abstract
The cultivation of flooding-tolerant grasses on wet or rewetted peatlands is a priority in climate change mitigation, balancing the trade-off between atmospheric decarbonisation and biomass production. However, effects of management intensities on greenhouse gas (GHG) emissions and the global warming potential (GWP) are widely unknown. This study assessed whether intensities of two and five annual harvest occurrences at fertilisation rates of 200 kg nitrogen ha− 1 yr− 1 affects GHG exchange dynamics compared to a ‘nature scenario’ with neither harvest nor fertilisation. Fluxes of carbon dioxide (CO2), methane (CH4), and nitrous oxide (N2O), using opaque and transparent chambers, were measured on a wet fen peatland with a mean water table depth of -10 cm below soil surface. Overall, no treatment effect was found on biomass yields and GHG emissions. Annual cumulative CH4 emissions were low, ranging between 0.3 and 0.5 t CO2-C eq ha− 1 yr− 1. Contrary to this, emissions of N2O were high, ranging between 1.1 and 1.5 t CO2-C eq ha− 1 yr− 1. For magnitudes of CH4 and N2O, soil moisture conditions and electrical peat properties were critical proxies. Atmospheric uptake of CO2 by net ecosystem exchange was higher for the treatments with management. However, this benefit was offset by the export of carbon in biomass compared to the treatment without management. In conclusion, the results highlighted a near-equal GWP in the range of 10.5–11.5 t CO2-C eq t ha− 1 yr− 1 for all treatments irrespectively of management. In a climate context, a restoration scenario but also intensive paludiculture practices were equal land-use options.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Development regarding the cultivation of flooding-tolerant crops on wet or rewetted peatlands, known as ‘paludiculture’ (Tanneberger et al. 2021), was recently highlighted to play a major role as a key priority in mitigating climate change (Evans et al. 2021). This is due to a conflict in global needs in which the first aspect is societal dependence on arable land in times of a growing world population but the simultaneous need for ecological conservation (Mehrabi et al. 2018). On the other hand, decarbonisation is inevitable in order to limit global warming within the next 30 years to a peak warming of 2 °C (Iyer et al. 2022; Meinshausen et al. 2022). A nature-based solution to decarbonisation is peatland protection and rewetting, known to limit or, in some cases, stop or reverse radiative forcing due to the prevention of high carbon dioxide (CO2) emissions by waterlogging (Günther et al. 2020).
Though knowledge on greenhouse gas (GHG) emission factors (EF) for drained, rewetted, and pristine peatlands is fragmented, data exist with an increasing abundance. Contrary to this, there is a lack of disaggregated knowledge regarding emission dynamics for wet or rewetted peatlands under paludiculture practices (Bianchi et al. 2021). The main drivers of variation regarding CO2 emissions from peatlands are well known, being mainly dependent on the dynamics of water table depth (WTD), biomass growth, and biogeochemical stabilisation of soil organic matter (Moore and Dalva 1993; Skinner 2008; Wang et al. 2021; Kalisz et al. 2021). However, the determination of explicit drivers is even more complex for emissions of methane (CH4), where pivotal processes still are under debate. Considering the dynamics of anaerobic soil biogeochemistry, the selection of species for paludiculture was highlighted as essential (Noyce and Megonigal 2021). Typical semi-aquatic wetland plants, such as Typha spp. or Phragmites spp., are known to potentially contribute to increased CH4 emissions by allowing CH4 to bypass possible oxidation processes within the soil by aerenchymatous transport (Minke et al. 2016; Vroom et al. 2022). Contrary to this, bryophyte-dominated (i.e., Sphagnum spp.) peatlands are frequently associated with methanotrophic (i.e., methane-consuming) activity (Larmola et al. 2010; Kolton et al. 2022). However, no such direct association between vegetation composition and CH4 emissions exists for perennial grasses.
Phalaris arundinacea, or reed canary grass (RCG), is a bunchgrass species with a wide array of potential habitats. It is cultivated on dry mineral soils but also naturally invades floodplains and wetlands (Ustak et al. 2019). So far, the cultivation and biomass utilisation of RCG has been mainly for forage or as a bioenergy crop, while it recently gained attention as a feedstock in green protein biorefinery (Utama et al. 2018; Nielsen et al. 2021a; Næss et al. 2023). Furthermore, its versatility in various environmental conditions led to unique anatomical features, particularly in their culm and rhizomes, thereby distinguishing RCG from other Poaceae species and placing it structurally between mineral-soil adapted and semi-aquatic plants (Zhang et al. 2017). However, while the potential for cultivation of RCG on wet peatlands has been demonstrated, an assessment of associated GHG emissions is still lacking for management options beyond the commonly applied annual harvest in winter months or an experimental two-cut management (Kukk et al. 2010; Heinsoo et al. 2011; Järveoja et al. 2016; Karki et al. 2019). Therefore, the determination of a potential effect of different management intensities on annual GHG balances for RCG cultivation on wet or rewetted peatlands is critical in order to promote the development of knowledge to efficiently balance the needs for atmospheric decarbonisation and agricultural production (Mehrabi et al. 2018; Evans et al. 2021; Bianchi et al. 2021).
Thus, manipulation of the grass stand age is likely to be related to different magnitudes of ecosystem-atmosphere exchanges of GHG. In a previous study, variations in harvest intensities were found to affect root growth significantly and, therewith, the potential for instant belowground accumulation of carbon (C), with lesser harvests leading to increased C storage (Nielsen et al. 2021b). However, more frequent harvest occurrences have the potential to efficiently mitigate N2O emissions in summer months, where oxic conditions might occur due to reduced WTD. A dropdown in WTD has the potential to result in increased nitrogen (N) mineralisation, particularly in nutrient-rich fen peatlands (Minkkinen et al. 2020). There, regrowth of aboveground biomass (AGB) following harvest will not be delayed by N scarcity and the optimal partitioning mechanisms of plant growth, prioritising the development of plant organs critical to provide the most limiting resource (Fraser et al. 2015; Yang et al. 2018). Further, regrowth of juvenile plant material is associated with enhanced uptake of atmospheric CO2, due to enhanced photosynthetic activity, and N (Walker et al. 2014; Tejera et al. 2022).
Therefore, it was hypothesised that different intensities of harvest and fertilisation frequencies of RCG cultivated on a wet fen peatland will affect annual GHG emissions and the resulting global warming potential (GWP). In this context, the aim was to assess whether intensities of two and five annual harvest occurrences at fertilisation rates of 200 kg N and K per ha− 1 yr− 1 will lead to differing carbon balances and gas exchange dynamics as compared to a ‘nature scenario’ with neither harvest nor fertilisation.
Methods
Study Site and Biomass Harvest
We determined GHG emissions on four plots, cultivated with Phalaris arundinacea, cultivar: Lipaula, on a riparian fen peatland in Vejrumbro, Denmark (56°26’15.3"N, 9°32’44.1"E). The Vejrumbro field site has previously been used in various studies, e.g., regarding biomass productivity (Nielsen et al. 2021a) and the effects of flooding (Malinowski et al. 2015). The climate is temperate, with a long-term annual average temperature of 8.3 °C and an annual average precipitation of 675 mm for 1991–2022 (Aarhus University Viborg, Meteorological Station, Foulum). The Vejrumbro field site has been established in 2018 with various flood-tolerant perennial grass species as part of a biomass study. In the last decades, the site was classified as grassland and mainly used for grazing due to its wetness despite the establishment of drainage systems in the form of ditches and tile drains around the 1950s. Frequent flooding events on the study site and the relative distances of the GHG measurement plots to the Nørre Å river and drainage ditches created spatial heterogeneity of varying organic carbon (OC) contents and other environmental variables across the four plots (Table 1).
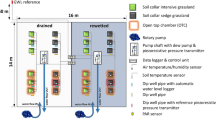
The four plots used for GHG measurements were subdivided into six subplots (Fig. 1) with differing management intensities represented by annual frequencies of harvest and fertilisation. Of these, treatments of zero (0-cut), two (2-cut) and five (5-cut) annual cuts were chosen for this study. The treatments with two and five annual cuts received split-fertiliser applications of in total 200 kg N and K ha− 1 yr− 1. Details on cultivation and seeding, as well as fertilisation and harvest occurrences, were described by Nielsen et al. (2021a). For this study, the subplots were equipped with permanently installed soil collars (55 cm x 55 cm), boardwalks, and relevant instrumentation for determining subplot-specific environmental parameters, as described below. The installation was performed in February and March 2020, and first test measurements started in April 2020. Biomass within the collars was harvested with a handheld grass trimmer (Makita, Anjō, Japan) in calendar weeks 24 and 36 (2-cut treatment), as well as 20, 24, 32, 36, and 42 (5-cut treatment). Harvested biomass was weighed, chopped, and oven-dried at 60 °C before further determination of organic carbon (OC) content on a Vario MAX CN (Elementar Analysesysteme GmbH, Hanau, Germany).

Drone picture of the Vejrumbro field site in the Nørre Å river valley. Plots used for the field trial are highlighted in red, with plot numbers indicated within the plots. A close-up shows a schematic illustration of the split-plot design, including infrastructure for greenhouse gas measurements. Drone pictures were taken by Jens Kjeldsen
Meteorological and Hydrological Parameters
The meteorological station of Aarhus University Viborg, approximately 7 km from the study site, continuously measured photosynthetically active radiation (PAR), air temperature, and precipitation. In addition, each subplot was equipped with a soil temperature (Tsoil) logger at -5 cm depth, logging soil temperature every hour. Further, for each GHG sampling occasion and subplot, measurements of Tsoil at -2 cm, -5 cm, and − 10 cm were taken manually with a digital thermometer (Weber Inc., IL, USA). The volumetric water content (θ, cm3 cm− 3) was measured during GHG sampling campaigns using a time domain reflectometry (TDR) system (Thomsen 2006). Further, subplots were equipped with electrodes for measurements of redox conditions (Eh) at -5 cm and − 25 cm depth, as well as one piezometer for manual measurements of water table depth (WTD), with screens between– 100 cm to -5 cm below ground surface. In addition, dipwells (perforated PVC tubes) of 100 cm in length were installed within the collar area and equipped with a water depth logger (Levelogger5; Solinst Canada Ltd., Ontario, Canada), measuring automatically every hour. Atmospheric pressure correction was based on barometric loggers of the same manufacturer. For all components, negative signs denote depths below the ground surface and vice versa.
Greenhouse Gas Sampling and Analysis from Opaque Chamber Measurements
White opaque PVC chambers (60 cm x 60 cm x 41 cm) were used for gas flux measurements of CO2, CH4, and N2O. Sampling campaigns were held every second week from the 5th of May 2020 to the 4th of May 2021. The chambers were equipped with a fan to mix headspace air, a vent to ensure pressure equilibrium, and a temperature probe measuring air temperature inside the chamber. During all GHG sampling campaigns, chambers were placed on separate middle pieces (60 cm x 60 cm x 41 cm), which were pre-positioned 30 min prior to sampling on the permanently installed soil collars. This procedure was in order to avoid methane ebullition as well as to make space for biomass. Five gas samples (11mL) were withdrawn at 0, 5, 10, 25, and 50 min after chamber closure, using a syringe (20 mL) connected to the chamber sampling port by a polypropylene tube of 1.2 m and 4 mm inner diameter. The sampling port inside the chamber had three air inlets, additionally ensuring the withdrawal of well-mixed air. After discarding 16 mL of dead air volume from the tube system, air from the syringe was transferred to pre-evacuated 6 mL glass exetainers (Labco Limited, UK). Those were stored dark until analysis on an Agilent 7890 gas chromatograph, equipped with an automatic injection system (CTC CombiPAL, Agilent A/S, Nærum, Denmark). Gas fluxes were calculated in R (R core team (2021), version 4.1.2 “Bird Hippie”), using the ‘gasfluxes’ package (Fuss et al., 2020), and a combination of linear and non-linear models for flux estimation with model selection based on the kappa.max technique for reduction of bias and uncertainty as in detail described by Hüppi et al. (2018), considering the GC system-specific precision limit (Petersen et al. 2012) and defining the minimum detectable fluxes to 3.5 g CO2 m− 2 h− 1, 3.2 µg CH4 m− 2 h− 1, and 1.9 µg N2O m− 2 h− 1. From a total of 341 fluxes, 83%, 89%, and 95% of CO2, CH4, and N2O fluxes were calculated using the robust linear model. 12 fluxes of CO2 and one flux of N2O were ‘NA’ while the remaining fluxes were determined using the HMR.fit method.
Aggregated annual fluxes of CH4 were calculated based on daily dependences with soil temperature using the R package flux (Jurasinski et al. 2014; version 0.3-0), using a similar approach as for CO2 (Eq. 1), while N2O emissions were aggregated to annual cumulative values using linear interpolation (Fuß et al., 2020). Fluxes of CO2 were only determined and calculated as a quality check (leak test) for CH4 and N2O and were not included in any further calculations.

In which RCH4 is the flux of CH4, Ϙ1 is the base respiration rate at 10℃, Ϙ2 is the bias coefficient, and Temp is the measured hourly soil temperature at -5 cm.
Measurements of Carbon Dioxide Using Transparent Chambers
In the same period from the 5th of May 2020 to the 4th of May 20,201, CO2 fluxes were measured in biweekly intervals using transparent chambers of the same size, including transparent middle pieces, as described for the opaque chambers in the section above. Ideally, measurements were performed the day following the opaque chamber measurements. However, this rhythm was subject to minor alterations of ± two days to ensure that measurements were performed in sampling windows (between 10:00 and 15:00) without precipitation. CO2 concentrations were measured using a Li-Cor 840 A infrared gas analyser (Li-Cor Inc., Lincoln, USA) connected to an automatic datalogger (Campbell CR850; Campbell Scientific Inc., Logan, USA), over chamber closure times of 120 s. The chambers were equipped with (1) air-temperature sensors, measuring temperature in- and outside the chamber, (2) a temperature control unit, starting automatically when temperature differences exceeded 1 °C, (3) a PAR sensor (Li-Cor 190-SA; Li-Cor Inc., Lincoln, USA), (4) as well as an H2O sensor– later used to correct CO2 for water vapour concentrations according to Webb et al. (1980). More details on the design for transparent chambers were described by Elsgaard et al. (2012).
To cover different ranges of naturally occurring PAR, artificial PAR blocking was applied by shrouding the chambers with meshes. For each collar placement, first, net ecosystem exchange (NEE) measurements were conducted without shrouding at natural PAR, followed by 50% PAR-blocking and 75% PAR-blocking. Finally, one set of Reco measurements was conducted by blocking 100% PAR. Before each PAR-blocked measurement, plants were given 2 min to adapt photosynthesis rates. This procedure resulted in four measurements per collar placement. In addition, the order of transparent chamber measurements was randomised for each sampling campaign.
Annual Carbon Balances
Based on the flux calculation, hourly fluxes for Reco and gross primary production (GPP) were determined by gap-filling for each plot and treatment. Here, a Tier 2 method, as described by Liu et al. (2022), including vegetation height (GH) but using soil temperature (Ft), was applied. This resulted in the following Lloyd and Taylor Arrhenius-type models for Reco (Eq. 2) and a Michaelis-Menten-based equation for GPP (Eq. 3).
Values for GH were interpolated between manual measurement campaigns at each GHG sampling occasion, with a cut set to the stubble height of 7 cm after each harvest occurrence. In those equations, TRef is the reference temperature of 283 K, and T0 is the temperature constant of 227 K, indicating the start of biological processes. Rref denotes respiration at TRef, while E0 are estimated values for the activation energy. GPPmax is the maximum rate of carbon fixation at a PAR of 2000 expressed in mg CO2-C m− 2 h− 1, whereas alpha (α) denotes the light use efficiency. Both equations were previously described in detail by Liu et al. (2022) and Oestmann et al. (2022) and are commonly applied. NEE values for each hour were derived by summing Reco and GPP.
Following the approach of Liu et al. (2022) and Hoffmann et al. (2015), the fitted models were controlled for performance using the comparison between modelled and measured values by hands on a variety of performance indicators: Mean absolute error (MAE), observations standard error (RSR) based on root mean square error, coefficient of determination (R2), modified index of agreement (md), per cent bias (PBIAS), and Nash-Sutcliff’s model efficiency (NSE). Thresholds for performance ratings were adapted from Hoffmann et al. (2015). Models were accepted if 5 out of 6 performance indicators were at least “satisfactory” and two indicators outweighed the “unsatisfactory” performance with a “good” performance rating. Uncertainties for all GHGs and annual balances were determined using a combined bootstrap and Monte Carlo jackknife method with 1000 iterations. Details on this method were previously described by, e.g., Köhler et al. (2012), Beetz et al. (2013), and Günther et al. (2015). To calculate the GWP of CH4 and N2O in terms of CO2eq, we applied conversion factors of 28 and 265 (Myhre et al. 2013), also adopted by the United Nations Framework Convention on Climate Change (UNFCCC) in 2021. For all GHGs and balances, the atmospheric sign convention was applied where negative values indicate uptake and positive values emissions.
In terms of global warming potential (GWP), annual GHG balances (Eq. 4) for each treatment and plot were calculated considering the export of C by biomass harvest (CExport):
For better comparability, all annual balances were expressed as CO2-C eq.
Statistics
For all parameters and values, observations are reported as means with standard errors (n = 4) to present the data distribution. The significance of differences between means was tested by one-way ANOVA with post-hoc Tukey’s HSD at a confidence level of 95%. Effects of co-variates on CH4 and N2O fluxes were assessed using generalised additive models (GAMs) in the package mgcv (Wood, Version 1.8–39,2022) in R (R Core Team (2020) Version 4.1.2– “Bird Hippie”), capable of accounting for linear and non-linear relationships (Marra and Wood 2011; Wood 2011; Wood et al.,2016). Effects of co-variables and categorical treatments on the annual cumulative emissions of NEE, CH4, N2O and the resulting GWP were derived using ANOVA-evaluated linear regression models.
Results
Meteorological and Hydrological Conditions
The study year 2020/2021 had an annual average of 8.6 °C and a total precipitation of 593 mm and was, therefore, warmer (+ 0.2 °C) and dryer (-82 mm) than the long-term average over the past 30 years. The annual average WTD for the site was − 0.10±-0.03 m, with no distinct differences between plots. However, from late May to October, WTD dropped to values below − 20 cm, dipping down to more than − 40 cm in August 2020. From November to February, all plots were inundated (Fig. 2a). θ varied on an annual basis between 59.9 ± 2.2% (plot 12, treatment 2) to 66.7 ± 1.8% (plot 13, treatment 5), with a site average of 64.1 ± 1.9%. The pH-corrected redox conditions in both depths (-5 cm and − 25 cm) differed among plots and subplots without a link to a potential treatment effect. For instance, redox conditions at -5 cm ranged between 227.8 ± 11.2 (plot 13) to 291.7 ± 17.7 (plot 12) Eh, with no distinct treatment-specific differences (Table 1).

Redox conditions, expressed as the for pH corrected redox potential (Eh), in the soil layer of -5 cm to -25 cm depth (left y axis) over the course of the study period for (a) each plot, and (b) each treatment. Blue lines indicate the volumetric water content (θ) in % (right y axis), while black lines show the fluctuation of water table depth (WTD) at cm soil depth (left y axis). For illustration, the green line shows interpolated emissions of CH4(in 1/100 µg m-2h-1, left y axis). Red dashed lines indicate zero
Biomass Yields and Carbon Content
Mean harvested biomass yields were similar for the treatments with two (10.6 ± 1.1 t DM ha− 1 yr− 1) and five annual cuts (9.4 ± 1.0 t DM ha− 1 yr− 1), resulting in the export of 4.7 ± 0.5 and 4.2 ± 0.5 t of C, and 0.25 ± 0.03 and 0.29 ± 0.03 t of N by biomass harvest (Table 2).
Carbon Dioxide
NEE for all treatments was positive on an annual basis, ranging between 3.8 ± 3.2 (2-cut) to 9.6 ± 2.2 (0-cut) t CO2-C ha− 1 yr− 1 with no statistical difference between the annual values (Table 3). However, the treatments with two and five annual cuts were both characterised by negative (net uptake) NEE during the early growing season in April-June (Fig. 3). This observation was absent for the 0-cut treatment. A similar pattern was observed for GPP, which was higher for the treatments subject to harvest. Here the 2-cut treatment showed the highest GPP, with peaks prior to harvest seen on a plot basis (Figure S1). Reco, however, was similar for all treatments, averaging 21.5 ± 2.7 t CO2-C ha− 1 yr− 1.

Averaged gap filled daily carbon (C) fluxes, partitioned into gross primary productivity (GPP), net ecosystem exchange (NEE), and ecosystem respiration (Reco), per treatment as indicated by the number of annual biomass cuts (0, 2, 5) throughout the study period from May 2020– May 2021
Methane Emissions
Methane emissions were low for all treatments (Table 3), with any significant difference. However, the highest value of cumulative emissions was found for the 2-cut treatment (0.5 ± 0.13 t CO2-C eq ha− 1 yr− 1) compared to the 0- and 5-cut treatments (0.3 ± 0.09 and 0.3 ± 0.05 t CO2-C eq ha− 1 yr− 1, respectively). Across plots and treatments, the highest CH4 emissions were observed at low Eh values, typically following increases in θ (Figure S2). In addition, while CH4 fluxes per measurement campaign correlated with soil temperature and GH, no effects on annual cumulative values were found (Tables S1, S2).
Nitrous Oxide
Both fluxes and annual cumulative emissions of N2O were high for all treatments, with no significant difference, despite 0.4 less N2O (in t CO2-C eq ha− 1 yr− 1) emitted from the treatment with five annual cuts (Table 3). In this context, no effect of harvest and fertilisation frequency was detected on neither N2O fluxes per measurement campaign nor annual cumulative emissions (Tables S1, S2). Inconsistent directions of treatment effects of harvest and fertilisation frequency also depicted this lack of correlation. Here, for instance, N2O emissions increased with management intensification in plot 19, while the opposite was observed in plot 12 (Fig. 4).

Cumulative annual values for methane (CH4), nitrous oxide (N2O), net ecosystem exchange (NEE) and the global warming potential (GWP). Values are given as means per treatment (bars) and individual values per plot (coloured dots), including the standard error (n = 4) of the mean. Red dashed lines indicate zero
Annual Greenhouse Gas Balances
The annual GHG balances of all treatments were dominated by NEE and the export of harvested C in biomass, together accounting for 81% (2-cut) to 88% (5-cut) of the GWP. NEE alone accounted for 84% of the GWP for the 0-cut treatment. For the treatment with five annual cuts, this resulted in the highest GWP of all treatments (11.5 ± 2.61 t CO2-C eq ha− 1 yr− 1), despite the lowest contribution of N2O and CH4. However, there was no significant difference regarding the GWP of the three treatments, all of which ranged around approximately 11 t CO2-C eq ha− 1 yr− 1 (Fig. 4).
Discussion
Management Intensity did not Affect the Global Warming Potential
In this study, the magnitudes of annual GHG emissions for RCG cultivation on a wet fen peatland under different intensities of harvest and fertilisation, with either zero, two, or five annual cuts, were assessed. Annual biomass production for the treatments including harvests was similar and within the range of previously reported yields for RCG in a temperate climate (Tilvikiene et al. 2016) but lower compared to a study in the same area and similar WTD (Karki et al. 2019). The similarity in biomass yields was also depicted by near-equal Reco emissions for all treatments, both aspects previously being correlated by Liu (2022). Given that gap-filled annual Reco on both the 0-cut and 2-cut treatment were equal (both 21.4 t CO2-C ha− 1 yr− 1), we estimated that aboveground biomass production on the 0-cut treatment also must have been similar. This assumption is supported by the previously reported yield of 8.3 t DM ha− 1 yr− 1 for RCG in the same study area, with one annual summer cut (Nielsen et al. 2021a). However, despite similar biomass growth and Reco, we found differing CO2 dynamics for partitioned fluxes of GPP and, subsequently, NEE. Here, the treatment with two annual cuts resulted in the smallest annual CO2 emissions of 3.8 t CO2-C ha− 1 yr− 1, which is in line with the IPCC 2013 EF for shallow-drained temperate grassland on nutrient-rich peat (IPCC 2014), but high in comparison to other studies. For instance, Schrier-Uijl et al. (2014) found an annual cumulative NEE of 1.1 t CO2-C ha− 1 yr− 1 for intensively and extensively managed sites in the Netherlands with similar drops in WTD over summer months as on our study site. However, a point of discussion in this context is the applicability of the annual mean WTD for comparisons between sites. For instance, according to Wilson et al. (2016a) and the IPCC (2014), our study site would be classified as rewetted due to the annual average WTD of -10 cm. Nevertheless, it is a site with an existing shallow drain system and reduced WTD during summer months. Another point of discussion is whether emissions or net carbon balances are reported. For instance, we found NEE emissions in the range of 3.8–9.6 t CO2-C ha− 1 yr− 1, depending on management, therewith being similar to the range of EFs for drained cropland or deeply-drained intensively used grassland (Wilson et al. 2016a; Weideveld et al. 2021). If also accounting for the export of biomass, known as the net ecosystem carbon balance (NECB), nowadays commonly applied following the IPCC guidelines (IPCC 2014), all our treatments were similar with regard to CO2 emissions, having a NECB of 8.5 (2-cut) to 10.1 (5-cut) t CO2-C ha− 1 yr− 1. However, the results were far above the recently defined EF for German peatlands, which were within the range of 0–4 t CO2-C ha− 1 yr− 1 for the same average WTD as on our study site (Tiemeyer et al. 2020). Thus, defining a universal range of CO2 emissions for wet peatlands under the land use of permanent grass or paludiculture is not a straightforward task due to the dependence on biomass yields as a response to soil nutrient and mineral status.
The observation of near-equal emission magnitudes for all management intensities did not change when also including annual emissions of CH4 and N2O. CH4 emissions were very low, while N2O emissions were unexpectedly high. However, annual emissions for both gases were close to equal among different management intensities. For instance, annual CH4 emissions were 0.3 t CO2-C eq ha− 1 yr− 1 for both extremes of assessed treatments: no harvest and fertilisation, as well as intensive management with five annual cuts. In the case of the 0-cut treatment, this might be explained by a combination of enhanced root growth (Nielsen et al. 2021b) in combination with potential CH4 oxidation by aerenchymatous oxygen transport to the root zone. However, an explanation the other way around is more likely: that dry soil in periods with enhanced soil temperature led to increased CH4 oxidation over the entire study area, disregarding of treatment. In addition, in cases of high captured CH4 fluxes, mainly on the 2-cut treatment, a lag-time effect has been observed– meaning that higher CH4 fluxes were detected delayed with regard to the occurrence of optimal conditions of θ and soil temperature. This lag-time effect has also been observed in a study by Yuan et al. (2021), who found that methanogenic bacteria are unable to utilise C substrate availability immediately after optimal soil moisture conditions are reached. According to a review by Abdalla et al. (2016) on CH4 emission in northern peatlands of different stages of naturalness, our observed CH4 emissions were way below values reported for natural fens (1.77 t CH4 in CO2-C eq t ha− 1 yr− 1), but above CH4 dynamics from drained fen peatlands (0.05 t CH4 in CO2-C eq t ha− 1 yr− 1). Thus, for all treatments, it was found that annual cumulative emissions of CH4 that were more within the range of dynamics for natural bog peatlands (0.1–1.0 t CH4 in CO2-C eq t ha− 1 yr− 1), usually associated with high rates of methanotrophy (Kolton et al. 2022). However, compared to a compilation of annual CH4 emissions for German grasslands (Tiemeyer et al. 2020), our observed values from a wet fen peatland with an annual WTD of -10 cm were higher than values reported for a similar mean WTD (0.05 t CH4 in CO2-C eq t ha− 1 yr− 1), as well as across all WTDs and under grassland-use (0.007 t CH4 in CO2-C eq t ha− 1 yr− 1).
Even regarding emissions of N2O, we did not find significant differences between treatments. This, however, is curious since the 0-cut treatment did not receive any N fertiliser application– and yet, emissions were equal to those from the 2-cut treatment, having received 200 kg of N h− 1 yr− 1. In other words, fertilisation did not result in higher N2O emissions for the 2-cut and 5-cut treatments. Overall, N2O emissions were high and with 1.1 (5-cut) and 1.5 (0-cut, 2-cut) t CO2-C eq t ha− 1 yr− 1 within the range of emissions for drained peatlands under the land-use of fertilised cropland (Wilson et al. 2016a). Considering the IPCC default EF (IPCC 2019) for direct N2O emissions from agricultural soil, a fixed percentage of approximately 1% of fertiliser-N applied, as well as priors of approximately 7.9 and 4.6 kg N2O-N ha− 1 yr− 1 for fertilised and unfertilised organic soils, respectively (Tiemeyer et al. 2020; Mathivanan et al. 2021), our results were exceeding average EFs. In detail, it was observed that the treatments with two and five annual cuts emitted 7.5% and 5.5% of the N applied. In the case of the 0-cut treatment without fertiliser application, the observed high emissions of N2O exceeded expectable prior emissions by 30% and were likely caused by N mineralisation during summer months with low WTD (Minkkinen et al. 2020). Considering the large observed N2O emissions from the prior on the unfertilised plots, the harvest of biomass is likely to have mitigated potentially higher N2O emissions for the fertilisation treatments due to plant rejuvenation (Walker et al. 2014; Tejera et al. 2022).
Although our methods effectively determined treatment effects, the sampling procedure may not represent actual GHG budgets due to its temporal constraints. For instance, Dooling et al. (2018) found that annual cumulative emissions for CH4 are underestimated if only based on daytime measurements, even if this might be an artefact resulting from nocturnal stratification (Stieger et al. 2015). Further, due to the bi-weekly measurement campaigns, it is without much doubt that we have not adequately captured hot moments of CH4 and N2O emissions, which contributed to 140 and 45% of mean annual fluxes in a study by Anthony and Silver (2021). In addition, also modelled daily values of CO2 fluxes, including their partitioning, are likely to not be flawless due to artificially derived PAR reduction by shrouding. Even though plants were given time to acclimate their photosynthesis rates to the reduced light levels before measurements were started, we could not capture fluxes under different temperature levels. Instead, a diurnal sampling campaign approach over naturally occurring levels of PAR and temperature would have been of benefit.
Water Table Depth is Only One Potential Proxy
For both main aspects related to this study, (1) biomass yields, including their accumulation of C and N, as well as (2) GHG emissions in dependence on management intensity, we did not find a treatment effect. However, we found intra-site variations that were distinct on a plot basis. First, in dependence on the distance to the adjacent Nørre Å river, organic carbon contents between plots with differences of up to 20% OC. This plot-specific difference was also found for pH and redox conditions. The most pronounced inter- and intra-plot specific differences were observed when relating the hydrological conditions of WTD, θ, and Eh to GHG fluxes. Typically, mean WTD levels are used as proxies for annual emission rates and are proposed to be utilised in national inventory reports (Tiemeyer et al. 2020). However, WTD is only one proxy, not necessarily being strictly correlated to two other critical hydrological aspects: the capillary fringe intensity (Gnatowski et al. 2002; Macrae et al. 2013) and the soil’s water holding capacity, which are also affected by precipitation rates (Irfan et al. 2020; Dai et al. 2022). As also observed in a study by Estop-Aragonés et al. (2012), soil moisture contents for our plots were not continuously developing parallel to changes in WTD. With both θ and Eh being related to suitable conditions for fluxes of CH4 and N2O (van den Pol-van Dasselaar et al. 1998; Wang et al., 2018b; Zhao et al. 2019; Korkiakoski et al. 2022), we found that soil-moisture related differences between plots explained the variation of observed emission magnitudes, irrespectively of treatment. Säurich et al. (2019) found the highest N2O production rates from denitrification at a water-filled pore space of 80–95%, which is similar to the average measured volumetric water contents of 60–67% during summer months for our site, but with distinct differences between sub-plots. Nonetheless, Jungkunst et al. (2008) and Tiemeyer et al. (2016) also found high N2O emissions of up to 12 kg N2O-N ha− 1 yr− 1 on sites with a similarly intermediate WTD of around − 20 cm to -10 cm. Furthermore, Berendt et al. (2022) concluded their study on the influence of peatland rewetting on N2O fluxes by stating that emissions were highly variable across states of soil wetness and related to hot moments and hot spots. In addition, the assessment of peat Eh revealed how quickly the border between aerated (Eh > + 400 mV, pH 7) and anaerobic (Eh < + 350 mV, pH 7) was fluctuating in the top 25 cm of soil depth, even in periods with low WTD. In our study, measurements of peat Eh supported the findings of both dynamics: high N2O flux rates and low CH4 emissions due to the lack of extended anaerobic conditions of Eh< -150 mV (pH 7). Thus, in accordance with previous studies (e.g.Masscheleyn et al. 1993; Pezeshki and DeLaune 2012; Wang et al. 2018 a,b; Zhang and Furman 2021), we found that peat Eh conditions were a better proxy explaining the variation in GHG dynamics than WTD.
To Harvest or Not to Harvest?
Our results showed that irrespective of management: (1) with and without fertilisation and (2) under different intensities of harvest, the GWP for all scenarios was similar. In the first instance, this implies that both extremes, a restoration scenario and intensive paludiculture practices were similarly climate-affecting land-use options. In areas where arable land is not a restricted resource, rewetting and restoration is likely to be the most cost-efficient option for atmospheric decarbonisation and ecological restoration (Mehrabi et al. 2018; Evans et al. 2021; Bianchi et al. 2021). In addition, previous research on belowground biomass (BGB) development under different harvest intensities showed significantly higher root biomass development for non-harvested RCG stands (Nielsen et al. 2021b). Therewith, direct C inputs from BGB, and long-term storage under water-logged conditions, are likely to be higher in non-harvested areas in the long run. However, in areas under pressure regarding issues of concurrent production and sustainable peatland management, intensive paludiculture practices might offer a solution to socioeconomically and environmentally responsible peatland agriculture (Wijedasa et al. 2016). Various options exist for paludiculture, ranging from the production of bioenergy plants to forage and protein concentrates for livestock feed (Damborg et al., 2019; Nielsen et al. 2021a; Martens et al. 2021). In this context, a frequent harvest of paludiculture biomass might add the benefit of nutrient removal and thus to the long-term biological restoration of peat environments (Hinzke et al. 2021; Vroom et al. 2022; Zak and McInnes 2022). However, our results are only based on one year of data and under hydrological conditions with a clear potential for improvements in WTD and its stability. Considering that peatlands have shown the potential for sudden developments regarding their C sink function (Roulet et al. 2007; Wilson et al. 2016b), our results only represent a snapshot in time regarding the effect of management intensities on the GWP for wet or rewetted fen peatlands.
Conclusion
This study aimed to assess whether intensities of two and five annual harvest occurrences at fertilisation rates of 200 kg N and K per ha− 1 yr− 1 will lead to differing carbon balances and gas exchange dynamics compared to a ‘nature scenario’ with neither harvest nor fertilisation.
First, CH4 emissions were low for a wet fen with a mean WTD of -10 cm but characterised by a drop in WTD in August of down to -40 cm. This was found for all plots and treatments due to soil moisture conditions and the associated redox potential, providing pathways for CH4 oxidation in the upper 25 cm of the peat layer when WTD was low and temperatures optimal. Contrary to the low emissions of CH4, we found substantially high flux rates of N2O, translating to EFs in the range of agricultural cropland. Here also, the unfertilised treatment was characterised by unforeseen high fluxes, which was interpreted as prior emissions from N mineralisation and denitrification during summer months, characterised by low WTD and an Eh of between + 300 and + 500 mV. Overall, it was found that peat Eh and θ conditions were better proxies explaining the variation in GHG dynamics than WTD.
Second, fertilisation and harvest did in no case of management intensity result in higher cumulative emissions of N2O due to the rejuvenescence of biomass. In addition, biomass yields, and their composition, were equal for both harvest intensities. Thus, while GPP was higher and NEE lower for the treatments with harvest occurrences, these C benefits were offset by the export of C in biomass as compared to the treatment without management. Based on these observations, our results highlighted a near-equal GWP in the range of 10.5–11.5 t CO2-C eq t ha− 1 yr− 1 for all scenarios irrespectively of management. In a climate context, our findings supported that both extremes, a restoration scenario but also intensive paludiculture practices, were similar land-use options regarding their climate impact. Finally, this indicates that site-specific hydrological and electrical peat properties are more critical regarding their climate impact than paludiculture management practices.
Data Availability
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
References
Anthony TL, Silver WL (2021) Hot moments drive extreme nitrous oxide and methane emissions from agricultural peatlands. Glob Chang Biol 27(20):5141–5153. https://doi.org/10.1111/gcb.15802
Berendt J, Jurasinski G, Wrage-Mönnig N (2022) Influence of rewetting on N2O emissions in three different Fen types. Nutr Cycl Agrosyst. https://doi.org/10.1007/s10705-022-10244-y
Bianchi A, Larmola T, Kekkonen H, Saarnio S, Lång K (2021) Review of Greenhouse Gas emissions from Rewetted Agricultural soils. Wetlands 41(8):108. https://doi.org/10.1007/s13157-021-01507-5
Buendia C, Tanabe E, Kranjc K, Baasansuren A, Fukuda J, Ngarize M, Osako S, Pyrozhenko A, Shermanau Y, P. and, Federici S (eds) (2019) 2019 refinement to the 2006 IPCC guidelines for National Greenhouse Gas inventories. IPCC, Switzerland
R Core Team (2021) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL: https://www.R-project.org/
Dai L, Fu R, Guo X, Du Y, Zhang F, Cao G (2022) Soil moisture variations in response to Precipitation Across different vegetation types on the northeastern Qinghai-Tibet Plateau. Front Plant Sci 13:854152. https://doi.org/10.3389/fpls.2022.854152
Dooling GP, Chapman PJ, Baird AJ, Shepherd MJ, Kohler T (2018) Daytime-Only Measurements Underestimate CH < sub > 4 Emissions from a Restored Bog. 25%J Ecoscience (3), 259–270, 212
Elsgaard L, Görres C-M, Hoffmann CC, Blicher-Mathiesen G, Schelde K, Petersen SO (2012) Net ecosystem exchange of CO2 and carbon balance for eight temperate organic soils under agricultural management. Agric Ecosyst Environ 162:52–67. https://doi.org/10.1016/j.agee.2012.09.001
Estop-Aragonés C, Knorr K-H, Blodau C (2012) Controls on in situ oxygen and dissolved inorganic carbon dynamics in peats of a temperate Fen. 117. https://doi.org/10.1029/2011JG001888
Evans CD, Peacock M, Baird AJ, Artz RRE, Burden A, Callaghan N, Morrison R (2021) Overriding water table control on managed peatland greenhouse gas emissions. Nature 593(7860):548–552. https://doi.org/10.1038/s41586-021-03523-1
Fraser LH, Pither J, Jentsch A, Sternberg M, Zobel M, Askarizadeh D, Zupo T (2015) Worldwide evidence of a unimodal relationship between productivity and plant species richness. 349(6245):302–305. https://doi.org/10.1126/science.aab3916
Fuss R, Hueppi R, Asger RP (2020) Gasfluxes: Greenhouse gas flux calculation from chamber measurements. R Package Version 0.4-4
Gnatowski T, Szatyłowicz J, Brandyk T (2002) Effect of peat decomposition on the capillary rise in peat-moorsh soils. 16(2):97–102
Günther A, Huth V, Jurasinski G, Glatzel S (2015) The effect of biomass harvesting on greenhouse gas emissions from a rewetted temperate Fen. 7(5):1092–1106. https://doi.org/10.1111/gcbb.12214
Günther A, Barthelmes A, Huth V, Joosten H, Jurasinski G, Koebsch F, Couwenberg J (2020) Prompt rewetting of drained peatlands reduces climate warming despite methane emissions. Nat Commun 11(1):1644. https://doi.org/10.1038/s41467-020-15499-z
Heinsoo K, Hein K, Melts I, Holm B, Ivask M (2011) Reed Canary grass yield and fuel quality in Estonian farmers’ fields. Biomass Bioenergy 35(1):617–625. https://doi.org/10.1016/j.biombioe.2010.10.022
Hinzke T, Tanneberger F, Aggenbach C, Dahlke S, Knorr K-H, Kotowski W, Kreyling J (2021) Can nutrient uptake by Carex counteract eutrophication in Fen peatlands? Sci Total Environ 785:147276. https://doi.org/10.1016/j.scitotenv.2021.147276
Hoffmann M, Jurisch N, Borraz A, Hagemann E, Drösler U, Sommer M, M., Augustin J (2015) Automated modeling of ecosystem CO2 fluxes based on periodic closed chamber measurements: a standardized conceptual and practical approach. Agric for Meteorol 200:30–45. https://doi.org/10.1016/j.agrformet.2014.09.005
Hüppi R, Felber R, Krauss M, Six J, Leifeld J, Fuß R (2018) Restricting the nonlinearity parameter in soil greenhouse gas flux calculation for more reliable flux estimates. PLoS ONE 13(7):e0200876. https://doi.org/10.1371/journal.pone.0200876
IPCC (2014) 2013 supplement to the 2006 IPCC guidelines for national greenhouse gas inventories: Wetlands
Irfan M, Setya OC, Virgo F, Sutopo, Ariani A, Sulaiman M, Iskandar I (2020) Is there a correlation between rainfall and soil moisture on peatlands in South Sumatra? J Phys: Conf Ser 1572(1):012040. https://doi.org/10.1088/1742-6596/1572/1/012040
Iyer G, Ou Y, Edmonds J, Fawcett AA, Hultman N, McFarland J, McJeon H (2022) Ratcheting of climate pledges needed to limit peak global warming. Nat Clim Change. https://doi.org/10.1038/s41558-022-01508-0
Järveoja J, Peichl M, Maddison M, Soosaar K, Vellak K, Karofeld E, Mander Ü (2016) Impact of water table level on annual carbon and greenhouse gas balances of a restored peat extraction area. Biogeosciences 13(9):2637–2651. https://doi.org/10.5194/bg-13-2637-2016
Jungkunst HF, Flessa H, Scherber C, Fiedler S (2008) Groundwater level controls CO2, N2O and CH4 fluxes of three different hydromorphic soil types of a temperate forest ecosystem. Soil Biol Biochem 40(8):2047–2054. https://doi.org/10.1016/j.soilbio.2008.04.015
Jurasinski G, Koebsch F, Guenther A, Beetz S (2014) flux: Flux rate calculation from dynamic closed chamber measurements. R package version 0.3-0. https://CRAN.R-project.org/package=flux
Kalisz B, Urbanowicz P, Smólczyński S, Orzechowski M (2021) Impact of siltation on the stability of organic matter in drained peatlands. Ecol Ind 130:108149. https://doi.org/10.1016/j.ecolind.2021.108149
Karki S, Kandel T, Elsgaard L, Labouriau R, Lærke PJM, Peat (2019) Annual CO 2 fluxes from a cultivated fen with perennial grasses during two initial years of rewetting. 25
Köhler W, Schachtel G, Voleske P (2012) Biostatistik: Eine Einführung für Biologen Und Agrarwissenschaftler. Springer-
Kolton M, Weston DJ, Mayali X, Weber PK, McFarlane KJ, Pett-Ridge J, Kostka JE (2022) Defining the < i > Sphagnum Core Microbiome across the North American Continent reveals a central role for Diazotrophic Methanotrophs in the Nitrogen and Carbon Cycles of Boreal Peatland Ecosystems. 13(1):e03714–03721. https://doi.org/10.1128/mbio.03714-21
Korkiakoski M, Määttä T, Peltoniemi K, Penttilä T, Lohila A (2022) Excess soil moisture and fresh carbon input are prerequisites for methane production in podzolic soil. Biogeosciences 19(7):2025–2041. https://doi.org/10.5194/bg-19-2025-2022
Kragbæk Damborg V, Krogh Jensen S, Johansen M, Ambye-Jensen M, Weisbjerg MR (2019) Ensiled pulp from biorefining increased milk production in dairy cows compared with grass–clover silage. J Dairy Sci 102(10):8883–8897. https://doi.org/10.3168/jds.2018-16096
Kukk L, Astover A, Roostalu H, Rossner H, Tamm IJAR (2010) The dependence of reed canary grass (Phalaris arundinacea L.) energy efficiency and profitability on nitrogen fertilization and transportation distance. 8(1):123–133
Larmola T, Tuittila ES, Tiirola M, Nykänen H, Martikainen PJ, Yrjälä K, Fritze H (2010) The role of Sphagnum mosses in the methane cycling of a boreal mire. Ecology 91(8):2356–2365. https://doi.org/10.1890/09-1343.1
Liu Y (2022) Grazing rest during spring regreening period promotes the ecological restoration of degraded alpine meadow vegetation through enhanced plant photosynthesis and respiration. Front Plant Sci 13:1008550. https://doi.org/10.3389/fpls.2022.1008550
Liu W, Fritz C, Weideveld STJ, Aben RCH, van den Berg M, Velthuis M (2022) Annual CO2 Budget Estimation from Chamber-based flux measurements on intensively drained Peat Meadows: Effect of gap-filling strategies. 10. https://doi.org/10.3389/fenvs.2022.803746
Macrae ML, Devito KJ, Strack M, Waddington JM (2013) Effect of water table drawdown on peatland nutrient dynamics: implications for climate change. Biogeochemistry 112:661–677
Malinowski R, Groom G, Schwanghart W, Heckrath G (2015) Detection and delineation of localized flooding from WorldView-2 Multispectral Data. 7(11):14853–14875
Marra G, Wood SN (2011) Practical variable selection for generalized additive models. Comput Stat Data Anal 55(7):2372–2387. https://doi.org/10.1016/j.csda.2011.02.004
Martens M, Karlsson NPE, Ehde PM, Mattsson M, Weisner SEB (2021) The greenhouse gas emission effects of rewetting drained peatlands and growing wetland plants for biogas fuel production. J Environ Manage 277:111391. https://doi.org/10.1016/j.jenvman.2020.111391
Masscheleyn PH, DeLaune RD, Patrick WH (1993) Methane and nitrous oxide emissions from laboratory measurements of rice soil suspension: Effect of soil oxidation-reduction status. Chemosphere 26(1):251–260. https://doi.org/10.1016/0045-6535(93)90426-6
Mathivanan GP, Eysholdt M, Zinnbauer M, Rösemann C, Fuß R (2021) New N2O emission factors for crop residues and fertiliser inputs to agricultural soils in Germany. Agric Ecosyst Environ 322:107640. https://doi.org/10.1016/j.agee.2021.107640
Mehrabi Z, Ellis EC, Ramankutty N (2018) The challenge of feeding the world while conserving half the planet. Nat Sustain 1(8):409–412. https://doi.org/10.1038/s41893-018-0119-8
Meinshausen M, Lewis J, McGlade C, Gütschow J, Nicholls Z, Burdon R, Hackmann B (2022) Realization of Paris Agreement pledges may limit warming just below 2°C. Nature 604(7905):304–309. https://doi.org/10.1038/s41586-022-04553-z
Minke M, Augustin J, Burlo A, Yarmashuk T, Chuvashova H, Thiele A, Hoffmann M (2016) Water level, vegetation composition, and plant productivity explain greenhouse gas fluxes in temperate cutover fens after inundation. Biogeosciences 13(13):3945–3970. https://doi.org/10.5194/bg-13-3945-2016
Minkkinen K, Ojanen P, Koskinen M, Penttilä T (2020) Nitrous oxide emissions of undrained, forestry-drained, and rewetted boreal peatlands. For Ecol Manag 478:118494. https://doi.org/10.1016/j.foreco.2020.118494
Moore TR, Dalva M (1993) The influence of temperature and water table position on carbon dioxide and methane emissions from laboratory columns of peatland soils. 44(4):651–664. https://doi.org/10.1111/j.1365-2389.1993.tb02330.x
Myhre G, Shindell D, Bréon F-M, Collins W, Fuglestvedt J, Huang J, Koch D, Lamarque J-F, Lee D, Mendoza B, Nakajima T, Robock A, Stephens G, Takemura T, Zhang H (2013) Anthropogenic and natural Radiative forcing. In: Qin TFD, Plattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM (eds) Climate Change 2013: the physical science basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [Stocker. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA
Næss JS, Hu X, Gvein MH, Iordan C-M, Cavalett O, Dorber M, Cherubini F (2023) Climate change mitigation potentials of biofuels produced from perennial crops and natural regrowth on abandoned and degraded cropland in nordic countries. J Environ Manage 325:116474. https://doi.org/10.1016/j.jenvman.2022.116474
Nielsen CK, Stødkilde L, Jørgensen U, Lærke PE (2021a) Effects of Harvest and Fertilization frequency on protein yield and Extractability from Flood-Tolerant Perennial Grasses cultivated on a Fen Peatland. 9. https://doi.org/10.3389/fenvs.2021.619258
Nielsen CK, Jørgensen U, Lærke PE (2021b) Root-To-Shoot ratios of Flood-Tolerant Perennial grasses depend on Harvest and Fertilization Management: implications for quantification of Soil Carbon Input. 9. https://doi.org/10.3389/fenvs.2021.785531
Noyce GL, Megonigal JP (2021) Biogeochemical and plant trait mechanisms drive enhanced methane emissions in response to whole-ecosystem warming. Biogeosciences 18(8):2449–2463. https://doi.org/10.5194/bg-18-2449-2021
Oestmann J, Tiemeyer B, Düvel D, Grobe A, Dettmann U (2022) Greenhouse Gas Balance of Sphagnum Farming on highly decomposed Peat at former Peat extraction sites. Ecosystems 25(2):350–371. https://doi.org/10.1007/s10021-021-00659-z
Petersen SO, Hoffmann CC, Schäfer CM, Blicher-Mathiesen G, Elsgaard L, Kristensen K, Greve MH (2012) Annual emissions of CH < sub > 4 and N < sub > 2 O, and ecosystem respiration, from eight organic soils in Western Denmark managed by agriculture. Biogeosciences 9(1):403–422. https://doi.org/10.5194/bg-9-403-2012.
Pezeshki SR, DeLaune RD (2012) Soil oxidation-reduction in wetlands and its impact on plant functioning. Biology (Basel) 1(2):196–221. https://doi.org/10.3390/biology1020196
Roulet NT, Lafleur PM, Richard PJH, Moore TR, Humphreys ER, Bubier J (2007) Contemporary carbon balance and late Holocene carbon accumulation in a northern peatland. 13(2):397–411. https://doi.org/10.1111/j.1365-2486.2006.01292.x
Säurich A, Tiemeyer B, Dettmann U, Don A (2019) How do sand addition, soil moisture and nutrient status influence greenhouse gas fluxes from drained organic soils? Soil Biol Biochem 135:71–84. https://doi.org/10.1016/j.soilbio.2019.04.013
Schrier-Uijl AP, Kroon PS, Hendriks DMD, Hensen A, Van Huissteden J, Berendse F, Veenendaal EM (2014) Agricultural peatlands: towards a greenhouse gas sink– a synthesis of a Dutch landscape study. Biogeosciences 11(16):4559–4576. https://doi.org/10.5194/bg-11-4559-2014
Skinner RH (2008) High biomass removal limits Carbon Sequestration potential of mature temperate pastures. 37(4):1319–1326. https://doi.org/10.2134/jeq2007.0263
Stieger J, Bamberger I, Buchmann N, Eugster W (2015) Validation of farm-scale methane emissions using nocturnal boundary layer budgets. Atmos Chem Phys 15(24):14055–14069. https://doi.org/10.5194/acp-15-14055-2015
Tanneberger F, Appulo L, Ewert S, Lakner S, Brolcháin Ó, Peters N, J., Wichtmann W (2021) The power of Nature-based solutions: how Peatlands can help us to Achieve Key EU sustainability objectives. Adv Sustainable Syst 5(1):2000146. https://doi.org/10.1002/adsu.202000146
Tejera M, Boersma NN, Archontoulis SV, Miguez FE, VanLoocke A, Heaton EA (2022) Photosynthetic decline in aging perennial grass is not fully explained by leaf nitrogen. J Exp Bot. https://doi.org/10.1093/jxb/erac382
Thomsen AJIRPP (2006) ManTDR software for making manual TDR measurements. 3
Tiemeyer B, Albiac Borraz E, Augustin J, Bechtold M, Beetz S, Beyer C, Zeitz J (2016) High emissions of greenhouse gases from grasslands on peat and other organic soils. 22(12):4134–4149. https://doi.org/10.1111/gcb.13303
Tiemeyer B, Freibauer A, Borraz EA, Augustin J, Bechtold M, Beetz S, Drösler M (2020) A new methodology for organic soils in national greenhouse gas inventories: data synthesis, derivation and application. Ecol Ind 109:105838. https://doi.org/10.1016/j.ecolind.2019.105838
Tilvikiene V, Kadziuliene Z, Dabkevicius Z, Venslauskas K, Navickas K (2016) Feasibility of tall fescue, cocksfoot and reed canary grass for anaerobic digestion: analysis of productivity and energy potential. Ind Crops Prod 84:87–96. https://doi.org/10.1016/j.indcrop.2016.01.033
Ustak S, Šinko J, Munoz JJJ, o. CEA (2019) Reed Canary grass (Phalaris arundinacea L.) as a promising energy crop. 20(4):1143–1168. https://doi.org/10.5513/JCEA01/20.4.2267
Utama DT, Lee SG, Baek KH, Chung WS, Chung IA, Kim DI, Lee SK (2018) Blood profile and meat quality of Holstein-Friesian steers finished on total mixed ration or flaxseed oil-supplemented pellet mixed with reed canary grass haylage. Animal 12(2):426–433. https://doi.org/10.1017/S1751731117001707
van den Pol-van Dasselaar A, van Beusichem ML, Oenema O (1998) Effects of soil moisture content and temperature on methane uptake by grasslands on sandy soils. Plant Soil 204(2):213–222. https://doi.org/10.1023/A:1004371309361
Vroom RJE, Geurts JJM, Nouta R, Borst ACW, Lamers LPM, Fritz C (2022) Paludiculture crops and nitrogen kick-start ecosystem service provisioning in rewetted peat soils. Plant Soil 474(1):337–354. https://doi.org/10.1007/s11104-022-05339-y
Vroom RJE, van den Berg M, Pangala SR, van der Scheer OE, Sorrell BK (2022) Physiological processes affecting methane transport by wetland vegetation– A review. Aquat Bot 182:103547. https://doi.org/10.1016/j.aquabot.2022.103547
Walker AP, Beckerman AP, Gu L, Kattge J, Cernusak LA, Domingues TF, Woodward FI (2014) The relationship of leaf photosynthetic traits– vcmax and jmax– to leaf nitrogen, leaf phosphorus, and specific leaf area: a meta-analysis and modeling study. 4(16):3218–3235. https://doi.org/10.1002/ece3.1173
Wang J, Bogena HR, Vereecken H, Brüggemann N (2018) Characterizing redox potential effects on Greenhouse Gas Emissions Induced by Water-Level Changes. Vadose Zone J 17(1):170152. https://doi.org/10.2136/vzj2017.08.0152
Wang D, Zang S, Wu X, Ma D, Li M, Chen Q, Zhang N (2021) Soil organic carbon stabilization in permafrost peatlands. Saudi J Biol Sci 28(12):7037–7045. https://doi.org/10.1016/j.sjbs.2021.07.088
Webb EK, Pearman GI, Leuning R (1980) Correction of flux measurements for density effects due to heat and water vapour transfer. 106(447):85–100. https://doi.org/10.1002/qj.49710644707
Weideveld STJ, Liu W, van den Berg M, Lamers LPM, Fritz C (2021) Conventional subsoil irrigation techniques do not lower carbon emissions from drained peat meadows. Biogeosciences 18(12):3881–3902. https://doi.org/10.5194/bg-18-3881-2021
Wijedasa LS, Page SE, Evans CD, Osaki M (2016) Time for responsible peatland agriculture. 354(6312):562–562. https://doi.org/10.1126/science.aal1794
Wilson D, Blain D, Couwenberg J, Evans C, Murdiyarso D, Page S, Tuittila E (2016a) Greenhouse gas emission factors associated with rewetting of organic soils
Wilson D, Farrell CA, Fallon D, Moser G, Müller C, Renou-Wilson F (2016b) Multiyear greenhouse gas balances at a rewetted temperate peatland. Glob Chang Biol 22(12):4080–4095. https://doi.org/10.1111/gcb.13325
Wood SN (2011) Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J Royal Stat Society: Ser B (Statistical Methodology) 73(1):3–36. https://doi.org/10.1111/j.1467-9868.2010.00749.x
Wood SN (2022) Mgcv: Mixed Gam computation vehicle with automatic smoothness estimation. R package version 1.8–39. https://cran.r-project.org/web/packages/mgcv/mgcv.pdf
Wood SN, Pya N, Säfken B (2016) Smoothing parameter and model selection for general smooth models. J Am Stat Assoc 111(516):1548–1563. https://doi.org/10.1080/01621459.2016.1180986
Yang Y, Dou Y, An S, Zhu Z (2018) Abiotic and biotic factors modulate plant biomass and root/shoot (R/S) ratios in grassland on the Loess Plateau, China. Sci Total Environ 636:621–631. https://doi.org/10.1016/j.scitotenv.2018.04.260
Yuan F, Wang Y, Ricciuto DM, Shi X, Yuan F, Brehme T, Xu X (2021) Hydrological feedbacks on peatland CH4 emission under warming and elevated CO2: a modeling study. J Hydrol 603:127137. https://doi.org/10.1016/j.jhydrol.2021.127137
Zak D, McInnes RJ (2022) A call for refining the peatland restoration strategy in Europe. 59(11):2698–2704. https://doi.org/10.1111/1365-2664.14261
Zhang Z, Furman A (2021) Soil redox dynamics under dynamic hydrologic regimes - a review. Sci Total Environ 763:143026. https://doi.org/10.1016/j.scitotenv.2020.143026
Zhang X, Hu L, Yang C, Zhou C, Yuan L, Chen Z, Seago JL (2017) Structural features of Phalaris arundinacea in the Jianghan Floodplain of the Yangtze River, China. Flora 229:100–106. https://doi.org/10.1016/j.flora.2017.02.016
Zhao J-F, Peng S-S, Chen M-P, Wang G-Z, Cui Y-B, Liao L-G, Tan Z-H (2019) Tropical forest soils serve as substantial and persistent methane sinks. Sci Rep 9(1):16799. https://doi.org/10.1038/s41598-019-51515-z
Acknowledgements
The authors would like to thank the technical and laboratory staff at the Departments of Agroecology, Aarhus University, for excellent guidance as well as assistance with crop management and analyses. The concept for this paper was developed at the workshop titled “Peatlands for climate change mitigation in agriculture” that took place in Aarhus, Denmark, on 4–5 October 2022, and which was sponsored by the Organisation for Economic Co-operation and Development (OECD) Co-operative Research Programme: Sustainable Agricultural and Food Systems. The opinions expressed and arguments employed in this publication are the sole responsibility of the authors and do not necessarily reflect those of the OECD or of the governments of its Member countries.
Funding
This study was financially supported by the PEATWISE project (https://www.eragas.eu/en/eragas/Research-projects/PEATWISE.htm) in the frame of the ERA-NET FACCE ERA-GAS, and the projects RePeat DK and INSURE. FACCE ERA-GAS and INSURE received funding from the European Union’s Horizon 2020 research and innovation programme under the grant agreement no. 696356 and 862695, respectively. Further, the study was partly supported by the Aarhus University Center for Circular Bioeconomy (CBIO, https://cbio.au.dk/en/). In addition, CN received funding by the European Union under Horizon Europe (WET HORIZONS, grant agreement no. 101056848).
Open access funding provided by Aarhus Universitet
Author information
Authors and Affiliations
Contributions
All authors contributed to the study design, reading and revision of the manuscript, and approved the final version. CN developed and performed the study design and experimental work, analysis of the data, and writing of the manuscript. WL developed the script for modelling. MK contributed significantly to data collection in the field. PL was involved in gathering of funds.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nielsen, C.K., Liu, W., Koppelgaard, M. et al. To Harvest or not to Harvest: Management Intensity did not Affect Greenhouse Gas Balances of Phalaris Arundinacea Paludiculture. Wetlands 44, 79 (2024). https://doi.org/10.1007/s13157-024-01830-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13157-024-01830-7