
Abstract
In this study, the influence of geographic isolation on the levels of morphological and genetic differentiation among populations of the karst forest-adapted Pilea pteridophylla (Urticaceae) was evaluated. Based on our results, there is a strong morphological differentiation among the only two know populations of Pilea pteridophylla. Both populations can be clearly differentiated based on vegetative and reproductive characters, and the observed variation is geographically structured. Moreover, our genetic data from the chloroplast genome show strong levels of population differentiation and no shared haplotypes were observed between populations. Also, our estimation of divergence times supports a long-term isolation hypothesis between populations. Based on our results, dispersal seed limitation, long-term forest isolation and topographic heterogeneity (local adaptation) are related to high levels of endemism and species turnover in the mountain karst forests of Mexico.
Similar content being viewed by others


Introduction
Tropical karst forests are considered among the most diverse biomes worldwide (Geekiyanage et al., 2019; Molina-Paniagua et al., 2023). Their huge diversity is related to their extreme soil characteristics (e.g., high levels of Ca, Mg and K, very slow rates of soil formation and absence of surface water), high heterogeneity of microhabitats (e.g., limestone outcrops, cliffs, caves and chasms) and their archipelago-like distribution (Clements et al., 2006). Thus, plant lineages that have invaded and diversified in these forests are specialized flora, with small or disjunct distribution ranges and notorious morphological innovations (Wendt, 1987). Tropical karst forests are an ideal model to study the effects of its discontinuous distribution on the morphological and genetic variation of species; however, there are relatively few phylogeographic studies focused on plants adapted to karstic soils (e.g., Zhu et al., 2022). Despite that, the results of the studies published so far are consistent in showing that allopatric speciation is one of the major evolutionary processes causing diversity in these forests (Gao et al., 2015; Tseng et al., 2019; Wang et al., 2017; Zhu et al., 2022).
In Mexico, tropical karst forests occupy 20% of its total surface (~ 391,700 km2) in the southeast, along its eastern slope, and in the Yucatán Peninsula (Espinasa-Pereña, 1990, 2007). Two main types of karst forests can be recognized in Mexico, the platform karst and the fold mountains karst (Lugo-Hubp, 1990). Each type is differentiated in terms of its geological structure and climatic characteristics (Espinasa-Pereña, 1990, 2007; Lugo-Hubp, 1990). The fold mountains karst (named hereafter as mountain karst forest sensu Molina-Paniagua et al., 2023) is mainly covered by tropical rainforests and ranges from the southwest of Tamaulipas to southern Veracruz, in the north-east portion of Oaxaca, in southern Tabasco and several regions of the state of Chiapas (Fig. 1A). Instead, platform karst is well represented, but almost restricted to the Yucatán Peninsula (Karst of the Yucatán Peninsula sensu Espinasa-Pereña, 2007). Thus, the mountain karst forests cover 2.07% of Mexico’s surface, harbouring some of the most diverse forests in the country (Molina-Paniagua et al., 2023; Fig. 1A). Recently, it was estimated that around 11,000 species of vascular plants occur within the mountain karst forests of Mexico, just over 50% of the species estimated for the country (Molina-Paniagua et al., 2023).

Mountain karst forests of Mexico and the studied species Pilea pteridophylla A. K. Monro (Urticaceae). A Geographic distribution of the Mountain karst forests of Mexico. B Individual from the Chiapas population. C Individual from the Tabasco population
As stated, allopatric speciation has been invoked as one of the main processes that sustains high diversity within the mountain karst forests of Mexico (Wendt, 1987; Pérez‐García & Meave, 2005; Pérez‐García et al., 2009). However, there are no studies in the region that properly evaluate the role of geographic isolation on the patterns of morphological and genetic differentiation of the species. The latter should be a priority; tropical karst forests of Mexico are naturally fragmented but frequently surrounded by a matrix of agricultural fields, pastures, roads and cities, so elucidating the main evolutionary processes that underpin their species diversity is critical for improved conservation actions.
Here, we studied the genetic and morphological differentiation within Pilea pteridophylla (Urticaceae), a species endemic to Mexico with a restricted distribution to two allopatric populations located in the mountain karst forests of Tabasco and Chiapas in southern Mexico. Specifically, we assessed the influence of geographic isolation on the levels of morphological and genetic differentiation among populations.
Materials and methods
Study organism
Pilea pteridophylla A. K. Monro is a calcicole herbaceous plant endemic to southern Mexico (Fig. 1B–C). The species was described based on specimens collected in Tabasco, Mexico (Monro, 1999). It is easily distinguished from the rest of the Mexican species of Pilea by its pectinate leaves, which resemble those of a fern in colour, shape and texture (Monro, 1999; Fig. 1B–C). In fact, it is the only species of Pilea with pectinate leaves known from America (Monro, 1999). The species is restricted to the tropical rain forest, and grows on karstic limestone soils. For a long time, this species was known only from specimens collected (all before the year 2000) at the type locality in the Sierra Madrigal, Tabasco, Mexico (Monro, 1999). However, Ortiz-Rodriguez et al. (2012) reported a new locality for this species in the state of Chiapas, just over 80 km away from the type locality. Those authors indicated notable morphological differences between the individuals of both populations, mainly in the size of the plants and the depth of the margin teeth, but no formal analysis was performed. Recent explorations around the type locality have failed to locate living individuals of Pilea pteridophylla, which does not mean that the species is locally extinct, but it is probably restricted to less accessible regions. Thus, a new locality in Cerro Tortuguero, Macuspana, Tabasco, was recently located; the individuals present in this locality have morphological characteristics, such as the pectinate leaves, that conform to those of the type specimen (Fig. 1C).
Both populations currently recognized for Pilea pteridophylla are part of the mountain karst forests of Mexico, and although their populations consist of several individuals (more than 100 mature plants), their distribution is extremely localized. Specifically, the individuals from Chiapas occur at altitudes above 1000 m above sea level and appear associated with a few rocky mounds at ground level, while individuals from Macuspana, Tabasco, occur at altitudes between 500 and 600 m above sea level and appear to be restricted to a single limestone walls or cave entrance.
Sample collection
Collections were made in the two populations of Pilea pteridophylla to obtain fresh samples for DNA extraction (leaves) and complete specimens for morphological differentiation analysis (Table 1). A total of 36 herbarium specimens (17 individuals from Macuspana, Tabasco, and 17 individuals from Berriozábal, Chiapas) were analysed. Of these, 30 samples were used for DNA extraction (15 samples from Macuspana, Tabasco, and 15 samples from Berriozábal, Chiapas). We also had access to two specimens of Pilea pteridophylla from the type locality, but samples for DNA extraction were not available and were only included in the morphological analyses.
Morphological differences among populations
In order to explore morphological differentiation among individuals of Pilea pteridophylla, we measured seven morphological traits (vegetative and reproductive), including those traditionally used for species delimitation (Monro, 2001; Table 2). A principal components analysis (PCA) was performed on individuals using a correlation matrix with PAST v.3.06 (Hammer et al., 2001). We used the first two principal components (PC) to evaluate differences among groups of populations through an analysis of variance performed in R (Chambers et al., 1992; R Core Team, 2021). The graphic representation of the PCA results analyses were handled with the “cowplot” and “ggpubr” packages in R (Alboukadel, 2020; Claus, 2020).
Genetic differences among populations
We extracted DNA from leaves tissue using a CTAB protocol (Cota-Sánchez et al., 2006). The trnL-trnF and trnQ-rps16 intergenic spacer regions from the chloroplast genome (cpDNA) were amplified by PCR and sequenced for the 30 individuals of Pilea pteridophylla. These markers have been screened for variability in several plant lineages at the species and population levels (e.g. Shepherd & Heenan, 2017). Since the chloroplast genome is maternally inherited in most angiosperms, cpDNA sequences may provide signals about the routes and distances of seed movement and their impacts on gene flow.
For trnL-trnF amplifications, we used the universal primers (trnL-e, trnL-f) designed by Taberlet et al. (1991), whereas for the trnQ-rps16, we used the primers (trnQ-5’, rps16 × 1) described by Shaw et al. (2007). Protocols for DNA amplification by PCR assays and for sequencing the PCR products with some minor modifications were described elsewhere (Ornelas et al., 2016). PCR products were sequenced bidirectionally using the BigDye Terminator Cycle Sequencing kit (Applied Biosystems, Foster City, CA, USA, and analysed on a 310 automated DNA sequencer (Applied Biosystems) at the INECOL’s sequencing facility, or at University of Washington High Throughput Genomics Unit, Seattle, Washington. Edited sequences were aligned manually with PhyDE v.0.9971 (Müller et al., 2010: http://www.phyde.de/). The resulting sequences are available as alignments at Ortiz-Rodriguez (2023a), and available individually in the GenBank database (trnL-F, GenBank accession numbers: PP151098–PP151127; trnQ-rps16, GenBank accession numbers: PP151128–PP151157).
We calculated haplotype number (h), haplotype diversity (Hd) and nucleotide diversity (π) for both Tabasco and Chiapas localities using DnaSP v.5.10 (Librado & Rozas, 2009). Since both locations have the same number of individuals (15), estimates of genetic diversity are probably not influenced by sample size. To assess genetic differentiation, we considered each locality as a population, which is a simple way to investigate the spatial distribution of genetic variation. Thus, Wright’s (1965) fixation index was calculated in the Arlequin v.3.5.1.2 program (Excoffier et al., 2005) with 16,000 replicates and a 0.05 level of significance. This index (pairwise FST-values) has values between 1 and zero, where 1 is indicative of strong population differentiation, that is, presence of genetic structure and without gene flow between populations (genetic isolation), while a value of zero indicates no genetic differentiation between populations.
Relationships among P. pteridophylla haplotypes are presented as statistical parsimony networks generated using the program TCS v.1.2.1 (Clement et al., 2000), with gaps treated independently as missing data and as fifth state, and a 95% connection probability limit. Loops were resolved following the criteria given by Pfenninger and Posada (2002). We report herein results for each marker independently and for the combined trnL-trnF and trnQ-rps16 data set.
Phylogenetic relationships and divergence time estimation
Divergence times were estimated under a Bayesian approach as implemented in BEAST v.1.8.1 (Drummond & Rambaut, 2007) using the trnL-F dataset. We include various representatives of Pilea and related genera of Urticaceae as available in the supplementary material of Fu et al. (2022). The 76-accession dataset (available at Ortiz-Rodriguez, 2023b) was prepared for BEAST using BEAUti with the optimum model setting determined by the results of jModelTest (Darriba et al., 2012).
As our DNA database contains a mix of inter- and intraspecies sampling, we used pure-birth, birth–death and constant-size coalescent tree priors as models for dating estimates (Ritchie et al., 2017). The first two are designed for species-level data (speciation-based tree priors) and the last one is most suitable for trees describing the relationships between individuals in the same population/species (coalescent-based tree priors). Thus, non-coalescent methods could overestimate the age of lineages, while coalescent methods could bias the time estimation of inter-species divergences towards young ages (Liu et al., 2022; Mello et al., 2021). Consequently, we used three calibration schemes as follows: (1) we used a lognormal calibration prior distribution, which assigned the highest probabilities to substantially older ages than the calibrated ages accounting for errors associated with the incompleteness and limited knowledge of the Urticaceae fossil record. In this way, the Urticaceae crown group was calibrated using the estimated mean age for the family (68.7 Ma; Wu et al., 2018) as offset and the shape of the lognormal distribution was defined with a median of 72.8 Ma and a 95% probability interval between 69 and 136.4 Ma, using the unequivocal angiosperm crown group pollen grain fossils (Friis et al., 2010; Hughes, 1994) as soft upper bound (offset: 68.7, mean: 11.4, log (Stdev): 30). An uncorrelated lognormal relaxed molecular clock (UCLD) and the Yule speciation tree prior were applied to the trnL-F dataset. (2) The branching process prior (‘tree’ prior) was changed to use a birth–death model that considers the probability of frequent extinction processes throughout evolutionary history of the genus Pilea (not solely a continuous speciation as in the Yule model). (3) Lastly, the ‘tree’ prior was changed to use a constant-size coalescent model and with the rest of the parameters as in the previous dating strategies. We ran each scheme 20 million generations for the cpDNA data set of the MCMC chain, with a sampling frequency of 1000 generations. Convergence of the posterior distributions of the parameters was evaluated by monitoring the effective sample sizes (ESS > 200) and trace plots in Tracer (http://tree.bio.ed.ac.uk/softw are/tracer/). A maximum credibility tree, which represents the maximum posterior topology, was calculated in TreeAnnotator (Drummond & Rambaut, 2007) after discarding the first 10% of trees as burn-in.
Results
Morphological differences among populations
The first two PC components of the PCA explained 82.6% of the total variance in morphological traits (Table 2). The PC1 explained 67.7% of the variance was positively associated with the plant length, stem width, leaf length and leaf width, and negatively associated with the depth of margin teeth in proportion to leaf width. The PC2 (14.8% of the variance) was associated only with the proportion of leaf surface without teeth on margin (Table 2). Significant differences between groups (Chiapas vs. Tabasco) were observed only in PC1 (t = 19.166, df = 35.892, p-value < 2e-16; Fig. 2). Table 3 summarizes the main morphological differences between individuals from each of the studied localities.

Morphological variation among individuals of Pilea pteridophylla sampled along its distribution range in the tropical karst forest of southern Mexico. Plot of individual scores for the first two components of the principal component analysis using morphological data. Coloured symbols represent the two populations recognized for the species: red circles, Tabasco; and blue circles, Chiapas. Ellipses correspond to the 95% confidence intervals estimated for each population. The lines represent the dispersion of the individuals within each population
Genetic differences among populations
A matrix of 462 bp for trnL-F and 870 bp for rps16-trnQ was obtained. Thus, the concatenated matrix (rps16-trnQ + trnL-F) consisted of 1332 bp. The number of haplotypes varied from 4 to 8 per dataset with gaps considered missing data and from 7 to 15 when treated as 5th state (Table 4, Supplemental Fig. S1). The haplotype diversity values were relatively high and the nucleotide diversity values were low in all cases (Table 4). There are no shared haplotypes between populations (Fig. 3), and a strong population genetic structure is assumed. Pairwise FST values ranged from 0.33 to 0.89 and with significant differences in all comparisons (Table 4). The nucleotide diversity and FST values were always higher in the rps16-trnQ dataset and in the concatenated dataset (rps16-trnQ + trnL-F). In summary, both populations (Chiapas and Tabasco) show strong genetic differentiation with FST values close to 1 (Fig. 3, Table 4). Haplotype diversity was relatively high overall, with more haplotypes in Chiapas compared to Tabasco (Table 4). The nucleotide diversity was low in general, with haplotypes differentiated by few mutational steps within each of the populations.

Statistical parsimony networks of rps16-trnQ, trnL-trnF and rps16-trnQ + trnL-trnF dataset using the gaps as missing data. Coloured symbols represent the two populations recognized for the species: red circles, Tabasco; and blue circles, Chiapas. Open-white circles represent the number of mutational steps between haplotypes. The size of the circles is proportional to the frequency of each haplotype. Loops (dashed lines) were resolved following the criteria given by Pfenninger and Posada (2002). The grey area refers to the regions with the presence of mountain karst forests in southern Mexico [shape file available at CENAPRED (http://www.atlasnacionalderiesgos.gob.mx/archivo/visor-capas.html)]
Phylogenetic relationships and divergence time estimation
The maximum clade credibility tree that resulted from the BEAST analysis using the trnL-F dataset and the Yule speciation tree prior is shown in Fig. 4. Mean ages of major clades with 95% HPD intervals for the three calibration schemes are indicated in Table 5. The maximum clade credibility tree retrieved the genus Pilea (PP value = 0.99) as monophyletic, sister group of Lecanthus (PP value = 1.0) and Achudemia (PP value = 0.99). The genus Pilea is divided into two main clades and 8 sections (Fig. 4). The first clade (PP value = 0.27) corresponds to sections Tetraphyllae (Pilea tetraphylla only), Trimeris (PP value = 1.0), Lecanthoides (PP value = 1.0), Angulatae (PP value = 1.0) and Tetrameris (PP value = 1.0), and the last four nested in a strongly supported subclade (PP value = 1.0). The second clade contains the sections Verrucosae (PP value = 1.0), Plataniflorae (PP value = 1.0) and Pilea (PP value = 1.0). Pilea pteridophylla (PP value = 1.0) appears as a member of the section Pilea, along with Pilea pleuroneura, Pilea mexicana and Sarcopilea domingensis (= Pilea fairchildiana) where the Chiapas and Tabasco populations form two reciprocally monophyletic clades (Chiapas lineage with PP value = 0.25; and Tabasco lineage with PP value = 0.96).

Maximum clade credibility tree derived from the BEAST analysis. Posterior estimates of divergence times were inferred using a trnL-trnF dataset. Nodes are posterior mean ages (Mya), with blue node bars representing the 95% HPD intervals. Bayesian PP < 0.95 are indicated above branches. Geological time-scale abbreviations: PLIO, Pliocene; PL, Pleistocene
No major differences in the age estimates can be attributed to the use of different calibration tree priors (Table 5). The estimated median age of diversification for the Urticaceae family was between 71.5 and 75.2 Ma (95% HPD Yule model: 89.5–68.7 Ma; 95% HPD birth–death model: 85.8–68.7 Ma; 95% HPD constant-size coalescent model: 92.0–68.7 Ma). The estimated age for the diversification of the genus Pilea was estimated between the Eocene and early Miocene [Yule model: mean = 38.1 Ma (95% HPD 54.5–24.5 Ma); birth–death model: Mean = 34.4 Ma (95% HPD 48.1–18.6 Ma); constant-size coalescent model: Mean = 33.7 Ma (95% HPD 56.8–12.3 Ma)], so its current distribution could be associated with a boreotropical origin. According to the analysis of the trnL-F chloroplast marker, the two populations of Pilea pteridophylla could have diverged during the middle Miocene and late Pliocene [Yule model: mean = 7.2 Ma (95% HPD 13.5–3.5 Ma)] or later between the late Miocene and the Pleistocene [birth–death model: mean = 4.95 Ma (95% HPD 7.4–1.7 Ma); constant-size coalescent model: mean = 3.9 Ma (95% HPD 7.5–1.0 Ma)], in any case, a deep divergence and isolation between individuals restricted to different vegetation patches (Fig. 4).
Discussion
In this study, the influence of geographic isolation on the levels of morphological and genetic differentiation among populations of Pilea pteridophylla was evaluated. Based on our results, there is a strong morphological differentiation among populations of Pilea pteridophylla (Fig. 2, Table 3). Both populations can be clearly differentiated based on vegetative and reproductive characters (Table 3), and the observed variation is geographically structured (Fig. 2). Moreover, our genetic data from the chloroplast genome show strong levels of population differentiation and no shared haplotypes were observed between populations (Fig. 3, Table 4). Also, our estimation of divergence times supports a long-term isolation hypothesis between Chiapas and Tabasco populations (Fig. 4, Table 5). Thus, our study supports the hypothesis of allopatric speciation as one of the main evolutionary processes triggering diversity within the mountain karst forests of Mexico (Wendt, 1987). Based on the evidence presented here, some evolutionary factors involved in the separation of Mexican karst-adapted plant lineages are discussed below.
Low seed-mediated gene flow and habitat heterogeneity promotes genetic differentiation among populations adapted to karstic soils
Based on the low substitution rates attributed to chloroplast markers, the estimated deep age of divergence between the Chiapas and Tabasco populations of Pilea pteridophylla is somewhat unusual. However, the deep divergence suggests that seed-mediated migration rates are actually low in Pilea pteridophylla. This result is quite interesting since long-distance dispersal through water currents has been hypothesized as the main dispersal mechanism in Urticaceae (Wu et al., 2018). According to Wu et al. (2018), seeds of many Urticaceae floated for > 220 days, and remained viable after 10 months in water. Pilea pteridophylla is an herb that grows exclusively in cave entrances or on rocky knolls, where surface water is scarce but with large runoff during the rainy season. However, seed dispersal through water currents is rather local. In Chiapas, groups of 10 or more individuals of Pilea pteridophylla are widely dispersed throughout the forest, suggesting that water currents can move seeds long distances locally. This is supported by the higher number of haplotypes observed in Chiapas compared to those recorded in Tabasco. In Tabasco, the population is rather dense, with all individuals separated by a few centimetres, where the effect of water on seed dispersal and on the level of genetic differentiation among individuals is low. Thus, it is likely that the genetic flow between Chiapas and Tabasco is currently interrupted.
Our results also suggest that topographic complexity is an important feature for population differentiation. First, the geographic distance between populations (more than 80 km2) and their differences in altitude suppose ecological barriers to genetic exchange between populations. Second, local-scale habitat preferences (rocky knolls vs. caves and walls) are probably more related to their local distribution and their morphological characteristics (Liu et al., 2021). More specifically, individuals from Chiapas have larger stems because in the rocky knolls the soil can be deeper, resulting in greater support for their roots and stems, while for individuals from Tabasco, the rocky walls could limit plant growth (in stems and roots), probably associated with less soil and water (Matthes-Sears & Douglas, 1995; Liu et al., 2020). However, field experiments are needed to test this hypothesis.
The karst-endemic species as relict lineages of ancient vegetation
Our estimation of divergence times shows a deep divergence (between middle Miocene and Pleistocene) among Pilea pteridophylla populations. This level of divergence is consistent with the strong phylogeographic structure and the clear morphological differentiation between individuals restricted to each population. Wendt (1987) suggests that the different levels of morphological differentiation between endemic sister species in the karst forests of Mexico could be related to the time of isolation between patches of vegetation to which they are restricted, in addition to the fact that karst forests have functioned as floristics refuges at different times and in different places between the Miocene and Pleistocene (Wendt, 1987, 1993). Our data support this idea. Specifically, the strong differentiation between populations of Pilea pteridophylla could be related to a prolonged geographic isolation driven by the past climate fluctuations occurred between middle Miocene and Pleistocene. Also, Pilea pteridophylla is the only species with fern-like leaves within the genus Pilea, and its calcareous habit, restricted distribution and its strong population differentiation suggest a long-term adaptation to a karstic habitat. Based on this evidence, it is possible to hypothesize that the species endemic to the karst forests of Mexico are relicts of an ancient vegetation, while the non-endemic elements could be more recent invaders (Wendt, 1987, 1993). Although little is known about the time in which the plant immigration events to the karst relief occurred (see Li et al., 2022), the data published so far (focused on Asian forests) suggest that primary immigration events occurred during the Miocene, which is supported in this study; however, it could also be related to the expansion and diversification during the Miocene of many lineages of lowland tropical forest plants in general (Becerra, 2005; Ornelas et al., 2018; Ortiz-Rodriguez et al., 2020; Rull, 2020; Gutiérrez-Ortega et al., 2024). Interestingly, floristic data suggest that only a small part (less than 4%) of the species currently present in the mountain karst forest of Mexico are exclusive (endemic) to this biome (see Molina-Paniagua et al., 2023), and for another small percentage of species (6%), between 50 and 90% of their records are distributed exclusively within this forest. This suggests that adaptation to karst soils is a slow evolutionary process. Viewed this way, our data support the idea of endemic karst species as ancient forest elements, but also show that plant diversification in karst forests has not been constant (as suggested by long-term forest stability), and has occurred to a lesser extent towards the present.
Long-term geographic isolation promotes speciation
Based on our results, the individuals of Pilea pteridophylla restricted to each population can be morphologically and genetically differentiated. These differences are associated with a long-term geographic isolation without apparent gene flow and contrasting ecological conditions. This evidence shows convincingly that the individuals from Chiapas are an independent lineage and should be recognized as a new species to science. Consequently, a new Mexican species of Pilea is described below.
Taxonomic section
Pilea monroi Ortiz-Rodr. & Lagos-Báez sp. nov. (Fig. 5).

Pilea monroi Ortiz-Rodr. and Lagos-Báez (Urticaceae). A Habit, plant with pistillate inflorescences. B Stem, a leafless portion with inflorescences. C Flower bud. D Staminate flower with four stamens and four tepals. E Infructescence. F Stem portion with pistillate inflorescences. Drawn by Robin Pérez Lucas based on Lagos-Báez 57 (MEXU)
Etymology
Pilea monroi named in honour of Dr Alex Monro, taxonomist, specialist in the genus Pilea (Urticaceae) and especially interested in the study of the plant diversity in the karstic forests of America and China.
Similar to Pilea pteridophylla in general fern-like appearance, but clearly differentiated by its dentate (vs. pectinate) leaves, and larger stem and leaf size. Furthermore, they are allopatrically distributed and under different environmental conditions.
Type: MEXICO: Chiapas; Berriozábal, Zona sujeta a protección ecológica “La Pera”. Predio “Flor de Café”, 16°51′50.6″ N, 93°19′51.7″ W, alt. 1068 m., May 2019, K. Lagos-Báez 57 (Holotype, MEXU; isotypes: MO, KEW).
Description
Herbs 254–740 mm, perennial, epipetric. Stems erect, sparsely branched, greenish in vivo, blackened when dry, glabrous, cystoliths linear, X or Y shaped, not clearly visible when dry, the internodes 7.5–54 × 1.5–2.4 mm, irregularly circular or slightly angled in cross-section, striate. Leaves petiolate, distichous, laminae and petioles at the same node of equal length, glabrous, laminae 80–180 × 12–22 mm, linear lanceolate, chartaceous, deeply dentate above de basal 1/7 to 1/3, adaxial surface dark green when dry, glabrous, cystoliths fusiform, V-, X-, Y-shaped, abaxial surface greyish or olive, rarely light brown, glabrous, cystoliths fusiform, V-, X-, Y-shaped, glandular punctate, base symmetrical or nearly so, decurrent, attenuated, rounded, apex cuspidate, primary venation pinnate, secondary veins 26–42 pairs at 90° to midrib; petioles 3.5–5.0 × 0.8–2.5 mm, stipules 0.4–1.0 mm, deltate, membranous, often persistent. Inflorescences ca. 15 per stem, 2.0–4.0 mm, unisexual, pistillate inflorescences preceding and concurrent with staminate inflorescences, peduncle and pedicels subtended by stipuliform, minute bracts, staminate flowers, cream or slightly pink, tepals 4, 0.8–1.2 mm, ovate, glabrous, stamens 4, filaments 0.5–0.8 mm, anthers 0.3–0.4 mm, pistillate flowers, cream, tepals 3, 0.5–0.8 mm, the central one slightly longer than the two lateral, ovate, glabrous. Infructescences of 2–3 mm, peduncle 0.35 mm diam., fruits 0.4–0.6 × 0.2–0.3 mm, asymmetric, subcompressed ovoid, orange-brown, margin broadening and flattened towards the apex.
Distribution
Pilea monroi is known only from the type locality in Chiapas, Mexico. The species inhabits within the mountain karst forest at ca. 1000 m altitude.
Notes
Pilea monroi is the only species in Mexico with linear lanceolate, deeply dentate leaves, and like Pilea pteridophylla, its fern-like appearance makes it easily distinguishable from other species. However, the leaf margins of Pilea monroi are dentate and not pectinate as in Pilea pteridophylla.
Pilea plumosa from Costa Rica, also have linear lanceolate leaves with a dentate margin, but it is distinguished from Pilea monroi by its narrower leaf blades (1.5–4.0 mm vs 12–22 mm), by the abaxial leaf surface covered by orbicular cystoliths (vs V-, X-, Y-shaped) and by its leaves with fewer secondary veins (4–13 pairs vs 26–42 pairs).
Additional specimens examined
MEXICO: Chiapas; Berriozábal, Zona sujeta a protección ecológica “La Pera”. Predio “Peña Flor”, 16°51′50.6″ N, 93°19′51.7″ W, alt. 1068 m., Ortiz-Rodríguez A. E. 320 (HEM).
Data availability
The authors declare the availability of data in the supplementary section of the manuscript.
References
Alboukadel, K. (2020). ggpubr: 'ggplot2' based publication ready plots. R package version 0.4.0. https://CRAN.R-project.org/package=ggpubr
Becerra, J. (2005). Timing the origin and expansion of the Mexican tropical dry forest. Proceedings of the National Academy of Sciences of the United States of America, 102, 10919–10923. https://doi.org/10.1073/pnas.0409127102
Chambers, J. M., Freeny, A., & Heiberger, R. M. (1992). Analysis of variance; designed experiments. Chapter 5 of Statistical Models in S. J. M. Chambers and T. J. Hastie (eds), Wadsworth & Brooks/Cole. Claus, O. W. (2020). cowplot: Streamlined Plot Theme and Plot Annotations for 'ggplot2'. R package version 1.1.1. https://CRAN.R-project.org/package=cowplot
Clement, M., Posada, D., & Crandall, K. A. (2000). TCS: A computer program to estimate gene genealogies. Molecular Ecology, 9(10), 1657–1660
Clements, R., Sodhi, N. S., Schilthuizen, M., & Ng, P. K. L. (2006). Limestone karsts of Southeast Asia: Imperiled arks of biodiversity. BioScience, 56(9), 733–742. https://doi.org/10.1641/0006-3568(2006)56733:LKOSAI2.0.CO2
Cota-Sánchez, J. H., Remarchuk, K. & Ubayasena, K. (2006). Ready-to-use DNA extracted with a CTAB method adapted for herbarium specimens and mucilaginous plant tissue. Plant Molecular Biology Reporter 24, 161. https://doi.org/10.1007/BF02914055
Darriba, D., Taboada, G. L., Doallo, R., & Posada, D. (2012). jModelTest 2: More models, new heuristics and parallel computing. Nature Methods, 9, 772
Drummond, A. J., & Rambaut, A. (2007). BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evolutionary Biology, 7, 214. https://doi.org/10.1186/1471-2148-7-214
Espinasa Pereña, R. (1990). Propuesta de clasificación del karst de la República Mexicana. Universidad Nacional Autónoma de México
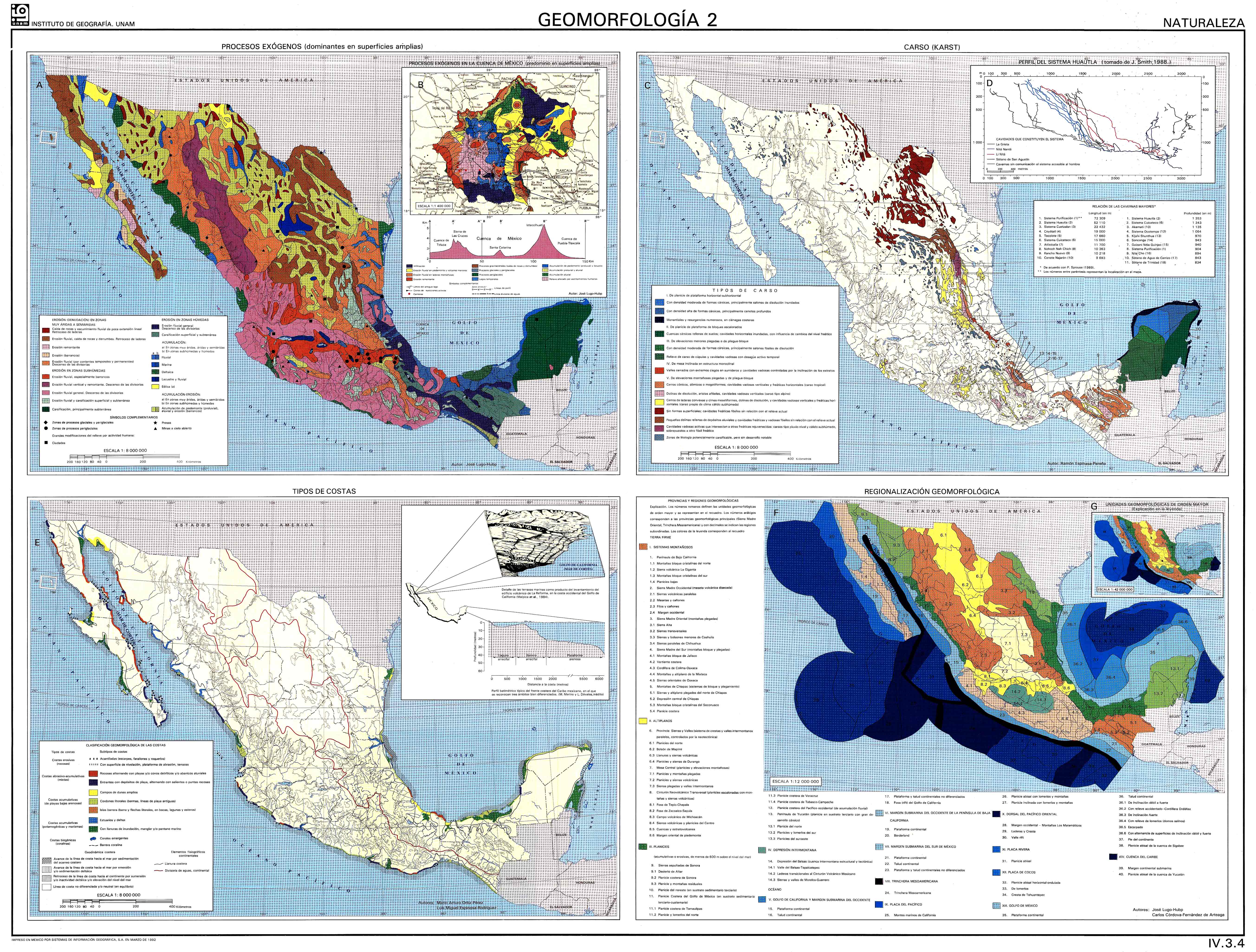
Espinasa Pereña, R. (2007). El Karst de México. Mapa NA III 3. Nuevo Atlas Nacional de México (Instituto de Geografía, Universidad Nacional Autónoma de México).https://geodigital.geografia.unam.mx/atlas_nacional/index.html/grals/Tomo_II/IV.Naturaleza/IV.3.Relieve/IV.3.4.jpg
Excoffier, L., Laval, G., & Schneider, S. (2005). Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol Bioinform, 1, 47–50
Friis, E. M., Pedersen, K. R., & Crane, P. R. (2010). Diversity in obscurity: Fossil flowers and the early history of angiosperms. Phil. Trans. R. Soc. B, 365, 369–382. https://doi.org/10.1098/rstb.2009.0227
Fu, L. F., Wen, F., Maurin, O., Rodda, M., Gardner, E. M., Xin, Z. B., Wei, Y. G., & Monro, A. K. (2022). A revised delimitation of the species-rich genus Pilea (Urticaceae) supports the resurrection of Achudemia and a new infrageneric classification. Taxon, 71(4), 796–813. https://doi.org/10.1002/tax.12711
Gao, Y., Ai, B., Kong, H., Kang, M., & Huang, H. (2015). Geographical pattern of isolation and diversification in karst habitat islands: A case study in the Primulina eburnea complex. Journal of Biogeography, 42, 2131–2144. https://doi.org/10.1111/jbi.12576
Geekiyanage, N., Goodale, U. M., Cao, K., & Kitajima, K. (2019). Plant ecology of tropical and subtropical karst ecosystems. Biotropica, 51, 626–640. https://doi.org/10.1111/btp.12696
Gutiérrez-Ortega, J. S., Pérez-Farrera, M. A., Matsuo, A., Sato, M. P., Suyama, Y., Calonje, M., Vovides, A. P., Kajita, T., & Watano, Y. (2024). The phylogenetic reconstruction of the Neotropical cycad genus Ceratozamia (Zamiaceae) reveals disparate patterns of niche evolution. Molecular Phylogenetics and Evolution, 190, 107960. https://doi.org/10.1016/j.ympev.2023.107960
Hammer, Ø., Harper, D. A. T, & Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Paleontol Electronica 4, 4. http://palaeo-electronica.org/2001_1/past/issue1_01.htm
Hughes, N. F. (1994). The enigma of angiosperm origins. Cambridge University Press
Li, X-Q, Xiang, X-G, Zhang, Q., Jabbour, F., Ortiz, R.C., Erst, A. S., Li, Z-Y, & Wang, W. (2022). Immigration dynamics of tropical and subtropical Southeast Asian limestone karst floras. Proc. R. Soc. B.2892021130820211308. https://doi.org/10.1098/rspb.2021.1308
Librado, P., & Rozas, J. (2009). DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics, 25, 1451–1452
Liu, L., Hu, J., Chen, X., Xu, X., Yang, Y., & Ni, J. (2022). Adaptation strategy of karst forests: Evidence from the community-weighted mean of plant functional traits. Ecology and Evolution, 12, e8680. https://doi.org/10.1002/ece3.8680
Liu, Y., Chen, J., Xiaojing, Z., Qiaoyu, L., Yu, D., Jinchun, L., & Jianping, T. (2021). Herbaceous community species composition and productivity are affected by soil depth and funnel effect in a simulated karst experiment. Global Ecology and Conservation, 22, e01033. https://doi.org/10.1016/j.gecco.2020.e01033
Lugo-Hubp, J. (1990) El relieve de la República Mexicana. Revista Mexicana de Ciencias Geológicas 474 (9), 82–111. http://rmcg.geociencias.unam.mx/index.php/rmcg/article/view/1255
Matthes-Sears & Douglas, W. L. (1995). Rooting characteristics of trees in rock: A study of Thuja occidentalis on cliff faces. International Journal of Plant Sciences, 156(5), 679–686
Mello, B., Tao, Q., Barba-Montoya, J., & Kumar, S. (2021). Molecular dating for phylogenies containing a mix of populations and species by using Bayesian and RelTime approaches. Molecular Ecology Resources, 21(1), 122–136. https://doi.org/10.1111/1755-0998.13249
Molina-Paniagua, M. E., Alves de Melo, P. H., Ramírez-Barahona, S., Ortiz-Rodriguez, A. E., Monro, A.K., Burelo-Ramos, C. M., & Gómez-Domínguez H (2023) How diverse are the mountain karst forests of Mexico? bioRxiv 2023.03.01.530643; doi: https://doi.org/10.1101/2023.03.01.530643
Monro, A. K. (1999). Seven new species of Pilea Lindley (Urticaceae) from Mesoamerica. Novon, 9(3), 390–400. https://doi.org/10.2307/3391738
Monro, A. K. (2001). Synopsis of Mesoamerican Pilea (Urticaceae), including eighteen typifications and a key to the species. Bulletin of the Natural History Museum, Botany Series 31, 9–25. https://www.biodiversitylibrary.org/part/190603
Müller, K., Müller, J., & Quandt, D. (2010). PhyDE (Phylogenetic Data Editor). http://www.phyde.de/
Ornelas, J. F, Licona-Vera, Y., & Vásquez-Aguilar, A. A. (2018). Genetic differentiation and fragmentation in response to climate change of the narrow endemic Psittacanthus auriculatus. Tropical Conservation Science 11. https://doi.org/10.1177/19400829187555
Ornelas, J. F., Gándara, E., Vásquez-Aguilar, A. A., et al. (2016). A mistletoe tale: Postglacial invasion of Psittacanthus schiedeanus (Loranthaceae) to Mesoamerican cloud forests revealed by molecular data and species distribution modeling. BMC Evolutionary Biology, 16, 78. https://doi.org/10.1186/s12862-016-0648-6
Ortiz-Rodriguez, A. E, Lorea-Henández, F., Pérez-Farrera, M. A., & Monro, A. K. (2012). First record of Pilea pteridophylla A.K.Monro (Urticaceae) and Ocotea heribertoi T.Wendt (Lauraceae) for the flora of Chiapas. Botanical Sciences 90(2), 203–206. https://doi.org/10.17129/botsci.485
Ortiz-Rodríguez, A. E., Licona-Vera, Y., Vásquez-Aguilar, A. A., Hernández-Soto, M., López-Huicochea, E. A., & Ornelas, J. F. (2020). Genetic differentiation among Psittacanthus rhynchanthus (Loranthaceae) populations: Novel phylogeographic patterns in the Mesoamerican tropical lowlands. Plant Systematic and Evolution, 306, 10. https://doi.org/10.1007/s00606-020-01638-y
Ortiz-Rodriguez, A. E. (2023a). “rps16-trnQ+ trnL-F cpDNA sequences from Pilea pteridophylla (Urticaceae)”, Mendeley Data, V1. https://doi.org/10.17632/xxbkfcdnnc.1
Ortiz-Rodriguez, A. E. (2023b). “trnL-trnF cp DNA sequences from Pilea (Urticaceae)”, Mendeley Data, V1. https://doi.org/10.17632/xhzgb287vd.1
Pérez-García, E. A., & Meave, J. A. (2005). Heterogeneity of xerophytic vegetation of limestone outcrops in a tropical deciduous forest region in southern México. Plant Ecology, 175, 147–163. https://doi.org/10.1007/s11258-005-4841-8
Pérez‐García, E. A., Sevilha, A. C., Meave, J. A., & Scariot, A. (2009). Floristic differentiation in limestone outcrops of southern Mexico and central Brazil: A beta diversity approach. Boletín de la Sociedad Botánica de México 84, 45–58. https://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S0366-21282009000100005
Pfenninger, M., & Posada, D. (2002). Phylogeographic history of the land snail Candidula unifasciata (Helicellinae, Stylommatophora): Fragmentation, corridor migration, and secondary contact. Evolution 56 (9), 1776–88. https://www.jstor.org/stable/3094736
R Core Team. (2021). R: A language and environment for statistical computing. R Foundation for statistical computing. Vienna, Austria. Available at http://www.R-project.org
Rull, V. (2020). Neotropical diversification: Historical overview and conceptual insights. In: Rull, V., Carnaval, A. (eds) Neotropical Diversification: Patterns and Processes. Fascinating Life Sciences. Springer, Cham. https://doi.org/10.1007/978-3-030-31167-4_2
Shaw, J., Lickey, E. B., Schilling, E. E., & Small, R. L. (2007). Comparison of whole chloroplast genome sequences to choose non-coding regions for phylogenetic studies in angiosperms: The tortoise and the hare III. American Journal of Botany, 94, 275–288. https://doi.org/10.3732/ajb.94.3.275
Shepherd, L. D., & Heenan, P. B. (2017). Evidence for both long-distance dispersal and isolation in the Southern Oceans: Molecular phylogeny of Sophora sect Edwardsia (Fabaceae). New Zealand Journal of Botany, 55, 334–346. https://doi.org/10.1080/0028825X.2017.1353527
Taberlet, P., Gielly, L., Pantou, G., & Bouvet, J. (1991). Universal primers for amplification of three non-coding regions of plastid DNA. Plant Molecular Biology, 17, 1105–1105. https://doi.org/10.1007/BF00037152
Tseng, Y.-H., Huang, H.-Y., Xu, W.-B., Yang, H.-A., Peng, C.-I., Liu, Y., et al. (2019). Phylogeography of Begonia luzhaiensis suggests both natural and anthropogenic causes for the marked population genetic structure. Botanical Studies, 60, 20. https://doi.org/10.1186/s40529-019-0267-9
Wang, J., Ai, B., Kong, H., & Kang, M. (2017). Speciation history of a species complex of Primulina eburnea (Gesneriaceae) from limestone karsts of southern China, a biodiversity hot spot. Evolutionary Applications, 10, 919–934. https://doi.org/10.1111/eva.12495
Wendt, T. (1987). Las selvas de Uxpanapa, Veracruz-Oaxaca, México: Evidencia de refugios florísticos cenozoicos. Anales Instituto De Biología, Universidad Nacional Autónoma De México, Serie Botánica, 58, 29–54
Wendt, T. (1993). Composition, floristic affinities, and origins of the canopy tree flora of the Mexican Atlantic slope rain forests. In T. P. Ramamoorthy, R. Bye, A. Lot, & J. Fa (Eds.), Biological Diversity of Mexico (pp. 595–680). Oxford University Press.
Wright, S. (1965). The interpretation of population structure by F-statistics with special regard to systems of mating. Evolution, 19(3), 395–420. https://doi.org/10.2307/2406450
Wu, Z.-Y., Liu, J., Provan, J., Wang, H., Chen, C.-J., Cadotte, M. W., Luo, Y.-H., Amorim, B. S., Li, D.-Z., & Milne, R. I. (2018). Testing Darwin’s transoceanic dispersal hypothesis for the inland nettle family (Urticaceae). Ecology Letters, 21, 1515–1529. https://doi.org/10.1111/ele.13132
Zhu, X., Liang, H., Jiang, H., Kang, M., Wei, X., Deng, L., & Shi, Y. (2022). Phylogeographic structure of Heteroplexis (Asteraceae), an endangered endemic genus in the limestone karst regions of southern China. Frontiers in Plant Science, 13, 999964. https://doi.org/10.3389/fpls.2022.999964
Acknowledgements
We sincerely thank Christopher Davidson and Sharon Christoph for their support through the project “KARSTBIO: Biodiversity, Evolution and Conservation of the tropical rain forest on karstic zones of Latin America”. Also, we thank the anonymous reviewers for providing many useful comments on the manuscript. This work forms part of the research project with which the first author obtained her bachelor's degree.
Author information
Authors and Affiliations
Contributions
KLB and AEOR conceived the ideas; KLB, HGD and AEOR conduct the field work; KLB and AAVA conducted the lab work; KLB and AEOR analysed the data; AEOR wrote the main manuscript text; YLV, LHR and JFO reviewed the manuscript and contribute substantially to the structure and writing of the final version. All authors read and approved the final version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
Cite this article
Lagos-Báez, K., Licona-Vera, Y., Gómez-Domínguez, H. et al. Genetic and morphological differentiation among populations of the narrowly endemic and karst forest-adapted Pilea pteridophylla (Urticaceae). Org Divers Evol (2024). https://doi.org/10.1007/s13127-024-00638-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13127-024-00638-x