
Abstract
Mammalian adipose tissue is traditionally categorized into white and brown relating to their function and morphology: while white serves as an energy storage, brown adipose tissue acts as the heat generator maintaining the core body temperature. The most recently identified type of fat, beige adipocyte tissue, resembles brown fat by morphology and function but is developmentally more related to white. The synthesis of beige fat, so-called browning of white fat, has developed into a topical issue in diabetes and metabolism research. This is due to its favorable effect on whole-body energy metabolism and the fact that it can be recruited during adult life. Indeed, brown and beige adipose tissues have been demonstrated to play a role in glucose homeostasis, insulin sensitivity, and lipid metabolism—all factors related to pathogenesis of type 2 diabetes. Many agents capable of initiating browning have been identified so far and tested widely in humans and animal models including in vitro and in vivo experiments. Interestingly, several agents demonstrated to have browning activity are in fact secreted as adipokines from brown and beige fat tissue, suggesting a physiological relevance both in beige adipocyte recruitment processes and in maintenance of metabolic homeostasis. The newest findings on agents driving beige fat recruitment, their mechanisms, and implications on type 2 diabetes are discussed in this review.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Traditionally, adipose tissues have been divided into subcategories of white adipose tissue (WAT) and brown adipose tissue (BAT) according to their function and morphology. BAT functions in thermogenesis generating heat through combustion of nutrients that is uncoupled from ATP production by uncoupling protein (UCP1). Besides, the main functions of energy storage (WAT) and heat production (BAT) are considered endocrine tissues due to secretion of adipokines that participate in metabolic regulation of the body [14]. Newest addition, beige adipose tissue, has been established as an intermediate between BAT and WAT. The tissue shares the ability to thermogenesis as well as morphological features. Beige fat tissue derives from precursors of white adipocytes [27, 45] and is found in clusters scattered within the areas of white adipose tissue as opposed to occurrence of BAT in separate and distinct depots.
An aspect of interest in beige adipose tissue is that beige adipocytes have been demonstrated to be recruitable. Beige adipocytes exhibit low basal expression level of UCP1 and other thermogenesis related genes that are inducible upon stimulus, such as cold stimulation [26, 84, 91]. Theories behind the origin of beige adipocytes include (i) trans-differentiation from mature white adipocytes, (ii) existence of a distinct beige fat cell precursor, and (iii) differentiation from brown or white adipocyte precursors [17, 60, 61]. It was previously suggested that browning is a feature of WAT instead of beige adipocytes being a distinct cell type, but this theory was heavily questioned by the findings on beige adipocyte gene expression pattern by Wu et al. [88].
Brown and beige adipocytes have raised an interest in health sciences with their capability to counteract different metabolic diseases, including obesity and type 2 diabetes. Thermogenesis without ATP production, orchestrated by UCP1, is an effective way of assisting energy wastage, thus creating a negative energy balance. While some metabolic disorders and obesity are commonly linked to type 2 diabetes, brown adipose tissue contributes directly to the disease onset and permanence. Obesity resistance has also been linked to activation of these tissues in several mouse models [9, 34, 66]. In addition, brown adipose tissue might exert a beneficial effect on whole-body metabolism via other routes, such as lipid and glucose clearance from circulation. Taking into account the beneficial effects of BAT occurrence, the emergence of thermogenesis-able beige adipocytes and browning of WAT hold an interest and potential towards treating type 2 diabetes. In addition, beige adipocytes are hypothesized to have a distinct role in metabolic regulation and exhibit different physiological purposes compared to classical brown adipocytes [45]. This review discusses different browning agents that have been discovered and studied so far in the context of diabetes type 2 treatment and/or effect of browning to glucose metabolism and insulin sensitivity.
Development of adipose tissues on the molecular level
Brown and white fat develop from mesoderm through separate differentiating lineages. Mesenchymal stem cells (MSCs; multipotent stem cells of the adipose tissue extracellular matrix) can commit to either adipogenic or myogenic lineage, leading to emergence of either white adipocytes, brown adipocytes, or myocytes. Classical brown adipocytes and myocytes develop along myogenic lineage and derive from myogenic factor 5 positive cells (Myof5+), whereas precursors of white and beige adipocytes are Myof5− and thus adipogenic [60, 67]. Despite sharing a precursor, beige fat cells exhibit distinct gene expression compared to either WAT or BAT, combining expression pattern of genes characteristic to WAT (adiponectin, adipocyte protein 2 gene (aP2), adipsin) and BAT (Ucp1, Cidea, Pgc-1α). In addition, beige fat cells express genes specific to the fat tissue, such as a developmental transcription factor T-box1 (Tbx1), a fatty acid transporter solute carrier family 27 member 1 (Slc27a1), and transmembrane protein 26 (TMEM26) as well as molecules of immune and inflammatory response pathways, such as CD40 and CD137 [88]. Distinct genetic loci drive the regulation of BAT and beige adipocytes, suggesting that different pathways are responsible for regulation of these tissues.
The differentiation processes of adipogenic and myogenic lineages are orchestrated with surprisingly similar transcriptional cascades. Peroxisome proliferator-activated receptor-γ (PPARγ) is the key switch determining the cell fate and differentiation to either a brown or a white adipocyte [29]. PPARγ drives the adipogenesis transcriptional program in co-operation with a set of different co-activators and regulators, such as CCAAT/enhancer binding proteins (C/EBS), PPARγ co-activator-1α (PGC-1α) and PRDM16. PGC-1α is the major regulator of mitochondrial biogenesis and oxidative metabolism in brown fat and other cell types, such as skeletal muscle, and one of the most important factors of brown adipocyte development [62]. PRDM16, a transcriptional co-regulator, is one of the key switches between determining the development into a brown adipocyte or a myocyte. PRDM16 functions by interacting with multiple DNA-binding transcriptional factors, such as PPARα, PPARγ, and several C/EBP protein family members and has the capability of inducing PGC-1α expression, while C/EBS participate in the upkeep of the differentiation state [29, 67]. In addition to major transcriptional switches PPARγ and PRDM16 that determine the cell fate between myocyte, WAT, and BAT, other overlapping cascades and molecular factors are also involved in adipogenesis and adipocyte differentiation.
BAT and beige connections to type 2 diabetes
BAT intakes glucose and lipids from circulation aiding in glucose clearance and relieving the demand for insulin secretion by β-cells, thus improving their function [45]. Fasting glucose concentrations in individuals with detectable BAT were demonstrated to be lower than in those without active BAT [38]. The predominant source of energy for brown adipose tissue metabolism is fatty acids stored in brown adipocytes [46], while only 10% of the energy is derived from plasma glucose. However, the clearance of this glucose has been demonstrated to be effective in diabetics. In humans, boosting of glucose uptake into BAT consists of two different mechanisms: insulin-dependent and insulin-independent [57]. Heat production-related glucose uptake into BAT has been suggested to be independent from insulin signaling, thus bypassing all the normal parameters regulating the blood glucose level [15, 50, 56]. BAT is one of the most sensitive target tissues for insulin action and is considered to be the most important metabolically active organ for glucose. BAT activation enhances insulin signaling in BAT itself by combining the insulin-independent glucose uptake with thermogenesis and uptake of the glucose administered by insulin signaling [45].
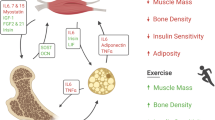
Many of the effects responsible for glucose and lipid clearance from plasma are indirectly improving the state of insulin resistance and β-cell failure. While glucolipotoxicity is one of the major pathologies behind insulin resistance, peripheral lipid and glucose clearance benefits β-cells and aids in restoring peripheral insulin sensitivity [59]. In practice, BAT-related treatment methods include (i) increasing the BAT mass, (ii) recruitment of beige adipocytes by browning of WAT, and (iii) boosting the effects of activation of already existing BAT by targeting the regulation of fuel uptake and utilization [57]. Formation of beige adipocytes has been demonstrated to have ameliorating effects to body weight as well as to glucose and lipid homeostasis [7, 50]. Physiological stimuli traditionally considered to induce the formation of beige adipocytes include cold exposure, diet, and exercise. In addition to activation of classical BAT, prolonged cold exposure, as well as β-adrenergic agonist treatment, has been demonstrated to be able to brown WAT [24, 25, 91].
Browning agents
In the following paragraphs and Table 1, we list and discuss a comprehensive and updated set of browning agents as reported in the literature.
Cold exposure
Cold exposure is one of the oldest and most studied physiological stimuli leading to WAT browning. In early studies in rodents, both acute and chronic cold exposure has been reported to increase oxygen consumption, stimulate BAT insulin signaling, and to lead to the emergence of Ucp1-positive cells and morphological changes in mouse parametrial fat bad [26, 84, 91]. Cousin et al. (1992) demonstrated increase in UCP1 expression in rodent periovarian WAT in response to prolonged cold exposure in + 4 °C [13]. Cold exposure induced browning in mice has also been studied by Vitali et al. (2012) and they suggest that the density of sympathetic nerves innervating WAT is one of the key features for the browning effect to take place [81]. However, very little is known about SNS innervation in WAT browning and beige adipocytes, underlining it as one important future research line [21].
β-3 adrenergic receptor agonists
In early rodent models of Ghorbani et al. (1997), chronic treatment with beta3-adrenoceptor agonist CL 316243 was shown to lead to occurrence of brown adipocytes within traditional WAT depots including mesenteric, inguinal, epididymal, and retroperitoneal fat depots [24]. Cousin et al. (1992) reported increase in UCP1 expression in periovarian WAT depots in rodents as a result for treatment with BRL 26830A [13].
Short-chain fatty acids
These are end products of microbial fermentation of carbohydrates and starch by colon and have been linked to changes in body weight, appetite, and adiposity. Namely, acetate, butyrate, and propionate have been the compounds of interest while demonstrating to have protective effect against diet-induced obesity and insulin resistance in mice models [41]. Acetate has been demonstrated with WAT browning capacity in mice model [65].
Dietary factors and organic compounds
Several plant-based compounds and dietary factors have been demonstrated to have browning effects based on studies done on mice or in vitro [85]. Capsaicin (and related capsinoids) has been demonstrated to initiate WAT browning in mice by several different mechanisms, including modulation of expression of PPARγ 1 co-activator α, facilitation of PPARγ–Prdm16 interaction, and promotion of sirtuin-1 (SirT1) expression [4]. Plant-produced resveratrol has been demonstrated to induce Ucp1 and Pgc1α gene expression, having similar effects to butein in a mice model [2]. Plant-derived berberine (BBR) has been shown to induce development of brown-like adipocytes in inguinal, but not epididymal adipose depots in mice through PGC-1α signaling [94]. Fish oil intake has been shown to lead to upregulation of UCP1 and the β3 adrenergic receptor in both interscapular BAT and inguinal WAT of mice [31]. In the study of Chen et al. (2017), decaffeinated green tea extract supplemented for mice induced expression of variety of browning-related biomarkers, including PPARγ, PRDM-16, BMP-7, FGF-21, UCP-1, CPT-1 CIDEA, and PGC-1α [10].
In cell culture models, cinnamon extract has been demonstrated to increase expression of brown adipocyte markers, decrease those of WAT, and induce BAT phenotype in cell culture models done with 3T3-L1 adipocyte cell line. In addition, cinnamon effected UCP1 expression in adipocytes isolated from mouse subcutaneous WAT [35]. Similarly to cinnamon, several other compounds have been studied to promote browning of 3T3-L1 cells as observed by significant increases in mRNA expressions of UCP-1, PGC-1α, and PRDM16, including ginsenoside Rb1, curcumin, and quercetin [40, 43, 51].
Nuclear receptors and ligands
Bile acid-activated nuclear receptor, farnesoid X receptor (FXR) regulates bile acid, lipid and glucose homeostasis, and several other regulatory functions in the intestinal tract. FXR activation with fexaramine antagonist has been demonstrated to enhance thermogenesis and browning of subcutaneous WAT [19]. Liver X receptors (LXRs), nuclear receptors that control lipid and glucose metabolism, have been reported to be able to regulate the browning of WAT, activation of mitochondria, and increased energy expenditure [49].
MicroRNAs
Different microRNA types and their roles have been studied also in terms of WAT browning and adipose tissue physiology. MicroRNAs have been shown to regulate both BAT activation and subcutaneous WAT browning in cold exposure [70]. miRNA-32 inhibition in vivo has been demonstrated to reduce FGF21 expression and compromise tolerance to cold, thus having a role in cold-induced WAT browning [52]. MiRNA-455 has been studied to enhance WAT browning response to cold and norepinephrine stimulation [92]. In contrast, several miRNA types act as negative regulators of WAT browning and BAT activity [70].
Drug agents
Chronic treatment with thiazolidinediones (TZDs) has been demonstrated to induce browning of WAT by activating PPARγ and PRDM16 [60]. Prostaglandin E2 (PGE2) is a PPARγ co-operator that has been demonstrated to divert preadipocyte differentiation in WAT to beige/brite mature adipocytes and exert upregulation of UCP1 [23]. Another PPARγ-signaling affecting drug, PPARγ antagonist Gleevec has been demonstrated to significantly increase brown/beige fat thermogenic and mitochondrial genes, including Ucp-1, Pgc-1α, and cox-5b in WAT and interscapular BAT in mice [11]. Slit 2 derived secretory product, a member of the Slit extracellular protein family, has been demonstrated to regulate beige adipocyte induction via Prdm16 and robust activation of PKA signaling. Slit2-C fragment has an active thermogenic property having an effect on energy expenditure, and glucose homeostasis in vivo [73]. Artepillin C (ArtC) has been demonstrated to induce brown-like adipocytes in mice and primary inguinal WAT-derived adipocytes in vitro through PPARγ and PRDM16 signaling [54].
Adrenomedullin 2 (AM2) has been demonstrated to induce browning of rat primary adipocytes in vitro along with the variety of metabolism ameliorating effects through PGC1α and PRDM16 signaling [93]. Beta-lapachone (BLC) has been reported with the ability to stimulate browning of WAT, increase expression of UCP1, and ameliorate metabolic parameters of mice fed a high-fat diet. The effects of BLC were partially controlled via the regulation of miR-382 targeting deionidase iodothyronine type II (Dio2), but the overall mechanism remains unclear [12].
Inflammatory factors
In a setting of post burn injury, bone marrow originated interleukin 6 (IL-6) has been demonstrated to regulate WAT browning [1]. Interleukin 4 (IL-4) has been reported with a role in BAT activation responding to cold exposure in a macrophage-dependent manner [53]. In addition, deficiency of immediate early response gene X-1 (IEX-1), a downstream effector of Nf-Kb signaling pathway, was demonstrated to induce browning and activate thermogenic gene program in WAT through promotion of alternative activation of adipose macrophages [69].
Hormonal factors
Many different hormones released from variety of organs also exert a crosstalk with adipose tissue physiology and related processes. Thyroid hormones are well known to be able to induce WAT browning through both peripheral and central mechanisms [87]. Triiodothyronine (T3) metabolite triiodothyracetic acid (TRIAC) has been demonstrated with the ability to regulate WAT browning process and induce ectopic expression of UCP1 in abdominal WAT depot of rats [48]. In human cell culture model, T3 treatment enhanced UCP1 expression and mitochondrial biogenesis in human adipose-derived stem cells [37]. Circulating T4 levels have been shown to correspond to UCP1 expression in WAT in humans [47]. While deionidase type II (Dio2) is a key enzyme regulating thyroid hormone signaling and plays a crucial role in thyroid-hormone-related activation of BAT, it is also suggested to be one of the major players in thyroid hormone induced WAT browning [86]. Parathyroid hormone (PTHr) and tumor-derived parathyroid hormone-related protein (PTHrP) has been demonstrated to lead WAT browning in cancer-cachexia models studied in mice [32, 75]. Glucagon-like peptide 1 (GLP1) has been demonstrated to stimulate BAT thermogenesis and adipocyte browning in mice [5, 44]. Leptin together with insulin has been reported to drive WAT browning and weight loss through action on hypothalamic neurons [16]. Melatonin induces browning of WAT in inguinal WAT of rats as observed by increased expression of PGC-1α and UCP-1 [28]. Natriuretic peptides (NPs), endocrine hormones released from the heart, are demonstrated to possess WAT browning capacity and effect on BAT metabolism through several different pathways [6, 42].
Genetic factors
A set of genes have been associated with WAT browning including PTEN, Cox2, Foxc2, folliculin, and Gq. Expression of Gq, a type of G protein-coupled receptor in human WAT, has been demonstrated to inversely correlate with UCP1 expression [33]. Modulation of TGF-β/Smad3 signaling is proposed to activate a brown adipocyte like phenotype in rodent WAT [90].
Batokines
Batokines, such as fibroblast growth factor 21 (FGF21), interleukin 6 (IL-6), and prostaglandin D2 synthase (PTGDS), have been demonstrated to regulate glycaemia and metabolism in animal models by further enhancing glucose and lipid clearance [59, 79]. Fisher et al. demonstrated that in addition to having ameliorating effect on classical BAT and metabolism, FGF21 expression is also necessary for the cold-induced recruitment of beige adipocytes in WAT. Browning is connected with induction of PGC-1α protein levels having only little effect on mRNA expression. Since circulating levels of FGF21 are not elevated upon cold exposure, it has been suggested that FGF21 exerts its action on adipose tissue browning via para- and autocrine mechanisms [20]. However, Keipert and Jastroch demonstrated that FGF21 secreted from muscle acts as an endocrine hormone, targeting other tissues than the tissue of origin. In WAT, FGF21 secreted from skeletal muscle manifests effects that lead to the increased browning of epididymal and subcutaneous fat depots in mice [30]. Apelin, a newly identified adipocyte-derived hormone that is secreted by both brown and white adipocytes, stimulates adipose browning [74]. In their studies, Than et al. demonstrated that apelin increases the activity of BAT, induces browning of WAT, and stimulates metabolic activity both in vitro and in vivo [74].
Exercise
Capability of exercise to induce browning has been a controversial issue. Older studies in humans and rodents report negative results for BAT activation and browning in response to exercise [68, 72, 82]. However, exercise has also been demonstrated to increase expression of PGC-1α [80, 89] and a recently identified exercise adipomyokine irisin has been shown to induce browning in human and rodents [7, 39]. Irisin is cleaved from fibronectin type III domain-containing protein 5 (FNDC5) expressed on the surface of skeletal muscle and WAT, and secreted into circulation. In their studies, Boström et al. demonstrated the browning capability of irisin in in vitro experiments in mouse subcutaneous WAT as well as described the elevated level of plasma irisin in humans in response to exercise [7].
PPAR ligands and signaling
In mice, treatment with PPAR agonists has been demonstrated to increase UCP1 expression in different WAT depots, particularly in the inguinal depot in vivo [22, 36]. In addition, Petrovic et al. demonstrated browning of epididymal WAT depot in mice in response to prolonged treatment with PPARγ agonist rosiglitazone [60]. Study by Ohno et al. (2012) suggested that browning effect of PPAR ligands is centered to stabilization and expression of PRDM16, the main factor controlling classical BAT development [55]. On the other hand, other suggested regulators of PPARγ signaling include α/β-hydrolase domain 6 (ABHD6) that has been demonstrated to negatively regulate adipose tissue browning [95]. In cell culture models of human and mice white adipocytes, a selective PPARα-agonist WY14643 was shown to stimulate mitochondrial biogenesis and UCP1 expression along with the PPAR co-activator PGC-1α [3, 76, 78]. Since PPARγ signaling is one of the main switches in adipocyte development, its involvement in pathways related to WAT browning is highly likely.
BMPs
Bone morphogenetic protein 7 (BMP7) was demonstrated to be able to induce BAT selective genes and induce UPC1 mRNA expression in brown preadipocytes [77]. It also drives the browning of mesenchymal progenitor cells by inducing the expression of PRDM16. This leads to the full activation of BAT gene expression pattern, suggesting that BMP7 is one of the factors determining the cell fate between brown and white [78]. Both BMP7 and BMP4 were demonstrated to induce white-to-brown shifting in primary human adipose stem cells [18]. Bone morphogenetic proteins (BMPs) have been suggested to facilitate adipogenetic differentiation by induction of PRDM16 and PGC-1α [77] and by negatively regulating skeletal myogenesis [71]. The Wingless pathway (Wnt) is crucial in embryonic muscle formation. Activation of this pathway represses brown and white adipogenesis by suppressing the induction of PPARγ and C/EBSα in the precursor cells [64].
Metabolites
Lactate and β-hydroxybutyrate have been demonstrated to increase UCP1 gene expression in human and murine WAT by intracellular redox modifications [8]. Beta-aminoisobutyric acid (BAIBA) has been demonstrated to increase expression of BAT marker genes in WAT of mice both in vitro and in vivo through PPARα-mediated mechanism. In humans, plasma levels of BAIBA are increased with exercise and compound is able to induce BAT-like phenotype in human pluripotent stem cells [63]. Eicosapentaenoic acid has been demonstrated with potential to activate and increase marker gene expression in BAT depots of mice, but no effect on WAT depots has been described [58]. Retinoic acid, a bioactive metabolite of vitamin A, has been reported to induce WAT browning via activating vascular endothelial growth factor (VEGF) signaling in vitro. In co-operation with VEGFA, retinoic acid also was shown to activate p38MAPK signaling pathway and to bind to Prdm16 promoter enhancing its transcription [83].
Concluding remarks
Browning of white adipose tissue has been—and remains to be—an active field of research due to its demonstrated connection and potential to contribute to treatment and prevention of diabetes type 2 and other prevalent metabolic disorders. From the continuously expanding list of novel browning agents, naturally occurring compounds such as metabolites, dietary, hormonal, and stimulated physiological factors hold a special interest due to lack of side effects and non-invasive treatment possibilities of obesity and metabolic disorders.
References
Abdullahi A, Chen P, Stanojcic M, Sadri AR, Coburn N, Jeschke MG (2017) IL-6 signal from the bone marrow is required for the browning of white adipose tissue post burn injury. Shock 47(1):33–39
Azhar Y, Parmar A, Miller CN, Samuels JS, Rayalam S (2016) Phytochemicals as novel agents for the induction of browning in white adipose tissue. Nutr Metab (Lond) 13:89–016–0150-6 eCollection 2016
Barbera MJ, Schluter A, Pedraza N, Iglesias R, Villarroya F, Giralt M (2001) Peroxisome proliferator-activated receptor alpha activates transcription of the brown fat uncoupling protein-1 gene. A link between regulation of the thermogenic and lipid oxidation pathways in the brown fat cell. J Biol Chem 276(2):1486–1493
Baskaran P, Krishnan V, Ren J, Thyagarajan B (2016) Capsaicin induces browning of white adipose tissue and counters obesity by activating TRPV1 channel-dependent mechanisms. Br J Pharmacol 173(15):2369–2389
Beiroa D, Imbernon M, Gallego R, Senra A, Herranz D, Villarroya F, Serrano M, Ferno J, Salvador J, Escalada J, Dieguez C, Lopez M, Fruhbeck G, Nogueiras R (2014) GLP-1 agonism stimulates brown adipose tissue thermogenesis and browning through hypothalamic AMPK. Diabetes 63(10):3346–3358
Bordicchia M, Liu D, Amri EZ, Ailhaud G, Dessi-Fulgheri P, Zhang C, Takahashi N, Sarzani R, Collins S (2012) Cardiac natriuretic peptides act via p38 MAPK to induce the brown fat thermogenic program in mouse and human adipocytes. J Clin Invest 122(3):1022–1036
Bostrom P, Wu J, Jedrychowski MP, Korde A, Ye L, Lo JC, Rasbach KA, Bostrom EA, Choi JH, Long JZ, Kajimura S, Zingaretti MC, Vind BF, Tu H, Cinti S, Hojlund K, Gygi SP, Spiegelman BM (2012) A PGC1-alpha-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 481(7382):463–468
Carriere A, Jeanson Y, Berger-Muller S, Andre M, Chenouard V, Arnaud E, Barreau C, Walther R, Galinier A, Wdziekonski B, Villageois P, Louche K, Collas P, Moro C, Dani C, Villarroya F, Casteilla L (2014) Browning of white adipose cells by intermediate metabolites: an adaptive mechanism to alleviate redox pressure. Diabetes 63(10):3253–3265
Cederberg A, Gronning LM, Ahren B, Tasken K, Carlsson P, Enerback S (2001) FOXC2 is a winged helix gene that counteracts obesity, hypertriglyceridemia, and diet-induced insulin resistance. Cell 106(5):563–573
Chen LH, Chien YW, Liang CT, Chan CH, Fan MH, Huang HY (2017) Green tea extract induces genes related to browning of white adipose tissue and limits weight-gain in high energy diet-fed rat. Food Nutr Res 61(1):1347480
Choi SS, Kim ES, Jung JE, Marciano DP, Jo A, Koo JY, Choi SY, Yang YR, Jang HJ, Kim EK, Park J, Kwon HM, Lee IH, Park SB, Myung KJ, Suh PG, Griffin PR, Choi JH (2016) PPARgamma antagonist Gleevec improves insulin sensitivity and promotes the browning of white adipose tissue. Diabetes 65(4):829–839
Choi WH, Ahn J, Jung CH, Jang YJ, Ha TY (2016) Beta-lapachone prevents diet-induced obesity by increasing energy expenditure and stimulating the browning of white adipose tissue via downregulation of miR-382 expression. Diabetes 65(9):2490–2501
Cousin B, Cinti S, Morroni M, Raimbault S, Ricquier D, Penicaud L, Casteilla L (1992) Occurrence of brown adipocytes in rat white adipose tissue: molecular and morphological characterization. J Cell Sci 103(Pt 4):931–942
Cypess AM, Lehman S, Williams G, Tal I, Rodman D, Goldfine AB, Kuo FC, Palmer EL, Tseng YH, Doria A, Kolodny GM, Kahn CR (2009) Identification and importance of brown adipose tissue in adult humans. N Engl J Med 360(15):1509–1517
Dallner OS, Chernogubova E, Brolinson KA, Bengtsson T (2006) Beta3-adrenergic receptors stimulate glucose uptake in brown adipocytes by two mechanisms independently of glucose transporter 4 translocation. Endocrinology 147(12):5730–5739
Dodd GT, Decherf S, Loh K, Simonds SE, Wiede F, Balland E, Merry TL, Munzberg H, Zhang ZY, Kahn BB, Neel BG, Bence KK, Andrews ZB, Cowley MA, Tiganis T (2015) Leptin and insulin act on POMC neurons to promote the browning of white fat. Cell 160(1–2):88–104
Elabd C, Chiellini C, Carmona M, Galitzky J, Cochet O, Petersen R, Penicaud L, Kristiansen K, Bouloumie A, Casteilla L, Dani C, Ailhaud G, Amri EZ (2009) Human multipotent adipose-derived stem cells differentiate into functional brown adipocytes. Stem Cells 27(11):2753–2760
Elsen M, Raschke S, Tennagels N, Schwahn U, Jelenik T, Roden M, Romacho T, Eckel J (2014) BMP4 and BMP7 induce the white-to-brown transition of primary human adipose stem cells. Am J Phys Cell Physiol 306(5):C431–C440
Fang S, Suh JM, Reilly SM, Yu E, Osborn O, Lackey D, Yoshihara E, Perino A, Jacinto S, Lukasheva Y, Atkins AR, Khvat A, Schnabl B, Yu RT, Brenner DA, Coulter S, Liddle C, Schoonjans K, Olefsky JM, Saltiel AR, Downes M, Evans RM (2015) Intestinal FXR agonism promotes adipose tissue browning and reduces obesity and insulin resistance. Nat Med 21(2):159–165
Fisher FM, Kleiner S, Douris N, Fox EC, Mepani RJ, Verdeguer F, Wu J, Kharitonenkov A, Flier JS, Maratos-Flier E, Spiegelman BM (2012) FGF21 regulates PGC-1alpha and browning of white adipose tissues in adaptive thermogenesis. Genes Dev 26(3):271–281
Fraçois M, Qualls-Creekmore E, Berthoud HR, Münzberg H, Yu S (2018) Genetics-based manipulation of adipose tissue sympathetic innervation. Physiol Behav 190:21–27
Fukui Y, Masui S, Osada S, Umesono K, Motojima K (2000) A new thiazolidinedione, NC-2100, which is a weak PPAR-gamma activator, exhibits potent antidiabetic effects and induces uncoupling protein 1 in white adipose tissue of KKAy obese mice. Diabetes 49(5):759–767
Garcia-Alonso V, Claria J (2014) Prostaglandin E2 signals white-to-brown adipogenic differentiation. Adipocyte 3(4):290–296
Ghorbani M, Himms-Hagen J (1997) Appearance of brown adipocytes in white adipose tissue during CL 316,243-induced reversal of obesity and diabetes in Zucker fa/fa rats. Int J Obes Relat Metab Disord 21(6):465–475
Guerra C, Koza RA, Yamashita H, Walsh K, Kozak LP (1998) Emergence of brown adipocytes in white fat in mice is under genetic control. Effects on body weight and adiposity. J Clin Invest 102(2):412–420
HART JS, HEROUX O (1956) Utilization of body reserves during exposure of mice to low temperatures. Can J Biochem Physiol 34(3):414–421
Hocking S, Samocha-Bonet D, Milner KL, Greenfield JR, Chisholm DJ (2013) Adiposity and insulin resistance in humans: the role of the different tissue and cellular lipid depots. Endocr Rev 34(4):463–500
Jimenez-Aranda A, Fernandez-Vazquez G, Campos D, Tassi M, Velasco-Perez L, Tan DX, Reiter RJ, Agil A (2013) Melatonin induces browning of inguinal white adipose tissue in Zucker diabetic fatty rats. J Pineal Res 55(4):416–423
Kajimura S, Seale P, Kubota K, Lunsford E, Frangioni JV, Gygi SP, Spiegelman BM (2009) Initiation of myoblast to brown fat switch by a PRDM16-C/EBP-beta transcriptional complex. Nature 460(7259):1154–1158
Keipert S, Jastroch M (2014) Brite/beige fat and UCP1 - is it thermogenesis? Biochim Biophys Acta 1837(7):1075–1082
Kim M, Goto T, Yu R, Uchida K, Tominaga M, Kano Y, Takahashi N, Kawada T (2015) Fish oil intake induces UCP1 upregulation in brown and white adipose tissue via the sympathetic nervous system. Sci Rep 5:18013
Kir S, White JP, Kleiner S, Kazak L, Cohen P, Baracos VE, Spiegelman BM (2014) Tumour-derived PTH-related protein triggers adipose tissue browning and cancer cachexia. Nature 513(7516):100–104
Klepac K, Kilic A, Gnad T, Brown LM, Herrmann B, Wilderman A, Balkow A, Glode A, Simon K, Lidell ME, Betz MJ, Enerback S, Wess J, Freichel M, Bluher M, Konig G, Kostenis E, Insel PA, Pfeifer A (2016) The Gq signalling pathway inhibits brown and beige adipose tissue. Nat Commun 7:10895
Kopecky J, Clarke G, Enerback S, Spiegelman B, Kozak LP (1995) Expression of the mitochondrial uncoupling protein gene from the aP2 gene promoter prevents genetic obesity. J Clin Invest 96(6):2914–2923
Kwan HY, Wu J, Su T, Chao XJ, Liu B, Fu X, Chan CL, Lau RHY, Tse AKW, Han QB, Fong WF, Yu ZL (2017) Cinnamon induces browning in subcutaneous adipocytes. Sci Rep 7(1):2447–017–02263-5
Laplante M, Sell H, MacNaul KL, Richard D, Berger JP, Deshaies Y (2003) PPAR-gamma activation mediates adipose depot-specific effects on gene expression and lipoprotein lipase activity: mechanisms for modulation of postprandial lipemia and differential adipose accretion. Diabetes 52(2):291–299
Lee J-Y, Takahashi N, Yasubuchi M, Kim Y-I, Hashizaki H, Kim M-J, Sakamoto T, Goto T, Kawada T (2012) Triiodothyronine induces UCP-1 expression and mitochondrial biogenesis in human adipocytes. Am J Phys Cell Physiol 302(2):C463–C472
Lee P, Greenfield JR, Ho KK, Fulham MJ (2010) A critical appraisal of the prevalence and metabolic significance of brown adipose tissue in adult humans. Am J Physiol Endocrinol Metab 299(4):E601–E606
Lee P, Linderman JD, Smith S, Brychta RJ, Wang J, Idelson C, Perron RM, Werner CD, Phan GQ, Kammula US, Kebebew E, Pacak K, Chen KY, Celi FS (2014) Irisin and FGF21 are cold-induced endocrine activators of brown fat function in humans. Cell Metab 19(2):302–309
Lee SG, Parks JS, Kang HW (2017) Quercetin, a functional compound of onion peel, remodels white adipocytes to brown-like adipocytes. J Nutr Biochem 42:62–71
Lin HV, Frassetto A, Kowalik EJ Jr, Nawrocki AR, Lu MM, Kosinski JR, Hubert JA, Szeto D, Yao X, Forrest G, Marsh DJ (2012) Butyrate and propionate protect against diet-induced obesity and regulate gut hormones via free fatty acid receptor 3-independent mechanisms. PLoS One 7(4):e35240
Liu D, Ceddia RP, Collins S (2018) Cardiac natriuretic peptides promote adipose 'browning' through mTOR complex-1. Mol Metab 9:192–198
Lone J, Choi JH, Kim SW, Yun JW (2016) Curcumin induces brown fat-like phenotype in 3T3-L1 and primary white adipocytes. J Nutr Biochem 27:193–202
Lopez M, Dieguez C, Nogueiras R (2015) Hypothalamic GLP-1: the control of BAT thermogenesis and browning of white fat. Adipocyte 4(2):141–145
Loyd C, Obici S (2014) Brown fat fuel use and regulation of energy homeostasis. Curr Opin Clin Nutr Metab Care 17(4):368–372
Ma SW, Foster DO (1986) Uptake of glucose and release of fatty acids and glycerol by rat brown adipose tissue in vivo. Can J Physiol Pharmacol 64(5):609–614
Martínez-Sánchez N, Moreno-Navarrete JM, Contreras C, Rial-Pensado E, Fernø J, Nogueiras R, Diéguez C, Fernández-Real JM, López M (2016) Thyroid hormones induce browning of white fat. J Endocrinol 232(2):351–262
Medina-Gomez G, Calvo RM, Obregon M-J (2008) Thermogenic effect of triiodothyroacetic acid at low doses in rat adipose tissue without adverse side effects in the thyroid axis. Am J Physiol Endocrinol Metab 294(4):E688–E697
Miao Y, Wu W, Dai Y, Maneix L, Huang B, Warner M, Gustafsson JA (2015) Liver X receptor beta controls thyroid hormone feedback in the brain and regulates browning of subcutaneous white adipose tissue. Proc Natl Acad Sci U S A 112(45):14006–14011
Mossenbock K, Vegiopoulos A, Rose AJ, Sijmonsma TP, Herzig S, Schafmeier T (2014) Browning of white adipose tissue uncouples glucose uptake from insulin signaling. PLoS One 9(10):e110428
Mu Q, Fang X, Li X, Zhao D, Mo F, Jiang G, Yu N, Zhang Y, Guo Y, Fu M, Liu JL, Zhang D, Gao S (2015) Ginsenoside Rb1 promotes browning through regulation of PPARgamma in 3T3-L1 adipocytes. Biochem Biophys Res Commun 466(3):530–535
Ng R, Hussain NA, Zhang Q, Chang C, Li H, Fu Y, Cao L, Han W, Stunkel W, Xu F (2017) miRNA-32 drives brown fat thermogenesis and trans-activates subcutaneous white fat browning in mice. Cell Rep 19(6):1229–1246
Nguyen KD, Qiu Y, Cui X, Goh YP, Mwangi J, David T, Mukundan L, Brombacher F, Locksley RM, Chawla A (2011) Alternatively activated macrophages produce catecholamines to sustain adaptive thermogenesis. Nature 480(7375):104–108
Nishikawa S, Aoyama H, Kamiya M, Higuchi J, Kato A, Soga M, Kawai T, Yoshimura K, Kumazawa S, Tsuda T (2016) Artepillin C, a typical Brazilian propolis-derived component, induces brown-like adipocyte formation in C3H10T1/2 cells, primary inguinal white adipose tissue-derived adipocytes, and mice. PLoS One 11(9):e0162512
Ohno H, Shinoda K, Spiegelman BM, Kajimura S (2012) PPARgamma agonists induce a white-to-brown fat conversion through stabilization of PRDM16 protein. Cell Metab 15(3):395–404
Olsen JM, Sato M, Dallner OS, Sandstrom AL, Pisani DF, Chambard JC, Amri EZ, Hutchinson DS, Bengtsson T (2014) Glucose uptake in brown fat cells is dependent on mTOR complex 2-promoted GLUT1 translocation. J Cell Biol 207(3):365–374
Orava J, Nuutila P, Lidell ME, Oikonen V, Noponen T, Viljanen T, Scheinin M, Taittonen M, Niemi T, Enerback S, Virtanen KA (2011) Different metabolic responses of human brown adipose tissue to activation by cold and insulin. Cell Metab 14(2):272–279
Pahlavani M, Razafimanjato F, Ramalingam L, Kalupahana NS, Moussa H, Scoggin S, Moustaid-Moussa N (2017) Eicosapentaenoic acid regulates brown adipose tissue metabolism in high-fat-fed mice and in clonal brown adipocytes. J Nutr Biochem 39:101–109
Peirce V, Vidal-Puig A (2013) Regulation of glucose homoeostasis by brown adipose tissue. Lancet Diabetes Endocrinol 1(4):353–360
Petrovic N, Walden TB, Shabalina IG, Timmons JA, Cannon B, Nedergaard J (2010) Chronic peroxisome proliferator-activated receptor gamma (PPARgamma) activation of epididymally derived white adipocyte cultures reveals a population of thermogenically competent, UCP1-containing adipocytes molecularly distinct from classic brown adipocytes. J Biol Chem 285(10):7153–7164
Pisani DF, Djedaini M, Beranger GE, Elabd C, Scheideler M, Ailhaud G, Amri EZ (2011) Differentiation of human adipose-derived stem cells into “Brite” (brown-in-white) adipocytes. Front Endocrinol (Lausanne) 2:87
Puigserver P, Wu Z, Park CW, Graves R, Wright M, Spiegelman BM (1998) A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell 92(6):829–839
Roberts LD, Bostrom P, O'Sullivan JF, Schinzel RT, Lewis GD, Dejam A, Lee YK, Palma MJ, Calhoun S, Georgiadi A, Chen MH, Ramachandran VS, Larson MG, Bouchard C, Rankinen T, Souza AL, Clish CB, Wang TJ, Estall JL, Soukas AA, Cowan CA, Spiegelman BM, Gerszten RE (2014) Beta-aminoisobutyric acid induces browning of white fat and hepatic beta-oxidation and is inversely correlated with cardiometabolic risk factors. Cell Metab 19(1):96–108
Ross SE, Hemati N, Longo KA, Bennett CN, Lucas PC, Erickson RL, MacDougald OA (2000) Inhibition of adipogenesis by Wnt signaling. Science 289(5481):950–953
Sahuri-Arisoylu M, Brody LP, Parkinson JR, Parkes H, Navaratnam N, Miller AD, Thomas EL, Frost G, Bell JD (2016) Reprogramming of hepatic fat accumulation and ‘browning’ of adipose tissue by the short-chain fatty acid acetate. Int J Obes 40(6):955–963
Seale P, Conroe HM, Estall J, Kajimura S, Frontini A, Ishibashi J, Cohen P, Cinti S, Spiegelman BM (2011) Prdm16 determines the thermogenic program of subcutaneous white adipose tissue in mice. J Clin Invest 121(1):96–105
Seale P, Kajimura S, Yang W, Chin S, Rohas LM, Uldry M, Tavernier G, Langin D, Spiegelman BM (2007) Transcriptional control of brown fat determination by PRDM16. Cell Metab 6(1):38–54
Segawa M, Oh-Ishi S, Kizaki T, Ookawara T, Sakurai T, Izawa T, Nagasawa J, Kawada T, Fushiki T, Ohno H (1998) Effect of running training on brown adipose tissue activity in rats: a reevaluation. Res Commun Mol Pathol Pharmacol 100(1):77–82
Shahid M, Javed AA, Chandra D, Ramsey HE, Shah D, Khan MF, Zhao L, Wu MX (2016) IEX-1 deficiency induces browning of white adipose tissue and resists diet-induced obesity. Sci Rep 6:24135
Shamsi F, Zhang H, Tseng YH (2017) MicroRNA regulation of brown adipogenesis and thermogenic energy expenditure. Front Endocrinol (Lausanne) 8:205
Shen JJ, Huang L, Li L, Jorgez C, Matzuk MM, Brown CW (2009) Deficiency of growth differentiation factor 3 protects against diet-induced obesity by selectively acting on white adipose. Mol Endocrinol 23(1):113–123
Shibata H, Nagasaka T (1987) The effect of forced running on heat production in brown adipose tissue in rats. Physiol Behav 39(3):377–380
Svensson KJ, Long JZ, Jedrychowski MP, Cohen P, Lo JC, Serag S, Kir S, Shinoda K, Tartaglia JA, Rao RR, Chedotal A, Kajimura S, Gygi SP, Spiegelman BM (2016) A secreted Slit2 fragment regulates adipose tissue thermogenesis and metabolic function. Cell Metab 23(3):454–466
Than A, He HL, Chua SH, Xu D, Sun L, Leow MK, Chen P (2015) Apelin enhances brown adipogenesis and browning of white adipocytes. J Biol Chem 290:14679–14691
Thomas SS, Mitch WE (2017) Parathyroid hormone stimulates adipose tissue browning: a pathway to muscle wasting. Curr Opin Clin Nutr Metab Care 20(3):153–157
Tiraby C, Tavernier G, Lefort C, Larrouy D, Bouillaud F, Ricquier D, Langin D (2003) Acquirement of brown fat cell features by human white adipocytes. J Biol Chem 278(35):33370–33376
Tseng YH, Kokkotou E, Schulz TJ, Huang TL, Winnay JN, Taniguchi CM, Tran TT, Suzuki R, Espinoza DO, Yamamoto Y, Ahrens MJ, Dudley AT, Norris AW, Kulkarni RN, Kahn CR (2008) New role of bone morphogenetic protein 7 in brown adipogenesis and energy expenditure. Nature 454(7207):1000–1004
Tupone D, Madden CJ, Morrison SF (2014) Autonomic regulation of brown adipose tissue thermogenesis in health and disease: potential clinical applications for altering BAT thermogenesis. Front Neurosci 8:14
Villarroya J, Cereijo R, Villarroya F (2013) An endocrine role for brown adipose tissue? Am J Physiol Endocrinol Metab 305(5):E567–E572
Villena JA, Carmona MC, Rodriguez de la Concepcion M, Rossmeisl M, Vinas O, Mampel T, Iglesias R, Giralt M, Villarroya F (2002) Mitochondrial biogenesis in brown adipose tissue is associated with differential expression of transcription regulatory factors. Cell Mol Life Sci 59(11):1934–1944
Vitali A, Murano I, Zingaretti MC, Frontini A, Ricquier D, Cinti S (2012) The adipose organ of obesity-prone C57BL/6J mice is composed of mixed white and brown adipocytes. J Lipid Res 59(10):619–629
Vosselman MJ, Hoeks J, Brans B, Pallubinsky H, Nascimento EB, van der Lans AA, Broeders EP, Mottaghy FM, Schrauwen P, van Marken Lichtenbelt WD (2015) Low brown adipose tissue activity in endurance-trained compared with lean sedentary men. Int J Obes
Wang B, Fu X, Liang X, Deavila JM, Wang Z, Zhao L, Tian Q, Zhao J, Gomez NA, Trombetta SC, Zhu MJ, Du M (2017) Retinoic acid induces white adipose tissue browning by increasing adipose vascularity and inducing beige adipogenesis of PDGFRalpha(+) adipose progenitors. Cell Discov 3:17036
Wang X, Wahl R (2014) Responses of the insulin signaling pathways in the brown adipose tissue of rats following cold exposure. PLoS One 9(6):e99772
Wankhade UD, Shen M, Yadav H, Thakali KM (2016) Novel browning agents, mechanisms, and therapeutic potentials of brown adipose tissue. Biomed Res Int 2016:2365609
Weiner J, Kranz M, Klöting N, Kunath A, Steinhoff K, Rijntjes E, Köhtle J, Zeisig V, Hankir M, Gebhardt C, Deuther-Conrad W, Heiker JT, Kralisch S, Stumvoll M, Blüher M, Sabri O, Hesse S, Brust P, Tönjes A, Krause K (2016) Thyroid hormone status defines brown adipose tissue activity and browning of white adipose tissues in mice. Sci Rep 6:38124
Weiner J, Hankir M, Heiker JT, Fenske W, Krause K (2017) Thyroid hormones and browning of adipose tissue. Mol Cell Endocrinol 458:156–159
Wu J, Bostrom P, Sparks LM, Ye L, Choi JH, Giang AH, Khandekar M, Virtanen KA, Nuutila P, Schaart G, Huang K, Tu H, van Marken Lichtenbelt WD, Hoeks J, Enerback S, Schrauwen P, Spiegelman BM (2012) Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 150(2):366–376
Wu Z, Puigserver P, Andersson U, Zhang C, Adelmant G, Mootha V, Troy A, Cinti S, Lowell B, Scarpulla RC, Spiegelman BM (1999) Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 98(1):115–124
Yadav H, Rane SG (2012) TGF-beta/Smad3 signaling regulates brown adipocyte induction in white adipose tissue. Front Endocrinol (Lausanne) 3:35
Young P, Arch JR, Ashwell M (1984) Brown adipose tissue in the parametrial fat pad of the mouse. FEBS Lett 167(1):10–14
Zhang H, Guan M, Townsend KL, Huang TL, An D, Yan X, Xue R, Schulz TJ, Winnay J, Mori M, Hirshman MF, Kristiansen K, Tsang JS, White AP, Cypess AM, Goodyear LJ, Tseng YH (2015) MicroRNA-455 regulates brown adipogenesis via a novel HIF1an-AMPK-PGC1alpha signaling network. EMBO Rep 16(10):1378–1393
Zhang H, Zhang SY, Jiang C, Li Y, Xu G, Xu MJ, Wang X (2016) Intermedin/adrenomedullin 2 polypeptide promotes adipose tissue browning and reduces high-fat diet-induced obesity and insulin resistance in mice. Int J Obes 40(5):852–860
Zhang Z, Zhang H, Li B, Meng X, Wang J, Zhang Y, Yao S, Ma Q, Jin L, Yang J, Wang W, Ning G (2014) Berberine activates thermogenesis in white and brown adipose tissue. Nat Commun 5:5493
Zhao S, Mugabo Y, Ballentine G, Attane C, Iglesias J, Poursharifi P, Zhang D, Nguyen TA, Erb H, Prentki R, Peyot ML, Joly E, Tobin S, Fulton S, Brown JM, Madiraju SR, Prentki M (2016) Alpha/beta-hydrolase domain 6 deletion induces adipose browning and prevents obesity and type 2 diabetes. Cell Rep 14(12):2872–2888
Funding
Open access funding provided by University of Oulu including Oulu University Hospital.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Kaisanlahti, A., Glumoff, T. Browning of white fat: agents and implications for beige adipose tissue to type 2 diabetes. J Physiol Biochem 75, 1–10 (2019). https://doi.org/10.1007/s13105-018-0658-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13105-018-0658-5