
Abstract
Over 300,000 ha of forested wetlands have undergone restoration within the Mississippi Alluvial Valley region. Restored forest successional stage varies, providing opportunities to document wetland functional increases across a large scale restoration chronosequence using the Hydrogeomorphic (HGM) approach. Results from >600 restored study sites spanning a 25 year chronosequence indicate that: 1) wetland functional assessment variables increased toward reference conditions; 2) restored wetlands generally follow expected recovery trajectories; and 3) wetland functions display significant improvements across the restoration chronosequence. A functional lag between restored areas and mature reference wetlands persists in most instances. However, a subset of restored sites have attained mature reference wetland conditions in areas approaching or exceeding tree diameter and canopy closure thresholds. Study results highlight the importance of site selection and the benefits of evaluating a suite of wetland functions in order to identify appropriate restoration success milestones and design monitoring programs. For example wetland functions associated with detention of precipitation (a largely physical process) rapidly increased under post restoration conditions, while improvements in wetland habitat functions (associated with forest establishment and maturation) required additional time. As the wetland science community transitions towards larger scale restoration efforts, effectively quantifying restoration functional improvements will become increasingly important.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wetland Restoration in the Mississippi Alluvial Valley
Wetlands provide a variety of well-established hydrologic, biogeochemical, and habitat functions linked to ecosystem services that prove beneficial to society (Smith et al. 1995; Novitski et al. 1996). In particular, bottomland hardwood (BHW) forests within the Mississippi Alluvial Valley exhibit wetland functions including the retention and temporary storage of precipitation and floodwater; nutrient cycling, transformation of elements and compounds, and organic carbon export; and the support of faunal and floral habitat (Smith and Klimas 2002). Over 10 million ha of BHW once occurred in the region; however extensive alteration to the landscape have resulted in a 70% decline in BHW habitat (The Nature Conservancy 1992; King et al. 2006). Notably, rates of BHW alteration has outpaced impacts to other wetland types, creating an area of concern in terms of reduction in wetland extent and function (Hefner and Brown 1995). Wetland impacts resulted from a variety of factors including habitat fragmentation, settlement expansion, agriculture and forestry, and flood control activities (Gardiner and Oliver 2005).
Public and private organizations recognized the negative impacts of BHW wetland degradation during the 1970s and 1980s and began promoting wetland restoration via afforestation to repair degraded ecosystem functions (US Congress 1985; Haynes et al. 1995; Hobbs and Cramer 2008). In response, an estimated 300,000 has undergone reforestation under the US Department of Agriculture Natural Resources Conservation Service Wetland Reserve Program and other initiatives (King and Keeland 1999; Allen et al. 2000; King et al. 2006). Additionally, the US Army Corps of Engineers (USACE) restored >11,000 ha of BHW wetlands to offset unavoidable adverse impacts associated with construction of flood control and navigation projects (Haynes and Moore 1988; USACE 1989). Restoration projects reclaim BHW forests previously converted to agriculture, many of which exhibited marginal production due to seasonal high water tables and/or the need for extensive drainage (Stanturf et al. 1998; Berkowitz 2013). Restoration activities include planting desirable BHW tree species, selected for their capacity to thrive under wetland soil and hydrologic conditions (Stanturf et al. 2000; Humphrey et al. 2004). Characteristic species utilized for restoration include Fraxinus pennsylvanica, Quercus nuttallii, Quercus lyrata, Carya aquatica, and other flood-tolerant hydrophytes associated with high value wetland habitats (Smith and Klimas 2002). Afforestation typically occurs via row planting at typical seedling spacing of 3–4 m2. Restoration locations often contain access roads but remain free of other alterations, managed as natural areas as designated through fee title agreements with state and local agencies or private landholders. Recreational birding, hunting, and other activities occur on a subset of restoration locations, providing additional public benefits (Jenkins et al. 2010).
The USACE wetland restoration initiative began in 1990, representing some of the oldest large-scale restored BHW wetland tracts in the region, with periodic additional land acquisition occurring over time (Lin 2009). Approximately 6500 ha underwent restoration by the year 2000, with an additional 3800 ha completed restoration by 2010. Recent efforts continue to expand the extent of BHW restoration projects with the addition of 1550 ha since 2011. The periodicity of afforestation provides a mechanism for examining restoration success across a chronosequence exhibiting wetland functions under various conditions as forest succession occurs. Establishing the restoration chronosequence also allows for an evaluation of project benefits, development of predictive restoration trajectories, as well as the establishment and monitoring of restoration project milestones (Wigginton et al. 2000; Walker et al. 2010). Berkowitz (2013) previously utilized the restoration chronosequence, identifying restoration assessment metrics providing rapid response indicators of restoration success. That work demonstrated that readily available indicators of restoration trajectory such as shrub-sapling density and ground vegetation cover responded to restoration during short (<5) and mid-term (<10 year) timeframes, while other indicators (e.g., tree diameter maturation) required longer time periods (>15–20 years) to provide useful insight into wetland functional outcomes. The current study returned to the chronosequence to further evaluate restoration conditions, with an increased focus on quantifying large scale wetland functional increases as restored BHW wetlands approach canopy closure thresholds.
Wetland Functional Assessment
To support USACE wetland restoration initiatives in the region, Smith and Klimas (2002) developed a Hydrogeomorphic (HGM) wetland functional assessment method specifically designed for the Mississippi Alluvial Valley. The HGM method addressed seven wetland functions provided by BHW wetlands, and included approaches to determine conditions in both mature and restored areas (Smith et al. 2013). The HGM approach utilizes geomorphic, vegetative, and structural measurements to approximate wetland functions (Brinson 1993, 1995; Rowe et al. 2009). Specifically, in the HGM approach readily attainable structural metrics (e.g., tree diameter, ground cover) are combined using simple empirical arithmetic relationships to produce functional capacity index scores ranging from zero (a lack of wetland function) to 1.0 (fully functional) (Smith et al. 1995). The use of HGM techniques maintains several advantages over other ecological assessment approaches by incorporating 1) ecosystem classification, 2) collection of quantitative data, and 3) scaling based on reference data (Clairain 2002). As a result, the HGM approach was approved for use by several US federal resource management agencies and has been upheld in several recent US court decisions as a legally defensible and acceptable methodology to assess ecological resources (Federal Register 1997; Ohio Valley Environmental Coalition, Inc, et al. vs. US Army Corps of Engineers et al. 2012; Noble et al. 2014). Bauder et al. (2009) described the HGM approach as more accurate than other wetland assessment methods, and Cole (2006) while generally critical of wetland functional assessments identified HGM as the best available method for rapidly determining wetland function. Recent research demonstrated the applicability of the HGM methodology to restoration progress over time, with Berkowitz and White (2013) validating the wetland assessment approach by linking wetland assessment metrics with direct measurements of wetland function. The HGM assessment provided a reliable proxy for direct measurements of nutrient and biogeochemical cycling functions (e.g., microbial biomass flux), and documented that wetland functions increased with restoration stand age across the chronosequence described above. Similar validation results were reported following evaluation of a HGM functional assessment method applied to the Appalachian mountain region in locations exhibiting a range of alterations (Berkowitz et al. 2014; Noble et al. 2014).
Despite numerous publications addressing afforestation and restoration within the Mississippi Alluvial Valley, questions persist regarding the mid- to long-term success of large scale restoration efforts designed to improve BHW wetland function (Stanturf et al. 2001) and the applicability of existing assessment approaches designed to quantify those functions (Cole 2016). In response, the current study 1) evaluated changes in wetland assessment metrics and function across the restoration chronosequence, 2) examined restoration trajectories incorporating the oldest available restored BHW wetlands in the region, and 3) compared the levels of restored wetland function with mature BHW forested wetlands.
Methods
Study Locations
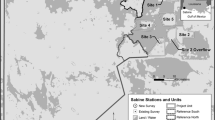
The current effort focused on 12 restored bottomland hardwood restoration study locations within the Mississippi Alluvial Valley (Fig. 1). Restoration tracts were selected by identifying poorly drained, frequently flooded agricultural lands within the region that historically supported BHW forested wetlands. Potential restoration tracts were selected based upon their reforestation potential, proximity to existing BHW habitat, and opportunities to connect or expand existing state or federally managed conservation lands. The restored BHW forests examined herein are subject to overbank, backwater flooding from rivers, streams, and direct precipitation at typical wetland hydrology return intervals of 5 yrs or less (Smith and Klimas 2002; Berkowitz and White 2013). Backwater flooding describes inundation resulting from impeded drainage, usually due to high water in downstream systems. Typically, backwater flooding occurs when a large stream in flood stage inhibits drainage within adjacent tributaries. Impeded drainage leads to increasing water tables and surface inundation. In the Mississippi Alluvial Valley additional backwater flooding results from structures associated with flood control projects such as levees. Many BHW forests in the study area occupy backswamp deposits and other low-lying areas near tributary networks. Prior to restoration, study locations were managed for row crop production including corn, soybeans, cotton, and rice varieties. Hydric soils dominating the study area include Sharkey clay (Very-fine, smectitic, thermic Chromic Epiaquerts), Alligator clay (Very-fine, smectitic, thermic Chromic Dystraquerts), Forestdale silty clay loam (Fine, smectitic, thermic Typic Endoaqualfs), and related series (United States Department of Agriculture, Natural Resources Conservation Service 2016; Soil Survey Staff 2017). Restored BHW wetland ages ranged from 5 to 25 years post restoration, providing a chronosequence of post restoration conditions. Additionally, data from 26 mature BHW forest locations within the region was selected for comparison with the restored wetland areas (Berkowitz and White 2013). Tree core analysis indicated that the canopy trees within each mature BHW forests were > 80 years old.

Location of BHW wetland restoration sites within the Mississippi Alluvial Valley region
Wetland Functional Assessment Approach
The wetland functional assessment completed at each sample transect location applied the HGM method developed by Smith and Klimas (2002) who provide a thorough description of field data collection and analysis. Briefly, sample transects were established at each restoration location during the period of May through September 2016. Along sample transects, multiple sample sites were established. The distribution of sample transects and sampling intensity varied depending upon the size and heterogeneity of restoration locations, with additional data collected in larger, more diverse locations (Table 1). At each sample site the HGM assessment approach utilized 17 variables incorporating both remote sensing and ground based measurements (Table 2). Remote sensing resources determined the assessment scores for the core area; habitat connection; wetland tract; and flood frequency variables. A combination of remote sensing and ground based observations were used to determine cation exchange capacity and soil integrity assessment variables, which documented the degree and extent of soil disturbance.
Direct field measurements addressed each of the remaining variables using data collected within a 0.04 ha sample plot and associated sub-plots and transects. Micro-depressional ponding quantified the percentage of each sample plot characterized by concave features capable of retaining water. Tree basal area examined the diameter of each tree with a diameter at breast height (DBH) ≥10 cm within the 0.04 ha sample plot. Notably, most trees (estimated at >95%) with DBH ≥10 cm observed at restoration sites were planted and rows remained visible at many sample locations. Natural recruitment of other woody species was observed, but few of those individuals had reached a DBH of 10 cm. The density of trees and snags exhibiting DBH ≥10 within the sample plot was determined, along with the species composition of the highest strata within the 0.04 ha plot. Shrub-sapling density measurements utilized replicate nested 0.004 ha subplots. Woody debris and log biomass measurements were conducted along replicate linear transects. Finally, four replicate one m2 subplots were used to characterize ground vegetation cover as well as the thickness of both soil O- and A-horizons.
Following the measurement of each assessment variable, the variables are standardized and converted into variable subindex scores varying from zero to 1.0 as described in Smith and Klimas (2002). Once generated, variable subindex scores are combined using assessment equations to determine wetland functional capacity scores which range from zero representing a lack of wetland function to 1.0 indicating the highest achievable level of wetland function in the region (Table 3; Smith et al. 2013).
Statistical analysis of changes in wetland functional assessment variables and functional capacity scores were conducted across the restoration chronosequence using analysis of variance (ANOVA) where assumptions of normality (Shapiro and Wilk 1965) and homogeneity of variance (Levene 1960) were met. For the ANOVA, sample plot age classes were utilized to group restoration locations because many restoration plantings span calendar years; age classes and the number of replicate sample plots [n] were as follows: 5 [97], 10 [61], 13 [209], 20 [115], 25 [124]. Wetland functional assessment variables and functional capacity scores were considered dependent factors, with restoration chronosequence age classes applied as grouping variables. Data were square root or log transformed as needed to meet model assumptions. Post-hoc testing utilized the Tukey test. The non-parametric Kruskal-Wallis test evaluated differences between restoration age classes when data failed to meet model assumptions, followed by Dunn-Bonferroni post hoc testing (Dunn 1964). All statistical significance was evaluated at the α = 0.05 level using SPSS version 20.0 (IBM, Inc). All statistical testing results (p-values) evaluating each of the wetland assessment variables and functions across the restoration chronosequence are provided in Supplemental Table 1.
Results and Discussion
Wetland Assessment Variable Changes Across the Restoration Chronosequence
At each restoration site, the 17 assessment variables were evaluated to complete the HGM wetland functional assessment. Based upon the findings of Berkowitz (2013), assessment variables were previously classified as 1) rapid response variables, 2) response variables requiring additional time to display a measureable effect, and 3) stable variables that remain fixed over time. Those studies documented changes in shrub-sapling density and ground vegetation cover within a few years after restoration occurred, while variables such as tree basal area often require longer time periods to display significant increases (15 or more years; Smith and Klimas 2002; Berkowitz 2013). Conversely, stable assessment variables including flood frequency, wetland tract size, and habitat connectivity often remain constant over time due to landscape factors (i.e., property boundaries, presence of levees, etc). As a result, the following analysis focused on the subset of assessment variables expected to change within the restoration chronosequence project timeframes examined including tree basal area, tree density, woody debris biomass, ground vegetation cover, shrub-sapling density, and the development of soil horizons.
During the 2016 sampling effort, the authors observed substantial increases in tree diameter, initiation of canopy closure, and other factors related to forest succession across the oldest restoration sites (>25 years) compared to site evaluations conducted in previous monitoring events. Evaluating the restoration chronosequence, statistically significant increases in tree basal occurred (Fig. 2a; Supplemental Table 1). The HGM assessment technique evaluates trees with a diameter at breast height (DBH) ≥10 cm. This diameter was first encountered at a subset of 10 year stands, with significant increases documented in 13–20 year old stands, and further improvements at the 25 year post restoration increment following a linear increase in diameter over the restoration chronosequence (Fig. 1a). Tree density data displayed similar results, with significant incremental increases in tree density occurring within the restoration sites after 13 years, followed by further significant increases at 20 and 25 year time intervals (Fig. 1b). Because the HGM approach focuses on trees with DBH ≥10 cm and includes smaller tress in shrub-sapling density measurements, observed increases in tree density likely represent a combination of 1) incorporation of planted trees entering the 10 cm size class and 2) recruitment of naturally regenerated trees into restored areas over time. The observed increases in tree basal area and density support the predictions of Berkowitz (2013), who categorized them as response variables, providing useful measures of restoration success after a period of approximately 15 years post restoration. The increases in both tree diameter and density followed linear trend lines throughout the 25 year chronosequence, which are expected to begin conversion to a more logarithmic pattern as succession continues after approximately 40 years (Allen et al. 2000).

Mean wetland assessment variable measurements observed across the restoration chronosequence. Error bars represent one standard deviation of the mean; entries with different letter designations were significantly different at α = 0.05
Woody debris biomass inputs were expected to increase in response to higher basal area and tree density values, which provide additional sources of woody debris (e.g., loss of branches during storms and self-pruning). Monitoring results indicate significant linear increases in woody debris across the restoration chronosequence (Fig. 1c). Further additions of woody debris biomass are anticipated as stand development continues into the stem exclusion phase of forest succession (Oliver and Larson 1996). Ground vegetation cover follows anticipated patterns following restoration in which coverage increases with the cessation of agricultural activities (e.g., plowing, crop removal), then decreases as transient species are recruited to taller strata and canopy cover approaches closure, reducing the quantity and quality of available light supporting herbaceous plant growth (Fig. 1d; Bigelow et al. 2011). These values also coincide with results reported in Berkowitz (2013), in which ground cover displayed initial increases to approximately 75% after 15 years of restoration, with older (e.g., >20 year) forests exhibiting significantly lower ground cover values near 20%. Shrub-sapling density data displayed similar patterns, with initial increases followed by sharp declines as restoration site succession progressed. Within 25 years, shrub-sapling densities approximated values observed during initial post restoration conditions (Fig. 1e). Both ground vegetation cover and shrub-sapling density were identified as rapid response variables in Berkowitz (2013). The current dataset supports those findings, providing a series of short- to mid-term restoration monitoring milestones.
Soil horizon development increased over the restoration chronosequence, with significant differences in O-horizon thickness detected 25 years post restoration (Fig. 1f). However, soil horizon thickness measurements remained highly variable, with standard deviation ranges encompassing approximately 25% of the average values. The development of thicker O-horizons has been linked with soil development (i.e., age), organic matter inputs, and hydroperiod (Buol et al. 2001). These factors potentially account for the observed degree of variability as the restoration sites display differences in time since soil disturbance (i.e., agricultural plowing activities), vegetation cover and litterfall organic matter sources, and soil inundation which increases organic matter content via decreased soil microbial decomposition efficiency (Reddy and DeLaune 2008).
Examining the entire dataset the wetland assessment variables evaluated generally displayed expected responses to restoration, with assessment metrics following anticipated patterns across the restoration chronosequence. Rapid response variables (e.g., ground vegetation cover, shrub-sapling density) show initial increases followed by anticipated decline, providing short- and mi-term opportunities for monitoring restoration progress. Also, response variables such as tree basal area and density continue to display significant increases across the restoration chronosequence, especially after tree diameter thresholds are exceeded. As a result restoration assessment efforts must focus on the subset of variables with the potential for change detection within project monitoring timeframes, guiding the selection of restoration success metrics during project planning, implementation, and monitoring design. As an example scenario, the sapling shrub density variable data predicts stem densities between 400 and 700 stems/ha after 5 years, followed by subsequent increases (>900) at 10–13 years and declines (<900) in later years (Fig. 2e). This allows for development of specific restoration success milestones that can be monitored over time. However, the values presented herein may need to be adjusted due to regional conditions and practical application of these results may remain limited by mitigation and restoration monitoring periods which commonly occur over 5 to 10 year timeframes (Zedler and Callaway 1999).
Restoration Trajectory Curves Across the Restoration Chronosequence
Smith and Klimas (2002) developed anticipated restoration (i.e., recovery) trajectory curves for a subset of the assessment variables included in the HGM wetland functional assessment (Fig. 3). The estimated trajectories represent generalized approximations based upon expected forest successional conditions, not specific to wetland type or restoration technique. Thus the analysis below compares estimated trajectory curves with monitoring results, providing for evaluation of the theoretical trajectory curves and promoting a discussion of trajectory curve applicability. Additionally, results are compared with data from mature BHW locations in the region. Average tree basal area measurements largely agreed with or exceeded expected results across the 25 year restoration chronosequence. Tree basal areas of 0.53±0.26 m2/ha (average ± standard deviation) were observed in 10 year old stands, outpacing the predicted establishment of trees with a DBH ≥10 cm (Fig. 3a). Only three years later, average basal areas increased to 5.8±0.5 m2/ha, exceeding the predicted value of 3.6 m2/ha. Basal areas continue to increase with stand age, with average values of 12.1±3.1 m2/ha corresponding well with the anticipated basal areas of 15 m2/ha within 25 years of restoration. The rapid increase of tree diameter within three to five year intervals provides additional potential restoration milestone during mid-range post restoration time frames. Notably, the tree basal area values observed at 80 year old reference locations remain below the theoretical values estimated in Smith and Klimas (2002).

Estimated (●) and observed (+) wetland restoration trajectory curves and mature BHW forest conditions (estimated data adapted from Smith and Klimas 2002)
Examining tree density recovery trajectory curves, measurements follow predicted results in the early years after restoration planting (Fig. 3b). For example, 13 year old stands displayed average tree densities of 250±36 stems/ha, which compares well with the predicted value of 240 stems/ha. During natural forest successional patterns tree densities decrease in subsequent years as forests near canopy closure and stem exclusion occurs (Summers 2010; Bigelow et al. 2011) and canopy closure was observed at a subset of older restoration sites during the 2016 data collection effort. However, since the restoration planting scheme planted trees at lower densities than typically observed under nature forest regeneration regimes the degree of stem exclusion and senescence in restoration sites remains unknown and tree density results may vary from the estimated trajectory patterns (Oliver and Larson 1996; Stanturf et al. 2001). For example, the average tree density of 376 ± 36 stems/ha documented at 25 year old restored sites only slightly exceeds the tree densities reported in fully mature reference wetlands in the region (~300 stems/ha; Lockhart et al. 2010) and the measurements of 336 stems/ha collected at mature BHW locations as part of the current study. This data suggests that restoration plantings were conducted at the appropriate density to maximize wetland functions as quickly as possible following restoration activities. However, additional monitoring will be required to assess whether tree densities remain constant as the restored BHW wetland mature, or increase toward the higher values predicted by Smith and Klimas (2002) followed by a decrease back toward reference values.
Woody debris biomass data lag behind predicted values across the restoration chronosequence, with average values of 9±2 m3/ha after 25 years, well below the anticipated 20 m3/ha anticipated (Fig. 3c). Woody debris measurements are in agreement with mature BHW forest values (8.9 m3/ha), however the amount of woody debris is expected to continue increasing as tree growth continues and self-pruning is initiated during further succession (Oliver and Larson 1996). Conversely, the ground vegetation cover data (Fig. 3d) largely agreed with predicted trends, with 13 and 20 year old restoration sites (55±10% and 56±15%, respectively) in alignment with the 58% and 53% groundcovers predicted by Smith and Klimas (2002). Further, the 25 year restoration sites (30±8%) appear to outpace the expected declines in ground vegetation cover (50%) with older restoration sites trending towards the predicted 20% ground vegetation cover and the measured values <5% ground vegetation observed in mature BHW forests as part of the current study.
Shrub-sapling densities also follow the pattern outlined in Smith and Klimas (2002), in which densities increase following restoration, with a subsequent decline (Fig. 3e). However, the field sampling data remain well below the predicted values at all restoration stand age classes. The fully mature BHW forest shrub-sapling densities (1690 stems/ha) generally agree with the estimated value (2000 stems/ha) provided in Smith and Klimas (2002). The development of O-horizon biomass is expected to occur over 40 years post restoration, resulting in O-horizon ≥2 cm thick. Soil data collected 10 years post restoration agree with projected results, with average O-horizon thickness of 0.41 ± 0.1 cm corresponding with the 0.33 cm expected (Fig. 3f). Yet, older restoration sites lagged behind anticipated findings, with O-horizons averaging 0.79±0.2 cm after 25 years, compared to the expected value near 1.0 cm. Despite the lag, a subset of monitoring locations exceed soil horizon thickness projects, with values reaching as high as 2.8 cm which exceeds the 15 year/cm O-horizon development rate reported by Buol et al. (2001). Both the predicted values and the restoration chronosequence measurements remain below the O-horizon thickness values collected at mature BHW forested sites (average 4.8 cm). Additional monitoring will be needed to track progression of soil horizons as restoration progresses in the absence of soil disturbance (e.g., plowing).
In summary, available data suggests that several of the assessment variables meet (e.g., tree basal area) or exceed (e.g., ground vegetation cover) the anticipated milestones based on estimated recovery trajectory curves. While additional research is recommended, this study suggests that restoration assessment metrics are trending toward conditions at mature forested wetlands in the region. For example each of the variables display appropriate patterns (e.g., sapling shrub density), although in some cases, observed values fail to reach the predicted trajectory curve values. The assessment variables are expected to further develop as forest succession continues and trajectory curves appear to provide a useful tool for evaluating restoration progress and predicting future conditions. Additionally, tree density data demonstrate that appropriate stocking rates were implemented during restoration, resulting in tree densities corresponding to mature forests as reported in Lockhart et al. (2010) and measured at mature field sites. This highlights the fact that restoration trajectories may deviate from natural patterns of forested wetland succession and in some cases, restored areas may mimic desired end-state conditions over truncated time periods. However, estimated trajectory curves may require revision as restoration sites continue to mature and more is learned about how assessment variables progress under real world scenarios. Future research should focus on instances in which patterns of forest succession under restored regimes deviate from natural patterns with an emphasis on opportunities to accelerate restored wetland maturation (and associated functional increases) through restoration design, implementation, and post restoration management. While the predicted values provided by Smith and Klimas (2002), Berkowitz (2013) and others remain valuable, a knowledge gap in anticipated restoration outcomes remains with few restoration sites in the region spanning the 25-80 year post restoration period. Further, additional trajectory data examining snag density (few snags occur in the 0-25 year old restoration sites) and other variables can be incorporated into future monitoring efforts as restoration sites enter the stem exclusion phase of forest succession.
Wetland Functional Increases Across the Restoration Chronosequence
Seven wetland functions were evaluated at each restoration site based upon the assessment variables and equations outlined above (Tables 2 and 3). The following examines changes in each function across the restoration chronosequence, provides a discussion of the drivers behind functional increases, and compares results with measured mature BHW wetland functions. A discussion of wetland assessment data interpretation and application is also included. The detain floodwater function displayed statistically significant increases across the restoration chronosequence, most notably within the 20 and 25 year age classes (Fig. 4a; Supplemental Table 1). The increases in functional scores result from improvements in ground vegetation, shrub-sapling density, and tree density assessment variable scores. The functional outcomes are anticipated to increase further as these variables, which provide physical obstructions within the wetland that decrease overland flow velocity, continue to develop over time. Additional improvements in the detain floodwater function are expected to result from the incorporation of the log biomass variable as forest succession progresses, as the absence of downed logs at restoration sites decreases floodwater functional scores by 25% within the current 25 year dataset. Also, a small decrease in functional scores was observed in 10 year old study sites due to a lower flood frequency return intervals at those locations. As discussed above, the interplay between rapid response, response, and static assessment variables dictates the current and future conditions of restoration sites. As a result a 10 year old restoration site with an infrequent flood return interval may display equivalent or lower detain floodwater functions than a 5 year old site that experiences more frequent flood events despite the presence of more aboveground biomass accumulation. The detain floodwater functional outcomes remain below those observed in mature BHW locations, with the oldest restoration sites exhibiting an average 20% deficit compared with measurements obtained at >80 year old wetlands.

Mean wetland functional scores (FCI = Functional Capacity Index) observed across the restoration chronosequence. Error bars represent one standard deviation of the mean; entries with different letter designations were significantly different at α = 0.05
The detain precipitation function showed significant increases after 10 years across the restoration chronosequence (Fig. 4b), followed by small improvements throughout the remainder of the 25 year chronosequence although subsequent increases were not statistically significant. Further improvements in the detain precipitation functions remain limited by the time required for additional organic matter accumulation in surface soil horizons and increases in surface roughness (i.e., microtopography) to occur via tree throw, bioturbation, and other mechanisms (Stolt et al. 2000). The lack of substantial increases in the function after 10 years highlights the need to conduct restoration at locations with appropriate soil and surface characteristics or conduct site enhancement during restoration via organic matter amendments and/or mechanically increasing surface roughness. The detention of precipitation represents a largely physical process chiefly occurring via micro-depressional storage, infiltration and retention by organic material and soils (Smith and Klimas 2002). As a result, the detain precipitation function has the potential to yield functional benefits immediately after restoration occurs, without the necessity for tree maturation to occur as required for several other functions including fish and wildlife habitat maintenance. Notably a subset (13%) of the 606 restoration sample sites examined exhibited functional scores >0.80 functional capacity index, yielding results comparable to conditions observed at mature BHW forests (average of 0.81±0.02 functional capacity index). These high-scoring detain precipitation function sites encompassed the entire range of the restoration chronosequence (i.e., 5-25 years) further demonstrating the capacity of restoration projects to display substantial benefits to physically dominated wetland functions over short time periods.
Previous studies examined the nutrient cycling function across the restoration chronosequence, comparing the HGM assessment results with direct measures of wetland nutrient cycling. Those studies linked measures of soil carbon and nitrogen with nutrient processing mechanisms including soil microbial biomass and potentially mineralizable nitrogen (Berkowitz and White 2013). Results indicated that higher rates of nutrient cycling function (i.e., higher nutrient content and processing capacity) corresponded with increased HGM assessment outcomes, validating the wetland functional assessment approach. Examining the updated chronosequence data, the cycle nutrients function displayed significant increases during both the 20 and 25 year restoration intervals (Fig. 4c). Seven assessment variables related to carbon accumulation and processing (e.g., tree basal area, O-horizon thickness) drive the nutrient cycling functional score. As a result, continued tree growth, soil horizon development, and the generation of additional woody debris will likely result in higher nutrient cycling functional outcomes in the future. The incorporation of snags (currently absent from restored forests; decreasing functional scores by 12.5%) as forest succession continues will further increase nutrient cycling functions toward conditions observed in mature BHW wetlands. The current dataset suggests that a subset (5%) of restored sites are providing nutrient cycling functions at equivalent observed in mature BHW forests clustered in the older restoration age classes (20-25 years post restoration).
The export organic carbon function were similarly validated by linking soil organic carbon concentrations with direct measurements of inundation frequency and duration, thus providing a mechanism for organic carbon export to occur (Berkowitz and White 2013). Incorporating the most current data into the chronosequence yields export organic carbon scores that remain significantly higher in 25 year old restoration sites (Fig. 4d). The export organic carbon function combines flood frequency data with assessment variables that serve as proxy measures of carbon sources (e.g., woody debris biomass) available for export to downstream environments. Note that the flood frequency variable has significant implications for the export organic carbon function, representing a switch effect (or switch index; Smith et al. 2013) with the capacity to either turn the function on/off or weight the impact of other assessment variables on the level of wetland function. If a BHW forest is not subject to flooding then the export of organic carbon to downstream environments cannot occur and the resultant function capacity will remain zero. In contrast if flooding (and potential organic carbon export) does occur, the functional capacity is weighted based upon the frequency of flood events with locations exhibiting flood frequencies ≤2 years having that capacity to achieve the highest possible level of function (i.e., 1.0 functional capacity index). For example some recently (e.g., 5 year) restored locations with flood frequency return intervals ≤2.0 years yielded higher functional scores than lower flood frequency 10 and 13 year old sites with more carbon available for export. With increasing site maturity, additional accumulation of above and below ground carbon stocks is anticipated, resulting in higher export organic carbon functional scores over time. However, few study sites (<1%) currently display levels of function observed in mature BHW locations and substantial improvements in site carbon content will be required for restored sites to approach reference values.
The remove elements and compounds function also utilizes the flood frequency return interval as a multiplicative component (or switch index) for determining functional assessment capacities (Table 3; Fig. 4e). The remaining variables associated with the function (i.e., soil cation exchange capacity, O- and A-horizon thickness) provide proxy measures of a restored wetlands ability to improve water quality through the sequestration of nutrients, heavy metals pesticides and other imported materials from floodwaters. As a result, restored locations with frequent flood return intervals and favorable soil conditions display increased capacity for provide the remove elements and compounds function. This accounts for the high scores observed within the 5 and 10 year restoration intervals and the overall higher functional outcomes compared to most of the other functions examined herein. Notably, a subset of the older restoration site intervals display statistically significant increases in remove elements and compounds functional scores due to improvements in the soil horizon variables despite the presence of estimated flood frequencies >2 years. This demonstrates the impact of site maturation and accumulation O- and A- soil horizon biomass over time despite initial limitations associated with flood frequency conditions. These findings again highlight the need for targeted restoration site selection in order to maximize wetland functions, especially in the short-term following restoration.
The maintain plant communities function (Fig. 4f) exhibited statistically significant increases across the restoration chronosequence, with 30% (>200) of study sites exhibiting functional capacity index values ≥0.80. Steady increases in functional assessment scores resulted from improvements in tree basal area and density and additional functional score increases are expected as additional tree growth occurs. The high initial level of wetland plant community function can be attributed to the species composition variable, which are maximized at restoration sites through selective planting of highly desirable species as outlined in Smith and Klimas (2002). As a result, plant community functions exceeded the values observed in mature BHW forests in the region (0.71 ± 0.02) at >50% of the 606 study sites. These factors result in the highest scores for the seven functions examined in the wetland assessment and emphasize the benefits of utilizing appropriate plant communities during restoration design and implementation. In contrast, the provide fish and wildlife habitat (i.e., habitat) functional scores display initial functional capacity index values of zero for the first 5 years post restoration due to the absence of trees, snags, and logs (Fig. 4g). Scores steadily increase with the onset of tree development, with significant functional increases occurring after 10, 13, and 25 years after restoration. The incorporation of snags and log biomass along with additional tree growth will drive further functional increases as forest succession proceeds, however the lack of wetland functional benefits observed in early years underscores the need to maximize other functions (e.g., remove elements and compounds) during the initial post restoration period.
Interpretation of Wetland Functional Assessment Results
A variety of wetland assessment approaches exist (>60), including methodologies that 1) evaluate wetlands based upon a single output (e.g., function/condition) or average of outputs (Fennessy et al. 2004), 2) analyze multiple outputs as described above (Smith et al. 2013), or 3) examine a suite of outputs broadly encompassing habitat, hydrology, and biogeochemical cycling functions (Noble et al. 2017; Berkowitz et al. 2017). In some cases, resource managers prefer to evaluate restoration success based upon one output or average of multiple outputs. While this approach has advantages, including simplicity and ease of comparison between project alternatives, relying on a single result may limit practitioners’ ability to target specific functions of interest (e.g., habitat for a particular species) or implement strategies to increase one or more desired functional outcome (Wakeley and Smith 2001). For example, wetland functional scores that include tree basal area, and log biomass require substantial time periods prior to showing improvement, while other functions (e.g., remove elements and compounds) respond over much shorter timeframes. Further, some functions may benefit from ongoing restoration site management, such as forest thinning to improve tree growth rates. Conversely, the detain precipitation function (based upon microdepressional ponding and flood frequency) remains difficult to improve through post restoration management activities, emphasizing the importance of site selection during restoration planning and implementation.
Examining the chronosequence data, the average wetland functional capacity across the chronosequence follows anticipated patterns with significantly higher functions detected after 13 and 25 years of restoration (Fig. 4h). The rate of functional capacity improvements also increases across the chronosequence as more mature restoration sites surpass tree diameter and canopy closure thresholds and stores of carbon (e.g., woody debris and O-horizon biomass) accumulate in older restoration sites. As a result, the average functional outputs are anticipated to show additional improvements over time. However as noted in the individual functions discussed above, restored study sites generally lag behind mature BHW forests conditions with few 20 and 25 year old sites achieving the levels of wetland functions observed in reference areas despite a subset of functions approaching or exceeding reference conditions.
The approach of averaging HGM function score has previously generated debate in the scientific literature (Berkowitz et al. 2018). However, during application some resource managers prefer to evaluate restoration success based upon a single value, as opposed to examining changes in each functional score independently. This approach has advantages (simplicity), but also limits a resource manager’s ability to target specific functions of interest (e.g., habitat) or implement strategies to further improve one or more functional outcomes. As previously noted, some functional scores (e.g., habitat) may benefit from restoration site management, while others remain more difficult to improve through forest management activities.
In summary, these findings reiterate the positive effects of 1) sub-canopy and canopy tree development and 2) above and below ground biomass production (e.g., woody debris accumulation, soil horizon development) on functional outcomes. However, restored ecosystems often fail to achieve the level of function observed in their mature un-impacted counterparts, especially in the early years following restoration and with regard to carbon accumulation (Malakoff 1998; Moreno-Mateos et al. 2012). While not unexpected, the delay in initial functional response in young restored wetlands results from a time lag required for BHW forests to progress towards maturity (Gardiner and Oliver 2005). In fact, Smith and Klimas (2002) suggest that BHW forests in the region may require as many as 100 years to reach a steady state. In response, Berkowitz and White (2013) examined wetland functions across the 20 years post restoration chronosequence reporting that functional scores remained an average of 31% below the levels observed at mature control sites. The current study suggests that conditions are improving, with 25 year old restored areas displaying a lower average functional lag of 20%. Future research will be required to continue tracking changes in wetland functions over time and to address the knowledge-gap in anticipated restoration outcomes as restored forested wetlands progress through the 25–100 year post restoration period. Additionally, a region-wide analysis of restored wetland functions is needed to quantify and extrapolate the benefits of large scale wetland restoration (~300,000 ha) projects implemented throughout the Mississippi Alluvial Valley and compare those benefits to the loss of wetland functions resulting from the 70% decline in historical BHW extent.
References
Allen, JA, Keeland, BD, Stanturf, JA, Clewell, AF, Kennedy Jr, HE (2000) A guide to bottomland hardwood restoration. US Forest Service. General Technical Report SRS-40
Bauder ET, Bohonak AJ, Hecht B, Simovich MA, Shaw D, Jenkins DG, Rains M (2009) A draft regional guidebook for applying the Hydrogeomorphic approach to assessing wetland functions of vernal Pool Depressional wetlands in Southern California. San Diego State University, San Diego
Berkowitz JF (2013) Development of restoration trajectory metrics in reforested bottomland hardwood forests applying a rapid assessment approach. Ecological Indicators 34:600–606
Berkowitz JF, White JR (2013) Linking wetland functional rapid assessment models with quantitative hydrological and biogeochemical measurements across a restoration chronosequence. Soil Science Society of America Journal 77:1442–1451
Berkowitz JF, Noble CV, Summers E, White JR, DeLaune RD (2014) Investigation of biogeochemical functional proxies in headwater streams across a range of channel and catchment alterations. Environmental Management 53(3):534–548
Berkowitz JF, Beane N, Philley K, Ferguson M (2017) Operational draft regional guidebook for the rapid assessment of wetlands in the North Slope region of Alaska. ERDC/EL TR-17-14. US Army Engineer Research and Development Center, Vicksburg
Berkowitz JF, Evans DE, Philley KD, Pietroski JP, Ehorn C, Beane N (2018) Restoring bottomland hardwood forests on U.S. Army Corps of Engineers lands: 2016 monitoring report. ERDC/EL TR-18-4
Bigelow SW, North MP, Salk CF (2011) Using light to predict fuel reduction and group select effects in Sierran mixed conifer forest. Canadian Journal of Forest Research 41:2051–2063
Brinson MM (1993) A Hydrogeomorphic approach to wetland functional assessment. Technical Report WRP-DE-4. Waterways Experiment Station, U.S. Army Corps of Engineers, Vicksburg
Brinson MM (1995) The HGM approach explained. National Wetlands Newsletter 17(6):7–13
Buol SW, Southard RJ, Graham RC, McDaniel PA (2001) Soil genesis and classification, 6th edn. Wiley, West Sussex
Clairain EJ (2002) Hydrogeomorphic approach to assessing wetland functions: guidelines for developing regional guidebooks; chapter 1, Introduction and overview of the Hydrogeomorphic approach. ERDC/EL TR-02-3. US Army Engineer Research and Development Center, Vicksburg
Cole CA (2006) HGM and wetland functional assessment: six degrees of separation from the data. Ecological Indicators 6:485–493
Cole CA (2016) HGM: a call for model validation. Wetlands Ecology and Management 24(5):579–585
Dunn OJ (1964) Multiple comparisons using rank sums. Technometrics 6:241–252
Federal Register (1997) National action plan to implement the Hydrogeomorphic approach to assessing wetland function. Federal Register 62(119):33607–33620
Fennessy MS, Jacobs AD, Kentula ME (2004) Review of rapid methods for assessing wetland condition. EPA/620/R-04/009. U.S. Environmental Protection Agency, Washington, DC
Gardiner ES, Oliver JM (2005) Restoration of bottomland hardwood forests in the lower Mississippi Alluvial Valley, USA. In: Stanturf JA, Madsen P (eds) Restoration of boreal and temperate forests. CRC Press, Boca Raton, pp 235–251
Haynes RJ, Moore L (1988) Reestablishment of bottomland hardwoods within national wildlife refuges in the southeast. In: Proceedings of a conference: increasing our wetland resources. Washington, DC: National Wildlife Federation. 95-l03
Haynes RJ, Bridges RJ, Gard SW, Wilkins TW, Cook HR Jr (1995) Bottomland hardwood reestablishment efforts for the US Fish and Wildlife Service: Southeast Region. In: Fischenich JM, Lloyd CM, Palmero MR (eds) Proceedings of the National Wetlands Engineering Workshop. Technical Report-WRP-RE-8. US Army Corps of Engineers, Waterways Experiment Station, Vicksburg
Hefner JM, Brown JD (1995) Wetland trends in the southeastern United States. Wetlands 4:1–11
Hobbs RJ, Cramer VA (2008) Restoration ecology: interventionist approaches for restoring and maintaining ecosystem function in the face of rapid environmental change. The Annual Review of Environment and Resources 33:39–61
Humphrey ML, Lin JP, Kleiss BA, Evans DE (2004) Monitoring wetland functional recovery of bottomland hardwood sites in the Yazoo Basin, MS. Tech Note ERDC/EL TN-04-01. US Army Engineer Research and Development Center, Vicksburg
Jenkins WA, Murray BC, Kramer RA, Faulkner SP (2010) Valuing ecosystem services from wetlands restoration in the Mississippi Alluvial Valley. Ecological Economics 69(5):1051–1061
King SL, Keeland BD (1999) Evaluation of reforestation in the lower Mississippi River alluvial valley. Restoration Ecology 7(4):348–359
King SL, Twedt DJ, Wilson RR (2006) The role of the wetland reserve program in conservation efforts in the Mississippi River Alluvial Valley. Wildlife Society Bulletin 34(4):914–920
Levene H (1960) Robust testes for equality of variances. In: Olkin I (ed) Contributions to probability and statistics. Stanford University Press, Palo Alto, pp 278–292
Lin J (2009) Results of 2009 monitoring/functional assessment of selected USACE reforested bottomland hardwood sites in the Yazoo Basin. US Army Corps of Engineers, Engineer Research and Development Center, Vicksburg
Lockhart BR, Guldin JM, Foti T (2010) Tree species composition and structure in an old bottomland hardwood forest in south-central Arkansas. Castanea 75(3):315–329
Malakoff D (1998) Restored wetlands flunk real-world test. Science 280(5362):371–372
Moreno-Mateos D, Power ME, Comín FA, Yockteng R (2012) Structural and functional loss in restored wetland ecosystems. PLoS Biology 10(1):e1001247
Noble CV, Summers E, Berkowitz JF (2014) Validating the operational draft regional guidebook for the functional assessment of high-gradient ephemeral and intermittent headwater streams. ERDC/EL TR-14-7. US Army Engineer Research and Development Center, Vicksburg
Noble CV, Summers E, Berkowitz JF, Spilker F (2017) Operational draft regional guidebook for the functional assessment of high-gradient headwater streams and low-gradient perennial streams in Appalachia. ERDC/EL TR-17-1
Novitski RP, Smith RD, Fretwell JD (1996) Wetland functions, values, and assessment. In: Fretwell JD, Williams JS, Redman PJ (eds) National water summary on wetland resources, USGS water-supply paper 2425. US Department of the Interior, US Geological Survey, Washington, DC
Oliver CD, Larson BC (1996) Forest stand dynamics. Wiley, New York
Ohio Valley Environmental Coalition, Inc, et al. vs. US Army Corps of Engineers et al (2012) Civil Action number 3:11-0149. United States District Court for the Southern District of West Virginia, HuntingtonDivision, Huntington, WV
Reddy KR, DeLaune RD (2008) Biogeochemistry of wetlands: science and applications. CRC Press, Boca Raton
Rowe D, Parkyn S, Quinn J, Collier K, Hatton C, Joy M, Maxted J, Moore S (2009) A rapid method to score stream reaches based on the overall performance of their Main ecological functions. Environmental Management 43(6):1287–1300
Shapiro SS, Wilk MB (1965) An analysis of variance test for normality (complete samples). Biometrika 52:591–611
Smith RD, Klimas CV (2002) A Regional Guidebook for Applying the Hydrogeomorphic Approach to Assessing Wetland Functions of Selected Regional Wetland Subclasses, Yazoo Basin, Lower Mississippi River Alluvial Valley. ERDC/EL TR-02-4. US Army Engineer Research and Development Center, Vicksburg
Smith RD, Ammann A, Bartoldus C, Brinson MM (1995) An approach for assessing wetland functions using hydrogeomorphic classification, reference wetlands and functional indices. Technical Report TR-WRP-DE-9. Waterways Experiment Station, Army Corps of Engineers, Vicksburg
Smith D, Noble CV, Berkowitz JF (2013) Hydrogeomorphic (HGM) approach to assessing wetland functions: Guidelines for developing guidebooks. ERDC/EL TR-13-11. US Army Engineer Research and Development Center, Vicksburg
Soil Survey Staff (2017) Natural Resources Conservation Service. United States Department of Agriculture. Web Soil Survey. Available online at http://websoilsurvey.nrcs.usda.gov/. Accessed 09/01/16
Stanturf JA, Schweitzer CJ, Gardiner ES (1998) Afforestation of marginal agricultural land in the lower Mississippi River Alluvial Valley, USA. Silva Fennica 32:281–297
Stanturf JA, Gardiner ES, Hamel PB, Devall MS, Leininger TD, Warren ME (2000) Restoring bottomland hardwood ecosystems in the lower Mississippi Alluvial Valley. Journal of Forestry 98(8):10–16
Stanturf JA, Schoenholtz SH, Schweitzer CJ, Shepard JP (2001) Achieving restoration success: myths in bottomland hardwood forests. Restoration Ecology 9(2):189–200
Stolt MH, Genthner MH, Lee Daniels W, Groover VA, Nagle S, Haering KC (2000) Comparison of soil and other environmental conditions in constructed and adjacent palustrine reference wetlands. Wetlands 20(4):671–683
Summers EA (2010) Evaluating Ecological Restoration in Tennessee Hardwood Bottomland Forests. Masters Thesis. University of Tennessee, Knoxville
The Nature Conservancy (1992) Restoration of the Mississippi River Alluvial Plain as a functional ecosystem. Southeastern Louisiana University, Hammond
United Stated Army Corps of Engineers (USACE) (1989) Yazoo backwater area mitigation lands, Lake George property land acquisition, Yazoo Basin, Mississippi: environmental assessment. US Army Corps of Engineers, Vicksburg District, Vicksburg
United States Congress (1985) The Food Security Act of 1985. Public Law. US Congress, Washington, DC, pp 100–233
United States Department of Agriculture, Natural Resources Conservation Service (2016) Field Indicators of Hydric Soils in the United States, Version 8.0. Vasilas LM, Hurt GW, Berkowitz JF (eds) USDA, NRCS, in cooperation with the National Technical Committee for Hydric Soils. https://www.nrcs.usda.gov/Internet/FSE_DOCUMENTS/nrcs142p2_053171.pdf. Accessed 15 Dec 2017
Wakeley JS, Smith RD (2001) Hydrogeomorphic approach to assessing wetland functions: Guidelines for developing regional guidebooks; Chapter 7: Verifying, field testing, and validating assessment models. ERDC/EL TR01–31 Waterways Experiment Station, Army Corps of Engineers, Vicksburg
Walker LR, Wardle DA, Bardgett RD, Clarkson BD (2010) The use of chronosequences in studies of ecological succession and soil development. Journal of Ecology 98(4):725–736
Wigginton JD, Lockaby BG, Trettin CC (2000) Soil organic matter formation and sequestration across a forested floodplain chronosequence. Ecological Engineering 15:S141–S155
Zedler JB, Callaway JC (1999) Tracking wetland restoration: do mitigation sites follow desired trajectories? Restoration Ecology 7(1):69–73
Acknowledgements
Funding support was provided by the USACE Vicksburg District. Thanks to Darrell Evans, Jason Pietroski, Kevin Philley, Nathan Beane, Casey Ehorn and others who endured countless hours of extreme heat, humidity, insects, and head-high Toxicodendron radicans during field data collection. Kenneth Parrish, Gary Young, and Daniel Sumerall provided comments on the draft manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic Supplementary Material
Supplemental Table 1
(DOCX 26 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Berkowitz, J.F. Quantifying Functional Increases Across a Large-Scale Wetland Restoration Chronosequence. Wetlands 39, 559–573 (2019). https://doi.org/10.1007/s13157-018-1103-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-018-1103-9