
Abstract
The fossil record of the lygaeoid true bug family, Geocoridae so far consisted of a compression fossil from the Florissant Formation, Geocoris infernorum, and a mould fossil from the Izarra Formation, G. monserrati. Here, we report the third taxon, Protogeocoris arcanus gen. et sp. nov. from Cenomanian Burmese amber along with revision of the two previously mentioned species. The study of the specimens resulted in erecting a new genus, Eocenogeocoris gen. nov. to accommodate G. infernorum [ = Eocenogeocoris infernorum (Sudder, 1890) comb. nov.], because of its unique combination of characters compared to extant geocorine true bugs. Furthermore, the interpretation of the novel morphological data acquired from the study of the fossil geocorids in the context of the knowledge on extant representatives of the family suggests that characteristic groups of the family may have been evolved until the mid-Cretaceous, whilst the species-level diversity of the nominotypical genus, Geocoris Fallén, 1814, and closely allied genera is possibly resulted by quick radiation event after a climatic changes at the Eocene-Oligocene boundary.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The family Geocoridae (Heteroptera: Pentatomomopha: Lygaeoidea), or commonly known as big-eyed bugs, is a peculiar lygaeoid true bug family in terms both morphological and ecological aspects. Representatives of the family display a series of highly specialised exoskeletal morphological features that are unique among lygaeoid true bugs, e.g. enlarged, reniform, often stylate eyes and medially curved sutures of abdominal tergites 4/6 and 5/6 (Henry, 1997). Furthermore, omnivorous feeding with a preference on predaceous behaviour is also unusual within the superfamily Lygaeoidea (lygaeoids are mostly seed and sap feeding except the hematophagous tribe Cleradini), which has consequently made some of the species promising candidates for biocontrol research (Sweet 2000; Kóbor 2020). However, taxonomy, phylogenetic relationships between and within taxon groups, and evolution of the representatives of the family are not well understood and virtually unstudied.
Controversially, these deficiencies are resulted by the peculiar appearance of these insects. Diagnoses of taxa were mostly based on colouration patterns and easy-to-observe exoskeletal characters, e.g. arrangement of head and eyes or shape of pronotum. This resulted in taxonomic confusions and misconceptions, especially in the nominotypical genus, Geocoris Fallén, 1814 which is considered as non-monophyletic in its current sense as suggested by more recent revisionary works (Readio and Sweet, 1982; Malipatil, 1994).
The examination of fossils provides useful information regarding the phylogenies and evolution of the studied groups and the interpretation of the results in context of recent taxa (e.g. including fossil taxa in phylogenetic reconstructions of extant groups) is momentous in solving systematic problems. As for geocorid true bugs, such data is relatively scarce. In total two fossil representatives of the family are known so far: Geocoris infernorum Scudder, 1890 from the Eocene (Florissant Formation, approximately 34 Ma.); and Geocoris monserrati Ortuño and Arillo, 1997 from the Oligocene (Izarra Formation, approximately 33.9 to 23.03 Ma.). Furthermore, the taxonomic placement of G. infernorum is precarious as stated by the author himself in the original description (Scudder 1890).
In this study, we present the description of the oldest known taxon belonging to this family from mid-Cretaceous Burmese amber. Furthermore, we review and revise the knowledge on the previously described fossil representatives of big-eyed bugs, discuss their placement within the family based on the morphological data available on recent taxa and form hypotheses on their evolutionary significance.
Materials and methods
The amber piece containing the studied fossil originated from Noije Bum, near Tanai Village Hukawng Valley, Kachin State, northern Myanmar (26_21°33.41’N, 96_43°11.88’E) (Grimaldi et al. 2002: fig. 1, Cruickshank and Ko 2003: fig. 1; Bai et al. 2016). The studied piece of amber was purchased by and deposited in the collection of Paleoentomological Collection, Instituto de la Patagonia, Universidad de Magallanes in good faith for educational and research purposes and the authors of present study have no knowledge on unethical origin.
Observations, photomicrographs, and measurement for Protogeocoris arcanus gen. et sp. nov. were done with the use of Japan Optical Co. XLT-2310 stereoscopic microscope with Ricoh WG-50 digital camera adapted. Incident and transmitted lighting were used, mostly simultaneously.
Morphological terminologies were adapted from Tsai et al. (2011), Malipatil and Blackett (2013), and Kóbor (2019).
Instutional abbreviations: MCNA, Museo de Ciencias Naturales de Álava, Vitória, Spain; MCZC, Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts, USA; PCIP, Paleoentomological Collection, Instituto de la Patagonia, Universidad de Magallanes, Punta Arenas, Chile.
Systematic palaeontology
Order Hemiptera Linnaeus, 1758
Suborder Heteroptera Latreille, 1810
Infraorder Pentatomomorpha Leston, Pendergrast and Southwood, 1954
Superfamily Lygaeoidea Schilling, 1829
Family Geocoridae Baerensprung, 1860
Subfamily Geocorinae Baerensprung, 1860
A key to fossil Geocorinae
1. Eyes projected. Pronotum with a narrow, but distinct collar. Corium with R-M branching in apical third. … Protogeocoris arcanus gen. et sp. nov.
- Eyes not projected. Pronotum without distinct collar. R-M of corium appear to be fused at entire length. … 2.
2. Head pentagonal, eyes touching anterior edges of pronotum. Pronotum trapeziform. … Geocoris monserrati Ortuño and Arillo, 1997
- Head lunular, eyes encompassing anterior edges of pronotum. Pronotum conspicuously widened. … Eocenogeocoris infernorum (Scudder, 1890) comb. nov.
Genus Protogeocoris nov.

Protogeocoris arcanus gen. et sp. nov.: a dorsal habitus; b reconstruction (scale bar = 0.50 mm for both images).
Type species: Protogeocoris arcanus sp. nov.; by present designation and monotypy; gender: masculine.
Etymology: Generic name is derived from the name of genus Geocoris Fallén, 1814 (nominotypical genus of family Geocoridae) adding the prefix “proto-“ (Latin, meaning: “first of its kind”) which refers to the fact that this genus is the oldest known representative of the geocorid true bugs.
Differential diagnosis: Pronotum bilobate with narrow, but distinct collar and posterior margin concave (bilobate pronotum and distinct prothoracic collar can be observed in some representatives of Pamphantinae, but posterior margin is never concave); R-M of hemelytron basally fused, branching at 2/3 of length (basally fused, branching R-M can be observed in e.g. Engistus Fieber, 1864, Umbrageocoris Kóbor, 2019, but in these taxa pronotum always unilobed with straight or convex posterior margin); sutures between abdominal tergites 4/5-5/6 slightly and evenly curved posteriad at entire length (sutures of abdominal tergites 4/5-5/6 curved moderately to strongly medially in other geocorine genera; see Fig. 6a-b).
Description
Head pentagonal with vertex broad; eyes large, reniform, moderately stylate. Ocelli present, situated at middle of vertex, near base of eye stalks. Clypeus slightly pointed, appearing to exceed the mandibular plates. Antenniferous tubercles developed, but not visible is dorsal view. Antennomere I shortest, granuliform; antennomere II longest, cylindrical; antenno-meres III and IV subequal in length; antennomere III cylindrical, antennomere IV fusiform. Labial trough non-observable. Labiomere I shortest, labiomeres II-IV subequal in length; apex of labium reaching mesocoxae.
Thorax: Pronotum bilobate, trapeziform with narrow, but distinct collar; lateral margins moderately carinate and slightly constricted medially; posterior margin concave; pronotal callosities furrow-like, separated. Scutellum subequilateral, triangular with lateral margins slightly curved. Hemelytron appears to be membranous with clavus and exocorium more sclerotized than the rest of the hemelytron. Margins of clavus subparallel; claval commissure short, but distinct. Integument of corium appears to be wrinkled with simplified venation; Cu running near claval furrow; R-M basally fused, branching at approximately 2/3 of length; exocorium widening apically. Membrane with simplified, slightly visible venation lacking closed cells. Peritreme of metathoracic scent efferent apparatus bulbous without dorsal supportive process; orifice appears to be rounded; evaporatorium fine, matte, covering most of the metapleurite. Femora incrassate, fusiform; tibiae subcylindri-cal, slightly widened at apex. Tarsomere I longest, tarsomere II and III subequal in length; tarsomeres I and II cylindrical, tarsomere III fusiform. Claws simple, thin, evenly curved with thin parempodia-like appendages.
Abomen: Abdominal dorsum mostly obscured by wings. Sutures between abdominal sternites 4/5-5/6 appear to be slightly, evenly curved posteriad. Outer genital characters, e.g. posterior opening of pygophore, parameres in situ, not observable.
Protogeocoris arcanus sp. nov.

Protogeocoris arcanus gen. et sp. nov.: a head and pronotum in dorsal view (image not to scale; lettering: cal pronotal callosities, col prothoracic collar, oc ocelli); b labiomeres (image not to scale; lettering: l1-4 – labimeres I-IV); c hemelytron (image not to scale; lettering: cla clavus, Cu cubitus, EC exocorium, M Media, R-M fused Media and Radius, R radius); d tarsus and pretarsal structures (image not to scale; cl tarsal claw, p parempodia-like pretarsal appendage, t1-3 – tarsomeres I-III).
Material: Holotype, PCIP: curatorial number: type n° 0003.
Locality and horizon: Noije Bum, near Tanai Village Hukawng Valley, Kachin State, northern Myanmar; lowermost Cenomanian, mid-Cretaceous.
Etymology: Species epithet “arcanus” (Latin, meaning: mysterious) refers to the evolutionary importance of the species in terms of the family Geocoridae.
Differential diagnosis: Small insect (> 2mm) with characteristic pentagonal head, and slightly stylate reniform eyes. Hemelytra with clavus and exocorium more sclerotized and darker than rest of the hemelytron. Abdominal dorsum with a pair of dark longitudinal bands sublaterally. Other characteristics as in generic diagnosis.
Description: Almost complete male individual in amber piece, left hind leg missing; characters obscured by syninclusions and air bubbles denoted in description respectively.
Colouration: Vertex, eye stalks and thoracic dorsum with irregular darker markings, partly obscured by air bubbles. Antennomere I with irregular brownish spots and brownish annulation apically, antennomeres II-III with darker annulation apically, antennomere IV entirely darker (Fig. 1a). Hemelytra with clavus and exocorium entirely darker than rest of the hemelytron. Femora with irregular brownish spots. Abdominal dorsum with a pair of longitudinal, subparallel darker bands sublaterally. Abdominal sternites darker dorsolaterally.
Structure: General habitus moderately elongate, slightly flattened dorsoventrally.
Head: Eye stalks slightly projected, posterior edge of eyes not encompassing, nor touching anterior edges pronotum; ocular sulcus unobservable. Ratio of antennomeres: 1: 5.5: 5.0: 4.0. Ratio of labiomeres: 1: 6.5: 5.25: 0.30.
Thorax: Integument of thoracic dorsum densely punctate except at pronotal callosities. Anterior edges of pronotum angulate, slightly rounded; humeral angles acute anteriorly and rounded posteriorly. Median trifurcate carina not observable, obscured by air bubbles. Thoracic pleurites and sternites densely punctate. Femora appear to be maculate.
Measurements (in mm): total length of body: 1.85; length of head: 0.27; antennomeres I-IV: 0.02-0.11-0.10-0.8; labiomeres: 0.08-0.52-0.42-0.24; length of pronotum: 0.48; width of pronotum: 0.57; length of scutellum: 0.22; width of scutellum: 0.40; length of hemelytron: 1.10.
Remarks on palaeohabitat: The amber piece containing multiple syninclusions, mostly soil and plant debris, remains of other insects and a piece of skin suspected to be originating from an avian leg. The most notable syninclusions are three complete insects: two barklice (Psocoptera gen. sp.) and a snakefly (Nanoraphidiini gen.sp.). Considering the results of Christiansen and Nascimbene (2006), Lis et al. (2018, 2020) and Jiang et al. (2019) we suggest that the palaeohabitat of the insect was part of an abundant geophilous-arboreal environment in an amber forest. The preference of geophilous-arboreal habitats is similar to those of the extant representatives of the family Geocoridae (Slater 1977).
Genus Eocenogeocoris Kóbor nov.
(Figure 4)

Syninclusion of the holotype of P. arcanus: a Psocoptera gen. sp.; b Nanoraphidiini gen.sp. (images not to scale).
Type species: Geocoris infernorum Scudder, 1890 by present designation; gender masculine
Etymology: Generic name is derived from the name of genus Geocoris Fallén, 1814, adding the prefix “eoceno-“ referring to that the genus is known exclusively from the Eocene epoch.
Differential diagnosis: Eocenogeocoris Kóbor gen. nov. possesses a combination of characters as in the representatives of subgenus Piocoris Stål, 1872 and some Geocoris species e.g. G. flaviceps (Burmeister, 1834) or G. willey Kirkaldy, 1905: habitus ovoid; head lunular, compound eyes slightly stylate with posterior edge of eyes encompassing the anterior edges of pronotum; pronotum widened; ccutellum elongate triangular. However, margins of clavus subparallel; claval commissure short, but complete in Eocenogeocoris (margins of clavus converging in Geocoris sensu lato and allies; synapomorphy with Geocorinae incertae sedis, see Table 1). Fore femora strongly incrassate (autapomorphy; fore femora is moderately incrassate in other Geocorinae). Sutures between abdominal tergites 4/5-5/6 appear to be at most moderately curved posteriad medially (sutures between abdominal tergites 4/5-5/6 curved strongly in Geocoris sensu lato as in Fig. 6b; suspected synapomorphy with genus Germalus and allies).
Eocenogeocoris infernorum (Scudder, 1890) comb. nov.
(Fig. 4)

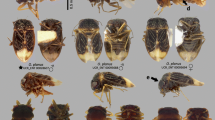
Eocenogeocoris infernorum (Scudder, 1890): a holotype; b paratype (scale bar = 1 mm for both images; lettering: a1-2: antennomeres I-II, abd5-6 – abdominal tergites V-VI; ce compound eye, cla clavus of hemelytron, cly clypeus, cor corium of hemelytron; f1 fore femora, gp genital plate; pr pronotum, sc scutellum).
Material examined: Holotype, MCZC: PALE-4411; paratypes, MCZC: PALE-4412, PALE-4413, PALE-4414, PALE-4415, PALE-4416.
Locality and horizon: Florissant Fossil Beds National Monument, Colorado, USA; uppermost Priabonian, late Eocene (approximately 34 Ma).
Redescription
Head lunular with vertex strongly widened; posterior edge of eyes encompassing anterior edges of pronotum. Eyes appear to be reniform, slightly stylate; ocelli present, situated near base of eye stalks. Antennomere I shortest, granuliform; antennomere II longest, cylindrical; antennomeres III and IV subequal in length; antennomere III cylindrical, antennomere IV fusiform.
Thorax: Pronotum conspicuously widened with anterior edges strongly rounded. Scutellum appears to be elongate triangular. Clavus, surroundings of venation and exocorium appear to be more sclerotized than the rest of the hemelytron. Clavus with margins subparallel; claval commissure complete, developed. Femora fusiform, fore femora appear to be strongly incrassated.
Abdomen. Abdominal tergites with sutures appear to be straight.
Remarks: The original description (Scudder, 1890) denotes that the identity and generic assignment of the species is problematic. This uncertainty stems from the nature of compression fossil (Azar et al., 2011; Chen et al., 2016) which can be distorted and mostly lacking detail. However, in case of G. infernorum there are characteristics to be observed based on which a diagnosis can be formulated. The convex general habitus, conspicuously lunular head and widened pronotum resembles to representatives of subgenus Piocoris Stål, 1872 and Geocoris s. str. species e.g. G. flaviceps (Burmeister, 1834) or G. willey Kirkaldy, 1905. However, these taxa are characterised by moderately incrassate fore femora, clavus with margins converging and claval commissure reduced, and sutures of abdominal tergites 4/5-5/6 strongly curved posteriad. In G. infernorum the fore femora are strongly incrassate, margins of clavus subparallel with a complete claval commissure and sutures of abdominal tergites 4/5-5/6 appear to be at most moderately curved. Similar arrangement of clavus and abdominal tergites can be found in Germalus and allies. Summarising, this species displays an amalgamation of characters present in various geocorine genera combined with the unusually strongly incrassate fore femora which is to be considered as autapomorphy in terms of the subfamily, thus its placement in Geocoris s. str. is not justified. Considering the above it is to be concluded that the species represents an extinct lineage of geocorine true bugs which lived during the Eocene epoch thus, the genus Eocenogeocoris Kóbor gen. nov. is suggested to be erected to accommodate the species.
Genus Geocoris Fallén, 1814
Type species: Cimex grylloides Linnaeus, 1761 (= Geocoris grylloides grylloides), fixed by Oshanin, 1912.
Subgenus Geocoris Fallén, 1814
Type species: As for genus.
Geocoris (Geocoris) monserrati Ortuño and Arillo, 1997
(Fig. 5)

Geocoris (Geocoris) monserrati Ortuño and Arillo, 1997 (holotype): a overview of fossil; b dorsal half; c ventral half (scale bar = 1 mm for images b and c).
Material examined: Holotype, MCNA: IZA-017, MCNA-5361.
Locality and horizon: Upper clayey member of the Izarra Formation, central part of the Basque-Cantabrian Basin, Spain; Oligocene epoch (33.9 – 23 Ma).
Redescription
Head pentagonal, eyes reniform, slightly stylate; ocular sulcus appears to be complete, but slightly defined; posterior edge of eyes touching anterior edges of pronotum. Ocelli situated near ocular sulcus, at base of vertex. Clypeus rounded, appearing to be exceeding the mandibular plates; margins of clypeus converging basally. Vertex with median longitudinal furrow extending from the middle of vertex to apex of clypeus. Antenniferous tubercle appears to be well-developed. Antennae tetramerous with antennomere I granuliform, shortest; antennomere II cylindrical, longest (but incomplete); antennomeres III and IV subequal; antennomere III cylindrical, antennomere IV fusiform. Labial furrow appears to be closed; labiomere I present but appears to be incomplete.
Thorax: Pronotum trapeziform, integument appears to be densely punctate except callosities, humeral angles and posterior margin. Pronotal callosities separated by punctures. Scutellum elongate triangular, integument punctate except well-developed median trifurcate carina. Clavus of hemelytron with margins converging towards apex; punctuation visible basally. Corium suspectedly macropterous or submacropterous (claval furrow present, clavus separable) with puctures along claval furrow and Cu. Thoracic pleurites desenly punctate except supracoxal lobes and surroundings of peritreme. Peritreme bulbous; orifice appears to be rounded. Femora of legs fusiform, fore femora slightly incrassate.
Abdomen: Characters of abdominal dorsum not observable. Ovipositor bisecting abdominal sternite VII.
Remarks: Ortuño and Arillo (1997) assigned the taxon to genus Geocoris and suggested a close relationship to G. ater (Fabricius, 1787), a species of subgenus Geocoris. According to our findings the genus assignment and the suspected relationship is established, thus G. monserrati is the first known representative of “modern geocorids”, i.e. Geocoris sensu lato and allied mono- and oligotypic genera.
Discussion
Superfamilial placement of Protogeocoris gen. nov.(Fig. 5)
The studied fossil possesses four-segmented piercing-sucking mouthpart i.e. rostrum and heteronomous forewing i.e. hemelytron consisting of clavus + corium and membrane, such characteristics are placing this insect in suborder Heteroptera (Schuh and Weirauch, 2020). The general appearance of the insect, especially the large and stylate compound eyes resemble to the representatives of Yuripopovinidae Azar et al., 2011 e. g. Caulisoculus Zhang and Chen, 2020 or Pseudocaulisoculus Kóbor and Roca-Cusachs, 2021. However, the venation of hind consists of three veins from which M-R are basally fused and venation of membrane is simplified, i.e. lacking closed cells. Contrastingly, hemelytron of the representatives of Yuripopovinidae possess three separate, longitudinal veins forming two closed cells, venation of membrane with closed cells. Furthermore, eyes unequivocally appear to be reniform, not rounded as in Yuripopovinidae. The presence of ocelli, incrassate fore femora and simplified venation of hemelytron with membrane lacking closed cells are shared synapomorphies of superfamily Lygaeoidea (Henry 1997). The studied fossil possesses all three of these characteristics, thus we ascribe this taxon to superfamily Lygaeoidea.
Placement of Protogeocoris gen. nov. within Lygaeoidea
This lygaeoid bug has pentagonal head with wide vertex and large, reniform, moderately stylate compound eyes which are characteristic to the family Geocoridae. Besides the arrangement of head and eyes, this peculiar true bug family is characterised by the dorsally situated abdominal spiracles II-IV and medially curved sutures between abdominal tergites 4/5-5/6 (Henry 1997) Abdominal sutures 4/5-5/6 appear to be slightly curved in Protogeocoris gen. nov. The degree of curvature of abdominal sutures considerably varies among geocorid taxa as demonstrated in Fig. 6a-b. Though abdominal spiracles are obscured in case of the specimen, supportive characters are to be observed which are displayed by various representatives of Geocoridae, e.g. placement of ocelli or trapeziform pronotum with furrow-like callosities (similar as in Unicageocoris Malipatil, 2013 and Nannogermalus Kóbor and Kondorosy, 2020).

Morphological characteristics of extant representatives of subfamily Geocorinae discussed in present study (indicated by arrow): a-b sutures of abdominal tergites 4/5-5/6 in Germalus victoriae Bergroth, 1895 (a) and Geocoris willeyi Kirkaldy, 1905 (b); c-d remnants of prothoracic collar (col) in Geocoris sp. in dorsal (c) and lateral view (d) (images not to scale).
The proportion of diameter of compound eyes to head length is similar as in subfamily Geocorinae Baerensprung, 1860. This similarity is supported by the arrangement of head which resembles to genus Germalus Stål, 1862 and allied genera. Furthermore, these geocorine taxa are characterised with subequilateral triangular scutellum and clavus with margins parallel and claval commissure present. Prothoracic collar as separate structure can be observed in genus Epipolops Herrich-Schaeffer, 1850 of subfamily Pamphantinae Barber and Bruner, 1933 (Rengifo-Correa et al. 2014). However, traces of such structure can be found in form of slightly elevated, impunctate region near the anterior margin of pronotum and prosternum in several species of subfamily Geocorinae (senior author’s unpublished notes, Fig. 6c-d). Based on the above discussed evidence we suggest ascribing this new true bug taxon in subfamily Geocorinae.
Relationship of Protogeocoris gen. nov. to other representatives of Geocorinae
The tribal classification of Geocorinae was proposed and later elaborated by Montandon (1907, 1913). Further improvement of the classification was later suggested by Parshley (1921); however, no revised classification was proposed subsequently. Though Montandon’s concept gained no further acceptance by the community, the additional evidence provided by the study of Slater and Hurlbutt (1957) on the morphology of the metathoracic wing of Lygaeoidea, along with the senior author’s unpublished findings suggests that two distinct major lineages are to be recognised within Geocorinae which are partly consonant with Montandon’s tribes. However, there are genera which display transitions or unique states of characters, thus cannot be classified in these two groups e.g. Apennocoris Montandon, 1907; Stylogeocoris Montandon, 1913 or Nannogermalus Kóbor and Kondorosy, 2020. Based on the comparison of diagnostic characters of these groups (Table 1) and the unique character state of prothorax with distinct collar it is to be assumed that Proto-geocoris represents a separate, extinct lineage within the subfamily. The basally fused and apically branching R-M of hemelytron can be observed in multiple geocorid genera e.g. Engistus Fieber, 1864 (Henestarinae) or Umbrageocoris Kóbor, 2019 (Geocorinae) and the exact systematic importance of the characteristics of the hemelytron is subject of further comparative study.
Evolutionary significance of fossil big-eyed bugs
According to Ross (2019, 2022), Protogeocoris is the first representative of the superfamily Lygaeoidea reported from mid-Creatceous Burmese amber and the oldest known taxon of family Geocoridae. Until now, only two extinct representatives of the family were known from compression and mould fossils: Eocenogeocoris infernorum (Scudder, 1890) from the Eocene (Florissant formation, approximately 34 Ma.); and Geocoris monserrati Ortuño and Arillo, 1997 from the Oligocene (Izarra formation, approximately 33.9 to 23.03 Ma.).
The original description of E. infernorum is cursory and the illustrations are lacking detail. It must be noted that characterisation and diagnosis of compression fossils is problematic in most cases (Azar et al. 2011; Chen et al. 2016). The examination of the available photo documentation of the fossils suggest that the species clearly identifies as a representative of family Geocorinae and can be considered as a representative of an extinct of lineage displaying a combination of characteristics of various geocorine genera and possessing a suspected autapomorphy, strongly incrassate fore femora, thus Eocenogeocoris gen. nov. is erected to accommodate the species. G. monserrati is described and illustrated accurately and it is to be confirmed that the placement suggested by Ortuño and Arillo (1997), i.e. the species is closely related to the extant species, Geocoris ater (Fabricius, 1787) and its Mediterranean and Palaearctic relatives, is justified.
The fossil record of big-eyed bugs currently consists of three distinct taxa from three different geographic era: Protogeocoris arcanus from the mid-Cretaceous, Eocenogeocoris infernorum from the late Eocene and Geocoris monserrati from the Oligocene. The two oldest of them, P. arcanus and E. infernorum, represent extinct lineages whilst G. monserrati is unambiguously displaying characteristics of the “modern” representatives of the subfamily Geocorinae, more specifically the genus Geocoris sensu stricto. This suggests that the major synapomorphies characterising the family Geocoridae, e.g. the pentagonal head with variably stylate, reniform eyes have been evolved until the lowermost-Cenomanian age of the mid-Cretaceous, thus the origin of geocorid subfamilies and characteristic lineages can be dated back to this epoch. Other character states e.g. the reduction of claval commissure or the variability in the degree of curvature of the sutures of abdominal sutures 4/5-5/6 (Fig. 6a-b), have been developed more recently and the taxa displaying character states which are suspectedly derived, i.e. the “geocorine lineage” (Geocoris sensu lato and closely allied mono- and oligotypic genera) appear latest in the Oligocene. The majority of geocorid species are representing this lineage. The species-level delimitation of these taxa is often problematic because the species can be diagnosed by a set of minor characters, which often display intraspecific variability of various degree (senior author’s unpublished findings). The Eocene-Oligocene boundary witnessed a cooling event, which resulted significant changes in the taxonomic composition, richness, and abundance distribution of insect groups (Smith et al. 2014). Taking the above into account it is to be assumed that the diversity of the “geocorine lineage” is resulted by a quick radiation event that can be dated back to the Oligocene epoch and the “uniform heterogeneity” can be attributed to the relatively young age (< 33.9 Ma) of these taxa. However, it must be noted that this hypothesis is based on indirect evidence and its proof requires further investigations. A promising method of proof is performing morphology-based phylogenetic reconstructions involving both extant and fossil taxa. Including fossil records in morphological phylogenies of extant taxa is considered to improve the accuracy of reconstructions and to test evolutionary hypotheses even if the data acquired from the study of fossils is incomplete or partly incorrect (Edgecombe 2010; Mongiardino Koch et al. 2021).
Data availability
Data sharing not applicable to this article as no datasets were generated or analysed during the current study.
Change history
15 May 2024
A Correction to this paper has been published: https://doi.org/10.1007/s12549-024-00609-2
References
Azar, D., Nel, A., Engel, M., Garrouste, R., & Matocq, A. (2011). A new family of Coreoidea from the Lower Cretaceous Lebanese Amber (Hemiptera: Pentatomomorpha). Polish Journal of Entomology, 80(4), 627–644. https://doi.org/10.2478/v10200-011-0049-5
Baerensprung F. V. (1860). Hemiptera Heteroptera Europaea systematice disposita. Berliner Entomologische Zeitschrift, 4 (appendix), 1–25.
Bai, M., Beutel, R.G., Klass, K.D., Zhang, W.W., Yang, X.K., & Wipfler, B. (2016). Alienoptera - a new insect order in the roach-mantodean twilight zone. Gondwana Research 39, 317e326. https://doi.org/10.1016/j.gr.2016.02.002
Chen, J., Wang, B., & Jarzembowski, E.A. (2016). Palaeontology: benefits of trade in amber fossils. Nature, 532, 441. https://doi.org/10.1038/532441a
Christiansen, K., & Nascimbene, P. (2006). Collembola (Arthropoda Hexapoda) from the mid Cretaceous of Myanmar (Burma). Cretaceous Research, 27(3), 318-363. https://doi.org/10.1016/j.cretres.2005.07.003.
Cruickshank, R.D., & Ko, K. 2003. Geology of an amber locality in the Hukawng Valley, northern Myanmar. Journal of Asian Earth Sciences 21, 441e455. https://doi.org/10.1016/s1367-9120(02)00044-5
Edgecombe, G. D. (2010). Palaeomorphology: fossils and the inference of cladistic relationships. Acta Zoologica, 91(1), 72-80. https://doi.org/10.1111/j.1463-6395.2009.00426.x
Fallen, C.F. (1814). Specimen novam Hemiptera disponendi methodum exhibens (dissertation). p. 10. https://doi.org/10.5962/bhl.title.106859
Grimaldi, D., Engel, M.S., & Nascimbene, P.C. (2002). Fossiliferous Cretaceous amber from Myanmar (Burma): its rediscovery, biotic diversity, and paleontological significance. American Museum Novitates, 3361, 1e71. https://doi.org/10.1206/0003-0082(2002)361%3C0001:fcafmb%3E2.0.co;2
Henry, T. J. (1997). Phylogenetic analysis offamily groups within the infraorder Pentatomomorpha (Hemiptera: Heteroptera), with emphasis on the Lygaeoidea. Annals of the Entomological Society of America, 90(3), 275–301. https://doi.org/10.1093/aesa/90.3.275
Jiang, T., Szwedo, J., & Wang, B. (2019). A unique camouflaged mimarachnid planthopper from mid-Cretaceous Burmese amber. Scientific Reports, 9(1), 1-11.
Kóbor, P. (2019). Umbrageocoris kondorosyi: a new genus and species of big-eyed bugs from New Guinea (Heteroptera: Lygaeoidea: Geocoridae). Acta Zoologica Academiae Scientiarum Hungaricae, 65(1), 1–8. https://doi.org/10.17109/azh.65.1.1.2019
Kóbor, P. (2020). A review on biology and agricultural significance of big-eyed bugs (Heteroptera: Lygaeoidea: Geocoridae). Hungarian Agricultural Research, 2020/2, 5–11.
Kóbor, P., & Kondorosy, E. (2020). Nannogermalus marmoratus: a new endemic big-eyed bug from New Caledonia (Heteroptera: Lygaeoidea: Geocorinae). Acta Zoologica Academiae Scientiarum Hungaricae, 66(4), 361–368. https://doi.org/10.17109/azh.66.4.361.2020.
Kóbor, P., & Roca-Cusachs, M. (2021). Pseudocaulisoculus longicornis: description of a new yuripopovinid true bug from mid-Cretaceous Burmese amber (Heteroptera: Coreoidea: Yuripopovinidae). Cretaceous Research, 125, 104849.
Latreille, P.A. (1810). Considérations générales sur l’ordre natureldes animaux composant les classes des Crustacés, des Arachnides, et des Insectes; avec un tableau méthodique deleurs genres, disposés en familles (pp. 1–444). Paris: F. Schoell, https://doi.org/10.5962/bhl.title.39620.
Leston, D., Pendergrast, J. G., & Southwood, T. R. E. (1954). Classification of the terrestrial Heteroptera (Geocorisae). Nature, 174(4419), 91-92. https://doi.org/10.1038/174091b0.
Linnaeus, C. (1758). Systema Naturae per regna tria naturae,secundum classes, ordines, genera, species cum characteribus,differentiis, synonymis, locis. Tomus I. Editio decima,reformata (pp. 1-823). Homiae: Laurentii Salvii, https://doi.org/10.5962/bhl.title.542.
Lis, J. A., Lis, B., & Heiss, E. (2018). Chilamnestocoris mixtus gen. et spec. nov., the first burrower bug (Hemiptera: Pentatomoidea: Cydnidae) in Upper Cretaceous Burmese amber. Cretaceous Research, 91, 257–262. https://doi.org/10.1016/j.cretres.2018.06.017
Lis, J. A, Roca-Cusachs, M., Lis, B., & Jung, S. (2020). Pullneyocoris dentatus gen. et sp. nov. (Hemiptera: Pentatomoidea: Cydnidae) the third representative of the subfamily Amnestinae from mid-Cretaceous amber of northern Myanmar. Cretaceous Research 115104562-S0195667120302482 104562. https://doi.org/10.1016/j.cretres.2020.104562.
Malipatil, M. B. (1994). Revision of Australian Geocoris Fallen and Stylogeocoris Montandon (Heteroptera: Lygaeidae: Geocorinae). Invertebrate Systematics, 8(2), 299–327. https://doi.org/10.1071/it9940299.
Malipatil, M. B., & Blackett, M. J. (2013). Review and revision of Australian Germalus Stål, with new genera and further new species of Australian Geocorinae (Hemiptera: Heteroptera: Geocoridae). Zootaxa, 3746(2): 257–300. https://doi.org/10.11646/zootaxa.3746.2.3.
Mongiardino Koch N., Garwood R. J., & Parry L. A. (2021). Fossils improve phylogenetic analyses of morphological characters. Proceedings of the Royal Society B: Biological Sciences, 288: 20210044. https://doi.org/10.1098/rspb.2021.0044.
Montandon, A. L. (1907). Nouveaux genres et espèces du groupe des Geocorinae. Annales Historico-Naturales Musei Nationalis Hungarici, 5, 89–97.
Montandon, A. L. (1913). Nouvelles contributions à l’étude des Geocorinae. Buletinul Societății Române de Științe București-România/Bulletin de la Société Roumaine des Sciences Bucarest-Roumanie, 22(2/3), 249–252.
Ortuño, V. M., & Arillo, A. (1997). Presencia del género Geocoris en el Oligoceno de Izarra (Álava, España). Descripción de Geocoris monserrati sp. nov.(Heteroptera, Lygaeidae). Nouvelle Revue d’Entomologie (NS), 14(4), 359–363.
Parshley, H. M. (1921). The Montandon Collection of Geocorinae (Lygaeidae, Hemiptera). Bulletin of the Brooklyn Entomological Society,16, 20–21.
Readio, J., & Sweet, M. H. (1982). A review of the Geocorinae of the United States east of the 100th meridian (Hemiptera: Lygaeidae). Miscellaneous Publications of the Entomological Society of America, 12, 1–91.
Rengifo-Correa, L., Brailovsky, H., Henry, T. J., & Morrone, J. J. (2014). Phylogenetics and evolutionary morphology of the Neotropical true bug genus Epipolops (Hemiptera: Heteroptera: Geocoridae). Systematic Entomology 39(1), 127–140. https://doi.org/10.1111/syen.12039
Ross, A. J. (2019). Burmese (Myanmar) amber checklist and bibliography 2018. Palaeoentomology, 2, 22e84. https://doi.org/10.11646/palaeoentomology.2.1.5
Ross, A. J. (2022). Supplement to the Burmese (Myanmar) amber checklist and bibliography, 2021. https://doi.org/10.11646/palaeoentomology.5.1.4
Schilling P. S. (1829). Hemiptera Heteroptera Silesiae systematice disposuit Schilling. Beiträge zur Entomologie, Berlin, 1, 34–92.
Schuh, R. T., & Weirauch, C. (2020): True bugs of the World (Hemiptera: Heteroptera): Classification and Natural History (Second Edition). Siri Scientific Press, Rochdale, United Kingdom.
Scudder, S. H. (1890). The Tertiary Insects of North America. Report of the United States Geological Survey of the Territories, 3,1–734. https://doi.org/10.5962/bhl.title.44698
Slater, J. A. (1977). The incidence and evolutionary significance of wing polymorphism in lygaeid bugs with particular refer-ence to those of South Africa. Biotropica, 217–229.
Slater, J. A., & Hurlbutt, H. W. (1957). A comparative study of the metathoracic wing in the family Lygaeidae (Hemiptera: Heteroptera). Proceedings of the Entomological Society of Washington, 59(2), 67–79.
Smith, D.M., Leckey, E. H., Walker, L. J., & Nufio, C. R. (2014). Insect response to Eocene-Oligocene Climate Change in Colorado, USA. In C. Simpson (Ed.), The Paleontological Society Special Publications, 13, 59–60.
Sweet, M. H. (2000). Economic Importance of Predation by Big-Eyed Bugs (Geocoridae). In C. W. Schaefer, & A. R. Panizzi, (Eds.) Heteroptera of economic importance (pp. 735–746). Boca Raton: CRC Press. https://doi.org/10.1201/9781420041859-32.
Tsai, J. F., Yang, M. M., Rédei, D., & Yeh, G. F. (2011). Jewel bugs of Taiwan (Heteroptera: Scutelleridae). Taichung: National Chung Hsing University.
Weirauch, C., & Schuh, R. T. (2011). Systematics and evolution of Heteroptera: 25 years of progress. Annual review of Entomology, 56, 487–510. https://doi.org/10.1146/annurev-ento-120709-144833.
Zhang, J., & Chen, J. (2020). A stalk-eyed true bug in mid-Cretaceous Burmese amber (Hemiptera, Pentatomomorpha, Yuripopovinidae). Cretaceous Research, 110, 104391.
Acknowledgements
The authors would like to express their gratitude towards Jesus Alonso Ramirez, curator at MCNA for locating and documenting holotype of G. monserrati and Professor Brian D. Farrell at MCZN for providing access to the photodocumentation of E. infernorum. Furthermore, we would like to thank the work of two anonymous reviewers for their help in the improvement of the manuscript.
Funding
Open access funding provided by ELKH Centre for Agricultural Research. Partial financial support was received from Project ANID-SA77210055 (Convocaria Nacional Subvención Instalación en la Academia).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. The specimens studied were examined by Péter Kóbor and Eudardo I. Faúndez. Photomicrographs were done by Eudardo I. Faúndez, line drawings were done by Marcos Roca-Cusachs. The first draft of the manuscript was written by Péter Kóbor and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is registered in Zoobank under: http://zoobank.org/urn:lsid:zoobank.org:pub:53B200AA-8B48-43C8-83A9-A83253DF6DC6
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kóbor, P., Faúndez, E.I. & Roca-Cusachs, M. The revision of fossil big-eyed bugs suggests a peculiar evolutionary history of a peculiar true bug family (Heteroptera: Lygaeoidea: Geocoridae). Palaeobio Palaeoenv 103, 531–543 (2023). https://doi.org/10.1007/s12549-022-00567-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-022-00567-7