
Abstract
The present study deals with microfacies analysis, paleoecology and sequence stratigraphy of the Lower–Middle Eocene rocks in central Nile Valley and west-central Sinai. Four rock units were marked (Minia, Darat, Samalut and Maghagha formations). Eight facies belts were interpreted including lagoon, back bank, main bank, fore bank and open marine outer bank facies and deposited on a gently dipping platform ramp. This ramp might be further divided to three main faunal associations depending on the depth of the water and associated fauna. Four major depositional sequences (DS1–DS4), reflecting third-order cycles and separated by two prominent sequence boundaries (SB1 and SB2) were marked. The record of these sequences reflects the changes in eustatic sea level plus the regional tectonism in the area. Every sequence includes facies associations, which constitute lowstand and/or transgressive and highstand systems tracts. Based on faunal elements and facies analysis, the water salinity, water temperature, nutrients, substrate, water depth and hydrodynamic behavior were discussed. The fine-grain size of the deposits, the loss of good connectedness and the occluding of the Nummulite tests by other minerals could give rise to low reservoir potential. The sedimentation was affected by the tectonic activity of the Syrian orogeny. Moreover, both external and internal mechanisms have controlled the carbonate sedimentation, basin construction and sequence bounding surfaces.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Nummulite buildups form paramount hydrocarbon reservoirs in Libya and Tunisia and are considered as promising exploration domains in Pakistan, Oman, Egypt and Italy (Racey et al. 2001). They dominate the continental shelf of Tethys (Fig. 1) and host larger benthic foraminifera (LBF) as alveolinid, nummulitid and orthrorphragminid (Buxton and Pedley 1989). Study of larger benthic foraminifera yields vital evidences for paleoenvironmental settings. Kiessling et al. (2003) mentioned that suitable paleogeography with warm climate resulted in increase of carbonate production in temperate areas lying at more than 45°N latitude with LBF and algae as the major sediment contributors. Nummulites are considered as rock-forming fauna and accumulate in typical banks, sometimes related to paleoreliefs. Both the initiation and paleoecology of these banks are still not completely understood (Bassi et al. 2013). Two kinds of processes are involved: biological (i.e., reproduction strategies) and physical (i.e., winnowing and sorting) are the main mechanisms for the building and structure of Nummulite banks, but Racey (2001) added that both bioturbation and compaction could modify the original fabric. Generally, LBF show a marked reaction even to subtle variations in environmental features. Therefore, they are good paleoenvironmental markers (i.e., salinity, water depth and oxygenation) and manifest the impact of these factors on LBF associations (Scott et al. 2001). Jorissen et al. (1995) mentioned that the number of LBF, their microhabitat structure, their species composition and their diversity mainly rely on the food and oxygen levels. The study area (Fig. 2) shows good exposures, with broad facies changes of carbonates and mixed carbonate–siliciclastics of the Eocene age. Several studies have dealt with the Eocene rocks in Egypt (i.e., Bandel and Kuss 1987; Hermina and Lindenberg 1989; Kulbrok 1996; Bignot and Strougo 2002; El Ayyat and Obaidalla 2016). The present work delineates the various sedimentary environments and their resultant facies and introduces paleoecological interpretations of the shallow, mixed carbonate–siliciclastic platform under investigation. This case study could be considered as a base for clarification of shallow marine depositional regimes and displays a frame for comparing with other Nummulite buildups in the Eocene sediments in and outside Egypt. It is also intended to highlight the significance of LBF and algae as sediment contributors and paleoenvironmental indicators and to perform a tentative first sequence stratigraphic analysis based on the facies association variations and their stacking patterns. This would enable correlating them with the sea-level oscillations in relative sea level and to provide an interpretation of the changes in depositional regime and facies with time and under the conditions of changing tectono-sedimentary boundaries.

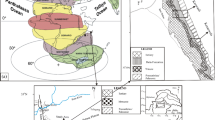
Paleogeographic map illustrating the spatial distribution of the Nummulite buildups during the Eocene period (modified after Racey 2001; Scotese 2001; Jorry 2004). Areas under the study have been highlighted in red ink stars, while black ones point to the positioning of the Nummulite buildups worldwide as: 1 = Senegal; 2 & 3 = Tunisia; 4, 5 & 6 = Libya; 7 = Egypt; 8 = Madagascar; 8 = Oman; 10 & 12 = Iran; 11 = Saudi Arabia; 13 = Cyprus; 14 & 15 = Turkey; 16 = Italy; 17, 18, 19, 20, 21, 22 & 23 = Southwestern Europe; 24 and 25 = Spain; 26 = Sri Lanka; 27 = India; 28 = Southeast Asia

Location map exhibiting the aerial distribution of the Eocene lithofacies in Egypt (after Said 1990) and the location of the measured sections in both the central Eastern Desert (Nile Valley) and west-central Sinai
Materials and methods
Field work
The field area occupies two sectors: the central division of the Eastern Desert and west-central Sinai (Fig. 2). They extend between 31°00′, 33° 30′ east and 27° 30′, 28° 30′ north, approximately lying in the Eastern Desert, east of Minia City and in west-central Sinai on the eastern shoulder of the Gulf of Suez (Fig. 2). Legend for explanatory symbols are shown in Fig. 3. Five surface sections were measured, described and sampled bed by bed, considering the lithology, texture, sedimentary structures and entombed biota. These stratigraphic sections are distributed from north to south (Fig. 2) as follows: Wadi Feiran section (33.430560 E, 28.789601 N); Wadi Tarfa section (30.986262 E, 28.789601 N); Wadi Tihna section (30.783010 E, 28.185396 N); Abu Qurqas section (30.885823 E, 27.72538 N) and Barsha section (30.90114 E, 27.72538 N). About 256 rock samples were picked up plus polished cut slabs (10 × 15 cm) were sawed, polished and photographed. Significant facies contacts and important stratigraphic boundaries were marked and photographed, while facies characteristics, stratal relationships and sample locations were noted.

Legend illustrating the symbols for rock types and faunal community in the area under consideration
Laboratory methods
The petrographic descriptions are based entirely on the microscopic investigations of 242 thin sections. Such investigations include visual determination of lithology; description of grain type, texture, fossil content and frequency estimation of bioclasts (Flügel 2004). The nomenclature of the facies follows that of Dunham (1962) and Embry and Klovan (1971) classification. To differentiate dolomites, thin sections were stained with alizarin-red S solution and potassium ferricyanide (Dickson 1965). Thin section photomicrography was performed by camera Zeiss MC 80. Petrographic composition was analyzed by point counting (300 points) on each thin section to develop grain size and quantities of grains and cements. Sedimentologic textures and structures were considered qualitatively. The paleoenvironmental reconfiguration and paleoecological elucidation such as water temperature, intensity of light, water salinity, nutrients, water depth, hydrodynamic energy of seawater and nature of substrate were established based on the works of Hohenegger (2004), Flügel (2010) and BouDagher-Fadel (2013). Literature on Lower Paleogene successions next to the goal areas were integrated and compared to clarify the syntectonism and to reset a more exact depositional history. Accordingly, the local analysis of rising, erosion, deposition and renewed subsidence was discussed taking into account the syntectonic instability, which formulated the sedimentary basins.
Geologic and stratigraphic setting
Geographically, the study area includes two salient realms: the Nile Valley realm and west-central Sinai realm. The first one is considered as a slice of the major limestone plateau paralleling the cultivated land on the eastern bank (El Maaza plateau). The scarp of the plateau strikes in the N–S direction (Fig. 2). Generally, the plateau slopes toward the NW direction. Therefore, folds and gentle warps are clearly observed along the scarp facing the Nile, while joints are generally vertical and common near the fold axis. Said (1990) considered it as simple structure and ascribed it to the Egyptian stable shelf. Brioussov (1968) recorded two distinct anticlines separated by a syncline in the midst of Samalut and Minia cities (in the Eastern Desert). Syn- and post-Eocene tectonics are considered as the main controlling parameter that influenced the depositional regime and patterns of the different Eocene lithofacies. The geographic occurrence of the Eocene lithofacies indicates that the vast Nile basin region might have formed an open gulf extending from the Tethys after the uplifting of the Red Sea area (Said 1990). The studied section in Sinai is located on the eastern side of the Suez Gulf near the central part of Sinai (Fig. 2). Shata (1956) divided the central part of Sinai to two structural units: the south stable foreland unit and the north, gently folded, central Sinai unit. The rift of the Gulf of Suez (Oligo-Miocene) cuts through the continuous Arabo-African plate. Since the pre-Cambrian, the whole area behaved as a platform. The Eocene sediments are limited in their geographical distribution in west-central Sinai (Fig. 2). They cover several tops in Wadi Feiran and exposed in the isolated outcrop in the extreme south of the Egma plateau.
Stratigraphically, several authors (e.g., Boukhary and Abdel-Malik, 1983; Mansour and Philobbos 1983; El Hinnawi et al. 2003; Hewaidy et al. 2015) proposed numerous formations spanning Late Ypresian–Late Lutetian interval in different areas in Egypt. The Formational names of the authors Mansour and Philobbos (1983) and El Hinnawi et al. (2003) have been followed in this study. Therefore, the study area can be subdivided into four formations: Minia Formation of Late Ypresian age, Darat Formation of Early Lutetian age, Samalut Formation of Early to Middle Lutetian age and Maghagha Formation of Late Lutetian age. In the following paragraphs, the lithology, thickness, areal distribution and faunal content for each formation are discussed.
Minia formation
In its type locality (Zawyet Sawada, opposite Minia city), the Minia Formation is described as composed mainly of carbonates with different structures and textures, thick-bedded, white, alveolinal limestone. It represents bar and ramp coastal depositional environments (Philobbos and Keheila 1979). There, it overlies the Thebes Group with seeming conformity and underlies the Samalut Formation. In the study area, it dominates the basal portion of the target succession (Figs. 4 and 5A) and its exposed thickness ranges between 23 and 74 m. It consists of grayish white to dark gray, biogenic limestone flooded with elongated alveolines, small and medium-sized Nummulites, bivalves and echinoderms. Chert boulders and nodules are scattered here and there approaching different sizes and shapes. Its contact with Samalut Formation is clear in the Nile Valley, but is rarely so inland. Due south, this contact coincides with a prominent marker bed of a relatively darker concretionary siliceous limestone bed (up to 1 m thick), which overlies a thinly laminated lime mudstone. Traveling north, this junction is expressed as siliceous limestone covered by marl or calcareous shale of 20–70 cm thick (Fig. 5A, photo inset) as paleosol horizon. The Minia Formation is missed at Wadi Feiran in Sinai (Fig. 6) and Wadi Tarfa (Fig. 7) in the Nile Valley because of lateral facies changes in Sinai or unexposed on the ground (being covered by younger sediments due north at Wadi Tarfa). The geologic age of Minia Formation is estimated as late Early Eocene according to Boukhary and Abdel-Malik (1983).

A Stratigraphic correlation for the studied rock units with that advocated by different authors in the surrounding areas. B Spatio-temporal correlation of the stratigraphic logs showing the distribution of the different rock units under study. locations in Fig. 2

A Field photograph displaying the Minia Formation rests unconformably under the Samalut Formation. The boundary is represented by paleosol horizon (photo inset, yellow arrow). Note the partition of the Samalut Formation into two parts (A and B). Nile Valley. B Field view exhibiting Samalut Formation rests unconformably above the Darat Formation. Note the two parts of the Samalut Formation (A and B). Wadi Feiran, Sinai. C Field photograph for the Darat Formation in Wadi Feiran. It includes yellowish brown, moderately hard, argillaceous limestone horizons intercalated with pale green, soft, thick-bedded shale. Note the increase of limestone on the expense of calcareous shale on the vertical scale. Westcentral Sinai. D Outcrop photo exhibiting a sequence of massive, ledge forming, poorly-bedded and amalgamated limestone, flooded with Nummulites gizehensis with cavernous, brownish weathered surfaces. Wadi Feiran, Sinai. E Field photo showing remnants of the Maghagha Formation overlying conformably Samalut Formation. Central Nile Valley, Wadi Tihna section. F Outcrop photo illustrating the interbedding between thin to medium bedded, soft, pale yellow, marly limestone and calcareous shale and marls of the Maghagha Formation. Central Nile Valley, Wadi Tihna section

Graphic stratigraphic log of Barsha section (Nile Valley) showing the distribution of the sedimentological characteristics, water energy and depositional environments (legend in Fig. 2)

Graphic stratigraphic log of Wadi Tihna section (Nile Valley) showing the distribution of the sedimentological characteristics, water energy, and depositional environments (legend in Fig. 2)
Darat formation
In its type locality (Wadi Nukhul, west-central Sinai), Viotti and El Demerdash (1968) described Darat Formation as a rock unit composed of alternating green and brown shales and marls with few limestone intercalations, flinty in the middle and chalky at the top with total thickness of 98 m thick. Herein, the upper levels of Darat Formation underlie unconformably the Samalut Formation. It crops out as moderately hard, yellowish brown, marly limestone interbedded with soft, pale green, medium- to thick-bedded calcareous shale (Fig. 5C). Chert balls and nodules are scattered almost parallel to bedding and limestone content increases at the expense of shale upsection. The faunal content indicates Middle Eocene age (El Heiny and Morsi 1986).
Samalut formation
In its type locality (opposite Samalut City, north of Minia, Eastern Desert), Bishay (1966) described this formation as the first Nummulites gizehensis beds of open bay environment and reefal in the upper part (Philobbos and Keheila 1979). There is thick and poorly bedded limestone of cream color and cavernous with scattered chert nodules, partly hard and nummulitic (up to 160 m thick). In the target area, it is recorded in Sinai and Nile Valley with residual thickness in the Nile Valley ranges from 10 to 52 m thick (Fig. 5A), but approaches nearly 43 m thick in Sinai (Fig. 5B). Due extreme south at Barsha section in the Nile Valley (Fig. 10), this formation and the upper part of the Minia Formation are absent due to erosion. The lithofacies and biota of the Samalut Formation, as recognized in Sinai, have similar affinities to those that occur in the Nile Valley and identified by Bishay (1961) in the Nile Valley. It includes calcareous shale at its base and rests unconformably over the marker silicified concretions of the underlying Minia Formation (Fig. 5A). In Sinai, Samalut Formation covers unconformably Darat Formation (Fig. 5B). It crops out as massive, ledge forming, poorly and amalgamated bedded limestone, crammed with Nummulites gizehensis (Fig. 5A), scattered chert nodules, pelecypods and echinoderms with sparse planktic and benthic foraminiferal fauna suggesting a Lutetian (Middle Eocene) age. Lithologically, it could be segmented into two informal parts (Fig. 5A and B). Its basal part (A) in the Nile Valley consists of grayish white, thin- to medium-bedded, moderately soft and occasionally burrowed nummulitic limestone. The upper one (B) consists chiefly of nummulitic limestone of pinkish white color, generally massive-bedded to crudely graded bedding, flooded with large Nummulites gizehensis in association with pelecypods and echinoderms near the base. In Sinai, the lower part (A) is argillaceous, soft, moderately bioturbated, bioclastic limestone (Fig. 5B), which tops unconformably the underlying Darat Formation. On the other hand, the upper one (B) includes a sequence of bedded limestone, crowded with Nummulites gizehensis and chert balls with nodules (Fig. 5B and D).
Maghagha formation
Bishay (1966) introduced Maghagha Formation to describe a rock sequence (110–165 m thick), which occurs to the east of Maghagha and Beni-Mazar towns. Maghagha Formation covers conformably the Samalut Formation and lies under the Qarara Formation of Late Lutetian age in the Nile Valley due north outside the study area (Fig. 5E). The boundary between Maghagha and Samalut formations is clearly traceable at the upper surface of the massive-bedded limestone enriched with large Nummulites gizehensis. It occurs in the Nile Valley and is completely absent in west-central Sinai. It is composed of thin- to medium-bedded, soft, pale yellow, marly limestone interbedded with calcareous shale and marls with a residual thickness of 7–9 m (Fig. 5F). According to its fossil content (Globorotalia centralis, G. lehneri and Globigerina venezuan), this formation (Fig. 4) has been dated Late Lutetian (Omara et al. 1978; Mansour and Philobbos 1983).
Sedimentology
The studied Eocene rocks were mostly accumulated over a gently, low-angle seaward dipping homoclinal ramp having a northern dip direction. This assumption is based on scarcity in reef-building organisms; the slump/slide structures and deposits, which would exist in high-angle platforms of steep slopes, are totally missed and the Nummulites occur almost in the middle and upper partitions of the investigated sequence (Figs. 6, 7, 8, 9, 10). Following Wright and Burchette (1996) classification of ramp profile, the studied Eocene rocks could be classified into outer ramp, mid ramp and inner ramp (Fig. 11). Faunistically, this ramp profile can be further classified into a sequence of depth-controlled faunal communities with the shallowest biota in the south and the deepest in the north. Furthermore, careful investigations led to identification of eight facies zones (Fig. 11). Summary of the characteristic features of the inferred depositional environments and their corresponding facies is introduced in Table 1. The facies belts have been differentiated based on composition and prevailing sedimentary structures. The depositional basin shows a steady change from relative shallow to deep parts of a platform setting, toward the southern and the northern sectors of the focus area, respectively. Modeling of the depositional basin illustrates that the facies display a gradual shallowing-upward trend. This shallowing upward is concurrent with a regional vertical tectonic activity in the north of Egypt at the start of the Eocene that was moreover accompanied by local syntectonics. Interbedded storm beds helped to estimate approximately the paleo-bathymetric depths of the depositional centers.

Graphic stratigraphic log of Abu Qurqas section (Nile Valley) showing the distribution of the sedimentological characteristics, water energy, and depositional environments (legend in Fig. 2)

Graphic stratigraphic log of Wadi Feiran section (west-central Sinai) showing the distribution of the sedimentological characteristics, water energy, and depositional environments (legend in Fig. 2)

Graphic stratigraphic log of Wadi Tarfa section (Nile Valley) showing the distribution of the sedimentological characteristics, water energy and depositional environments (legend in Fig. 2)

Schematic diagram for the Early to Middle Eocene studied succession (Nile Valley and Sinai), showing generalized depth-controlled distribution of the main larger and smaller groups of foraminifera and the location of the main Nummulite buildups in a mid-ramp domain
Paleoecology
Most extant LBF are neritic marine fauna, living largely in warm, nutrient-poor, reef and carbonate shelf environments, where they are the main producers of carbonate sediments (carbonate factories). It is concluded that LBF had similar distributions in the Mesozoic and Cenozoic. Modern taxa have geographic ranges equivalent to that of hermatypic kind of corals, in spite of some larger foraminifera showing wide distribution, north and south of the Equator. Hallock (1987) mentioned that integrating between studies on LBF and planktic foraminifera could help to have a rough guide to the main variations in the temperatures of the seawater during the last 66 million years (Hallock 1987).
Compositionally, the sediment grain types herein consist mainly of LBF, algae, mollusca and corals with subsidiary percents of bryozoa and echinoderms. All these components were accumulated in the carbonate sequence on a homoclinal carbonate ramp system (Fig. 12). BouDagher-Fadel (2008) claimed that the fossil foraminifera show many varieties of sizes as well as shapes and inhabit many varied environments, from the shallowest (near-shore) to the deepest (the deep sea). The distribution patterns of LBF are determined by a complex set of interrelated parameters including temperature, substrate, light, depth, nutrients, hydrodynamic energy and salinity (BouDagher-Fadel 2008). The most influential parameters, in shallow shelves, are nutrients sources and light, where primary production is most pronounced (Duxbury and Duxbury 1997).

Spatio-temporal effects of various paleoecological and environmental parameters on the distribution of larger and smaller foraminifera with algae
Light intensity
In the seawater, light intensity depends on water transparency and water depth. In other words, with increase in water depth, light intensity reduces and water transparency increases (Hallock 1987). Three groups of organisms, according to the dependence on the light, are considered as follows: (1) light-dependent organisms (algae, corals and porcelaneous foraminifers); (2) low light-dependent organisms (algae, Operculines or Lepidocyclines); (3) light-independent organisms (bryozoa, mollusca, crinoids and sponges).
Herein, different species of foraminifera are best indicators to reconstruct the paleo-depth conditions of seawater as porcelaneous forms, which are light dependent and abundant in shallow marine setting (restricted inner lagoon, Fig. 12A) of the Minia Formation, while they are rarely found in deep waters. On the contrary, the quantity of hyaline forams is mostly enhanced by increasing the water depth. Bulged and stout lenticular Nummulite shells predominate in shallow marine shelf (Fig. 12B), inhabiting the basal portion of the top of the photic zone (e.g., Nummulites bank), but thin-shelled tests (Fig. 12C) considerably occur in deep waters (Beavington-Penney et al. 2004) in fore-bank environment. Variation in the shell shapes signals lowering light grades at deeper parts of the sea floor (BouDagher-Fadel 2008). The green calcareous algae and subordinate corals recorded in the lagoon-bay sediments within the Minia Formation (Figs. 4, 5, 6 and 11) are typical of euphotic–mesophotic zone, with ultimate depth of prolific growth commonly about 20–30 m depth. On the other side, red algae span the shallow shelf from the shallowest (the lagoon) to the deepest (shelf break) and may extend down shelf, but thrive mostly between 40 and 90 m in the meso-oligophotic zone, where they are associated with larger foraminifera (Pomar 2001). Association of red algae with large foraminifera represents deposition in an oligophotic zone of a carbonate ramp under tropical to subtropical condition (Pomar 2001; Brandano and Corda 2002).
Planktic forams are also presented in the deepest parts of a carbonate ramp (Beavington-Penney et al. 2004) in open marine setting represented by Maghagha and Darat formations (Fig. 12D). The occurrence of planktic forams points to oligophotic situations, while LBF such as Nummulites, Heterostegines and Operculines reflect masochistic to oligophotic circumstances (BouDagher-Fadel 2008). Assembly of LBF and symbiont red algae is well dependent on the photic area (Barattolo et al. 2007). Thus, the investigated sedimentary sequence accumulated under oligophotic to mesophotic conditions.
Temperature
Temperature has impact on benthic fauna by affecting the metabolic performance and dissolution of CO2. A decrease in temperature decreases the metabolic performance and consequently the organism’s requirements of nutrient (Geel 2000).
In the focus area, it seems that each combination of organisms lives in a particular temperature range; for instance, most of the LBF (Fig. 12E) exist in the tropic and subtropic areas and enclosures at temperatures 18–23 °C (Wilson and Vecsei 2005). Herein, the gathering of porcelaneous foraminifera (Peneroplis and Alveolines) with corals and red coralline algae (Fig. 12F and G) may suggest environmental temperatures from 18 to 22 °C in tropical environment (Wilson and Vecsei 2005) and planktic forams (Fig. 12H) can exist in water temperatures < 14 °C (Geel 2000).
The presence of red algae resembles the Miocene coralline algal bioherm from Malta described by Bosence and Pedley (1979) as non-reefal algal limestone. In general, the presence of LBF in the rock record points to a warm environment, while their absence points to cooler or more nutrient-rich environments. Some extant larger foraminifera can tolerate water temperatures as low as 10–11 °C, including Amphistegines and Sorites in the Mediterranean, and Amphisorus and Amphistegines on the southwest Australian shelf (Li et al. 1999). LBF and corals, to some extent, would reside in warm waters having temperatures from 18–20 to 23–25 °C, respectively (Brandano et al. 2009b). All the mentioned indicators show that the studied Eocene rocks probably had a tropical–subtropical climate because of the extensive occurrence of benthic foraminifera, algae, corals and Bryozoa.
Salinity
It has a direct influence on the evolution of LBF, and therefore concerning the existence of large and long Discocyclines with flat, large Nummulite shells in a fore-bank silty to muddy floor (Fig. 11), which are pointers to a normal salinity of seawater with a low energetic condition (Scheibner et al. 2005), in the area of the study. Conversely, co-occurrence of imperforate porcelaneous foraminifera with dolomites of the tidal flats points to higher water salinities shoreward due south (Fig. 12).
Accordingly, due south (landward), water salinity can reach higher than 50 psu (Fig. 12) due to the existence of imperforate foraminifera and lack of red algae and corals (Mossadegh et al. 2009). Murray (2006) stated that Alveolines, Peneroplis and Orbitolites would not tolerate salinity higher than 50 psu (Fig. 12I).
The co-occurrence of imperforate foraminifera (Borelis, Archaias, and Peneroplis, Fig. 12J) with red algae and corals mirrors water salinity in the range of 42–51 psu (Mossadegh et al. 2009). The existence of porcelaneous foraminifera (i.e., peneroplidae and milolids), mollusca fragments, absence of hyaline foraminifera and other representatives of open marine biota and low frequency of calcareous agglutinated foraminifera mark the dominance of higher levels of hypersaline water in enclosed lagoon milieu. In hypersaline environments, miliolids are prevailing facies type, enriched with skeletal material (Mossadegh et al. 2009).
The occurrence of LBF with perforated wall (such as Nummulites, Operculines and Discocyclines) refers to normal seawater conditions (Fig. 12K) with salinity ranges from 32 to 38 psu (Mossadegh et al. 2009). Planktic foraminifera live in open ocean and also in inland seas (i.e., the Mediterranean). Their tolerance to variations in water salinity is very limited; indeed, most extant members are precisely stenohaline (Fig. 12L). No living planktic forams have been recorded in waters whose salinity was above 40 psu or beneath 32 psu (Murray 2006). Consequently, it could be mentioned that the upper levels of the investigated succession (Samalut and Maghagha Formation) accumulated in normal marine salinity conditions, but the lower and middle levels (Minia Formation) witnessed interrupted normal and higher water salinity circumstances.
Nutrients
LBF live in stabilized, nutrient-deficient, oligotrophic situations. On the other hand, LBF cannot react competitively if the nourishing supplies are plenty (Hallock 1985). The imperforate foraminifera (i.e., milliolids), which inhibit inner confined lagoon (Figs. 11 and 12M) indicate low grades of nutrients (Samankassou 2002). Coral reefs develop in submarine environments (i.e., open shelf lagoon) with the lowest level of nutrients (Figs. 11 and 12N) and easily die with excess of nutrients (Schlager 2005). Increase of nutrients gives rise to thriving red coralline algae and decrease in nutrients causes coral growth and development (James et al. 1999). The Nummulites bank under study (Fig. 12O) had a little associated micro- or macrofauna (mono-specific bank), suggesting that deposition was in a nutrient-poor (oligotrophic) environment and/or in a setting with significant hydrodynamic sorting. Therefore, the predominance of Nummulites indicates that the depositional environments were nutrient-poor or oligotrophic conditions (Hallock 1985). Generally, sediments rich in LBF are often inferred to represent deposition in oligotrophic settings, since such sediments are overwhelmingly accompanied with organisms tolerant of nutrient-poor conditions (i.e., calcareous algae and corals, Fig. 12G). The occurrence of Orbitolites, Milliolides, Peneroplis and Operculines with red coralline algae corals herein reflects mesotrophic to oligotrophic conditions (Fig. 12M and N). Zooxanthellate corals and the LBF assemblages of the goal area (imperforate porcelaneous and perforate hyaline) argue against persistently high nutrients, since these foraminifera grow in slightly mesotrophic (Halfar et al. 2004) to possibly oligotrophic water conditions (Langer and Hottinger 2000). This is confirmed by the missing of low-oxygen foraminiferal associations and the infrequent representation of filter-feeding fauna (serpulids and barnacles) that usually inhabit environments with relatively abundant organic matter. Open marine planktic foraminifera of both Maghagha and Darat formations (Fig. 12P) such as Globorotalia, Globigerina and Turborotalia (Omara et al. 1978; El Heiny and Morsi 1986) indicate surface trophic to intermediate mesotrophic water conditions (BouDagher-Fadel 2008).
Substrate
Nummulites possess many shapes, which resulted from the reproductive strategies and environmental conditions such as substrate and water hydrodynamic energies. These factors, of course, affect the thickness, size and the shape of the shells (Cosovic et al. 2004). Porcelaneous imperforate foraminifera (Fig. 12Q) live on substrate of comparatively low stability in shallow shelf (Geel 2000). Operculines (in the back-bank environment) inhabit soft substrates (Fig. 12R), while Heterostegines prefer to live on hard sediments (BouDagher-Fadel 2008). Generally, the biota suggests muddy, fine-grained substrate conditions. This interpretation is based mainly on the predominance of echinoids and burrowing bivalves in a muddy back-bank site behind the barrier of the Nummulites bank. Moreover, Archaias, Heterostegines and Operculines can live in environments with about 40% of clastic influx, but increase in clastic influx leads to reducing their number, while Miogypsines are less affected by clastic influx (Kumar and Saraswati 1997). Brasier (1975) mentioned that thick-shelled and conical forms (Fig. 12S) of LBF prefer housing coarse-grained sediments, but flattened shell forms (Fig. 12T) prefer soft substrates.
Water depth and hydrodynamic behavior
Changes in water depth could affect environmental changes such as sedimentation rate, turbulence, light intensity, organic change, water hydrodynamic energy and dissolved oxygen (Leckie and Olson 2003). LBF due to being sensitive to water depth in the early times were used to estimate water depth. Alveolinids (Fig. 12U) and miogypsines avoid very shallow water and live in sheltered habitats, away from oceanic wave energies, and thus are particularly common in deeper parts of restricted lagoonal settings (Murray 2006). Some LBF such as Borelis, Archaias and Peneroplis (Fig. 12M) are markers for shallow waters, tropical environments further down than 30 m (Brandano et al. 2009a). Some LBF such as Nummulites and Discocyclines (Figs. 12V and W) become flatter, with thinner outer walls, with increasing water depth (middle ramp Nummulites bank, Fig. 11) and decreasing light intensity. This reveals the dependence of morphology on the gradient of the depth (Pecheux 1995). Perforate forams (i.e., Nummulites, Operculines, Miogypsines, and Amphistegines) are abundant in subtropical to tropical environments reaching down to 40–70 m depth (Hottinger 1997). Herein, and near the base of the target succession, Miliolides, Peneroplis and Borelis are plentiful and mostly found in a shallower and restricted water depth. Conversely, within the middle and upper portions, Nummulites, Operculines, and Discocyclines (as perforate forams) are common and reflect open marine conditions (Fig. 12). Algae, such as Lithoporella, Lithophyllum and Lithothamnium (Fig. 12J), show subtropical to tropical environments, with depth of 20–80 m (Flügel 2010). Murray (2006) mentioned that the occurrence of planktic forams (i.e., in both Maghagha and Darat formations) and deficiency of benthic fauna (Fig. 12X) suggest deposition in tranquil deep water, in open marine outer shelf, commonly under the reach of current attack. Generally, the planktic foraminifera are noticeably abundant, indicating water depth environment was greater than 200 m (outer ramp environment). The hydrodynamic behavior of LBF, which depends on density, size and their shapes, is an essential factor controlling their transport (BouDagher-Fadel 2008). Depending on hydrodynamic water energy, some foraminifera develop thick lamellar crust shells (Fig. 12B) and others tend to develop a hard, conical shell shape (Fig. 12S) with abundant secondary cells (Rasser et al. 2005). The living nummulitids house symbiotic microalgae; they prefer calm water conditions and keep away from highly lighted realms close to the surface of the water, because the flat shells (Figs. 12K and O) could be readily damaged in a turbulent hydrodynamic regime (Hohenegger 2000). Herein, in the studied succession, Operculines and Discocyclines occurred in depth zones grading from high-energy, shallow water fore-bank facies (BouDagher-Fadel 2008) down to calm sea waters near the base of the photic zone (Chaproniere 1975). Orbitolites and Operculines prefer to house subtidal and lagoonal milieus that are dominated by low and medium levels of water energies (Hohenegger 2000 and 2004).
Accordingly, relying on the ecological influence of the aforementioned parameters on the spatio-temporal classification of the faunal elements (e.g., foraminifera and calcareous algae besides gastropods, pelecypods, echinoderms and scattered Bryozoa), the facial tabulation of the different groups of the Foraminifera and their coordinate depositional environments has been sketched (Figs. 11 and 12) taking into account the zones of distribution depth of recent foraminifera in seawater, benthic foraminiferal community toward the Paleocene–Eocene interval in the Campo section (Scheibner et al. 2007); distribution of Paleocene–Early Eocene LBF in SW Slovenia (Zamagni et al. 2008) and the Early Eocene foraminiferal-rich limestone in the Pyrenees area (Rasser et al. 2005).
Inner ramp faunal community (porcelaneous forams and green algae)
This association has been recorded in the Minia Formation (Fig. 11). Therefore, it builds the outer lagoon, shoals and restricted shelf lagoon. It contains low-diversity fauna (Alveolines, Textulariids, Orbitolites and Miliolides, Fig. 12A, F, I and M). Associated with them are codiacean and dasycladacean algae. Ecologically, this faunal association was the most suitable for existing marine conditions such as low rate of turbidity, intensive light and low-substrate constancy. The low turbidity is ascribed to the high degree of variation of the porcelaneous forams, which evolve in meso-to-oligotrophic settings, at shallow depths (Barattolo et al. 2007). Alternatively, lagoons of open shelf circumstances are housed by mixed normal marine and restricted biota such as Alveolines, Bryozoa, corals and echinoderms (Figs. 12F and N). According to Lee (1990), porcelaneous forams are prominent in shallow water settings in present-day tropics and subtropics, including dinoflagellate, rhodophycean and chlorophycean endosymbionts. In the studied inner ramp realm (Fig. 12), benthic forams indicated sedimentation inside the photic zone, in an area dominated by seagrass. Similarly, porcelaneous foraminiferal assemblages have been notified from Iraq and Iran (Sartorio and Venturini 1988), Turkey (Sirel 2003), southern Oman (Gallardo et al. 2001), and Italy (Bassi et al. 2007).
Middle ramp faunal community (perforate foraminifera and red algae)
This part of the ramp is colonized by larger foram red coralline algae (Buxton and Pedley 1989). In this division, larger perforate foraminifera (Nummulites, Operculines and Discocyclines), smaller foraminiferal perforate tests (e.g., rotaliids), red algae and echinoids are the principal faunal elements (Figs. 11 and 12B, I, N and V). The fossil content flourished in Nummulites bank and its associated lithofacies in the middle ramp (Figs. 11 and 12) and form the Samalut Formation. For the time being, coralline algae thrive in a vast extent of trophic cases (oligotrophic environments, Borowitzka and Larkum 1986); mesotrophic waters in different latitudes, Adey and MacIntyre 1973). According to the depth of the water, the shallow (proximal) middle ramp includes an association dominated by robust Nummulites with coralline algae (Fig. 12). The deep (distal) middle ramp is distinguished by a growing in the size and flatness of the perforate foraminiferal tests accompanied with an expansion of Bryozoa and echinoids. Accompanying coralline algae and LBF signal accumulation of the Samalut Formation in tropical to subtropical warm water conditions under oligotrophic to slightly mesotrophic conditions, but the lack of planktic forams, even in the deepest parts, indicates low nutrient supply and low productivity (Fig. 12).
Outer ramp faunal community (planktic foraminifera, bivalves and ostracods)
It dominates the northern parts and forms the Maghagha Formation (Nile Valley) and Darat Formation (Sinai) (Figs. 11 and 12), where deeper marine conditions prevailed compared with that in the south. Generally, it includes gray to greenish shales alternated with mudstones rich in planktic forams with ostracods, disseminated calcareous benthic fauna, thin-shelled gastropods and skeletal fragments of bivalves. The deficiency in body and trace fossils proposes that the outer ramp facies was accumulated as hemipelagic sediment in quiet calm deep water, in open marine conditions, below the zone of current attack (Buxton and Pedley 1989). Similar faunal elements have been recorded by Amirshahkarami et al. (2007) in southwest Iran (Asmari Formation).
Sequence stratigraphic approach
The investigated sequence (50–75 m thick) corresponds nearly to the Ypresian and Lutetian time interval (Figs. 6, 7, 8, 9, 10, 11) and exhibits four depositional sequences of third order (Figs. 13 and 14), each of which shows retrogradational transgressive systems tract (TST) and aggradational highstand systems tract (HST) to progradational lowstand systems tract (LST) packages of facies.

Representative log for the studied Eocene rocks (Nile Valley) showing facies characteristics, sequence stratigraphy and sea-level changes compared with Haq et al. (1987) eustatic curves

Representative log for the studied Eocene rocks (west-central Sinai) showing facies characteristics, sequence stratigraphy and sea-level changes compared with Haq et al. (1987) eustatic curves
Sequence boundaries
Two prominent diachronous sequence boundaries (Figs. 13, 14 and 15) have been specified (SB1 and SB2). They have been marked as significant erosional hiatuses where paleosol beds (Fig. 15) exist and are considered as a result of sea-level drop that erodes the subaerially exposed sediment surface of the earlier sequences at the top of Minia and Darat formations (Figs. 5A and B).

Paleosol horizon between Minia and Samalut formations. A X-ray diffraction identified five minerals in the paleosol. Note that calcite is first abundant mineral, followed by dolomite. Nile Valley sector. B Field photograph illustrating the calcareous shale to marl (paleosol horizon) separating between Minia and Samalut formations (yellow arrow)
Late Ypresian–Early Lutetian sequence boundary (SB1)
This boundary has been marked at the contact between Minia and Samalut formations and represents the Ypresian–Lutetian time line (Figs. 5A and 13). It outcrops as erosional and irregular surface that terminated sedimentation of the Minia Formation and heralded the start of Samalut Formation. According to Haq et al. (1987) curve, there was a major sea-level regression and subaerial exposure at the Early–Middle Eocene boundary which corresponds to their TB2.9/TB3.1 cycle boundary (Fig. 13). Khalifa et al. (1993) located this boundary at the top of white limestone with siliceous concretions at the upper surface of the Minia Formation near the inlet of Wadi El Muweilih, west of the Nile Valley. The trait of this boundary varied laterally from one locality to the other in the Nile Valley. Due south, it has been delineated as irregular hardground stained with iron oxide and intensively bioturbated around Abu Qurqas section (Fig. 16A). Moving north, it has been marked as paleosol horizon (Figs. 5A and 15) overlying an irregular surface around both Wadi Tarfa and Wadi Tihna sections.

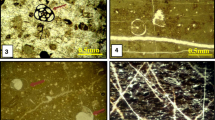
A Field photo shows Late Ypresian–Early Lutetian sequence boundary (SB1) in the Nile Valley at the contact between the top of the Minia Formation and the base of the overlying Samalut Formation. The boundary is irregular hardground streaked with iron oxide and intensively bioturbated. Nile Valley. B Middle Lutetian-Late Lutetian sequence boundary (SB2) in Sinai at the contact between the top of the Darat Formation and the basal part of the Samalut Formation as sharp erosional and bioturbated surface. Thin pebbly, discontinuous paleosol layer plastered on the sequence boundary. West-central Sinai. C Photomicrograph showing imperforate foraminifera floatstone of restricted lagoon at the base of TST of depositional sequence (SD1). Fauna include Orbitolites, Alveolines, Milliolides and other bioclasts. Minia Formation, Nile Valley. D Photomicrograph exhibiting algal packstone to floatstone microfacies of back bank environment at the top of HST of DS1. Pronounced fragmentation and spotty staining with iron oxide point to relative fall of sea level and start formation of sequence boundary (SB1). Nile Valley. E Photomicrograph illustrating perforate larger foraminifera (medium and large-sized Nummulites) with Bryozoa constitute float to packstone microfacies that signals maximum rise of relative sea level. Nile Valley. F Photomicrograph showing aggradational facies of HST of DS2. This facies is marked by increasing dominance of larger forams in coarser-grained, partially fragmented rudstone besides floatstone toward the top. Wadi Feiran section, Sinai. G Photomicrograph exhibiting encrustation of Nummulite shells by coralline algae (yellow arrow). Note a general increase of coarser-grained float- and rudstone besides boundstone denoting shallowing upward. The upper divide of the Samalut Formation (Nile Valley). H Photomicrograph showing bioturbated, creamy marly limestone (mud- to wackestone), which shows evidences of dolomitization indicating shallowing conditions followed by short-term hiatus (SB2) and represents HST deposits of DS3. Wadi Feiran, Sinai. I Photomicrograph illustrating bioturbated, bioclastic pelecypodal operculines wacke- to packstone microfacies of back-bank or lagoonal origin at the base of TST of DS4. Clasticity is relatively high. Wadi Feiran, Sinai. J Photomicrograph illustrating bryozoa Nummulites float- to packstone microfacies of Nummulites bank. Some tests were encrusted with coralline algae (now micritized) and filled partially with micrite. TST of DS4. Nile Valley. K Field photo displaying increase dominance of larger forams at the upper parts of the Samalut Formation (Nile Valley). This increase points to shallowing conditions. Wadi Tihna section. L Field photo displaying increase dominance of larger forams at the upper parts of the Samalut Formation in Wadi Feiran section, Sinai. Larger B-forms predominate over smaller A-forms. West-central Sinai
Middle Lutetian–Late Lutetian sequence boundary (SB2)
It is only recorded in Sinai at the formational boundary between Darat and Samalut formations as sharp erosional surface (Figs. 14 and 16B). The bed overlying this surface consists of ill-sorted, bioturbated pack- to floatstone (Fig. 16B, photo inset). On the sole of this bed, preserved horizontal, Y-shaped burrows represent the start of a transgression over the preceding rocks. Therefore, the depositional surface, accumulated upon the Samalut Formation, is regarded here as composite erosive surface (sequence boundary as well as transgressive surface). Thin pebbly, discontinuous paleosol horizon with argillaceous matrix plastered directly on the sequence boundary (Fig. 16B). Herein, this horizon is considered as lowstand deposits (Fig. 14). The overlying tabular plane beds of the Nummulites bank (Fig. 16B) have been considered as retrograding topsets across the Lutetian ramp. Benjamini (1984) stated that regressive facies developed initially during the sedimentation of the upper parts of Middle Lutetian in the Eocene of the Arava Valley (Negev, Israel). This regressive facies was succeeded by a regional hiatus nearly at the Middle and Late Lutetian boundary. Environmentally, the sudden arrival of nummulitic limestones at Wadi Feiran (Sinai) signals a significant increase in sedimentation rate, which is probably compatible with a major sea-level drop.
Depositional sequences
Sequentially, four depositional sequences (DS1–DS4), formed as echo for eustatic sea-level changes, have been delineated (Figs. 13 and 14). Drop of sea level characterizes the LST, deepening on trends considered as TST and shallowing trends indicate HST.
Depositional sequence (DS1)
It builds the upper divide of the Minia Formation (Nile Valley), nearly 22 m thick (Fig. 13). Its base is unexposed and bounded by SB1 at the top. It begins with imperforate forams floatstone of restricted inner lagoon (Fig. 16C), which is followed by retrogradational facies comprising a bioclastic imperforate foraminiferal grainstone of shoal bar origin as TST. The maximum flooding surface (MFS) differentiates between the TST (below) and the HST (above) as well as points to a variation from retrogradational to progradational parasequence stacking patterns. It is composed of thin bioturbated horizon, encrusted by iron oxide coloration with stained fragmented fauna. The HST starts with porcelaneous forams floatstone (restricted lagoon), followed by cross-bedded limestone (shoals) and topped by algal floatstone of back bank (Fig. 16D). Termination of this sequence with algal Nummulite packstone of back-bank facies points to relative of sea-level drop and initiation of sequence boundary (SB1) that represents the formational boundary between Samalut and Minia formations.
Depositional sequence (DS2)
The Nile Valley tops unconformably the Minia Formation (50 m thick). It is bounded at the base by the SB1 and topped by the present-day erosion surface. Three system tracts including LST, TST and HST have been known. The LST represents a progradational facies and appears as calcareous paleosol horizon of 20–30 cm thick, plastered directly over the SB1, which signals a regression in the relative sea level (Fig. 15B). X-ray analysis identifies five minerals in the paleosol: calcite, dolomite, iron oxides (hematite), quartz and clay minerals with calcite as a first abundant mineral, followed by dolomite (Fig. 15A). The calcite may be inherited or have an authigenic origin of formation. Allen and Hajek (1989) stated that a few calcretes contain more than 900 g–1 kg of calcite. As a fact, the transition from Early Eocene (Ypresian) to Middle Eocene (Lutetian) herein is clearly associated with a sea-level drop (Haq et al. 1987). This fall in sea level (Fig. 13) could lead to increase exposures of marine limestone and, in turn, increase in calcium available for calci-sol development. The source of quartz is probably windblown, either from quartz grains within the carbonate rocks or from extra provinces.
The TST is distinguished by a retrogradational facies (nearly 30 m thick) and recognized where there is a relative rise in sea level. It mainly consists of cyclic alternations of Nummulites bank and its back-bank facies and closed by fore-bank facies (Fig. 13). Perforated larger foraminifera with bryozoa constitute float- to packstone microfacies (Fig. 16E). These facies indicate the maximum relative rises in sea level. The HST is recognized as aggradational facies and marked by increasing dominance of larger forams in coarser-grained rudstone (Fig. 16F) besides floatstone toward the top (Fig. 13). At certain levels, test encrustation by coralline algae is noticeable giving rise to boundstone (Fig. 16G). By its position, this systems tract points to a highstand above the sea-level rise of the TST facies. On a regional scale, it is noticed that, the upper beds of the Samalut Formation in Sinai and the Nile Valley exhibit a general increase of larger foraminifera, in coarser-grained float- and rudstone besides boundstone (Figs. 13 and 14). This signals an overall shallowing; that is a Highstand (or regressive) Systems Tract.
Depositional sequence (DS3)
In Wadi Feiran (Sinai), this sequence builds the upper beds of the Darat Formation and it is not completely developed. Its basal divide is buried and unexposed and the upper part (9 m thick) may be partially eroded during the formation of SB2. It is topped by (SB2) that coincides with the time boundary between Middle and Late Lutetian and represented mainly by HST (Fig. 14). Predominant rock varieties are bioturbated, creamy marly limestone (mud- to wackestone) and pale green calcareous shale, which show evidences of dolomitization near the SB2 (Fig. 16H). Existence of dolomite above open marine facies of the Darat Formation has been interpreted as indication for shallowing conditions followed by short-term hiatus (SB2) and represents HST deposits formed by normal regression coupled with a sea-level highstand (Fig. 14). A highstand normal regression exists when the sediment supply rate, to a given shoreline, exceeds the rate of accommodation space (Posamentier et al. 1992). This sequence is ended with a drop of short-term in relative sea-level that led to emersion and initiated subaerial diagenetic processes.
Depositional sequence (DS4)
This sequence is represented by Samalut Formation in Sinai (Wadi Feiran). Stratigraphically, it rests on the unconformity surface (SB2) and coincides with the time line between late Middle Lutetian and early Late Lutetian (Fig. 14). Overall, its residual thickness measures nearly 39 m thick and topped by the present-day erosion surface. Sequentially, it could be distinguished to three distinctive systems tracts, namely LST, TST and HST.
The LST is a progradational sediment represented by a paleosol horizon (10–15 cm thick) with lithoclasts overriding erosive surface that points to a relative sea level fall (Figs. 14 and 16B). The following TST is characterized by a retrogradational facies (nearly 27 m thick) and recognized where there is a rise in relative sea level. Facies association starts with bioclastic pelecypodal operculines wacke- to packstone of back-bank or lagoonal origin (Fig. 16I), which is covered by Bryozoa Nummulite float to packstone of Nummulites bank origin (Fig. 16J). Perforated larger foraminifera represented by Nummulites and Operculines are dominant in large quantities. Catuneanu (2002) stated that the presence of shallow marine deposits above subaerial unconformity surface indicates a transgressive surface of erosion (Van Wagoner et al. 1988) or a ravinement surface. The HST is not completely preserved due to erosion (residual thickness 12 m). In the goal area, it is marked by a general shallowing with increasing dominance of larger forams (Figs. 14, 16K and L). Ecologically, the shallowing-up depositional sequence could reflect the impact of different environmental variables such as ecology, biological evolution and hydrodynamic processes.
Reservoir potentiality of the Nummulites bank in the focus area: general discussion
The hydrocarbon potential of Eocene carbonate sedimentary rocks yields about 20% worldwide petroleum resources, but constitutes more than 50% (Jorry 2004). The Eocene nummulitic limestone in Tunisia (Hasdrubal field) has an average porosity and permeability of 10.5% and 0.5 md, respectively (Racey 2001).
In Oman, the Nummulite buildups possess a porosity and permeability range between 0.7 and 14% and 0.95 md, respectively, in spite of their attack by post-depositional diagenetic processes (Racey 2001). Furthermore, the majority of the Nummulite tests are fragmented, reworked and transported by high-energy events. Adversely, the primary interparticle porosity, in the Ashtart field, is drastically occluded by calcite cements (Hmidi and Sadras 1991), which is quiet similar to some nummulitic horizons in the present work. The nummulitic limestone of the Ashtart field possesses a high intergranular porosity averaging 15%, but low permeability averaging 6 md within the Nummulite shells. The Nummulite accumulations in Libya are equivalent in time with those of Tunisia and have an average of porosity up to 16% (El Ghoul 1991). Added to that, similar Nummulite bodies have been deeply studied in Italy, former Yugoslavia and Egypt. Unfortunately, the available data about their reservoir potentiality are lacking (Racey 2001).
By comparison, it seems that, similar to the above-mentioned Nummulite banks, the carbonate ramp under consideration has a considerable amount of Nummulite accumulations. These accumulations display extensive impacts of fragmentation and reworking by high-energetic events (Fig. 16F and I). Accordingly, the present work could be used as analog study for other nummulitic limestones or Nummulite accumulations. Unfortunately, the present work might not be presented as a good analog because of some limitations such as limited scale of the target area and the sealing potential of the interbedded matrix-supported textures (mudstones, wackestones and even floatstones, Figs. 6, 7, 8, 9). Added to that, the dominant fine-grained matrix (i.e., micrite and/or Nummulite hash) could enter into the test interior cavities through the surface pores of shells that could drastically diminish the intraparticle porosity.
Taking into consideration the reservoir potentiality of the goal area, the total stratigraphic record of Nummulite banks of the Samalut Formation could be categorized into two pronounced parts (Fig. 5A and B) relying on the ratio of grains to matrix (grain supported or matrix supported, Figs. 6, 7, 8, 9). The architectural elements and facies of the lower part are entirely dominated by massive to burrowed mudstone to wackestone in the Nile Valley sector and argillaceous, soft, bioclastic packstone in Sinai. It shows a gradual increase in grain size upwards (Figs. 6 and 7) and accumulated under back-bank to lagoonal conditions. Rare Nummulite tests of coarser sizes were scattered randomly, and therefore, the deposit attains relatively a homogeneous characteristics. Even though this unit of fine-grained deposits is expected to have had a good porosity during deposition. As usual and by the drastic action of compaction, it is believed that this porosity have been lost during burial. Furthermore, since the deposits have fine-grain sizes (i.e., silt and clay), even if it could still preserve a certain porosity, the expected permeability would be very low (i.e., it could work as aquiclude), as the pores would be too small to permit possible fluids to go through them. Accordingly, this fine-grained part, in a general meaning, lacks any reservoir potential, especially for oil.
Moving upsection, facies forming the upper portion (Figs. 6 and 7), which attains a retrogradational pattern, shows a very heterogeneous reservoir characteristic. Its upward coarsening trend contains interbeds of mudstone and Nummulite-rich textures of coarser grain sizes, which have variable stacking pattern. The grain size of the deposits, at the base of this unit, varies from fine to medium, with thin interbedded mudstone beds that separate the Nummulite-rich coarser beds (Figs. 6 and 7). This kind of alternation leads to a poor vertical connectedness for the deposits and may also create possible flow discontinuities among the different beds. The Nummulite-rich beds are also observed, in most outcrops, to die out laterally. This could also limit the lateral interconnection. Only in some localities, the beds appear to be amalgamated (multi-event beds). At upper levels, the few beds that show moderate lateral continuity are also separated by thin mudstone beds. These beds may possibly work as permeability barriers that would form pronounced reservoir heterogeneity in the reservoir. As a general role, due to the lack of connectedness and their very fine grain size, the deposits of the lower division (Figs. 6, 7, 8, 9) altogether are expected to have poor reservoir quality.
Near the upper half of the Samalut Formation, the relatively coarsening upward trend, shoaling and the increase in Nummulite content (Figs. 6, 7 and 8) may point to a relative increase of the reservoir potential. However, the predominance of Nummulite tests could support the reservoir potential. On one side, many of the Nummulite shells are mostly fragmented (Fig. 16F and I), whereas the complete tests are bounded together by micritized coralline algae (i.e., boundstone textures, Fig. 16G and J), which negatively affect the interparticle permeability and porosity. On the contrary, the intact tests are filled with intraparticle fine-grained materials (micrite, iron oxides or clays) and minerals like calcite and even silica (Fig. 16G and J), which would have reduced the reservoir potential expected from the volume of Nummulites alone.
Reservoir type, quality and characteristics in the Nummulite bodies under study are a function of grain types, pore types, grain size, sorting, cementation and compaction, although predicting the location of high reservoir potential facies is hard to identify; exploration procedures should be directed to the main trend of the mid ramp where the nummulite accumulations are located.
Summing up, it is concluded that the fine- grain size of the deposits, the shortage of good connectedness in spatio-temporal scales, the filling of the shells of the Nummulites by other minerals and bounding together by coralline algae could result in low reservoir potential. Add to that, the initial porosity is expected to have been lost by the impact of the compaction and diagenetic effects (burial cementation). Here, the biostratigraphy (especially Nummulites) and its general effect on the porosity distribution is not well known but other studies in different parts of the Tethyan region (i.e., Tunisia and Libya) demonstrated that Nummulite accumulations can form good reservoirs for both gas and oil. The overall evaluation of the reservoir potentiality in the target area, the impact of diagenesis, which could have further adversely affected the porosity and thus the reservoir potential of the deposits, has not been seriously considered.
Summary and conclusion
Four rock units have been recognized in the goal area (Minia Formation of Late Ypresian age, Darat Formation of Early Lutetian age, Samalut Formation of Early to Middle Lutetian age and Maghagha Formation of Middle to Late Lutetian age). These units were deposited on a gentle-dipping carbonate ramp and eight facies belts have been identified based on their composition and dominant sedimentary structures. The depositional model exhibits that the facies associations point to a gradual shallowing upward. Ecologically, the distributions of LBF are confined to tropical and subtropical settings and this distribution is determined by a complex set of interrelated parameters (e.g., light, temperature, salinity, nutrients, depth, hydrodynamic energy and substrate). The majority of the LBF flourishes in the subtropical to tropical water conditions and trapped in temperatures of 18–25 °C. Herein, the combination of Peneroplis and Alveolines with red coralline algae and coral signals temperature values from 18 to 22 °C in tropical environment, while planktic forams can survive in less than 14 °C temperature. Accordingly, the studied Eocene sediments probably had a tropical–subtropical climate because of the extensive occurrence of benthic foraminifera, algae, corals and Bryozoa. Considering the water salinity, it could be noted that the higher levels of Samalut and Maghagha formations accumulated seawater of normal salinity, but the middle and lower levels (i.e., Minia Formation) suffered from sporadic higher to normal salinity water circumstances. The community of larger forams with coral and red coralline algae herein indicates meso- to oligotrophic conditions, while planktic forams indicate surface trophic to intermediate mesotrophic water conditions. Porcelaneous forams occur in substrate of low stability in more shallow milieus, while Operculines colonize soft sediment. Generally, the biota in the back-bank environment indicates muddy substrate (burrowing bivalves and echinoids). Thick-shelled and conical forms of LBF thrive on coarse-grained seabed, but flattened shell forms prefer soft substrates. Near the base of the studied sequence, Peneroplis and Miliolides (i.e., imperforate forams) inhibited a restricted shallower water depth, while in the middle and upper parts, Nummulites, Operculines, and Discocyclines (i.e., perforate foraminifera) are abundant reflecting open marine environment. On the contrary, planktic forams indicate water depth environment ˃200 m (open marine outer ramp). Sequentially, the studied succession has been classified to 4 superimposed depositional sequences (DS1–DS4) separated by two prominent diachronous sequence boundaries (SB1–SB2). The retrogradational facies (TST) are represented by the outer and inner lagoon packages; the aggradational one (HST) is exemplified by the tidal flat and shoal bar facies; but the progradational facies (HST) is represented by the presence of paleosol horizons overlying sequence boundaries. The sequence boundaries have been marked as significant erosional hiatuses where paleosol beds exist.
Many factors affect adversely its reservoir potentiality of the studied sequence such as the fine- grain size of the deposits, the lack of good connectedness, the filling of the Nummulite tests by minerals and bounding together by coralline algae. The pronounced spatial and temporal changes in the depositional settings and the occurrence of many hiatuses in the investigated rocks suggest that the sedimentary style has been impacted strongly by the tectonic activity rejuvenated episodically along the Syrian Arc Fold Belt. Both internal and external controls have played a major role in controlling the in situ carbonate production and the observed architectural style and the sequence bounding surfaces.
Data availability
All data generated or analyzed during this study have been completely included, inserted and used in this published article for description, analysis and interpretation of the obtained results.
References
Adey WH, MacIntyre IG (1973) Crustose coralline algae: a re-evaluation in the geological sciences. Geol Soc Am Bull 84:883–904
Allen B, Hajek B (1989) Mineral occurrence in soil environments. In: Dixon J, Weed S (eds) Minerals in soil environments. Soil Science Society of America, Madison, Wisconsin, pp 199–278
Amirshahkarami M, Vaziri-Moghaddam H, Teheri A (2007) Sedimentary facies and sequence stratigraphy of the Asmari Formation at Chaman-Bolbol, Zagros Basin, Iran. J Asian Earth Sci 29(5):947–959
Anketell JM, Mriheel IY (2000) Depositional environment and diagenesis of the Eocene Jdeir Formation, Gabes-Tripoli Basin, Western Offshore, Libya. J Pet Geol 23:425–447
Bandel K, Kuss J (1987) Depositional environment of the pre-rift sediments, Galala heights, Gulf of Suez, Egypt. Berliner Geowissenschaften Abhandlungen 78:1–48
Barattolo F, Bassi D, Romero R (2007) Upper Eocene larger foraminiferal-coralline algal facies from the Klokova Mountain (south continental Greece). Facies 53:361–375
Bassi D, Hottinger L, Nebelsick JH (2007) Larger foraminifera from the Late Oligocene of the Venetian area, north-eastern Italy. Palaeontology 50:845–868
Bassi P, Iryu Y, Braga J, Takayanagi H (2013) Bathymetric distribution of Ichnocoenosis from recent subtropical algal nodules off Fraser Island, eastern Australia. Paleo-3 369:58–66
Beavington-Penney SJ, Wright VP, Woelkerling W (2004) Recognizing macrophyte-vegetated environments in the rock record: a new criterion using ‘hooked’ forms of crustose coralline red algae. Sed Geol 166:1–9
Bebout DG, Pendexter C (1975) Secondary carbonate porosity as related to early tertiary depositional facies, Zelten field, Libya. Am Asso Petrol Geol Bull 59(4):665–693
Benjamini C (1984) Stratigraphy of the Eocene of the Arava Valley (eastern and southern Negev, southern Israel). Isr J Earth Sci 33:167–177
Bignot G, Strougo A (2002) Middle Eocene benthic foraminiferal assemblages from eastern Egypt, as biochronological and peritethyan lagoonal indicators. Rev Micropaléontol 45(2):73–98
Bishay Y (1961) Biostratigraphic study of the Eocene in the Eastern Desert between Samalut and Assiut by the larger foraminifera. 3rd Arab Petrol Congr Alexandria 2, p. 13
Bishay Y (1966) Studies on the larger foraminifera of the Eocene (the Nile Valley between Assiut and Cairo and SW Sinai). PhD Thesis, Alexandria University, Alexandria, Egypt
Borowitzka M, Larkum A (1986) Calcification in algae: mechanisms and the role of metabolism. Crit Rev Plant Sci 6(1):1–45
Bosence D, Pedley H (1979) Paleoecology of a Miocene coralline algal bioherm, Malta. Bull Cent Rech Explor Prod Elf-Aquitaine 3(1979):463–470
Boudagher-Fadel M (2008) Evolution and geological significance of larger benthic foraminifera. In: Wignall P (ed) Developments in paleontology and stratigraphy. Elsevier, Amsterdam, p 540
BouDagher-Fadel M (2013) Biostratigraphic and geological significance of planktonic foraminifera, 2nd edn. Office of the Vice Provost (Research), UCL, London, UK, p 287
Boukhary M, Abdel-Malik W (1983) Revision of the stratigraphy of the Eocene deposits of Egypt. N Jb Geol Paleont Mh 68(1983):321–337
Brandano M, Corda L (2002) Nutrients, sea level and tectonics: constrains for the facies architecture of a Miocene carbonate ramp in central Italy. Terra Nova 14:257–262
Brandano M, Frezza V, Tomassetti L, Pedley M, Matteucci R (2009a) Facies analysis and paleoenvironmental interpretation of the Late Oligocene Attard Member (Lower Coralline limestone formation), Malta. Sedimentology 56(4):1138–1158
Brandano M, Frezza V, Tomassetti L, Cuffaro M (2009b) Heterozoan carbonates in oligotrophic tropical waters, The Attard member of the lower coralline limestone formation (Upper Oligocene, Malta). Palaeo 274:54–63
Brasier M (1975) Ecology of recent sediment-dwelling and phytal foraminifera from the lagoons of Barbuda, West Indies. Jou Foram Res 5:42–46
Brioussov BA (1968) Tectonic division and basement relief in northern United Arab Republic (Egypt) according to gravity and magnetic data (Abstract). The geological society of Egypt, 6th annual meeting 3–9 May, 1968
Buxton MW, Pedley MH (1989) A standard model for Tethyan tertiary ramps. J Geol Soc Lond 146:746–748
Catuneanu O (2002) Sequence stratigraphy of clastic systems: concepts, merts and pitfalls. J Afr Earth Sci 35:1–43
Chaproniere C (1975) Paleoecology of Oligo-Miocene larger Foraminiferida, Australia. Alcheringa 1:37–58
Cosovic V, Drobne K, Moro A (2004) Paleoenvironmental model for Eocene foraminiferal limestones of the Adriatic carbonate platform (Istrian Peninsula). Facies 50(1):61–75
Dickson JA (1965) Carbonate identification and genesis as revealed by staining. J Sed Petrol 36:491–505
Dunham R (1962) Classification of carbonate rocks according to depositional texture. In: Ham W (ed) Classification of carbonate rocks. AAPG Bull, Tulsa, pp 108–121
Duxbury A, Duxbury A (1997) An Introduction to the world’s Oceans, 5th edn. WC Brown Publishers, Dubuque, p 504
El Ayyat AM, Obaidalla NA (2016) The impact of the Syrian arc orogeny on the early Paleogene rocks, western shoulder of the Gulf of Suez, Egypt. Paleo-3 454:30–53
El-Ghoul A (1991) A modified Farwah Group type section and its application to understanding stratigraphy and sedimentation along an E-W section through NC 35A, Sabratah Basin. Third Symposium on the Geology of Libya (4),Elsevier, Amsterdam 1637–1656
El Hinnawi EM, Samy Y, Zahran I (2003) Textural and mineralogical characteristics of coastal sediments along the Gulf of Aqaba (Abstract) 4th conf. Geol of Sinai for Development, Sinai (1995), (Ismailia)
Embry A, Klovan J (1971) Absolute water depth limits of Late Devonian paleoecological zones. Geol Rundsch 61:672–686
Flügel E (1982) Microfacies analysis of limestones. Springer, Berlin, p 633
Flügel E (2004) Microfacies of carbonate rocks: analysis, interpretation and application. Springer, Berlin Heidelberg, New York, p 976
Flügel E (2010) Microfacies of carbonate rocks. Springer, Berlin, p 984
Gallardo A, Serra-Kiel J, Ferrandez-Canadell C, Ph Roger J, Boix C, Caus E (2001) Macroforaminiferos porcelanados del Eoceno Superior-Oligoceno Inferior del Dhofar (Sultanato de Omán). 83–89. In: Melendez G, Herrera Z, Delevene G, Azanzazanza B (eds) Los fósiles y la paleogeografía. Actas de las XVII Jornadas de la Sociedad Española de Paleontología, Albarracín (Teruel), 5 (1). Publicaciones del Seminario de Paleontología de Zaragoza (SEPAZ), p 360
Geel T (2000) Recognition of stratigraphic sequences in carbonate platform and slope deposits: empirical models based on microfacies analysis of Paleogene deposits in southeastern Spain. Palaeo-3 155:211–238
Halfar J, Godinez-Orta L, Mutti M, Valdez-Holguin J, Borges J (2004) Nutrient and temperature controls on modern carbonate production: an example from the Gulf of California, Mexico. Geology 32:213–216
Hallock P (1985) Why are larger foraminifera large? Paleobiology 11:195–208
Hallock P (1987) Fluctuations in the trophic resource continuum: a factor in global diversity cycles. Paleo-3 2(5):457–471
Haq B, Hardenbol J, Vail P (1987) Chronology of fluctuating sea-levels since the Triassic. Science 235:1156–1166
El Heiny I, Morsi S (1986) Review of the Upper Eocene deposits in the Gulf of Suez, Egypt. Proceedings of the 8th Exploration Seminar, Cairo, 1986. Egyptian General Petroleum Corporation 1:117–130
Hermina M, Lindenberg H (1989) The Paleogene. In: Hermina M, Klitzsch E, List F (eds) Stratigraphic Lexicon and explanatory notes to the geological map of Egypt, 1:500,000. CCONOCO and EGPC, Cairo, pp 144–161
Hewaidy AA, El-Motaal EA, Sultan SA, Ramdan TA, El Khafif AA, Soliman SA (2015) Groundwater exploration using resistivity and magnetic data at the northwestern part of the Gulf of Suez, Egypt. Egypt J Pet 24(3):255–263
Hmidi Z, Sadras W (1991) The Ashtart Field. Tunisia Exploration Review (ETAP) 4:1–107
Hohenegger J (2000) Coenoclines of larger foraminifera. Micropaleontology 46(Supplement 1):127–151
Hohenegger J (2004) Depth coenoclines and environmental considerations of Western Pacific larger foraminifera. J Foram Res 34:9–33
Hottinger L (1997) “Foraminifera operculiniformes”. Memoires du Museum National D’histoire Naturelle, Nouvelle Serie, Serie C, Science de la Terre. Tome XL. p. 151, 66 pls, 57 figs, Paris
James N, Collins L, Bone Y, Hallock P (1999) Subtropical carbonates in a temperate realm: modern sediments on the southwest Australian shelf. J Sed Res 69:1297–1321
Jorissen F, Stigter H, Widmark JG (1995) A conceptual model explaining benthic foraminiferal microhabitats. Mar Microp 26:3–15
Jorry S (2004) The Eocene nummulite carbonates (Central Tunisia and NE Libya), sedimentology, depositional environments and application to oil reservoir (Ph. D. Thesis). Université De Genéve. Départment de Géologie et de Paléontologie, p 206
Khalifa M, Shamah K, Abu El-Ghar M (1993) Mixed carbonate siliciclastic ramp facies of the middle Eocene Midawara Formation, southeast of El Fayoum area, Western Desert, Egypt. Ann Geo Sur Egypt 19:335–348
Kiessling W, Flugel E, Golonka J (2003) Patterns of Phanerozoic carbonate platform sedimentation. Lethaia 36:195–226
Kulbrok F (1996) Biostratigraphie, Fazies und Sequenzstratigraphie einer Karbonaterampe in den Schichten der Oberkreide und des Alttertiars Nordost Agyptens (eastern Desert, N Golf von Suez, Sinai). Berrichte Fachbereich Geowiss Universität Bremen 81:1–216
Kumar A, Saraswati B (1997) Response of larger foraminifera to mixed carbonate-siliciclastic environments: an example from the Oligocene-Miocene sequence of Kutch, India. Paleo-3 136:53–65
Langer M, Hottinger L (2000) Biogeography of selected larger foraminifera. Micropaleontology 46(Supplement 1):105–126
Leckie R, Olson H (2003) Foraminifera as proxies for sea-level change on siliciclastic margins. SEPM Special Publication 75:1–19
Lee J (1990) Phylum Granuloreticulosa (Foraminifera). In: Margulis L, Corliss J, Melkonian M, Chapman D (eds) Handbook of Protoctista. Jones and Bartlett Publishers, Boston, pp 524–567
Li X, Liu W, Wang Y, Zhu Q (1999) The tectonic evolution and metallogenesis in Tethys of the Nujiang-Langcangjing-Jingsajing area, southwestern China. Geological Publishing House, Beijing, pp 1–276 (in Chinese)
Mansour H, Philobbos E (1983) Lithostratigraphic classification of the Eocene surface carbonates of the Nile Valley, Egypt: a review. Bull Fac Sci Assiut Univ 6(2):293–306
Mossadegh Z, Haig D, Allan T, Adabi M, Sadeghi A (2009) Salinity changes during Late Oligocene to Early Miocene Asmari Formation deposition, Zagros Mountains, Iran. Paleo-3 272:17–36
Murray J (2006) Ecology and applications of benthic foraminifera. Cambridge University Press, Cambridge, p 426
Omara S, Mansour H, Youssef M, Khalifa H (1978) Stratigraphy, paleoenvironment and structural features of the area east of Beni Mazar, Upper Egypt. Bull Fac Sci, Assiut Univ 3(5):23–54
Pecheux M (1995) Ecomorphology of recent large foraminifera, Operculina ammonoides. Geobios 28:529–566
Pedley HM (1996) Miocene reefs facies of the Pelagian region (central Mediterranean). In: Franseen EK, Esteban M, Ward WC, Rouchy J (eds) Models for carbonate stratigraphy from Miocene reef complexes of Mediterranean region. SEPM concepts in sedimentology and paleontology, Tulsa, pp 247–259
Philobbos ER, Keheila EA (1979) Depositional environment of the middle Eocene in the area southeast of Minia, Egypt. Ann Geol Surv Egypt 9:523–550
Pomar L (2001) Ecological control of sedimentary accommodation: Evolution from a carbonate ramp to rimmed shelf, Upper Miocene, Balearic Islands. Paleo 75:249–272
Posamentier HW, Allen GP, James DP, Tesson M (1992) Forced regressions in a sequence stratigraphic framework: concepts, examples, and exploration significance. AAPG Bulletin 76(1992):1687–1709
Racey A (2001) A review of Eocene Nummulite accumulations: structure, formation and reservoir potential. Jou Pet Geo 24:79–100
Racey A, Bailey H, Beckett L, Gallagher M, Hampton M, McQuilken J (2001) The petroleum geology of the Early Eocene El Garia Formation, Hasdrubal Field, offshore Tunisia. J Pet Geo 1(24):29–35
Rasser M, Scheibner C, Mutti M (2005) A paleoenvironmental standard section for Lower Ilerdian tropical carbonate factories (Pyrenees, Spain; Corbieres, France). Facies 51:217–232
Said R (1990) The geology of Egypt. Balkema Pub Comp, Amsterdam, New York, p 734
Samankassou E (2002) Cool-water carbonates in a paleoequatorial shallow-water environment: The paradox of the Auernig cyclic sediments (Upper Pennsylvanian, Carnic Alps, Austria-Italy) and its implications. Geology 30(7):655–658
Sartorio D, Venturini S (1988) Southern Tethys biofacies. San Donato Milanese, AGIP S.p.A 235 p
Scheibner C, Speijer R, Marzouk A (2005) Turnover of larger foraminifera during the Paleocene-Eocene thermal maximum and paleoclimatic control on the evolution of platform ecosystems. Geology 33(6):493–496
Scheibner C, Rasser M, Mutti M (2007) Facies changes across the Paleocene-Eocene boundary: the Campo section (Pyrenees, Spain) revisited. Paleo-3 248:145–168
Schlager W (2005) Carbonate Sedimentology and Sequence Stratigraphy. SEPM Concepts Sedimentol Paleontol 8:200
Scotese C (2001) Atlas of Earth History, Paleogeography, vol 1. PALEOMAP Project, Arlington, Texas, p 52
Scott T, Campbell K, Rupert F, Arthur J, Missimer T, Lloyd J, Yon J, Duncan J (2001) Geologic map of the State of Florida: Florida Geological Survey Map Series 146.
Shata A (1956) Structural development of Sinai Peninsula, Egypt. Bull Inst Desert Egypte 6(2):117–157
Sirele E (2003) Foraminiferal description and biostratigraphy of the Bartonian, Priabonian and Oligocene shallow-water sediments of the southern and eastern Turkey. Revue de Paléobiologie, Genève 22(1):269–339
Tucker ME, Wright VP (1990) Carbonate sedimentology. Blackwell, Oxford, UK, p 482
Tyrrell WW (1969) Criteria useful in interpreting environments of unlike but time-equivalent carbonate units (Transill-Capitan-Lamar), Capitan Reef Complex west Texas and New Maxico. In: Friedman GM (ed) Depositional environments in carbonate rocks, a symposium. SEPM, Tulsa, pp 80–97
Van Wagoner JC, Posamentier HW, Mitchum RM, Vail PR, Sarg JF, Loutit TS, Hardenbol J (1988) An overview of sequence stratigraphy and key definitions. In: Wilgus CK, Hastings BS, Kendall CJ, Posamentier HW, Ross CA, Van Wagoner JC (eds) Sea Level Changes- An Integrated Approach, Special Publication 42, Society of Economic Paleontologists and Mineralogists (SEPM) 39–45
Vaziri-Moghaddam H, Kimiagari MA, Taheri A (2006) Depositional environment and sequence stratigraphy of the Oligo-Miocene Asmari formation in SW Iran. Facies 52(1):41–51
Viotti C, El Demerdash G (1968) Studies on Eocene sediments of Wadi Nukhul area, east coast of Gulf of Suez. Proceedings of the 3rd African Micropaleontological Colloquium, Cairo, pp 403–423
Wilson JL (1975) Carbonate facies in geologic history. Springer, Berlin, Heidelberg, New York, p 471
Wilson JL, Jordan C (1983) Middle shelf. In: Scholle PA, Bebout DG, Moore CH (eds) Carbonate depositional environments. American Association of Petroleum Geologists Bulletin, Memoir, Tulsa, pp 297–343
Wilson M, Vecsei A (2005) The apparent paradox of abundant foramol facies in low latitudes: their environmental significance and effect on platform development. Earth Sci Rev 69:133–168
Wright V, Burchette T (1996) Shallow-water carbonate environments. In: Reading H (ed) Sedimentary environments: processes, facies and stratigraphy. Black-well, Oxford, pp 325–394
Zamgni J, Mutti M, Kosir A (2008) Evolution of shallow benthic communities during Late Paleocene-Earliest Eocene transition in the northern Tethys (SW Slovenia). Facies 54:25–43
Acknowledgements
The author thanks the editor-in chief and an anonymous referee for providing very detailed, critical and constructive reviews that improved the paper significantly. Funding by Assiut University (Egypt) has been highly acknowledged.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Contributions
The paper is single-author paper The corresponding author is AMEA.
Corresponding author
Ethics declarations
Conflict of interest
The author declares that this study was performed without any commercial or financial contributions that could be considered as a potential conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ayyat, A.M.E. Paleoenvironmental reconstruction, paleoecology and sequence stratigraphy of some Nummulites buildups in Egypt. Carbonates Evaporites 37, 60 (2022). https://doi.org/10.1007/s13146-022-00807-z
Accepted:
Published:
DOI: https://doi.org/10.1007/s13146-022-00807-z