
Abstract
Two stratigraphic sections are investigated, and a diverse larger benthic foraminiferal assemblage is recorded from the upper Paleocene–lower Eocene Southern Galala Formation at the Galala Plateaus, north Eastern Desert, Egypt. Twenty-eight larger foraminiferal species, belonging to thirteen genera, are identified and their comparative stratigraphic range with the Tethyan zonations of Hottinger (1960) and Serra-Kiel et al. (1998) is documented. Nine shallow benthic zones (SBZ3-6 and SBZ8-12) are designated in the studied interval, involving index zonal markers, e.g., Glomalveolina primaeva (Reichel), G. levis Hottinger, Nummulites atacicus (Leymerie), Alveolina vredenburgi Davies, and A. ellipsoidalis Schwager. Fifty-four thin sections are prepared and analyzed, yielding seven microfacies types in the present study, with larger benthic foraminifera as the dominant biotic components. These microfacies types indicate a deposition in restricted tidal flat inner ramp to open-marine middle ramp environments. The inner ramp environments are dominated by alveolinids, orbitolitids, and in part by miliolids and orthophragminids, reflecting euphotic to mesophotic, meso-oligotrophic, and normal to hyper salinity conditions. The middle ramp environment is characterized by nummulitids, implying meso-oligophotic, oligotrophic, and normal salinity conditions. The studied successions at the Galala Plateaus are devoid of corals in platform stages II and III, similar to the Pyrenean strata from middle latitudes at the northern Tethys, except for small coral patches in both stages in the latter due to the cooler temperature. The Early Eocene (Cuisian) Nummulites accumulations suggest a development on paleohighs in a distal inner ramp environment, and then a transportation by wave and current actions into the surrounding proximal middle ramp environment.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The most prominent long-term warming of the Cenozoic was reported in the Early Paleogene time (66.0–23.0 mya), including a pronounced short-term warming event, i.e., Paleocene-Eocene Thermal Maximum (PETM) occurred around 55.8 million years ago (Zachos et al. 2001; Gilmour et al. 2014). Previous studies from Spain and Egypt (e.g., Orue-Etxebarria et al. 2001; Pujalte et al. 2003; Baceta et al. 2005; Rasser et al. 2005; Scheibner et al. 2005; Scheibner and Speijer 2008) correlated the larger foraminiferal evolution to the PETM environmental changes. Due to the rising temperatures (~ 5–8 °C) during the Early Paleogene, the reef-building corals in the Tethyan domain were declined, and the larger benthic foraminifera, which show greater tolerance to higher temperatures, became expanded (Hottinger 1998; Orue-Etxebarria et al. 2001; Scheibner et al. 2005; Scheibner and Speijer 2008; Martín-Martín et al. 2020).
Larger benthic foraminifera are diverse and widely-distributed in various shallow carbonate platform environments, and are considered as excellent biostratigraphic and paleoenvironmental markers (Beavington-Penney and Racey 2004; Özgen-Erdem et al. 2007; Scheibner et al. 2007; Scheibner and Speijer 2009; Boudagher-Fadel et al. 2015; Sarkar 2019; Hadi et al. 2021). According to the studies of Hottinger (1960) and Schaub (1981) on the Early Paleogene alveolinids and nummulitids, twenty shallow benthic zones (SBZs) were documented in the Neo-Tethyan realm, and were used for high-resolution biostratigraphy in shallow-marine environments (Serra-Kiel et al. 1998, 2020; Beavington-Penney and Racey 2004; Scheibner et al. 2005; Özgen-Erdem et al. 2007; Scheibner and Speijer 2009; Drobne et al. 2011; Hadi et al. 2016; Amirshahkarami and Zebarjadi 2018).
The distribution pattern of larger foraminifera is influenced by a complex set of inter-related parameters, including temperature, light, nutrients availability, substrate, depth, hydrodynamic energy, and salinity (Hottinger 1983; Pecheux 1995; Duxbury and Duxbury 1997; Geel 2000; Scott et al. 2001; Leckie and Olson 2003; Beavington-Penney and Racey 2004; Rasser et al. 2005; Scheibner et al. 2005; Boudagher-Fadel 2008; Bassi et al. 2013; Tomassetti et al. 2016; Eder et al. 2018; Torres-Silva et al. 2019; Martín-Martín et al. 2020; El-Ayyat 2022). Nummulites banks are the most prominent larger foraminiferal accumulations, with a great distribution in the Eocene carbonate deposits along the northern and southern Tethyan margins (Racey 2001; Jorry 2004; Jorry et al. 2006; Martín-Martín et al. 2020, 2021; El-Ayyat 2022). The origin of these banks has been attributed to biological (i.e. reproduction strategies of larger foraminifera) and physical (i.e. winnowing) processes (Aigner 1982; Loucks et al. 1998; Racey 2001; Hasler 2004; Beavington-Penney et al. 2005, 2006; Jorry et al. 2006).
The study area represents the northern part of the Eastern Desert of Egypt, comprising the Northern and Southern Galala Plateaus (Fig. 1). The Paleocene–Eocene rocks in the study area were mostly deposited over a low-angle homoclinal ramp with a gentle, southward dipping (El-Ayyat 2022), represented at the Galala Plateaus by the Southern Galala Formation (Abdallah et al. 1970). The Southern Galala Formation underwent several studies from stratigraphical, paleontological, and paleoenvironmental aspects (e.g., Kuss et al. 2000; Ismail et al. 2007; Morsi and Scheibner 2009; Höntzsch et al. 2011a; El-Ayyat and Obaidalla 2013). Nevertheless, the paleonenvironmental reconstruction of the study area was based only on facies analysis (Scheibner et al. 2001, 2003; Höntzsch et al. 2011b; El-Ayyat and Obaidalla 2013), overlooking the paleoenvironmental and paleoecological significance of the included larger foraminifera. The larger foraminiferal content of the Southern Galala Formation has been previously reported (Aigner 1982; Scheibner et al. 2005; Scheibner and Speijer 2009). However, these studies focused on the Southern Galala Plateau and extended southward, neglecting the Paleocene-Eocene outcrops at the Northern Galala Plateau.

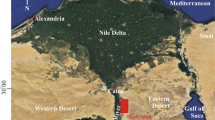
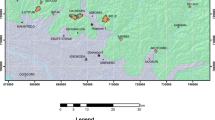
Location map and geological map of the study area, modified after Abd-Elhameed et al. (2020)
Therefore, the main objectives of the present study are; (1) to report the presence of the Thanetian–Cuisian larger benthic foraminiferal assemblages from the Southern Galala Formation at the Northern and Southern Galala Plateaus, (2) to correlate their stratigraphic distributions with the Tethyan biozonations of Hottinger (1960) and Serra-Kiel et al. (1998), (3) and to discuss the paleoenvironment and paleoecology through microfacies analysis of the studied interval, along with the included LBF communities.
Geologic setting and lithostratigraphy
Egypt was situated at the southern margin of the Tethys during the Late Cretaceous-Early Paleogene time, and a south-dipping carbonate platform developed in north Eastern Desert; Galala Plateaus (Scheibner and Speijer 2008; Mahfouz and Metwally 2020). In north Eastern Desert, on the western side of the Gulf of Suez, the Galala Plateaus include the Northern and Southern Galala Plateaus, separated by the 30 km-wide Wadi Araba (Fig. 1). These plateaus represent an isolated Late Cretaceous to Eocene carbonate platform at the southern margin of the Tethys, referred to as the unstable shelf of northern Egypt (Youssef 2003). The evolution of the platform is related to the tectonic activity of the ENE-WSW striking Wadi Araba Fault, which forms part of the Syrian Arc-Fold-Belt (Krenkel 1925; Moustafa and Khalil 1995; Hussein and Abdallah 2001). A major phase of tectonic activity occurred along the Syrian Arc-Fold-Belt during the Early Eocene (Shahar 1994). Regional uplift and subsidence triggered the formation of ENE–WSW striking basins, submarine swells and subaerially exposed plateaus on the unstable shelf (Said 1990; Schütz 1994).
The Early Eocene Galala Plateaus represent one of the southernmost plateaus of the unstable shelf. According to regional tectono-sedimentary constraints, three major depositional units can be distinguished; the Northern Galala/Wadi Araba High (NGWA), a transitional slope zone, and the Southern Galala Subbasin (SGS; Höntzsch et al. 2011b). The NGWA represents subaerially exposed to shallow-marine inner platform environments. Due to the synsedimentary monoclinal uplift and following erosion along the Wadi Araba Fault since the Late Cretaceous, major inner-ramp deposits were eroded or intensively altered (Moustafa and Khalil 1995). The connection between the NGWA and the SGS is represented by a transitional slope zone (mid ramp to outer ramp).
The study area stretches from the southern rim of the Northern Galala Plateau (Askhar El-Bahari section) to the northern rim of the Southern Galala Plateau (Umm Khayshar section), where the upper Paleocene–lower Eocene carbonate platform were subdivided into three biotic stages (I–III) based on the paleoclimatic changes associated with larger foraminiferal evolution (Scheibner et al. 2005).
The biotic stage I is characterized by the occurrence of coralgal reefs during the progradational phase of the carbonate platform, while the biotic stage II shows a change from coralgal reefs to larger foraminiferal shoals at the platform margin during the retrogradational phase of the carbonate platform, as a result of the increase in water temperature. The short-term effects of the Paleocene–Eocene Thermal Maximum (PETM) during the Early Eocene progradational phase (i.e., temperature increase, eutrophication) caused a change from the dominance of miscellanids and ranikothalids to alveolinids and nummulitids, characterizing the biotic stage III (Scheibner et al. 2005).
The upper Paleocene–lower Eocene deposits at the Galala Plateaus are represented by the Southern Galala Formation. It was first introduced by Abdallah et al. (1970) at Gebel Thelmet, on the western side of the Gulf of Suez to describe the Paleocene–Eocene rocks which has been affected by many tectonic activities related to the Syrian arc system. The Southern Galala Formation unconformably overlies the Campanian–Maastrichtian Sudr Formation (Fig. 2; Abdallah et al. 1970; Ismail et al. 2007). The base of the Southern Galala Formation is represented by a 3 m-thick white-yellow sandstone bed at Umm Khayshar area (Fig. 2), and a 3.5 m-thick conglomeratic bed at Askhar El-Bahari area (Fig. 2). Upwards at Umm Khayshar area, the sandstone bed is followed by about 65 m-thick yellowish-white to greyish-white fossiliferous limestones. These limestones are represented by bioclastic floatstone facies, followed by Nummulites grainstone and Nummulites packstone facies (Fig. 2). On the other hand, at Askhar El-Bahari area, the conglomeratic bed is overlain by about 60 m-thick white to greyish-white, fossiliferous limestones, represented by bioclastic rudstone facies, followed by Nummulites-Alveolina wacke-packstone and Nummulites grainstone facies. A 4 m-thick pinkish carbonate breccia (brecciated grainstones) bed caps the succession (Fig. 2).

Lithostratigraphic section of the Paleocene–Eocene Southern Galala Formation at Umm Khayshar and Askhar El-Bahari areas, with the distribution of the recognized microfacies types
Materials and methods
The upper Paleocene–lower Eocene successions are investigated from two sections; Umm Khayshar section at the northern scarp of the Southern Galala Plateau (Lat. 28° 55ʹ 34″ and Long. 32° 10ʹ 25″) and Askhar El-Bahari section at the southern scarp of the Northern Galala Plateau (Lat. 29° 03ʹ 04″ and Long. 32° 01ʹ 09″; Fig. 1). These sections are measured, described, and sampled for microfacies and benthic larger foraminifera analyses.
Fifty-four thin sections of standard size are prepared, analyzed, and photographed using Polarizing Microscope with attached digital camera from Beni-Suef University. Thin-section studies yield different microfacies types, as well as diverse benthic larger foraminiferal content. The recognized microfacies types are described and classified following the scheme of Dunham (1962), with modifications of Embry and Klovan (1972) and Scholle (2003).
Furthermore, twenty-eight larger benthic foraminiferal species belonging to thirteen genera are identified following the taxonomic descriptions of Schwager (1883), Leymerie (1846), Heim (1908), Reichel (1937), Davies (1937), de Cizancourt (1945), Henson (1948), Hottinger (1960, 2014), Drobne (1975), Kuss and Leppig (1989), Wan (1990), and Tosquella and Serra-Kiel (1998). All the materials are housed in the Museum of the Geology Department, Faculty of Science, Helwan University, Egypt.
Results
Biostratigraphy
Twenty-eight larger foraminiferal species are recorded from the studied upper Paleocene (Thanetian)-lower Eocene (Ilerdian-Cuisian) successions, and their stratigraphic distributions are correlated with the Tethyan biozonations of Hottinger (1960) and Serra-Kiel et al. (1998) (Figs. 3 and 4).

The distribution of the identified larger benthic foraminifera from the Paleocene-Eocene Southern Galala Formation at Umm Khayshar area

The distribution of the identified larger benthic foraminifera from the Paleocene-Eocene Southern Galala Formation at Askhar El-Bahari area
The lower part of the Thanetian has been described as the Glomalveolina primaeva zone by Hottinger (1960) or as SBZ3 by Serra-Kiel et al. (1998). This interval is represented by the total range of Glomalveolina primaeva in the two studied sections (Figs. 3 and 4). In addition, Elazigina lenticula and Discocyclina sp. in Umm Khayshar section and G. dachelensis, Periloculina slovenica, and Idalina sp. in Askhar El-Bahari section are associated with G. primaeva in this interval (Fig. 5).

a Glomalveolina primaeva (Reichel 1937), PEF-1. b Glomalveolina levis Hottinger 1960, PEF-2. c Glomalveolina dachelensis (Schwager 1883), PEF-3. d Idalina sp., PEF-5. e Periloculina slovenica Drobne, 1974, PEF-6. f Miscellanea rhomboidea Kuss and Leppig 1989, PEF-7. g Hottingerina lukasi Drobne 1975, PEF-13. h Nummulites catari Tosquella and Serra-Kiel 1998, PEF-16. i Elazigina lenticula (Hottinger 2014), PEF-17. j Discocyclina sp., PEF-18. k Nummulites atacicus (Leymerie 1846), PEF-23. l Nummulites globulus Leymerie 1846, PEF-36. m Orbitolites minimus (Henson, 1950), PEF-55. n Fabularia zitteli Schwager 1883, PEF-56
The upper part of the Thanetian has been described as the Glomalveolina levis zone by Hottinger (1960) or as SBZ4 by Serra-Kiel et al. (1998). It is defined by the total range of Glomalveolina levis in Askhar El-Bahari section and that of Nummulites catari in Umm Khayshar section (Figs. 3 and 4). Furthermore, Elazigina lenticula, Miscellanea rhomboidea, and Discocyclina sp. in Umm Khayshar section and G. dachelensis, Miscellanea rhomboidea, and Hottingerina lukasi in Askhar El-Bahari section are also recognized in this interval (Fig. 5).
The Early Ilerdian has been established as the Alveolina cucumiformis (junior synonym of A. vredenburgi; Hottinger et al. 1998) and A. ellipsoidalis zones by Hottinger (1960) or as SBZ5 and SBZ6, respectively, by Serra-Kiel et al. (1998). This interval is represented by the total range of Alveolina vredenburgi in Umm Khayshar section and that of A. ellipsoidalis in Askhar El-Bahari section (Figs. 3 and 4). Other Alveolina spp., e.g., A. minervensis, A. pasticillata, A. piper, A. rutimeyeri, A. subovata, and Fabularia zitteli are also included in Askhar El-Bahari section. Since the index species of SBZ5 and SBZ6 co-occur in the present study, the boundary between both zones cannot be traced (Figs. 5 and 6).

a Assilina subspinoza Davies, 1937, PEF-57. b Alveolina vredenburgi Davies, 1937, PEF-58. c Alveolina aragonensis Hottinger 1960, PEF-60. d Alveolina ellipsoidalis Schwager 1883, PEF-61. e Alveolina minervensis Hottinger 1960, PEF-62. f Alveolina pasticillata Schwager 1883, PEF-63. g Alveolina piper Hottinger 1960, PEF-66. h Alveolina rutimeyeri Hottinger, 1962, PEF-68. i Alveolina subovata Wan 1990, PEF-69. j Nummulites fossulatus de Cizancourt, 1946, PEF-70. k Nummulites sp., PEF-78. l Assilina major Heim 1908, PEF-107. m Assilina sp., PEF-108. n Lockhartia sp., PEF-113
The Middle Ilerdian has been described as the Alveolina corbarica zone by Hottinger (1960) or as SBZ8 by Serra-Kiel et al. (1998). However, A. corbarica is not recorded in the studied sections, and instead, the total range of Nummulites atacicus defines this interval (Figs. 3 and 4). In addition, Nummulites globulus in Umm Khayshar section and Orbitolites minimus and Nummulites globulus in Askhar El-Bahari section are also recognized in this interval (Figs. 5 and 6).
The Late Ilerdian has been established as the Alveolina trempina zone by Hottinger (1960) or as SBZ9 by Serra-Kiel et al. (1998). Nevertheless, A. trempina is not recognized in the present study. Therefore, the lower boundary of this interval is defined by the last occurrence of Nummulites atacicus and the upper boundary is defined by the last occurrence of Nummulites globulus (Figs. 3 and 4). Alveolina aragonensis is also associated with Nummulites globulus in Umm Khayshar section (Figs. 5 and 6).
The Early Cuisian has been described as the Alveolina oblonga zone by Hottinger (1960) or as SBZ10 by Serra-Kiel et al. (1998). A. oblonga is not reported in the studied sections, and therefore the total range of Nummulites fossulatus defines this interval (Figs. 3 and 4). Assilina subspinoza is also recorded, along with Nummulites fossulatus in Umm Khayshar section (Fig. 6).
The Middle Cuisian has been established as the Alveolina dainellii zone by Hottinger (1960) or as SBZ11 by Serra-Kiel et al. (1998). However, A. dainellii is not recognized in the present study. Therefore, the base of this interval is defined by the last occurrence of Nummulites fossulatus and the top is defined by the first occurrence of Assilina major (Figs. 3 and 4). Only Nummulites sp. is recorded from this interval in the studied sections (Fig. 6).
The Late Cuisian has been described as the Alveolina violae zone by Hottinger (1960) or as SBZ12 by Serra-Kiel et al. (1998). A. violae is not recorded in the studied sections and, therefore, the total range of Assilina major defines this interval (Figs. 3 and 4). Furthermore, Nummulites sp. and Lockhartia sp. are also included in this interval in the two studied sections (Fig. 6).
Microfacies analysis, paleoenvironmental and paleoecological interpretation
The microfacies analysis of the studied upper Paleocene–lower Eocene successions reveals seven microfacies types (MFT 1–7), reflecting various depositional environments (Fig. 9).
Calcareous sandstone (MFT-1)
This microfacies type is recorded from the lowermost part of the Southern Galala Formation in Umm Khayshar section (Fig. 2). It is made up of sub-angular to sub-rounded, fine to medium, sand-sized quartz grains. In addition, few sparry calcite crystals, ferruginous material, iron oxides, glauconite grains, and euhedral, zoned dolomite crystals are also included in this microfacies (Fig. 7a).

a Calcareous sandstone (MFT-1). b, c Sandy bioclastic rudstone (MFT-2), with larger benthic foraminifera (LBF), echinoids (Ec), and bivalves (Bv). d–f Brecciated grainstone (MFT-3), with echinoids, algal plates, and bivalves. f a rock sample of the brecciated grainstone facies. g, h Bioclastic floatstone (MFT-4), showing the dominant bivalve oyster shells and larger foraminifera
The complete absence of faunal components shown in the calcareous sandstone microfacies (MFT-1) and the presence of some glauconite grains reflect a near-shore environment, with restricted circulation conditions (Saber 2012). Based on the dominance of quartz grains and the presence of sparite and dolomite crystals, a deposition in a restricted tidal flat environment, with high water energy, is proposed for this microfacies (Fig. 9).
Sandy bioclastic rudstone (MFT-2)
This microfacies type is recorded from the lower part of the Southern Galala Formation, representing the Late Paleocene interval in Askhar El-Bahari section (Fig. 2). It is characterized by larger benthic foraminifera (e.g. Glomalveolina and Hottingerina), embedded in a groundmass of sparry calcite, with sub-angular-sub-rounded, fine- to medium-grained quartz. Moreover, bivalve shell fragments, gastropod shells, echinoid fragments, crinoid columnals, and halimedacean and dasycladacean algal plates are recorded in this microfacies (Fig. 7b, c).
The sandy bioclastic rudstone microfacies (MFT-2) are characterized by porcelaneous larger foraminifera, e.g. Glomalveolina, Hottingerina, Idalina, and Periloculina. Hottingerina lukasi has been reported from the shallow platform interior (Scheibner and Speijer 2009). Away from oceanic waves, alveolinids inhibit sheltered habitats, i.e., restricted lagoonal settings (Hottinger 1997; Murray 2006; Babazadeh and Alavi 2013; Coletti et al. 2021). The imperforate miliolid foraminifera (e.g. Idalina and Periloculina) also suggest an inner restricted lagoon environment (El-Ayyat 2022). The alveolinids and miliolids have been known as epiphytes, living within vegetated substrates (Brasier 1975; Beavington-Penny et al. 2006; Tomassetti et al. 2016; Martín-Martín et al. 2020; Coletti et al. 2021). They are also reflective of euphotic and moderately oligotrophic conditions (Hottinger 1997; Samankassou 2002; Barattolo et al. 2007; Sarkar 2019; Martín-Martín et al. 2020; El-Ayyat 2022). In addition, the abundance of miliolids suggests normal to hyper salinity conditions lower than 50 psu (Brasier 1975; Murray 2006; Mossadegh et al. 2009; Chan et al. 2017; Sarkar 2019). The presence of green algae characterizes euphotic, oligotrophic shallow-marine settings (Wray 1977; Brandano et al. 2015; Sarkar 2019; Mattern et al. 2021). Moreover, the presence of fine- to medium-grained quartz and sparitic matrix suggests a position close to a source area, with high water energy related to tidal currents or waves (Hadi et al. 2015). Therefore, this microfacies can be attributed to a restricted, high-energy inner ramp lagoon, with euphotic, oligotrophic, and normal to hypersaline waters (Fig. 9). Similar microfacies have been reported from Spain (Martín-Martín et al. 2020) and India (Sarkar 2019), and interpreted a restricted inner ramp lagoon. However, Martín–Martín et al. (2020) also recorded dasyclad and codial green algae, indicating a low-energy marine environment.
Brecciated grainstone (MFT-3)
It is recorded from the uppermost part of the Southern Galala Formation in Askhar El-Bahari section (Fig. 2). The ferruginous calcareous sandstone groundmass of this microfacies consists mainly of fine- to medium-grained, sub-angular to sub-rounded quartz, with few ferruginous material and sparite crystals. Sub-angular clasts of sandy bioclastic grainstone, with fragments of bivalves, echinoids, crinoids, and green algal plates, are embedded in the groundmass (Fig. 7d–f).
The allochthonous components of the brecciated grainstone facies (MFT-3) are represented by skeletal debris, e.g., fragments of bivalves, echinoids, and crinoids, in a sparitic matrix. Such skeletal debris and the sub-angular shape of the clasts suggest high water energy at the flanks of a reef/shoal (Schlagintweit and Gawlick 2008; Pleş et al. 2019). The low diversity of the recorded fauna reveals restricted circulation conditions. Hence, a deposition in a high-energy, restricted, inner ramp (back-shoal) environment is proposed (Fig. 9). Similar microfacies has been reported from Slovenia (Zamagni et al. 2008) and Iran (Bagherpour and Vaziri 2011; Amirshahkarami and Zebarjadi 2018), and interpreted as restricted, back-reef deposits.
Bioclastic floatstone (MFT-4)
It is recorded from the lower part of the Southern Galala Formation, representing the Late Paleocene interval in Umm Khayshar section (Fig. 2). This microfacies consists mainly of well-sorted bivalve oyster shells (Fig. 7g), with foliated structure, along with larger benthic foraminifera, e.g., Nummulites and Discocyclina (Fig. 7h). These skeletal components are embedded in a sparitic groundmass, with few fine- to medium-grained quartz and some concentric ooids.
The bioclastic floatstone microfacies (MFT-4) is characterized by abundant oyster fragments, larger foraminiferal tests, fine- to medium-grained quartz, and concentric ooids, indicating high-energy conditions above fair-weather wave base (FWWB), likely related to a shoal environment at a shelf-edge or bank-margin facies (Chablais et al. 2010; Sarkar 2019; Hadi et al. 2021). The co-occurrence of alveolinids (e.g. Glomalveolina) and nummulitids (e.g., Nummulites) suggests euphotic to mesophotic, meso-oligotrophic, open-marine, proximal to distal inner ramp environment, with normal salinity (32–38 psu) and high water energy, close to the FWWB (Geel 2000; Mossadegh et al. 2009; Hadi et al. 2016, 2021; Sarkar 2019). The orthophragminids (e.g., Discocyclina) have been reported from a wide array of environments at the lower limit of the photic zone, from inner ramp back- and fore-reef/shoal environments (Ghose 1977; Babazadeh and Alavi 2013; Hadi et al. 2015; Coletti et al. 2021) to deeper, middle-outer ramp environments (Eichenseer and Luterbacher 1992; Gilham and Bristow 1998; Beavington-Penney and Racey 2004; Babazadeh and Alavi 2013), with meso-oligotrophic and normal salinity conditions (Anketell and Mriheel 2000; Hadi et al. 2019; Martín–Martín et al. 2020). The orthophragminids recorded in this microfacies are represented by ovate forms, characterizing the inner ramp settings (Loucks et al. 1998; Zamagani et al. 2008). Discocyclina, Nummulites, and rotaliids (e.g., Elazigina) show preference for soft sandy and muddy substrates (Scheibner et al. 2005, 2007; Zamagani et al. 2008). Consequently, MFT-4 is attributed to a high-energy, distal inner ramp shoal environment, in agreement with their stratigraphic association with core- and back-bank deposits, with euphotic to mesophotic, meso-oligotrophic, and normal salinity conditions (Fig. 9). Although this microfacies show some features similar to that reported from Spain, France, and Slovenia (Rasser et al. 2005; Scheibner et al. 2007; Zamagani et al. 2008) with the predominance of larger benthic foraminifera, the corals and red algae are completely absent.
Nummulites-Alveolina wacke-packstone (MFT-5)
This microfacies type is recorded from the middle part of the Southern Galala Formation in Askhar El-Bahari section (Fig. 2). It is made up of micritic groundmass, encompassing various skeletal components. These components are dominated by larger benthic foraminifera, e.g., A-form Nummulites, Alveolina, and Orbitolites, with some bivalve shells and green algal plates (Fig. 8a, b).

a, b Nummulites-Alveolina wacke-packstone (MFT-5), with larger benthic foraminifera and algae. c, d Nummulites grainstone (MFT-6), showing the larger benthic foraminifera and echinoids. e, f Nummulites packstone (MFT-7), showing the larger benthic and planktonic foraminifera
The Nummulites-Alveolina wacke-packstone microfacies (MFT-5) are dominated by nummulitids, alveolinids, and orbitolitids. The alveolinids (e.g., Alveolina and Fabularia spp.) show a great tolerance to salinity and temperature fluctuations, and therefore, could inhibit any part within shallow-marine carbonate platforms (Drobne et al. 2011; Hadi et al. 2015, 2018; Sarkar 2019). They are also indicative of euphotic, meso-oligotrophic conditions (Tomassetti et al. 2016; Coletti et al. 2021). The wide distribution of alveolinids, orbitolitids (e.g., Orbitolites), and green algae, in a wacke-packstone matrix indicates euphotic, oligotrophic inner ramp environment, with moderate-high water energy (Wray 1977; Hottinger 1997, 1983; Scheibner et al. 2007; Babazadeh and Alavi 2013; Hadi et al. 2015; Coletti et al. 2021). Imperforate porcelaneous foraminifera reflect higher salinity conditions (< 50 psu; Mossadegh et al. 2009; El-Ayyat 2022). Hottinger (1977) and Hadi et al. (2016) interpreted the alveolinid-dominated carbonate successions as of back-bank origin. The co-occurrence of alveolinids and A-form Nummulites suggests euphotic to mesophotic, meso-oligotrophic, proximal to distal inner ramp settings, with high-energy conditions (Geel 2000; Beavington-Penney et al. 2006; Zamagani et al. 2008; Sarkar 2019; Hadi et al. 2021). It can be concluded that this microfacies has been deposited in a distal inner ramp (back-bank) environment, with moderate-high water energy, meso-euphotic, oligotrophic, and normal to hyper salinity conditions (Fig. 9). This microfacies type shows great similarity to that of Hadi et al. (2016), with high abundance of alveolinids, A-forms, and other porcelaneous foraminifera, that was indicator of the inner part of ramp with high-energy conditions.

Paleoenvironmental model for the Paleocene–Lower Eocene successions in the study area, with the distribution of the recognized microfacies types and the recorded larger benthic foraminiferal groups. FWWB Fair weather wave base
Nummulites grainstone (MFT-6)
This microfacies type is recorded from the upper part of the Southern Galala Formation in the two studied sections, representing the Early Eocene (Cuisian) interval (Fig. 2). It encompasses highly diversified biota in a sparitic groundmass, with some quartz grains and calcareous ooids. Larger benthic foraminifera, mainly Nummulites and Assilina spp., bivalve shells, gastropod shells, and echinoid fragments are the main skeletal components of this microfacies (Fig. 8c, d).
The Nummulites grainstone microfacies (MFT-6) contains abundant larger foraminifera (e.g. Nummulites and Assilina spp.) and echinoids, reflecting open-marine conditions (Racey 2001; Saber 2012; El-Ayyat 2022). The appearance of Assilina and Nummulites is indicative of an increase in water depth (Hadi et al. 2015). The presence of A-form and rare B-form nummulitids, along with the disappearance of the porcelaneous foraminifera, e.g., Alveolina, in a grainstone matrix reflects oligotrophic proximal to distal inner ramp palaeohighs, influenced by winnowing and current processes (Langer and Hottinger 2000; Zamagani et al. 2008; Hadi et al. 2021). The presence of calcareous ooids and fine- to medium-grained quartz suggests a higher water energy event, possibly related to tidal currents or waves (Hadi et al. 2015). The presence of nummulitids (e.g., Nummulites and Assilina) indicates preference for a soft substrate, and reflects meso-oligophotic and normal salinity conditions (32–38 psu; Hottinger 1997; Scheibner et al. 2005, 2007; Martín–Martín et al. 2020). The minor association of Nummulites with micro- and macrofauna confirms the deposition in nutrient-poor waters (Hallock 1985). Therefore, this microfacies reflect the development of Nummulites accumulations (banks) in a distal inner ramp environment, which were then transported into the proximal middle ramp environment, with considerable wave and current actions, meso-oligophotic, oligotrophic, and normal salinity conditions (Fig. 9). Similar microfacies type has been reported from Iran (Hadi et al. 2016), where the accumulations of A-form Nummulites-dominated with robust and ovoid tests are interpreted as palaeohighs developed in shallow inner-ramp setting.
Nummulites packstone (MFT-7)
It is recorded from the middle part of the Southern Galala Formation in Umm Khayshar section (Fig. 2). This microfacies consist of highly diversified faunal content, embedded in a groundmass of microcrystalline calcite. The faunal content is dominated by larger benthic foraminifera, e.g., Nummulites and Alveolina, planktonic foraminifera, bivalve shells, echinoid and crinoid fragments (Fig. 8e, f).
The Nummulites packstone (MFT-7) is dominated by Nummulites, co-occurring with rare larger porcelaneous foraminifera, e.g., Alveolina, suggesting downslope transport and deposition in distal inner ramp to proximal middle ramp settings, below the fair-weather wave base (Hadi et al. 2015, 2021). The B-form Nummulites of this microfacies can be indicative of deeper waters (Ćosović et al. 2004; Machaniec et al. 2011; Sarkar 2019) in a fore-bank environment (El-Ayyat 2022). These nummulitids indicate soft substrate, normal water salinity (32–38 psu), oligotrophic, and meso-oligophotic conditions (Scheibner et al. 2005; Mossadegh et al. 2009; Martín–Martín et al. 2020). Furthermore, the wide distribution of planktonic foraminifera reflects open-marine, middle to outer ramp settings, with oligotrophic settings and salinity between 32 and 40 psu (Beavington-Penney et al. 2004; Murray 2006). It can be concluded that this microfacies originated in a middle ramp (fore-bank) environment, with oligophotic, oligotrophic, and normal salinity conditions (Fig. 9). Martín–Martín et al. (2020) reported similar microfacies type from Spain with abundant lenticular nummulitids in packstone–wackestone beds. Nevertheless, the mixing of flattened tests of Assilina and operculiniform Assilina reflected an intermediate depth into the middle ramp environment.
Discussion
Paleocene–Eocene boundary
The Paleocene–Eocene boundary was delineated officially by the onset of the Carbon Isotopic Excursion (CIE; Aubry and Ouda 2003), corresponding to the base of P5b and NP9b zones (Dupuis et al. 2003; Scheibner and Speijer 2009; Zhang et al. 2013). On the other hand, there are two contrasting opinions concerning the position of P-E boundary with respect to the shallow benthic zones. Serra-Kiel et al. (1998) and Hottinger (1998, 2001) suggested that the P–E boundary spanned the time intervals from the lower part of SBZ6 to the middle part of SBZ7.
On the contrary, Scheibner et al. (205) and Scheibner and Speijer (2009) placed the P–E boundary at the transition between SBZ4 and SBZ5 in Egypt, coinciding with the base of the Ilerdian, with the index species for SBZ4 below the onset of the CIE and SBZ5 index species above it. This proposition is also consistent with data from the Pyrenean basin (Orue-Etxebarria et al. 2001; Pujalte et al. 2009a, b a, b; Serra-Kiel et al. 2020).
In the studied sections, the Paleocene–Eocene boundary is marked by the last occurrence of the index species for SBZ4, e.g., Glomalveolina levis and Hottingerina lukasi, and the first occurrence of the index species for SBZ5/6, e.g. Alveolina vredenburgi and Alveolina ellipsoidalis (Figs. 3 and 4).
Factors controlling the LBF distribution
Larger benthic foraminifera are the most prevailing faunal components of the recognized microfacies types. According to Scott et al. (2001), LBF are highly influenced by even subtle variations in environmental conditions. Therefore, they are considered as excellent markers for paleoenvironmental parameters, including temperature, light, nutrients availability, substrate, depth, hydrodynamic energy, and salinity (Hottinger 1977, 1983; Pecheux 1995; Duxbury and Duxbury 1997; Geel 2000; Scott et al. 2001; Leckie and Olson 2003; Beavington-Penney and Racey 2004; Rasser et al. 2005; Boudagher-Fadel 2008; Drobne et al. 2011; Tomassetti et al. 2016; Eder et al. 2018; Torres-Silva et al. 2019; El-Ayyat 2022). The most influential parameters, in shallow shelves, are nutrients sources and light intensity, where primary production is most pronounced (Duxbury and Duxbury 1997).
Light intensity
The light intensity relies on water depth and water transparency, i.e., light intensity reduces and water transparency increases as water depth increases (Hallock, 1987). According to light dependence, there are three groups of organisms; light-dependent organisms (e.g., corals, algae, and porcelaneous foraminifers), low light-dependent organisms (e.g., algae, Operculines, and Lepidocyclines), and light-independent organisms (e.g., bryozoa, mollusca, crinoids, and sponges). Different larger benthic foraminiferal taxa are good indicators to reconstruct the paleo-depth conditions of seawater. Porcelaneous forms, for example, are light dependent and abundant in shallow-marine settings (inner ramp environment; Fig. 9), while they are rare in deeper settings (Martín-Martín et al. 2020). On the other hand, the quantity of hyaline foraminifera (e.g. nummulitids) is mostly enhanced with increasing the water depth.
Lenticular shells of Nummulites are diverse in shallow-marine settings, inhabiting the basal portion of the top of the photic zone (e.g., Nummulites bank), but thin-shelled tests considerably occur in deep waters (fore-bank environment; Beavington-Penney et al. 2004). Such variation in shell shapes is indicative of decreasing light grades at deeper settings (BouDagher-Fadel 2008). The green calcareous algae recorded in the shallow-marine sediments within the Southern Galala Formation (Figs. 7 and 8) are typical of euphotic zone, with ultimate depth of prolific growth commonly about 20–30 m depth. Furthermore, planktonic foraminifera are recorded in a middle ramp environment in open-marine settings (Fig. 9), reflecting oligophotic conditions (BouDagher-Fadel 2008). It can be concluded that the decrease in light intensity affected the abundance and dominance of porcelaneous forms (e.g., alveolinids) and replaced them with hyaline forms (e.g., nummulitids).
Temperature
The impact of temperature on benthic fauna is illustrated through affecting the metabolic performance and dissolution of CO2, i.e., a decrease in temperature leads to a decrease in the metabolic performance and therefore the nutrient requirements of the organism (Geel 2000). In general, most of the LBF inhibit subtropical to tropical environments, at temperatures of about 18–23 °C (Lee 1990; Ungaro 1994; Hottinger 1997; Rasser et al. 2005; Wilson and Vecsei 2005; Brandano et al. 2009; El-Ayyat 2022), while their absence points to cooler or more nutrient-rich environments (El-Ayyat 2022). However, some extant larger foraminifera can tolerate water temperatures as low as 10–11 °C, e.g., Amphistegines, Sorites, Amphisorus, and Amphistegines (Li et al. 1999). The co-occurrence of larger benthic foraminifera and green calcareous algae, to some extent, would reside in warm waters having temperatures of about 18–20 (Brandano et al. 2009), proposing a tropical–subtropical climate for the studied succession.
Salinity
Salinity has a direct influence on the evolution of larger benthic foraminifera. The existence of porcelaneous foraminifera (e.g. alveolinids and miliolids) and the lack of hyaline foraminifera, red algae, and corals mark the dominance of higher levels of hypersaline water (higher than 50 psu) in enclosed lagoon setting (Mossadegh et al. 2009). The occurrence of larger benthic foraminifera with perforated wall (such as Nummulites) refers to normal water salinity conditions, with salinity ranges from 32 to 38 psu (Mossadegh et al. 2009). Planktonic foraminifera live in open oceans and also in inland seas (e.g., the Mediterranean). Their tolerance to variations in water salinity is very limited; indeed, most extant members are stenohaline. No extant planktonic foraminifera have been recorded from waters with salinity higher than 40 psu or lower than 32 psu (Murray 2006). It can be concluded that that the upper parts of the investigated successions accumulated in normal marine salinity circumstances as indicated by the high abundance of hyaline forms (e.g., nummulitids), while the lower parts witnessed interrupted normal and higher water salinity conditions according to the dominance of porcelaneous forms (e.g., alveolinids).
Nutrients
Larger benthic foraminifera live in stabilized, nutrient deficient, oligotrophic circumstances. On the other hand, LBF cannot react competitively in case of plenty nourishing supplies (Hallock 1985). The imperforate foraminifera (e.g., miliolids), which inhibit inner confined lagoon (Fig. 9), reflect low grades of nutrients (Samankassou 2002). In addition, the presence of green algae confirms the oligotrophic shallow-marine settings (Wray 1977). The presence of nummulitids (e.g., Nummulites and Assilina) indicates oligophotic conditions (Martín–Martín et al. 2020), while their association with alveolinids suggests meso-oligotrophic circumstances (Hadi et al. 2021). Concerning the recorded Nummulites bank, no associated micro- or macrofauna has been reported, reflecting a deposition in a nutrient-poor (oligotrophic) environment and/or in a setting with significant hydrodynamic sorting. In general, sediments rich in larger benthic foraminifera are often inferred to represent a deposition in oligotrophic settings, because such sediments are overwhelmingly accompanied with organisms tolerant of nutrient-poor conditions (e.g., green calcareous algae). The existence of open-marine planktonic foraminifera indicates surface trophic to intermediate mesotrophic water conditions (BouDagher-Fadel 2008).
Substrate
The different shapes of Nummulites resulted from the reproductive strategies and environmental conditions, such as substrate and water hydrodynamic energies. These factors affect the thickness, size and shape of these shells (Ćosović et al. 2004). Brasier (1975) mentioned that thick-shelled and conical forms of larger benthic foraminifera prefer housing coarse-grained sediments, but flattened shell forms prefer soft substrates. The alveolinids and miliolids have been known as epiphytes, living within vegetated substrates (Beavington-Penny et al. 2006; Tomassetti et al. 2016; Martín-Martín et al. 2020; Coletti et al. 2021). On the other hand, nummulitids (e.g., Nummulites and Assilina), rotaliids (e.g., Elazigina), and Discocyclina show preference for soft sandy and muddy substrates (Scheibner et al. 2005, 2007; Zamagani et al. 2008). Therefore, the substrate is an important factor affecting the abundance and dominance of different larger foraminiferal groups.
Water depth and hydrodynamic behavior
Changes in water depth could affect environmental changes such as sedimentation rate, turbulence, light intensity, organic change, water hydrodynamic energy, and dissolved oxygen (Leckie and Olson 2003). Due to their sensitivity to water depth, LBF were used in the early times to estimate water depth (El-Ayyat 2022). Alveolinids avoid very shallow water and live in sheltered habitats, away from oceanic wave energies, and thus are particularly common in deeper parts of restricted lagoonal settings (Murray 2006). Some LBF such as Nummulites and Discocyclina become flatter, with thinner outer walls, with increasing water depth (middle ramp Nummulites bank; Fig. 9) and decreasing light intensity. This reflects the dependence of morphology on the gradient of depth (Pecheux 1995).
Perforate foraminifera (e.g., Nummulites and Assilina) are abundant in subtropical to tropical environments, reaching a depth down to 40–70 m (Hottinger 1997). At the lower part of the studied successions, miliolids are plentiful and mostly found in a shallow and restricted water depth. In addition, green calcareous algae show subtropical to tropical environments, with a depth of 20–80 m (Flügel 2010). On the contrary, within the middle and upper parts, Nummulites, and Assilina (as perforate foraminifera) are common and reflect open marine conditions (Fig. 9). The occurrence of planktonic foraminifera and the deficiency of benthic fauna at the upper part of the successions suggest a deposition in tranquil deeper water (Murray 2006). Accordingly, increase in water depth affected the abundance of porcelaneous forms (e.g., alveolinids and miliolids), and led to the dominance of hyaline forms (e.g., nummulitids).
The hydrodynamic behavior of LBF, which depends on density, size and their shapes, is an essential factor controlling their transport (BouDagher-Fadel 2008). Some foraminifera develop thick lamellar crust shells and others tend to develop a hard, conical shell shape with abundant secondary cells, depending on hydrodynamic water energy (Rasser et al. 2005). The wide distribution of alveolinids and orbitolitids (e.g., Orbitolites) in the inner ramp back-bank setting indicates moderate water energy conditions (Coletti et al. 2021). On the other hand, the co-occurrence of alveolinids and nummulitids in the shallow-marine inner ramp settings suggests high water energy conditions, close to the FWWB (Sarkar 2019; Hadi et al. 2021).
Upon comparison of the Pyrenean strata from middle latitudes at the northern Tethys (Scheibner et al. 2007) with the Egyptian succession from the low latitudes at the southern Tethys (this study), the overall trend within the three platform stages proposed by Scheibner et al. (2005) is the same, while latitudinal differences exist, especially in platform stages II and III. The studied successions at the Galala Plateaus are devoid of corals in both platform stages II and III. This overall trend of a less frequent coral occurrence is also apparent in the Pyrenean succession, except for the small coral patches in the late Paleocene platform stage II and the early Eocene platform stage III (Baceta et al. 2005).
The reasons for these platform evolution stages were probably the same in both regions. However, the consequences were less severe in the middle latitudes of the Pyrenees, representing a latitudinal effect. In the low latitudes (this study), temperatures were too warm to sustain the growth of coral reefs and enabled the larger foraminifera to occupy this niche and flourish during the early Paleogene. On the other hand, in the middle latitudes (Pyrenees), the temperatures were cooler than in the tropics, so that coral reefs still could grow, but played only a minor role comparing with the dominant larger foraminifera (Scheibner et al. 2007).
Late Paleocene–Early Eocene paleoenvironmental evolution
The Early Palaeogene evolution of the Galala carbonate platform is strongly influenced and controlled by syndepositional tectonic activity along the Wadi Araba Fault (Höntzsch et al. 2011a). From the PETM to the latest Early Eocene, three tectono-sedimentary platform stages were classified and discussed by Scheibner et al. (2003) and Höntzsch et al. (2011b). The termination of the tectonic activity of the Wadi Araba Fault, related to the Syrian Arc System, is reflected by the absence of siliciclastics (Fig. 2). The microfacies analysis along the Late Paleocene–Early Eocene inner to middle ramp transect yields detailed information regarding environmental and biotic conditions of the platform system.
The shallow-marine inner ramp environment is dominated by photo-autotrophic larger benthic foraminiferal groups (e.g., alveolinids and nummulitids), smaller benthic foraminifera (e.g., miliolids), and halimedacean and dasycladacean green algae (Figs. 7–9). Typical low-latitude nutrient-depleted assemblages, such as corals, are absent. Other faunal components include bivalves and gastropods. The environmental impact of the PETM caused a major shift in the prevailing larger benthic foraminiferal taxa, where new taxa with different ecological strategies evolved (Orue-Etxebarria et al. 2001; Scheibner et al. 2005). Paleocene orthophragminids and miliolids were replaced by the first Eocene nummulitids. Porcelaneous alveolinids evolved from planspiral glomalveolinids to fusiform Eocene alveolinids (Hottinger 2001). Such major shift in the larger benthic foraminiferal assemblages is recorded in the middle ramp environment (Figs. 7–9). The diversification of LBF was linked to an overall increase in test size. The coeval evolving adult dimorphism of LBF is interpreted as an adaptation to the seasonality in warm and oligotrophic habitats (Hottinger 1998, 2001). The progressive increase in shell size and development of adult dimorphism are typical features of long-term maturation cycles within larger benthic foraminifera communities (Hottinger 1998; Zamagani et al. 2012). This Larger Foraminifera Turnover (LFT) was directly linked to the CIE of the PETM (Scheibner et al. 2005; Höntzsch et al. 2011a). The present study is in accordance with the studies of Scheibner et al. (2005), Scheibner and Speijer (2009), and Höntzsch et al. (2011b), except that the larger benthic foraminiferal assemblages shifted from the Paleocene ranikothalids, miscellanids, and glomalveolinids to the Eocene nummulitids and alveolinids.
Early Eocene Nummulites accumulations
Eocene Nummulites accumulations (banks) have been widely distributed along both margins of the Tethys, the northern margin (from Pyrenees to West Pacific) and the southern margin (from North Africa to Oman) (Racey 2001; Jorry 2004; Jorry et al. 2006; Martín-Martín et al. 2020, 2021; El-Ayyat 2022).
Concerning the origin of these accumulations, different processes have been proposed; biological (i.e., larger foraminiferal reproduction strategies) and physical (i.e., winnowing and sorting) processes (Aigner 1982; Loucks et al. 1998; Racey 2001; Hasler 2004; Beavington-Penney et al. 2005, 2006; Jorry et al. 2006).
The Early Eocene carbonate ramp of the Southern Galala Formation in the study area is dominated by larger benthic foraminifera (e.g., nummulitids). The Nummulites accumulations recorded in the present study have been developed on paleohighs in a distal inner ramp environment, and then transported into the surrounding proximal middle ramp environment with significant wave and current actions. This is comparable with the Eocene El-Garia Formation in Tunisia and the Ziarat Formation in Iran which were interpreted as nummulitic paleohighs in shallow-marine environments and transported into the surrounding deeper settings (Beavington-Penney et al. 2005; Hadi et al. 2021).
Conclusions
The upper Paleocene–lower Eocene Southern Galala Formation is exposed at the Northern and Southern Galala Plateaus, north Eastern Desert, Egypt, consisting mainly of fossiliferous limestones, with abundant larger benthic foraminiferal fauna. Twenty-eight larger foraminiferal species of thirteen genera are reported in the present study. In addition, their stratigraphic ranges are correlated with the Tethyan zonations of Hottinger (1960) and Serra-Kiel et al. (1998), designating nine shallow benthic zones (SBZ3-6 and SBZ8-12), involving index zonal markers, e.g., Glomalveolina primaeva, G. levis, Nummulites atacicus, Alveolina vredenburgi, and A. ellipsoidalis.
Based on the microfacies analysis of the studied interval and their fossil components, seven microfacies types are recognized, including calcareous sandstone, sandy bioclastic rudstone, brecciated grainstone, bioclastic floatstone, Nummulites-Alveolina wacke-packstone, Nummulites grainstone, and Nummulites packstone, mostly dominated by larger benthic foraminifera. The distribution of the recorded microfacies types and the paleoenvironmental and paleoecological interpretation of the dominant larger benthic foraminifera reflect depositional environments ranging from restricted inner ramp to open-marine middle ramp environments.
The larger foraminifera in the inner ramp environments are represented by alveolinids, orbitolitids, and in part by miliolids and orthophragminids, suggesting euphotic–mesophotic, meso-oligotrophic, and normal to hyper salinity conditions. Meanwhile, the middle ramp environment is dominated by nummulitids, reflecting meso-oligophotic, oligotrophic, and normal salinity conditions.
The Early Eocene (Cuisian) Nummulites accumulations that have been widely distributed along the northern and southern margins of the Tethys, reflect a development on paleohighs in a distal inner ramp environment and subsequent transportation by wave and current actions into the proximal middle ramp environment.
References
Abdallah AM, Sharkawi MA, Marzouk M (1970) The Campanian rocks of the geology of Mersa Thelmet area, Southern Galala, Plateau, A.R.E. Bull Fac Sci Cairo Univ 44:271–280
Abd-Elhameed S, Mahmoud A, El-Kazzaz Y, Salama Y (2020) Carboniferous brachiopod communities from Wadi Araba, western side of the Gulf of Suez, Egypt: paleontology, paleoecology and paleobiogeography. J African Earth Sci 162:1–28
Aigner T (1982) Event stratification in Nummulite accumulations and in shell beds from the Eocene of Egypt. In: Einsele G, Seilacher A (eds) Cyclic and Event Stratification. Springer-Verlag, Berlin, pp 248–262
Amirshahkarami M, Zebarjadi E (2018) Late Paleocene to Early Eocene larger benthic foraminifera biozones and microfacies in Estahbanate area, Southwest of Iran with Thetyan biozones correlation. Carbonates Evaporites 33(4):869–884
Anketell J, Mriheel I (2000) Depositional environment and diagenesis of the Eocene Jdeir Formation, Gabes-Tripoli Basin, Western Offshore. Libya J Petrol Geol 23:425–447
Aubry MP, Ouda K (2003) Introduction to the upper Paleocene-Lower Eocene of the Upper Nile Valley: Part I Stratigraphy. Micropaleontology 49(Suppl. 1):ii–iv
Babazadeh SA, Alavi M (2013) Paleonvironmental model for Early Eocene larger benthic foraminiferal deposits from south Birjand region, east Iran. Revnue De Paleobiologie, Geneve 32(1):223–233
Baceta JI, Pujalte V, Bernaola G (2005) Paleocene coralgal reefs of the western Pyrenean basin, northern Spain: new evidence supporting an earliest Paleogene recovery of reefal systems. Palaegeography, Palaeoclimatology, Palaeoecology 224:117–143
Bagherpour B, Vaziri MR (2011) Facies, paleoenvironment, carbonate platform and facies changes across Paleocene Eocene of the Taleh Zang Formation in the Zagros basin. SW-Iran Hist Biol 24(2):121–142
Barattolo F, Bassi D, Romano R (2007) Upper Eocene larger foraminiferal-coralline algal facies from the klokova mountain (southern continental Greece). Facies 53(3):361–375
Bassi D, Hottinger L, Nebelsick H (2007) Larger foraminifera from the Upper Oligocene of the Venetian area, northeast Italy. Palaeontology 50:845–868
Beavington-Penney SJ, Racey A (2004) Ecology of extant nummulitids and other larger benthic foraminifera: applications in palaeoenvironmental analysis. Earth Sci Rev 67:219–265
Beavington-Penney SJ, Wright VP, Racey A (2005) Sediment production and dispersal on foraminifera-dominated early Tertiary ramps: the Eocene El Garia formation. Tunisia Sedimentology 52(3):537–569
Beavington-Penney SJ, Wright VP, Racey A (2006) The Middle Eocene Seeb Formation of Oman: An investigation of a cyclicity, stratigraphic completeness, and accumulation rates in shallow marine carbonate settings. J Sediment Res 76(10):1137–1161
Boudagher-Fadel MK (2008) Evolution and geological significance of larger benthic foraminifera. In: Wignall P (ed) Developments in paleontology and stratigraphy. Elsevier, Amsterdam, p 540
Boudagher-Fadel MK, Price GD, Hu XM, Li J (2015) Late cretaceous to early Paleogene foraminiferal biozones in the Tibetan Himalayas, and a pan-Tethyan foraminiferal correlation scheme. Stratigraphy 12(1):67–91
Brandano M, Frezza V, Tomassetti L, Pedley M, Matteucci R (2009) Facies analysis and palaeoenvironmental interpretation of the late Oligocene Attard Member (lower Coralline Limestone Formation), Malta. Sedimentology 56:1138–1158
Brandano M, Tomassetti L, Frezza V (2015) Halimeda dominance in the coastal wedge of Pietra di Finale (Ligurian Alps, Italy): The role of trophic conditions. Sediment Geol 320:30–37
Brasier MD (1975) The ecology and distribution of Recent foraminifera from the reefs and shoals around Barbuda, West Indies. J Foramin Res 5:193–210
Chablais J, Onoue T, Martini R (2010) Upper Triassic reef-limestone blocks of southwestern Japan: new data from a Panthalassan seamount. Palaeogeogr Palaeoclimatol Palaeoecol 293:206–222
Chan SA, Kaminski MA, Al-Ramadan K, Babalola LO (2017) Foraminiferal biofacies and depositional environments of the Burdigalian mixed carbonate and siliciclastic Dam Formation, Al-Lidamarea, Eastern Province of Saudi Arabia. Palaeogeogr Palaeoclimatol Palaeoecol 469:122–137
Coletti G, Mariani L, Garzanti E, Consani S, Bosio G, Vezzoli G, Hu X, Basso D (2021) Skeletal assemblages and terrigenous input in the Eocene carbonate systems of the Nummulitic Limestone (NW Europe). Sed Geol 425:106005
Ćosović V, Drobne K, Moro A (2004) Paleoenvironmental model for Eocene foraminiferal limestones of the Adriatic carbonate platform (Istrian Peninsula). Facies 50(1):61–75
Davies LM, Pinfold ES (1937) The Eocene beds of the Punjab Salt Range. Memoirs of the Geological Survey of India, Palaeontologia Indica 24:1–79
de Cizancourt M (1945) Nummulites nouvelles ou peu connues d’Aquiiaine. B Soc Géol Fr 15:643–655
Drobne K (1975) Hottingerina lukasi n. gen., n. sp. (Foraminiferida) iz srednjega paleocena v severozahodni jugoslaviji. Radzprave 18:242–253
Drobne K, Ćosović V, Moro A, Buckovic D (2011) The role of the palaeogene adriatic carbonate platform in the spatial distribution of alveolinids. Turkish J Earth Sci 20:721–751
Dunham RJ (1962) Classification of carbonate rocks according to depositional texture. In: Ham, W.E. (Ed.), Classification of Carbonate Rocks. American Association of Petroleum Geologists Memoir, pp. 1108–1121.
Dupuis C, Aubry MP, Steurbaut E, Berggren WA, Ouda K, Magioncalda R, Cramer BS, Kent DV, Speijer RP, Heilmann-Clausen C (2003) The Dababiya Quarry Section: Lithostratigraphy, clay mineralogy, geochemistry and paleontology. Micropaleontology 49:41–59
Duxbury A, Duxbury A (1997) An introduction to the world’s Oceans, 5th edn. WC Brown Publishers, Dubuque, p 504
Eder W, Hohenegger J, Briguglio A (2018) Test flattening in the larger foraminifer Heterostegina depressa: predicting bathymetry from axial sections. Paleobiology 44:76–88
Eichenseer H, Luterbacher HP (1992) The marine Paleogene of the Tremp region (NE Spain): depositional sequences, facies history, biostratigraphy and controlling factors. Facies 27:119–152
El-Ayyat AM (2022) Paleoenvironmental reconstruction, paleoecology and sequence stratigraphy of some Nummulites buildups in Egypt. Carbonates Evaporites 37(4):1–28
El-Ayyat AM, Obaidalla NA (2013) Stratigraphy, sedimentology and tectonic evolution of the Upper Cretaceous/Paleogene succession in north Eastern Desert. Egypt J African Earth Sci 81:35–59
Embry AF, Klovan JE (1972) Absolute water depth limits of Late Devonian paleoecological zones. Geol Rundsch 61:672–686
Flügel E (2010) Microfacies of carbonate rocks. Springer, Berlin, p 984
Geel T (2000) Recognition of stratigraphic sequences in carbonate platform and slope deposits: empirical models based on microfacies analysis of Paleogene deposits in southeastern Spain. Palaeogeogr Palaeoclimatol Palaeoecol 155:211–238
Ghose BK (1977) Paleoecology of the Cenozoic reefal foraminifers and algae-a brief review. Palaeogeogr Palaeocl 22:231–256
Gilham RF, Bristow CS (1998) Facies architecture and geometry of a prograding carbonate ramp during the early stages of foreland basin evolution. Lower Eocene sequences. Sierra del Cadf, SE Pyrenees. Spain. In: Wright VP, Burchette T P (eds). Carbonate ramps. Geology of Society London Special Publication 149: 181–203.
Gilmour I, Jolley D, Kemp D, Kelley S, Gilmour M, Daly R, Widdowson M (2014) The early Danian hyperthermal event at Boltysh (Ukraine): Relation to Cretaceous- Paleogene boundary events. In: Keller G, Kerr AC (Eds.), Volcanism, Impacts, and Mass Extinctions: Causes and Effects. Geological Society of America, Special Papers 505: 133–146.
Hadi M, Mosaddegh H, Abbassi N (2015) Biostratigraphic interpretation and systematics of alveolina assemblages in the Ziarat Formation from soltanieh mountains (Western Alborz). Geosci J 95:39–44
Hadi M, Mosaddegh H, Abbassi N (2016) Microfacies and biofabric of nummulite accumulations (Bank) from the Eocene deposits of Western Alborz (NW Iran). J Afr Earth Sc 124:216–233
Hadi M, Vahidinia M, Hrabovsky J (2018) Larger foraminiferal biostratigraphy and microfacies analysis from the Ypresian (Ilerdian-Cuisian) limestones in the Sistan Suture Zone (eastern Iran). Turk J Earth Sci 27:122–145
Hadi M, Vahidinia M, Hrabovsky J (2019) Larger foraminiferal biostratigraphy and microfacies analysis from the Ypresian (Ilerdian-Cuisian) limestones in the sistan suture zone (eastern Iran). Turk J Earth Sci 28(1):122–145
Hadi M, Sarkar S, Vahidinia M, Bayet-Goll A (2021) Microfacies analysis of Eocene Ziarat Formation (eastern Alborz zone, NE Iran) and paleoenvironmental implications. All Earth 33(1):66–87
Hallock P (1985) Why are larger foraminifera large? Paleobiology 11:195–208
Hasler CA (2004) Geometry and Internal Discontinuities of an Ypresian Carbonate Reservoir (SIT Field, Tunisia). PhD Dissertation, Terre Environ 45. University of Geneva, Geneva, Switzerland, p. 230.
Heim ARN (1908) Die Nummuliten und Flyschbildungen der Schwei zeralpen. Versuch zu einer Revision der alpinen Eocaen-Stratigraphie. Abh Schweiz Paläont Ges 35:1–301
Henson FRS (1948) Larger imperforate foraminifera of South-Western Asia, families Lituolidae, Orbitolinidae and Meandropsinidae. British Museum (Natural History). London 11:1–127
Höntzsch S, Scheibner C, Guasti E, Kuss J, Marzouk AM, Rasser MW (2011a) Increasing restriction of the Egyptian shelf during the Early Eocene? New insights from a southern Tethyan carbonate platform. Palaeogeogr Palaeoclimatol Palaeoecol 302(3–4):349–366
Höntzsch S, Scheibner C, Kuss J, Marzouk AM, Rasser MW (2011b) Tectonically driven carbonate ramp evolution at the southern Tethyan shelf: The Lower Eocene succession of the Galala Mountains. Egypt Facies 57:51–72
Hottinger L (1960) Recherches sur les Alvéolines du Paléocène et de l’Eocène. Schweizerische Palaeontologische Abhandlungen 75(76):1–243
Hottinger L (1983) Processes determining the distribution of larger foraminifera in space and time. Utrecht Micropaleontol Bull 30:239–253
Hottinger L (1997) Shallow benthic foraminiferal assemblages as signals for depth of their deposition and their limitations. Bull Soc Geol France 168:491–505
Hottinger L (1998) Shallow benthic foraminifera at the Paleocene-Eocene boundary. Strata Serie 1(9):61–64
Hottinger L (2014) Paleogene larger rotaliid foraminifera from the western and central Neotethys. New York, Dordrecht, London, Springer Cham, Heidelberg, pp 1–196
Hottinger L (1977) Foraminifères operculiniformes. Mémoires Museum National Histoire Naturelle Sèr. C40: 159 pp.
Hottinger L (2001) Learning from the past? In: Box, E., Pignatti, J. (eds.). Volume IV: The living world. Part Two: Discovery and Spoliation of the Biosphere. San Diego, Academic Press, 449–477.
Hussein IM, Abdallah AMA (2001) Tectonic evolution of the northeastern part of the African continental margin. Egypt J Afr Earth Sci 33:49–68
Ismail AA, Hussein-Kamel Y, Boukhary M, Ghandour AA (2007) Campanian-Early Eocene Stratigraphy of the Southern Galala Plateau, Eastern Desert. Egypt Geologia Croatica 60(2):115–137
Jorry SJ, Hasler CA, Davaud E (2006) Hydrodynamic behaviour of Nummulites: implication for depositional models. Facies 52:221–235
Jorry SJ (2004) The Eocene Nummulites Carbonates (Central Tunisia and NE Libya): Sedimentology, Depositional Environments, and Application to Oil Reservoirs. PhD, Terre Environ 48. University of Geneva, Geneva, Switzerland, p. 226.
Krenkel E (1925) Geologie Afrikas. Bornträger, Berlin, p 461
Kuss J, Leppig U (1989) The early Tertiary (middle-late Paleocene) limestones from the western Gulf of Suez. Egypt Neues Jahrbuch Für Geologie Und Paläontologie 177:289–332
Kuss J, Scheibner C, Gietl R (2000) Carbonate platform to basin transition along an upper Cretaceous to lower Tertiary Syrian arc uplift, Galala Plateaus. Eastern Desert of Egypt Geoarabia 5(3):405–424
Langer MR, Hottinger L (2000) Biogeography of selected “larger” foraminifera. Micropaleontology 46(1):105–126
Leckie R, Olson H (2003) Foraminifera as proxies for sea-level change on siliciclastic margins. SEPM Special Publication 75:1–19
Lee J (1990) Phylum Granuloreticulosa (Foraminifera). In: Margulis L, Corliss J, Melkonian M, Chapman D (eds) Handbook of Protoctista. Jones and Bartlett Publishers, Boston, pp 524–567
Leymerie A (1846) Mémoire sue le terrain á Nummulites (épicrétacé) des Corbières et de la Montagne Noire. Mémoires De La Société Géologique De France 2:337–373
Li X, Liu W, Wang Y, Zhu Q (1999) The tectonic evolution and metallogenesis in Tethys of the Nujiang-Langcangjing-Jingsajing area, southwestern China. Geological Publishing House, Beijing, pp 1–276
Loucks RG, Moody RTJ, Bellis JK, Brown AA (1998) Regional depositional setting and pore network systems of the El Garia Formation (metlaoui group, lower Eocene), offshore Tunisia. In: Macgregor DS, Moody RTJ, Clark- Lowes DD (Eds.), Petroleum Geology of North Africa, pp. 355–374. Geol. Soc. London Spec. Publ. 132.
Machaniec E, Jach R, Gradziński M (2011) Morphotype variation of orthophragminids as a palaeoecological indicator: A case study of Bartonian limestones, pod capkami quarry, tatra mts, Poland. Ann Soc Geol Pol 81:99–205
Mahfouz KH, Metwally AA (2020) Maastrichtian-Paleocene successions at Kharga-Dakhla stretch, Western Desert, Egypt: paleoenvironmental and basin evolution interpretations. J Afr Earth Sc 162:103731
Martín-Martín M, Guerrera F, Tosquella J, Tramontana M (2020) Paleocene-lower eocene carbonate platforms of westernmost tethys. Sed Geol 404:105674
Martín-Martín M, Guerrera F, Tosquella J, Tramontana M (2021) Middle Eocene carbonate platforms of the westernmost tethys. Sed Geol 415:105861
Mattern F, Scharf A, Al-Sarmi M, Al-Sayigh AR, Al-Maktoumi M, Al-Omairi N, Al-Rawahi T, Al-Moqbali M (2021) Lithostratigraphy, microfacies and paleogeography of the shallow marine Middle Limestone Member of the Early Eocene Rusayl Formation, Oman: Relationship to the Early Eocene Climatic Optimum, sea-level changes and regional uplift. J Afr Earth Sc 184:104312
Morsi AMM, Scheibner C (2009) Paleocene-Early Eocene ostracodes from the Southern Galala Plateau (Eastern Desert, Egypt): taxonomy, impact of paleobathymetric changes. Rev Micropaléontol 52(2):149–192
Mossadegh Z, Haig D, Allan T, Adabi M, Sadeghi A (2009) Salinity changes during Late Oligocene to Early Miocene Asmari Formation deposition, Zagros Mountains. Iran Palaeogeogr Palaeoclimatol Palaeoecol 272:17–36
Moustafa AR, Khalil MH (1995) Superposed deformation in the northern Suez Rift, Egypt: relevance to hydrocarbons exploration. J Petrol Geol 18:245–266
Murray JW (2006) Ecology and Applications of Benthic Foraminifera. Cambrige University Press, Cambridge, p 426
Orue-Etxebarria X, Pujalte V, Bernaola G, Apellaniz E, Baceta JI, Payros A, Nunez-Betelu K, Serra-Kiel J, Tosquella J (2001) Did the Late Paleocene thermal maximum affect the evolution of larger foraminifers? Evidence from calcareous plankton of the Campo Section (Pyrenees, Spain). Mar Micropaleontol 41:45–71
Özgen-Erdem N, Akyazı M, Karabaşoğlu A (2007) Biostratigraphic interpretation and systematics of Alveolina assemblages from the Ilerdian-Cuisian limestones of Southern Eskişehir, Central Turkey. J Asian Earth Sci 29(5–6):911–927
Pecheux M (1995) Ecomorphology of recent large foraminifera, Operculina ammonoides. Geobios 28:529–566
Pleş G, Oprişa A, Bucur II, Săsăran E, Mircescu CV, Oltean G, Iacob RG (2019) The central-western Getic Carbonate Platform: Upper Jurassic to Lower Cretaceous biostratigraphy and sedimentary evolution of the Cioclovina-Băniţa sector (Southern Carpathians, Romania). Facies 65(3):1–25
Pujalte V, Schmitz B, Baceta JI, Orue-Etxebarria X, Bernaola G, Dinarès-Turell J, Payros A, Apellaniz E, Caballero F (2009a) Correlation of the Thanetian-Ilerdian turnover of larger foraminifera and the Paleocene-Eocene thermal maximum: confirming evidence from the Campo area (Pyrenees, Spain). Geol Acta 7(1–2):161–175
Pujalte V, Baceta JI, Schmitz B, Orue-Etxebarria X, Payros A, Bernaola G, Apellaniz E, Caballero F, Robador A, Serra-Kiel J, Tosquella J (2009b) Redefiniton of the Ilerdian Stage (early Eocene). Geol Acta 7(1–2):177–194
Pujalte V, Orue-Etxebarria X, Schmitz B, Tosquella J, Baceta JI, Payros A, Bernaola G, Caballero F, Apellaniz E (2003) Basal Ilerdian (earliest Eocene) turnover of larger foraminifera: Age constraints based on calcareous plankton and δ13C isotopic profiles from new southern Pyrenean sections (Spain). In: Wing SL, Gingerich PD, Schmitz B, Thomas E (eds.). Causes and consequences of globally warm climates in the early Paleogene. Geological Society of America 369: 205–221.
Racey A (2001) A review of Eocene nummulite accumulations: Structure, formation and reservoir potential. J Pet Geol 24(1):79–100
Rasser MW, Scheibner C, Mutti M (2005) A paleoenvironmental standard section for early Ilerdian tropical carbonate factories (corbieres, France; Pyrenees, Spain). Facies 51(1–4):218–232
Reichel M (1937) Étude sur les Alvéolines. Mémoires Société Paléontologie Suisse 57(59):1–147
Saber SG (2012) Depositional framework and sequence stratigraphy of the Cenomanian-Turonian rocks on the western side of the Gulf of Suez. Egypt Cretaceous Research 37:300–318
Said R (1990) Cenozoic. In: Said R (ed) The geology of Egypt. Balkema, Rotterdam, pp 451–486
Samankassou E (2002) Cool-water carbonates in a paleoequatorial shallow-water environment: The paradox of the Auernig cyclic sediments (Upper Pennsylvanian, Carnic Alps, Austria-Italy) and its implications. Geology 30(7):655–658
Sarkar S (2019) Does specialization imply rare fossil records of some benthic foraminifera: Late Palaeocene examples from the eastern Neo-Tethys (Meghalaya, NE India). Palaeogeogr Palaeoclimatol Palaeoecol 514:124–134
Schaub H (1981) Nummulites et Assilines de la Téthys paléogène. Taxinomie, phylogenèse et biostratigraphie. Schweizerische Palaeontologische Abhandlungen 104/105/106: 1–238.
Scheibner C, Speijer RP (2008) Late Paleocene-early Eocene Tethyan carbonate platform evolution: a response to long-and short-term paleoclimatic change. Earth-Sci Rev 90(3–4):71–102
Scheibner C, Speijer RP (2009) Recalibration of the Tethyan shallow-benthic zonation across the Paleocene-Eocene boundary: The Egyptian record. Geol Acta 7:195–214
Scheibner C, Marzouk A, Kuss J (2001) Shelf architecture of an isolated Late Cretaceous carbonate platform margin, Galala Mountains (Eastern Desert, Egypt). Sed Geol 145:23–43
Scheibner C, Reijmer JJG, Marzouk AM, Speijer RP, Kuss J (2003) From platform to basin: the evolution of a Paleocene carbonate margin (Eastern Desert, Egypt). Int J Earth Sci 92:624–640
Scheibner C, Speijer RP, Marzouk A (2005) Larger foraminiferal turnover during the Paleocene/Eocene thermal maximum and paleoclimatic control on the evolution of platform ecosystems. Geology 33:493–496
Scheibner C, Rasser MW, Mutti M (2007) The Campo section (Pyrenees, Spain) revised: implications for changing carbonate assemblages across the Paleocene-Eocene boundary. Palaeogeogr Palaeoclimatol Palaeoecol 248:145–168
Schlagintweit F, Gawlick HJ (2008) The occurrence and role of microencruster frameworks in Late Jurassic to Early Cretaceous platform margin deposits of the Northern Calcareous Alps (Austria). Facies 54:207–231
Scholle PA (2003) A color guide to the petrography of carbonate rocks: grains, textures, porosity, diagenesis. Am. Assoc. Pet. Geol. Mem. 77: 470 pp.
Schütz KI (1994) Structure and stratigraphy of the Gulf of Suez, Egypt. In: Landon SM (ed) Interior rift basins, AAPG Memoir 59: pp 57–96.
Schwager C (1883) Die foraminiferen aus dem Eocaenablegerungen der Libyschen wuste und Agyptens. Paleontographica 30:81–153
Scott T, Campbell K, Rupert F, Arthur J, Missimer T, Lloyd J, Yon J, Duncan J (2001) Geologic map of the State of Florida: Florida Geological Survey Map Series 146.
Serra-Kiel J, Hottinger L, Caus E, Drobne K, Ferrandez C, Jauhri AK, Less G, Pavlovec R, Pignatti J, Samso JM, Schaub H, Sirel E, Strougo A, Tambareau Y, Tosquella J, Zakrevskaya E (1998) Larger foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene. Bulletin De La Société Géologique De France 169:281–299
Serra-Kiel J, Vicedo V, Baceta JI, Bernaola G, Robador A (2020) Paleocene Larger Foraminifera from the Pyrenean Basin with a recalibration of the Paleocene Shallow Benthic Zones. Geol Acta 18:1–69
Shahar J (1994) The Syrian Arc System: an overview. Palaeogeogr Palaeoclimatol Palaeoecol 112:125–142
Tomassetti L, Benedetti A, Brandano M (2016) Middle Eocene seagrass facies from Apennine carbonate platforms (Italy). Sed Geol 335:136–149
Torres-Silva AI, Eder W, Hohenegger J, Briguglio A (2019) Morphometric analysis of Eocene nummulitids in western and central Cuba: taxonomy, biostratigraphy and evolutionary trends. J Syst Palaeontol 17:557–595
Tosquella J, Serra-Kiel J (1998) Nummulites catari: a new species from the Late Palaeocene of the Pyrenean basin. Slovenska Akademija Znanosti in Umetnosti 34(2):165–171
Ungaro S (1994) Nummulite morphological evolution. Bull Soc Paleont Ital Spec 2:343–349
Wan X (1990) Eocene Larger Foraminifera from Southern Tibet. Rev Esp Micropaleontol 22:213–238
Wilson ME, Vecsei A (2005) The apparent paradox of abundant foramol facies in low latitudes: their environmental significance and effect on platform development. Earth Sci Rev 69:133–168
Wray JL (1977) Calcareous Algae. Elsevier Publishers, Amsterdam, p 190
Youssef MM (2003) Structural setting of central and south Egypt: an overview. Micropaleontology 49:1–13
Zachos J, Pagani M, Sloan L, Thomas E, Billups K (2001) Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292:686–693
Zamagni J, Mutti M, Košir A (2008) Evolution of shallow benthic communities during the late Paleocene-earliest Eocene transition in the Northern Tethys (SW Slovenia). Facies 54(1):25–43
Zamagni J, Mutti M, Ballato P, Košir A (2012). The Paleocene-Eocene thermal maximum (PETM) in shallow-marine successions of the Adriatic carbonate platform (SW Slovenia). Bulletin 124(7–8): 1071–1086.
Zhang Q, Willems H, Ding L (2013) Evolution of the Paleocene-Early Eocene larger benthic foraminifera in the Tethyan Himalaya of Tibet, China. Int J Earth Sci 102:1427–1445
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Materials were prepared and data were collected by S. A. and Y. S.. S. A. and A. M. performed the literature search. The various analyses were performed by all the authors. The first draft of the manuscript was written by S. A. and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abd-Elhameed, S., Mahmoud, A.A. & Salama, Y. Late Paleocene–Early Eocene larger foraminifera from the Galala Plateaus, North Eastern Desert, Egypt: biostratigraphic, paleoenvironmental and paleoecological implications. Carbonates Evaporites 38, 84 (2023). https://doi.org/10.1007/s13146-023-00909-2
Accepted:
Published:
DOI: https://doi.org/10.1007/s13146-023-00909-2