
Abstract
The assemblage of small mammals from a site near Yeniköy (Anatolia) is described. The assemblage was collected by screen-washing a sample from a locality in the Selimye Formation (Sivas basin). The site has a magneto-stratigraphic calibrated age of 29 Ma (younger part of the early Oligocene). The murids dominate in this relatively small collection with Eucricetodon and Pseudocricetodon. In addition, a dipodid, an unidentified erinaceide and the new baluchimyin hystricognath genus and species Zorania milosi nov. gen. et nov. sp. are present. The study of Eucricetodon, Pseudocricetodon and Zorania nov. sp. includes an analysis of incisor enamel microstructure. The incisor microstructure of Eucricetodon shows that there are several parallel long-living lineages within the genus. The Anatolia-Balkans biogeography of the late Eocene-Oligocene is reviewed. Its rodent assemblages are characterised by dominance of murids and presence of taxa that are known from low-latitude Asia. The Paleogene Anatolia-Balkans biogeographic province, is therefore rather different from those of western European and the Indian subcontinent.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Palaeogeographic maps of the late Eocene-early Oligocene show that the Balkans and Anatolia formed a composite continental terrane, located south of the Paratethys sea and north of the Neotethys ocean. This area was part of a more or less interconnected stretch of land between Europe and the Indian sub-continent. Sea straits crossed this Anatolian-Balkans terrane at times, connecting the Paratethys seas to the Neotethys ocean. But, there is no consensus on location and timing, the published palaeogeographic maps show different configurations (Meulenkamp and Sissingh 2003, Popov et al. 2004, Gomes Rodrigues et al. 2012, Ozsvárt et al. 2016, Palcu and Krijgsman 2021).
In recent years several Oligocene and late Eocene assemblages of small mammals from Anatolia and from Serbia have been described (de Bruijn et al. 2003, 2017; Marković et al. 2017; Wessels et al. 2019; van de Weerd et al. 2021a, 2021b). These assemblages show compositional differences despite the relatively close proximity of the sites, indeed suggesting presence of geographic barriers. However, the fossil record for the different regions is far from complete as shown by recent chronostratigraphic schemes of the Balkans, Thrace basin and Anatolia (van de Weerd et al. 2021a, 2021b) illustrating the gaps in our knowledge and the age uncertainty ranges.
The Serbian and Anatolian assemblages contain many faunal elements unknown to Western and Central Europe. These assemblages, of which many species have Asian affinities, show that the Anatolia-Balkans land area is a complex biogeographic province separated from both Europe and from Asia, which is consistent with the palaeogeographic reconstructions put together by geologists.
The small mammals studied so far show that major differences in faunal composition between Western-Central Europe and Anatolia-Balkan persisted, testifying that the Alpine region endured as a barrier to mammal dispersal till late in the Oligocene. However, much more data are needed to complete this complex picture.
Here, we describe another part of the jigsaw puzzle: the small assemblage from the early Oligocene near Yeniköy (Sivas basin) in Anatolia (Fig. 1). It is one of the six late Eocene-early Oligocene faunas from this area known to date. It was found over thirty years ago; preliminary publications (Sümengen et al. 1990; Ünay et al. 2003) revealed its overall composition and biostratigraphic position. The assemblage is small with about fifty-five more or less complete teeth, but it is chronologically important because it has been dated magneto-stratigraphically (Krijgsman et al. 1996). Together with the nearby Inkonak site, Yeniköy can be positioned in the lithostratigraphic succession of the Sivas basin (Poisson et al. 1996, 2016). Last but not least, a faunal element in the Yeniköy association, previous thought to be a ctenodactyl, is now described as a new genus and species, Zorania milosi, and its taxonomic position as an hystricognath with Asian affinity has been established.

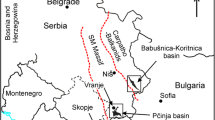
Palaeogeography of the Sivas basin for the middle part of the Oligocene (modified from Poisson et al. 2016, fig. 12d). Shown are major towns, the location of the sampled sites of Yeniköy and Inkonak (stars). The thin red broken lines indicate the boundaries between the main crustal blocks of Anatolia (Pontides, south Taurus, north Taurus, Kirşehir), the thick broken line is a major fault zone. The map differentiates between areas with shallow-marine deposits, areas with non-marine deposits (fluvial, fluvio-lacustrine and fluvio-deltaic. Selimye and Karayün Formations) and areas where Oligocene deposition did not occur. The inset map shows Turkey and the location of the palaeogeographic map
The Yeniköy and Inkonak sites in the Sivas basin
The large Sivas basin (Fig. 1) located east of Ankara, is the largest Cenozoic basin in the Anatolian interior. It has been extensively studied because of the spectacularly exposed Oligocene syn-depositional tectonic structures in its central part (Ribes et al. 2015; Kergaravat et al. 2016, 2017; Legeay et al. 2019). The Yeniköy and Inkonak sites with Oligocene vertebrates are situated south of the Tecer (=Deliler) Fault zone, near the south-western and southern margins of the basin where syndepositional tectonism has not been observed. The lithological succession of the Cenozoic fill of the Sivas basin near its southern margin may be simplified as follows. The basal unit is the Bozbel Formation, consisting of at least 2000m of deep-marine turbidites and volcano-clastic debris flows of Paleocene-Eocene age. The upper part of this formation is shallow-marine near Ortaköy (Gündogan et al. 2005). These deposits are overlain by late Eocene-early Oligocene evaporitic deposits included in the Hafik Formation (Poisson et al. 2016), other authors name it the Tuzhisar Formation (Pichat et al. 2016; Kergaravat et al. 2017). This evaporitic unit was thick in the central part of the Sivas basin, where it also included halite, responsible for intense syn-depositional deformation of the Oligocene (Ribes et al. 2015; Kergaravat et al. 2016, 2017; Legeay et al. 2018). The late Eocene-early Oligocene evaporitic unit is much thinner near the southern margin of the Sivas basin; only 90m has been logged near Yeniköy (Gündogan et al. 2005). The evaporitic formation is overlain by a thick succession of Oligocene fluviatile and limnic reddish silty-clays and sandstones (Fig. 1), these are included in the Selimye Formation and in its lateral equivalent the Karayün Formation. Lithologically these two units are similar, but the Karayün Formation, located in the centre of the Sivas basin, is characterised by syn-depositional deformation (Ribes et al. 2015; Kergaravat et al. 2016, 2017; Legeay et al. 2018). Estimates for the thickness of the Selimiye Formation are between 1500 and 2400m. The Oligocene Selimye Formation is stratigraphically the highest Tertiary unit in the southern part of the Sivas basin.
The Yeniköy vertebrate site is in the Selimye Formation near Ortaköy, in the south-western part of the Sivas basin (Fig. 1). Cater (1991), Gündogan et al. (2005) and Poisson et al. (2016) logged sections near Ortaköy. The late Eocene-early Oligocene gypsiferous unit wedges out near Yeniköy, it is absent east of the village. An about 800 m thick section near Yeniköy was measured and sampled for magneto-stratigraphy in the Selimye Formation during 1986-1987 by a joint team of the General Directorate of Mineral Research Exploration in Turkey (MTA) and the University of Utrecht (Krijgsnan et al. 1996). The location with coordinates 39°6'59.19"N; 36°22'58.38"E. is shown in Fig. 2. Near the middle of the section several test samples were taken, washed and the residue sorted. About 700 kg sediment was taken and processed from the best test sample. The small mammals obtained from the different samples do not show compositional differences and are not differentiated in this report.

Satellite image of the Yeniköy area, showing the town and the sampled Yeniköy site (white arrow; 39° 6'59.19"N; 36°22'58.38"E). The indicated formations and faults are after Gündogan et al. (2005). Copyrights CNES / Airbus & Maxar Technologies
Krijgsman et al. (1996) placed the Yeniköy vertebrate sample site in the lower part of chron C10r, with an approximate age of 29 Ma (Vandenberghe et al. 2012). Preliminary reports on the small mammals from this site are published by Ünay and de Bruijn (1987), Sümengen et al. (1990) and Ünay et al. (2003).
The late Oligocene Inkonak MR6 vertebrate site is located about 68 km to the north-east of Yeniköy (Fig. 1) in the same Selimye Formation (Poisson et al. 2016). Peter Steffens (BGR Hannover) found the first rodents in the section in 1970. Subsequently a large vertebrate assemblage was collected in 1986-87 by a joint team of the University of Utrecht and the General Directorate of Mineral Research Exploration in Turkey (MTA). The sample was taken from fluvio-lacustrine deposits originally named the Dokmetas Member of the Cevizcik Formation (Sümengen et al. 1990; de Bruijn et al. 1992). The sampled site is located in the east wall of the road-cut in the highway between Sivas and Kangal (Fig. 1; coordinates 39° 19' 41.14"N; 37° 8' 50.71"E). Poisson et al. (op. cit., his fig. 3) logged near the Inkonak MR6 site approximately 800 m post-Eocene section and included the sediments in the Selimye Formation. The magneto-stratigraphic investigation of the section with the vertebrate site did not result in an unambiguous correlation to the GTPS scale (Krijgsman et al. 1996). The vertebrate assemblage was initially considered earliest Miocene based on the evolutionary level of its faunal elements (de Bruijn et al. 1992), but was later revised to late Oligocene (Krijgsman et al. 1996; Ünay et al. 2003). To our knowledge no further mammals were collected from the Oligocene Selimye Formation after 1988-89, but it is certainly expected that detailed prospecting in the Selimye Formation will yield more strata with fossil small mammals.
Methods
The Yeniköy assemblage has a limited number of taxa, and Eucricetodon is dominant. Therefore, incisors can be relatively easily linked with the isolated molars of the same species. This prompted the second author to the study of incisors from Yeniköy during 2003. The upper and lower incisor of Eucricetodon and the lower incisor of Pseudocricetodon. were longitudinally and transversely sectioned. Because the original photographs of these specimens could not be located, these samples were re-photographed and studied during 2021 together with the lower incisor of Zorania milosi nov. gen. nov. sp. In order to take away doubts about the correct association of molars and incisors we prepared another set of incisors during 2022 and studied these too. These newly prepared samples confirmed that the original allocation to genus was correct. The incisors were studied and photographed with a Zeiss EVO15 scanning electron microscope of Utrecht University with magnifications up to 1250x and an acceleration voltage of 20 kV in HDBSD mode (high definition backscatter detection). Most of the illustrations of the molars from Yeniköy were on file, prepared years ago for preliminary publications and conferences, the first were published in Ünay and de Bruijn (1987). These are SEM images retouched by J. Luteyn. Despite the differences in grey tones, we decided to use these illustrations as is. A few photographs were taken recently with the JEOL Neoscope JCR6000. Lower case letters refer to the lower dentition, upper case letters refer to the upper dentition. Measurement are in millimeter units. Abbreviations in the descriptions are inferior (inf), superior (sup), number of specimens (N), standard deviation (SD), sinistral (sin) and dextral (dex). M1/2 indicates an M1 or M2. Abbreviations used in the description of the incisors: portio externe (PE), portio interne (PI), outer part portio externe (OPE), inner part portio externe (IPE), inner part portio interne (IPI), outerpart portio interne (OPI), Hunter-Schreger Bands (HSB), Intra Prismatic Matrix (IPM), portio externe without prisms (PLEX). The nomenclature of parts of Eucricetodon and Pseudocricetodon molars follows van de Weerd et al. (2021a, Fig. 12), that of the Dipodidae is after Daxner-Höck et al. (2014). The nomenclature of tooth parts of the Baluchimyinae are shown in the appropriate illustrations.
Systematic palaeontology
Suborder Supramyomorpha D’Elia, Fabre and Lessa, 2019
Infraorder Myodonta Schaub, 1958
Family Muridae Illiger, 1811
Subfamily Eucricetodontinae Mein and Freudenthal, 1971
Eucricetodon Thaler, 1966
Type species: Cricetodon collatum Schaub, 1925
Type locality and age: Küttigen, Germany; late Oligocene
Eucricetodon aff. ruber van de Weerd, de Bruijn and Wessels, 2021
(Figs. 3 a-k)
Type locality: Gözükızıllı-1b, late Oligocene
Synonomy: Eucricetodon Ünay and de Bruijn (1987)
Eucricetodon sp. 1 Ünay et al. (2003)
Measurements: see Table 1
Molars from Yeniköy: (code Y1):
M1sin: no. (101), (102). M1dex: no. (105), 106. (107), (108), 109, 110
M2sin: no. 111, 112, (113), (114), 115; M2dex: no. 121, 122, 123, 124, 125, 126, 127
M3sin: no. 131, 132, 133; M3dex: no. 136, 137, 138, 139, 140
m1sin: no. 141, (142), (143); m1dex: no. 146, 147
m2sin: no 151, 152, 153, 154, 155; m2dex: no. 161, (162), 163, 164, 165, 166
m3sin: no. (171), (172), (173); m3dex: no. (175), 176, 177, (178), 179, 180
(Incomplete or broken molars have numbers between brackets.)
Incisors from Yeniköy:
Prepared sample 27 (Y05), I sup longitudinal section.
Prepared sample 28 (Y05), I sup transverse section.
Prepared sample 29 (Y04), I inf longitudinal section.
Prepared sample 30 (Y04A), I inf transverse section.
Description: The molars (Fig. 3) are identical in morphology to those of Eucricetodon ruber from Gözükızıllı. There are minor differences in morphotype composition between the two assemblages, M1 and M2 morphotypes with spurs on the paracone or ectolophs are present in Gözükızıllı but are absent in Yeniköy; all Yeniköy M1 have weak anterior cingulums. The Yeniköy molars are on average a bit smaller (Fig. 4).

Eucricetodon aff. ruber from Yeniköy. a M3 sin Y1-132, b M2 dex Y1-126, c M1 dex Y1-109, d M3 dex Y1-138, e M2 dex Y1-122, f M1 dex. Y1-110, g m3 dex Y1-177, h m2 sin Y1-154, i m1 dex Y1-146, j m2 dex Y1-161, k m1 dex Y1-147

Measurements of Eucricetodon aff. ruber from Yeniköy compared with E. occulatus and E. ruber from Gözükızıllı. Similar scatter diagrams but without the Yeniköy specimens are shown in van de Weerd et al. (2021a)
I inf: (Fig. 5) The incisor material from Yeniköy consists of isolated and broken specimens. Transverse and longitudinal sections of the Eucricetodon lower incisor are shown in Fig. 5, the exterior side of the lower incisor is in Fig. 5c. The enamel surface shows a central rib (red arrow in Fig. 5) and two parallel less pronounced ribs. Enamel thickness is about 50 ųm at the level of the central rib, there the relative thickness of the portio externe (PE) is ~ 28%, that of the portio interne (PI) is ~72%. The PE is in two parts, the outer part (OPE) has no prisms (PLEX), the inner part (IPE) is tangential. The portio Interne (PI) consists of longitudinal HSB (Hunter-Schreger Bands). The outer part of the portio interne (OPI) has prism-parallel IPM (Intra Prismatic Matrix), in its inner part (IPI) the IPM is at an angle to the prisms. A reversal zone (syncline) is present below the median rib. This agrees with schmelzmuster type 10 of Kalthoff (2000).

Photographs of sections and exterior view of the lower incisor of Eucricetodon from Yeniköy: a, b longitudinal section of prepared sample 29; d, e cross section of prepared sample 30; c shows the anterior side of the incisor. The arrows in a and c point at the ribs, the reversal zone is below the red arrow. The white arrow in d points toward the tip of the incisor
I sup: (Fig. 6) Transverse and longitudinal sections of the Eucricetodon upper incisor are shown in Fig. 6. The exterior surface may have some indistinct ribs. Enamel thickness is ~70 ųm near its mesial side, the portio externe (PE), at this location measures ~20 ųm (~30%) and the portio interne (PI) ~50 ųm (~70%). The cross section shows that the Hunter-Schreger Bands (HSB) have a longitudinal orientation close to the mesial side, laterally changing to transverse over a short distance. The IPM is prism-parallel, but some IPM is at an angle to the prisms in the inner part of the PI near the mesial side of the tooth (Fig. 6d). The PE shows radial enamel, but in the specimen of Fig. 6, it is tangential very close to the mesial side (right side Fig. 6c). The longitudinal section shows weekly inclined transverse HSB's (Fig. 6c, d) and radial enamel in the PE. The enamel microstructure in the central and lateral part of the incisor corresponds to schmelzmuster type 1 of Kalthoff (2000).

Photographs of sections of the upper incisor allocated to Eucricetodon. a longitudinal section of prepared sample 27; b, detail of a; c transverse section of prepared sample 28; d and e details of c.
Discussion
The two Eucricetodon species E. ruber and E. occulatus from the late Oligocene of Gözükızıllı (Anatolia; van de Weerd et al. 2021a) differ in size only (Fig. 4). Eucricetodon from Yeniköy corresponds in size with E. ruber from Gözükızıllı (Fig. 4). However, one m2 in the Yeniköy assemblage (Fig. 4, no Y1-164) suggests that a smaller species could be present. The assemblages of E. ruber and E. occulatus from Gözükızıllı contain several M1 and M2 with ectolophs or spurs on the paracone, while none of the molars from Yeniköy show such features. Otherwise the morphology of the Eucricetodon molars is the same in the two sites.
Enamel microstructure of the incisors in Eucricetodon
In assemblages of isolated teeth it is tricky to associate incisors with molars of the same species. But in the small assemblage of Yeniköy with only a few murid taxa and dominance of one taxon, there is no uncertainty that the incisors are correctly associated to the Eucricetodon molars. As an extra check we have picked some other specimens from the Yeniköy assemblage and studied these using the SEM; the new sections show the same incisor structure. The derived type 10 enamel of the sectioned lower incisor allocated to Eucricetodon in Yeniköy is same as in those allocated to Eucricetodon in Gözükızıllı. However, there are minor differences in the development of the ridges of the exterior surface: there are two ridges in Gözükızıllı (van de Weerd et al. 2021a) and three in Yeniköy. Considering that in Gözükızıllı, the studied incisors were also not found in direct association with the molars, we double checked here as well. Lower incisors of murids are more progressive in structure than the upper incisor of the same species, that is why, in general, it is sufficient to investigate the lower incisor only. However, the upper incisor of Eucricetodon from Yeniköy warrants illustration (Fig. 6). It shows transverse HSB's (the primordial orientation in murids), but close to the mesial side these rotate to longitudinal (the derived orientation in murids). Kalthoff (2000) in her Fig. 7 mentioned HSB rotation in the incisor, but not in the manner observed here.

Pseudocricetodon sp. M1 and m2 from Yeniköy compared to selected species from the literature. The molars are shown at about the same length, the corresponding approximate enlargement factor is indicated between brackets; P. nawabi is the largest (28x), P. philippi the smallest (45x). P. philippi and P. orientalis are from the "middle" Oligocene of Kocayarma (Thrace basin; Ünay-Bayraktar, 1989); P. montalbanensis is from the early Oligocene of Montalban (Spain; Utrecht collection); P. heissigi is from the early Oligocene of Strelac 3 (Serbia; Marković et al. 2019) and P. nawabi from the "middle" Oligocene of Pakistan (Marivaux et al. 1999)
Van de Weerd et al. (2021a) reviewed history and status of Eucricetodon and the primitive small-sized early Oligocene species included in Atavocricetodon by Freudenthal (1996). Forty species were listed and two new species were added by these authors. De Bruijn et al. (2003) followed Freudenthal in grouping small primitive eucricetodontinids in Atavocricetodon, but considered this genus a “grade rather than a clade”. De Bruijn et al. (2003) included the much older late Eocene Eucricetodon leptaleos Wang and Meng, 1986 (Yunnan, southern China) in Atavocricetodon. Eucricetodon leptaleos is from sites in the Caijichong Formation considered Ulangochuan (39.9-37.2 Ma, see Wang et al. 2019). It seems to us that other late Eocene Asian species, such as Eucricetodon wangae Li, Meng and Wang, 2016 from northern China can be included too in Atavocricetodon. Considered this way, Atavocricetodon forms a group of late Eocene-early Oligocene primitive murids occurring in Asia. Atavocricetodon probably migrated from Asia into Anatolia using the land connection south of the Paratethys seas; it is present in the latest Eocene in the Anatolian site of Süngülü (de Bruijn et al. 2003). In the Anatolia-Balkans province Atavocricetodon was succeeded by Eucricetodon as evidenced in Yeniköy (this paper), Gözükızıllı (van de Weerd 2021a), the Thrace basin (Unay-Baytraktar, 1989) and Ugljevik (van de Weerd 2021b). Atavocricetodon reached western Europe from Asia immediately after the "grande coupure" (approximately at the Eocene-Oligocene boundary, Hooker et al. 2004), most likely using the land connection north of the Paratethys seas. In the earliest Oligocene site of Hoogbutsel (Belgium) Atavocricetodon is present with three species (Freudenthal 1988). Afterwards, Atavocricetodon is succeeded in Europe by more advanced species included in Eucricetodon, that migrated into the area during MP24 (Freudenthal and Martín-Suárez 2016).
The enamel microstructure of the lower incisor of species included in Atavocricetodon and Eucricetodon that have been investigated up to now are in Table 2. The table shows that groups of species can be distinguished. Early Oligocene (MP 22/23) Atavocricetodon atavus from Europe has type 1 of Kalthoff (2000). Type 1 is the primitive murid enamel structure, observed as well in Pappocricetodon antiquus from its middle Eocene type locality in China (Kalthoff 2000). A second group of species (with enamel type 4) is formed by Eucricetodon from the late Oligocene to Early Miocene (MP28 to MN3) of Western Europe. This is the "collatus group" of Freudenthal and Martin-Suarez (2016) that also includes the type species of the genus, E. collatus (Schaub, 1925). However, the enamel of E. collatus has not yet been investigated. The third group (with the derived enamel type 10) is formed by Eucricetodon from the geographically distinct Anatolia-Balkans province (sites Yeniköy MP 24, Gözükızıllı MP25/26 and Ugljevik MP27/28) that have the derived enamel type 10. Other groups may be present as well, as is suggested by Eucricetodon cf. huberi from Belgaric-1 (Quercy, France, MP25) that has enamel type 8. Thus incisor microstructure clearly shows that there are several parallel long-living lineages within Eucricetodon, but whether these can be distinguished just by the molars remains to be seen. De Bruijn et al. (2003) suggested for Atavocricetodon that it is conceivable that the limits of palaeontological classification based on molars have been reached, this statement seems equally valid for Eucricetodon.
Subfamily Pseudocricetodontinae Engesser, 1987
Genus: Pseudocricetodon Thaler, 1969
Type species: Pseudocricetodon montalbanensis Thaler, 1969
Type locality: Montalban, Spain (early Oligocene, MP23)
Remarks: For a recent introduction to the subfamily and a review of the genus Pseudocricetodon we refer to Marković et al. (2019). Nine species are included in the genus, occurring from Pakistan to Spain. The species differ mainly in size with additionally subtle details in dental morphology (Marković et al. 2019). It is not possible to allocate the limited material from Yeniköy to species level.
Pseudocricetodon sp.
Synonymy: Pseudocricetodon sp. Ünay et al. (2003)
Molars from Yeniköy: one M1 sin (Y1-111) 1.62 x 1.07; one m2 dex (Y1-182) 1.25 x 1.03
Incisors from Yeniköy:
Prepared sample 31 (Y01), I inf. longitudinal section.
Prepared sample 32 (Y01A), I inf. transverse section.
Description
M1 (Fig. 7c): The anterocone shows a minor anterior cleavage, suggesting an incipient division into two cusps. The anterior arm of the protocone is short and ends free. The protolophule bends towards the posterior arm of the protocone. The mesoloph is of medium length and somewhat irregular.
m2 (Fig. 7i): The m2 is characterised by the long posterior arm of the protoconid and a long mesolophid.
I inf. (Fig. 8): The front side of the incisor (Fig. 8b) shows many fine parallel running ribs, that are obliquely orientated in a fairly regular fashion. Twelve ribs have been indicated by arrows on Fig. 8d. The median ribs are best developed, other ribs become weaker towards the lateral sides of the tooth. Enamel thickness (Fig. 8d, e) is about 90 ųm, with the relative thickness of the PE ~ 14% and that of the PE ~ 86%. The thin PE is tangential and there is no PLEX. The PI is in two parts, the outer part with prism-parallel IPM, the inner part with the IPM at an angle of about 45° to the prisms. There is no reversal zone (“Umbiegung”, syncline). This enamel microstructure corresponds to schmelzmuster type 7 of Kalthoff (2000).

The lower incisor of Pseudocricetodon from Yeniköy. a longitudinal section of prepared sample 32, c detail of a, b anterior side of the incisor; d transverse section of prepared sample 32, e detail of d. Arrow in a point to tip of the incisor, arrows in d indicate ribs
Discussion
The two specimens from Yeniköy are relatively large and plot outside the size cluster of the assemblage from Montalban (type locality of Pseudocricetodon montalbanensis) present at the University in Utrecht (Fig. 2 in Marković et al. 2019). The two molars are close in size to those of P. nawabi Marivaux, Vianey-Liaud and Welcomme, 1999. Figure 7 compares the two molars from Yeniköy with the M1 and m2 of a selection of species, including the genotype P. montalbanensis and several Oligocene species from Pakistan, Thrace basin and Serbia. The selection of species in the figure is incomplete, but nevertheless illustrates that differences in morphology between the species are minute. The incipient cleavage of the anterocone of the Yeniköy M1 and presence of two long central lophids in the m2 are features that are, as far as we know, not seen in other Pseudocricetodon assemblages.
The incisors of three species of Pseudocricetodon investigated by Kalthoff (2000) are all type 7, thus similar to the Yeniköy specimens. The regular (sub-parallel) oblique ornamentation of the ribs is distinctive and, as far as we know, has not yet been illustrated. By contrast, the murid genera with a multitude of ribs illustrated by Kalthoff (2000, fig. 42) all have a relatively irregular rib ornamentation.
Subfamily Pappocricetodontinae
Genus: Witenia de Bruijn, Ünay, Saraç and Yïlmaz, 2003
Type species: Witenia flava de Bruijn, Ünay, Saraç and Yïlmaz, 2003
Species included in Witenia: Witenia fusca de Bruijn, Ünay, Saraç and Yïlmaz, 2003; Witenia yolua Gomes Rodrigues, Marivaux and Vianey-Liaud, 2012, Witenia europea de Bruijn, Marković, Wessels and van de Weerd, 2018.
cf. Witenia sp.
Material from Yeniköy: one left M2 (Y188), worn and corroded, 2.34 x 2.02 mm.
Description: Because the molar is strongly corroded, possibly by stomach acids of a predator, few details of its morphology are preserved. The molar has four roots and is slightly longer than wide. The lingual anteroloph is not pronounced. Unlike in many murids, its lingual sinus is not curved forward, but the anterior side of the sinus is at a right angle to the lingual tooth border, while its posterior side is oblique.
Discussion
This M2 together with some fragments show presence of a very large murid in the Yeniköy assemblage. Although few details of its morphology are preserved, its large size allows a comparison with Witenia. a genus known from the late Eocene of Süngülü (de Bruijn et al. 2003) and the early Oligocene of southern Serbia (de Bruijn et al. 2018). The M2 from Yeniköy is intermediate in size between that of the two species from Süngülü.
Infraorder: Myodonta Schaub, 1958
Family: Dipodidae Fischer, 1817
Introduction
There is no consensus on the classification above the genus level in the Dipodidae, discussions on this can be found in Daxner-Höck (1999), de Bruijn et al. (2003) and Wessels et al. (2019). Wessels et al. (2019) compiled a list of thirteen valid Paleogene genera of the Dipodidae, of which Banyuesminthus Tong, 1997 should be deleted because it has been synonomized with Allosminthus (Daxner-Höck et al. 2014). Earliest Dipodidae are present with four genera in the middle and late Eocene of Central Asia: Primisminthus, Sinosminthus, Allosminthus and Heosminthus. The family radiates in the early Oligocene with five new genera, while Heosminthus and Allosminthus survived from the Eocene. The Oligocene expansion is well demonstrated by the rich assemblages collected in Mongolia (Valley of Lakes, Daxner-Höck et al. 2014). Ten out of the twelve Paleogene genera were shown to be present in the Valley of Lakes representing 22 species; only Sinosminthus and Gobisminthus were not observed.
Genus Shamosminthus Huang, 1992
Shamosminthus aff. sodovis (Daxner-Höck, 2001)
(Fig. 9a-d)
Synonymy: Shamosminthus cf. sodovis in Ünay et al. 2003
Parasminthus cf. parvulus Bohlin, 1946 in Ünay and de Bruijn 1987.
Material from Yeniköy: one m1 sin (Y1-16, 0.95 mm x 0.65 mm); one m2 sin (Y1-17, 0.85 x 0.64); one m3 sin (Y1-18, 0.57 x 0.56); one M3 dex (Y1-19, 0.55 x 0.61).
Description
m1 (Fig. 9c): The m1 bears a tiny anteroconid that is located slightly lingually. Metalophid and posterior arm of the protoconid are well developed, the posterior arm is connected to the triangular mesoconid. The lingual mesoconid corner-point extends into the mesolophid, the labial corner-point into a long ectomesolophid. There is no connection between mesoconid and the anterior arm of the hypoconid; the hypolophid is oriented in posterior direction. The posterolophid connection to the entoconid is low.

Dipodidae from Anatolia and the Balkans juxtaposed with material from type areas in China and Mongolia. a-d Shamosminthus aff. sodovis from Yeniköy; i-l Heosminthus from Süngülü (Anatolia, de Bruijn et al. 2003); m-p Heosminthus from Valnis (Serbia, Wessels et al. 2019); e-h Shamosminthus sodovis, from Taatsin Gol (Mongolia, Daxner-Höck et al. 2014); q-t Heosminthus primiverus from the Caijichong Formation (China, Wang 1985)
m2 (Fig. 9b): The anterior arm of the protoconid and the metalophid are connected to the anterolophid. The labial part of the anterolophid and the protosinusid are well developed. The small mesoconid has well developed anterior and posterior connections to the protoconid posterior arm and to the hypoconid anterior arm. There is a long mesolophid that connects to the metaconid. The posterolophid reaches the entoconid, closing fossettid IV at a rather low level.
m3 (Fig. 9a): The m3 was damaged after photography, the hypoconid disappeared. Similar to the m2, the labial part of the anterolophid and the protosinusid are well developed and the mesolophid is long. The fossettids have low lingual closing ridges.
M3 (Fig. 9d): The rather large sinus between protocone and hypocone is almost closed, leaving a small opening between protocone and hypocone. There are five transverse parallel ridges: anteroloph, protoloph 1, mesoloph, metaloph and posteroloph. The connection between mesocone and hypocone is almost transverse. The hypocone is relatively large. Low ridges close the labial fossettes.
Discussion
The allocation of the Yeniköy material to genus and species is largely based on the studies of the material collected in Mongolia (Daxner-Höck et al. 2014). The ten dipodid genera present in the Oligocene of the Valley of Lakes (Mongolia) have been divided into four "groups". The material from Yeniköy is morphologically close to Allosminthus and Shamosminthus constituting the early Oligocene "Allosminthus group". The teeth from Yeniköy combine the small size of A. minutus with the more advanced dental morphology of A. sodovis, hence the assignment A. aff. sodovis (Fig. 9). The m1 from Yeniköy has an ectomesolophid, a feature absent in A. sodovis from the Valley of Lakes.
Figure 9 also includes the dipodid species from Süngülü (latest Eocene, Anatolia) and Valnis (early Oligocene. Serbia). The material from Süngülü was allocated to Heosminthus minutus by de Bruijn et al. (2003), but Wang (2009) and Daxner Höck (2014) transferred this species to Allosminthus. Wessels et al. (2019) include the Süngülü material and specimens from Serbia in Heosminthus primiverus. Figure 9 shows Süngülü and Serbian specimens together with type material of H. primiverus from the late Eocene of southern China. Heosminthus and Allosminthus are Asian elements that migrated into Anatolia and the Balkans, but did not reach Western and Central Europe. Plesiosminthus is the first dipodid to arrive in Western and Central Europe during the late Oligocene (Wessels et al., 2019).
Infraorder: Hystricognathi Tullberg, 1899
Subfamily: Baluchimyinae Flynn, Jacobs and Cheema, 1986
A cladistical analysis of the cheek teeth of the Hystricognathi by Marivaux and Boivin (2019) showed that the Baluchimyinae are polyphyletic, but this was not followed by a formal classification. Van de Weerd et al. (2021a) pragmatically maintained the Baluchimyinae to house the Asian hystricognaths, viz. six genera described from the Oligocene of Pakistan and three genera from the latest Eocene and Oligocene of Anatolia. Here we add the late Eocene Dianomys Wang, 1984 from southern China to the list as given by van de Weerd et al. (2021a). Below we describe another genus found in Oligocene deposits of Anatolia that we include in the Baluchimyinae. The nomenclature of tooth parts is shown in Figs. 10 and 11.

Upper incisor, upper molar, DP3 and partial DP4 of Zorania milosi nov. gen. nov. sp. juxtaposed with cheek teeth of Daxneria fragilis (Gözükızıllı) and Lophibaluchia pilbeami Flynn and Cheema, 1994 (site Y-GSP417, Pakistan) as shown in van de Weerd et al. (2021a). The teeth are illustrated as dextral and with different enlargement factors. b Nomenclature for parts of M1-2 of Zorania milosi

Lower insicor, lower premolars and molars of Zorania milosi nov. gen. nov. sp. juxtaposed with cheek teeth of Daxneria fragilis (Gözükızıllı) and Lophibaluchia pilbeami Flynn and Cheema, 1994 (site Y-GSP417, Pakistan; as shown in van de Weerd et al. 2021a). The teeth are illustrated as from the right jaw and with different scales. b, e nomenclature for parts of m1-2 and dp4 of Zorania milosi
Genus Zorania nov. gen.
Derivation nominis: named after Zoran Marković (Natural History Museum in Belgrade) in honour of his successful hunt for Paleogene small mammals in Serbia.
Type species: Zorania milosi nov. sp.
Diagnosis: Large lophodont baluchimyine with high-crowned molars, thin enamel and without cement. Cusps and ridges are seamlessly merged into strong and high lophs. The anterior half of the upper molar is a forked ridge incorporating protocone, anteroloph and paracone. A well-developed and deep central flexus separates the anterior and posterior parts of the molar. The posterior half consists of two ridges that connect at the hypocone, forming a V-shape with the relatively small rounded metacone positioned in its labial opening. The protocone is pinched. The lower molar consist of two strong and obliquely orientated ridges, which are both forked at the lingual side. The protoconid and hypoconid are pinched.
Differential diagnosis: Zorania differs from all known hystricognath genera by its large size and strongly lophodont and high-crowned molars.
Zorania milosi nov. sp.
Derivatio nominis: named after Miloš Milivojević (Natural History Museum in Belgrade) in honour of his search for Paleogene small mammals in Serbia.
Synonymy: Ctenodactylidae gen. B. sp. 1 (Ünay et al. 2003).
Holotype: M1 or M2 dex (Y1-191, 2.43 x 1.92; Fig. 10b-c).
Type locality: Yeniköy, Anatolia Turkey (Figs. 1 and 2).
Age of the type locality: late early Oligocene.
Paratypes: one dex (?) DP3 (Y1-200, 0.71 x 0.75); one partial DP4 dex (Y1-192, 1.09 x 2.03); dp4 sin (Y1-193, 2.03 x 1.30); m1-2 sin (Y1-194, 2.35 x 1.88).
Incisor: prepared sample I inf (Fig. 12) and several not numbered partial incisors.

Sections of the lower incisor of Zorania milosi nov. gen. nov. sp. a-c transverse sections; b enlargement of square in a; c enlargement of square in b; d-f sagittal sections. e enlargement of square in d; f enlargement of square in e
Diagnosis and differential diagnosis: as for the genus.
Description
Upper incisor (Fig. 10a): The upper incisor has a fairly wide, but shallow sulcus.
DP3 (Fig. 10d): The rounded premolar has a fairly large wear-facet indicating its posterior side. The position of the facet is indicate by a line in Fig. 10d. There is a single high cusp in antero-lingual position and there are well-developed cingulum ridges at the posterior and lingual tooth border. A single voluminous root is present.
DP4 or P4 (Fig. 10d): This partial tooth has been identified as the posterior part of a DP4 or P4. It shows a well-developed isolated metacone. The hypocone is large and merged with the straight posteroloph. The hypocone has a narrow connection to the metaconule, which tapers into the mesolophule.
M1 or M2 (Figs. 10b, c): Only a single, barely used upper molar is available. Cusps and ridges are slightly higher labially than lingually. Hypoflexus and mesoflexus form together a central valley that is deep at its lingual and shallow at its labial side. Paraflexus and the flexus between metacone and mesolophule are shallow. The posteroloph connects to the metacone nearly closing the labial entrance to the posteroflexus. The posteroloph is the weakest of the four lophs.
Lower incisor (Figs. 11a, 12): The lower incisors have a wide but very shallow sulcus, which is not apparent in the cross-section (Fig. 12) nor on Fig. 11a. The microscopic structure of the enamel of the lower incisor (Fig. 12) is very similar to that of Daxneria fragilis from Gözükızıllı (van de Weerd et al. 2021a). The enamel thickness is approximately 210 μm (Fig. 12a). The portio externa (PE) is thin (~10% of the total), and consists of radial enamel; in the illustrated specimen it is damaged by short cracks (Fig. 12c). The portio interna (PI) is thick (~90%) and consists of transverse multiserial Hunter-Schreger bands (HSB). The bands contain three to four prisms (Fig. 12e, f). Thin plates of IPM are present in the bands; its crystallites are about orthogonal to the prisms (arrowed in Fig. 12f). The HSB make an angle of about 30°with the normal on the enamel-dentine junction (EDJ; Fig. 12e). In cross-section the PE shows a characteristic wavy pattern of the IPM crossing the bands and enclosing the prisms (Fig. 12b, c). A PLEX (Prisma-Less External layer) is absent.
dp4 (Figs. 11de, e): The postero-labial side of the elongated premolar is damaged. The tooth consists of two forked ridges separated by a deep central valley. Metaconid and protoconid are connected by the transverse antero-cingulid, bearing a minor anterior bulge reminiscent of an anteroconid. The protoconid is pinched.
m1 or m2 (Figs 11b, c): Protoconid and hypoconid are both pinched and incorporated into the two forked ridges that make up this molar. Anterior and posterior cingulids are present, the anterior one is a sharp low ridge, the posterior cingulid is a low bulge.
Discussion
Zorania milosi nov. gen. et sp. has been compared with Daxneria fragilis, recently described from slightly younger deposits in Gözükızıllı in Asia Minor (van de Weerd et al. 2021a). At first sight the molars of the two genera are rather different, but the similarities in bauplan of the cheek teeth are apparent after closer inspection (Figs. 10 and 11). The two genera have more or less the same size and both have high crowns. Lophodonty is much more pronounced in Z. milosi, which has its cusps and ridges organised in two V-shaped ridges. In contrast to D. fragilis, the upper molar of Z. milosi has no connection (mure) between the anterior and the posterior V-shaped ridge. In addition, in Z. milosi, the mesolophule, metaconule and anterior arm of the hypocone are seamlessly incorporated into a ridge, in which the original constituting elements cannot be recognised. The differences between D. fragilis and Z. milosi in the lower teeth are similar to the differences seen in the upper molar. The lower premolar and molar of Z. milosi have cusps and ridges fused in two not-connected V-shapes and there is no ectolophid. Compared to D. fragilis, the tip of the metalophid II has a more posterior position in Z. milosi enforcing the V-morphology of the anterior ridge. The incisor structure of the two genera is nearly identical. The dental morphology of Z. milosi seems more evolved and specialised than that of the stratigraphically slightly younger Daxneria fragilis; but in any case the dental morphology of the two genera are sufficiently close for assuming a common origin.
Although Lophibaluchia is much smaller, its upper molars resemble those of Zorania and Daxneria (Fig. 10). However, lower molars of Lophibaluchia have a well-developed ectolophid, a feature absent in Zorania (Fig. 11). We suggest that Lophibaluchia may belong to the same clade.
Baluchimyinae have so far exclusively been found in low-latitude Asia and in Anatolia. Two species of Dianomys have been described from several late Eocene sites in the Caiijchong Formation in southern China (Wang 2001); the sites are Ulangochuan in age (Chinese Land Mammal Age, 39.9-37.2 Ma; age after Wang et al. 2019). Baluchimys has been found in the Krabi basin in Thailand (Marivaux et al. 2000). A combination of magnetostratigraphy and biostratigraphy dates the fossiliferous fluvio-lacustrine Krabi section to be early Oligocene (34-31 Ma; Bennami et al. 2001). Six genera of baluchimyines are know from three sites in the low-latitude Indian subcontinent (Z108, Bugti 417, Paali nala C2); these sits have an age between 30 and 28 Ma (Métais et al. 2009). By contrast, baluchmyines are not know from the many rich late Eocene-Oligocene rodent assemblages from northern China, Mongolia or Kazakhstan. The location of the sites in which baluchmyines are present thus strongly suggest that the subfamily has a preference for warm humid climates: viz. present in low-latitude Asia, but absent in northern-central Asia, presumably because of its relatively dry climates. The baluchimynes migrated from south Asia into Anatolia, following routes located south of the Paratethys seas. A variety of genera occurs in five late Eocene-early Oligocene sites of Anatolia and the Thrace basin (reviewed in van de Weerd et al. 2021a), suggesting that the climate of these areas, which are wedged between the Neotethys ocean and the Paratethys seas, was humid and warm as well. The geographic distribution pattern of the Diatomyidae is very similar: these have been found in the Oligocene of low latitude (tropical) Asia, including Pakistan, India, Thailand and southern China and the Balkans where these occur in five early Oligocene sites in Serbia (Marković et al. 2017).
Family Erinaceidae Fischer, 1817
Erinaceidae gen. et sp. indet.
Material from Yeniköy: One damaged P3sin (Y1-201, 1.15 x 1.11) and one m2 dex (Y1-203, 2.43 x 1.92).
Description
P3 (Figure 13b): The premolar is incomplete, with a part of the postero-labial side broken off. The premolar shows a high paracone and a single lingual cusp representing the protocone. There is a small parastyle integrated in a low U-shaped ridge.

Erinaceidae gen. et sp. indet. from Yeniköy compared with galericines and tupaiodontines. a-b Erinaceidae gen. et sp. indet. from Yeniköy, the P3 is damaged; c, d, h Galerix saratji van den Hoek Ostende, 1992 from Harami and Kilçak (early Miocene, from van den Hoek Ostende 1992); e Tetracus nanus Montalban (Oligocene, from Hugueney and Adrover 2003); f T. nanus Itardies (Oligocene, from Crochet 1995) g cf. Neurogymnurus sp. Inkonak (late Oligocene, from de Bruijn et al. 1992); i Anatolechus neimongolensis (late Eocene from Wang 2008); j Zaraalestus minutus (early Oligocene, from Ziegler et al. 2007)
The m2 (Fig. 13a) has a relatively short trigonid; protoconid and metaconid are fairly large.
Discussion
Allocation of these two isolated teeth from Yeniköy poses a problem as erinacides are relatively scarce in most Oligocene assemblages and isolated teeth often lack diagnostic features that allow allocation to genus or subfamily. We have compared the two teeth with European and Anatolian galericines. Given the presence of Asian taxa in the Yeniköy assemblage, we also have included in the comparison the erinaceide subfamily of the Tupaiodontinae that occur in the Far East. Galerix Pomel, 1848 from the latest Oligocene of nearby Inkonak MR6 have been described as G. cf. sarajti by de Bruijn et al. (1992). Tetracus Aymard, 1850 is known from the early and late Oligocene of France and Spain with two species (Crochet 1974, 1995; Hugueney and Adrover 2003). De Bruijn et al. (1992) compared isolated teeth from Inkonak to Neurogymnurus Filhol, 1877, a genus of large size. This genus occurs in the Oligocene of Europe (Crochet 1974, 1995; Maridet et al. 2019) and a species has been described from the early Oligocene of Kazakhstan (Lopatin 1999). A small galericine is present too in early Oligocene assemblages of Serbia (Wessels in prep.). Illustrations of the corresponding dental elements of these three genera and the two teeth from Yeniköy are compared in Fig. 13. The P3 of Tetracus nanus from Montalban (Spain, Fig. 13e) and Itardies (France, Fig. 13f) is of similar size as that from Yeniköy, but these have a low protocone and insignificant parastyle, giving it a primitive appearance. The P3 of cf. Neurogymnurus from Inkonak (Fig. 13g) is much larger, and its poster-lingual extension is relatively shorter than that of G. saratji. The P3 from Yeniköy differs from that of Galerix sarjati (Fig. 14d) in its slender lingual flange and in being slightly smaller in general.

Palaeogeographic map of the late Eocene-early Oligocene, modified from Ozvart et al. 2017. The land areas that together form the Anatolia-Balkans province are in dark green. The locations of the basins and sites with the main Paleogene rodent assemblages are indicated in light green. Present day coast lines are in red, red arrows indicate displacement of Africa-Arabia. References for the sites with fossil rodents: Transylvanian basin (Baciu and Hartenberger 2001); Babušnica-Koritnica and Pčinje Basins (de Bruijn et al. 2017); Thrace basin (Ünay-Bayraktar 1989, Doukas and Theocharopoulos 1999), Çankiri-Çorum basin (van de Weerd et al. 2021a), Sivas basin (this paper), Lesser Caucasus Süngülü (de Bruijn et al. 2003), Benara (Russel and Zhai 1987)
Tupaiodontinae have been described from the late Eocene and Oligocene of Mongolia and Nei Mongol (northern China) by Ziegler et al. ( 2007) and Wang (2008). The P3 of Zaraalestus minutus figured by Ziegler et al. (2007) and Anatolechus by Wang (2008) are shown in Fig 13i-j. Both specimen show a wide lingual flange bearing protocone and minor hypocone, not different from those of Galerix, but different from the Yeniköy specimen that has a narrow flange bearing only a protocone (Fig. 13b). The m2 of these tupaiodontines are not very characteristic and not shown in Fig. 13. Late Eocene and Oligocene insectivores from southern China and southern Asia remain poorly known. As far as we know only Rich et al. (1983) described erinaceide material from site 80026 in the Caijiachong Formation in southern China. The age of this site, originally considered early Oligocene, is late Eocene. However, these few isolated teeth and partial teeth, do not allow subfamily allocation.
In conclusion, the illustrated galericini and tupaiodontine P3 and m2 seem rather similar, and none of the P3 seem close in morphology that of Yeniköy, which are thus best classified as Erinaceidae gen. et sp. indet.
Biogeography, geochronology and fauna composition
Detailed palaeogeographic maps including Asia Minor and the Balkans region have been published by Rögl (1999), Popov (2004), Meulenkamp and Sissingh (2003), Ozvart et al. (2016) and Palcu and Krijgsman (2021). All the late Eocene-early Oligocene maps show a compound land mass between the Paratethys seas and the Neotethys ocean, but the maps differ in important aspects. Figure 14, modified from Ozvart et al. (2016), illustrates an area of great complexity, with different tectonic terranes and sea straits. By contrast, the maps published by Palcu and Krijgsman (2021) depict a continuous and uninterrupted landmass including Iran, Anatolia and the Balkans. Licht et al. (2022) reviewed the geological history of the Balkans and Anatolia. They suggest that the land area between the Paratethys seas and the Neotethys ocean (which they named Balkanatolia) came into being as a more or less continuous terrane in the middle Eocene and was populated by immigrating Asian large mammals in the late Bartonian. We have plotted all known sites with Paleogene fossil small mammals, including the Sivas basin with Yeniköy (Fig. 14). Two murid molars have been found in the northern-most sites Treznea and Bociu (Transylvanian basin) in a thin non-marine intercalation in a Priabonean marine section (Baciu and Hartenberger 2001). This proves the presence of late Eocene murids in the Balkans, while murids are completely absent in the late Eocene of Western and Central Europe. Unfortunately, the murids were not illustrated and further findings at or near these sites have not been reported. The localities in the Babušnica-Koritnica and Pčinje Basins are described in de Bruijn et al. (2018) and those in the Thrace basin by Ünay-Bayraktar (1989) and Doukas and Theocharopoulos (1999). Localities with Paleogene small mammals in Anatolia are in the Çankiri-Çorum basin (van de Weerd et al. 2021a), the Sivas basin (this paper) and in the Lesser Caucasus (Süngülu, see de Bruijn et al. 2003; Benara, see references in van de Weerd 2021a).
The chronology of sites with assemblages of small mammals in Asia Minor (including Yeniköy) and the Balkans was recently reviewed by van de Weerd et al. (2021a, 2021b, both fig. 17). The magneto-stratigraphic age of Yeniköy is ~29 Ma, i.e. the later part of the early Oligocene and about contemporaneous with European mammal age MP 24. The occurrence of an evolved Eucricetodon species in Yeniköy is in line with the timing of the immigration of evolved Eucricetodon species in Europe (Freudenthal and Martín-Suárez 2016). However, the incisor micro-structure shows that Eucricetodon species from the Anatolia-Balkans region and Europe do belong to different clades. Given that glirids migrated into Anatolia in the "middle Oligocene" (Unay et al. 2003), their absence in Yeniköy is no surprise, although the assemblage is too small to consider their absence significant. Glirids are absent too in the early Oligocene of Serbia. Süngülü is considered to be of late Eocene-early Oligocene age (de Bruijn et al. 2003) and given an age range of ~33-35 Ma in the published chronostratigraphic schemes (van de Weerd et al. 2021a, 2021b). There is thus a considerable time gap of about 4-5 Ma between Yeniköy and Süngülü, representing most of the early Oligocene, for which there are no data in Anatolia. However, the rodent assemblages of Valnis, Raljin and Strelac (Babusnica-Koritnica basin) in southern Serbia, are considered to represent this time interval.
The small assemblage of Yeniköy is dominated by murids with 23 out of the 27 first and second rodent molars (85%; see Table 3). This dominance of murids is similar in all the Eocene-Oligocene rodent assemblages from the area: 75% in Süngülü (late Eocene, de Bruijn et al. 2003), 85 to 95 % in the five "middle" Oligocene assemblages from the Thrace basin (Ünay-Bayraktar 1989), 81 % in Gözükızıllı (late Oligocene, van de Weerd et al. 2021a) and 98% in Inkonak (late Oligocene, de Bruijn et al. 1992). The situation is not different in the eight late Eocene and early Oligocene sites in Serbia where murids dominate with 71 to 100%. By contrast, murids are rare in the Oligocene of low-latitude Asia. The nearest sites with fossil small mammals in SE direction are in Pakistan: C2 Paali nala (Marivaux et al. 1999, 2002; Marivaux and Welcomme 2003) and Y-GSP 417 (Flynn et al. 1986). Both are "middle" Oligocene and were located at low latitudes (~20°, Metais et al. 2009) in a different, presumably tropical, climatic zone. Quantitative details of the composition of these have not been published, but numbers may be estimated by counting measured specimens in the papers dealing with these sites. The large C2 Paali nala assemblage, with an estimated 655 first and second rodent molars, is dominated by Diatomyidae (73%) and Baluchimyidae (15%), whereas murids are a small minority with 4%. From the Y-GSP 417 an estimated 181 first and second molars have been recovered, of which 22% belong to Diatomyidae and 77% to Baluchimyidae and none to murids.
Striking is the occurrence of baluchimyids in the murid-dominated late Eocene-Oligocene assemblages of Süngülü, Yeniköy, Gözükızıllı, Thrace basin and Benara. So far baluchimyids have not been found yet in the early Oligocene of Serbia. Equally notable is the occurrence of Diatomyidae in the early Oligocene sites of the Serbian Babušnica-Koritnica basin. The Diatomyidae originated in low-latitude Asia and must have migrated through Anatolia to reach the Balkans. The dipodids Heosminthus and Allosminthus found in Serbia and Anatolia, are of northern-central Asian origin.
Conclusions
Three murids, one new baluchimyid, a dipodid and a galericine were identified in the small mammal assemblage of Yeniköy, a site in the Oligocene fluvio-lacustrine Selimye Formation of the Sivas basin. The age of the site has been established by magnetostratigraphy at ~ 29 Ma, that is in the later part of the early Oligocene and about contemporaneous with the European mammal zone MP24. The small assemblage is dominated by murids The study of the microstructure of the lower incisor reveals presence of several long-living lineages within the murid Eucricetodon. The Yeniköy assemblage contains Zorania milosi, a new genus and species of the Baluchimyidae, representatives of this family have now been found in five of the six known Oligocene assemblages of Anatolia. The early Oligocene Yeniköy assemblage and the late Oligocene Inkonak assemblage (de Bruijn et al. 1992) are both recovered from the Selimye Formation. It is expected that further prospecting in this formation will yield more assemblages that could represent the time gap of an estimated 4-5 million years between Yeniköy and Süngülü. The Yeniköy assemblage confirms presence of a late Eocene-Oligocene Anatolian-Balkans biogeographic province characterised by dominance of murids and presence of Baluchimyidae, Diatomyidae and Dipodidae.
Data Availability statement
Data sharing not applicable to this article as no datasets were generated or analysed during the current study. The material of Yeniköy is temporarily housed in the Department of Earth Sciences (Utrecht University).
References
Aymard, A. (1850). Compte-rendu de la séance du 13 avril 1849, réponse à M. Robert sur les mammifères fossiles des calcaires du Puy. Annales de la Société d’Agriculture, Sciences, Arts et Commerce du Puy, T.14, 80-86
Baciu, C., & Hartenberger, J.-L. (2001). Un exemple de corrélation marin–continental dans le Priabonien de Roumanie. Remarques sur la Grande Coupure. Comptes Rendus de l'Academie de Sciences, Sciences de la Terre et des Planètes, 333, 441–446. https://doi.org/10.1016/S1251-8050(01)01659-7
Bennami, M., Chaimanee, Y., Jaeger, J.-J., Suteethorn, V., & Ducroq, S. (2001). Eocene Krabi basin (southern Thailand): Paleontology and magnetostratigraphy. Geological Society America Bulletin, 113(2), 265-273. https://doi.org/10.1130/0016-7606(2001)113<0265:EKBSTP>2.0.CO;2
Bohlin, B. (1946). The fossil mammals from the Tertiary deposit of Taben-Buluk, western Kansu. Part 2: Simplicidentata, Carnivora, Artiodactyla and primates. Palaeontologia Sinica, new series C, 8B, 1–259.
Bruijn, H. de, Ünay, E., Hoek Ostende, L.W. van den, & Saraç, G. (1992). A new association of small mammals from the Lowermost Miocene of Central Anatolia. Geobios, 25, 651-670.
Bruijn, H. de, Ünay, E., Saraç, G., & Yïlmaz, A. (2003). A rodent assemblage from the Eo/Oligocene boundary interval near Süngülü, Lesser Caucasus, Turkey. In N. López-Martínez, P. Peláez-Campomanes, & M. Hernández Fernández (Eds.), En torno a Fósiles de Mamíferos: Datación, Evolución y Paleoambiente. Coloquios de Paleontología, Volumen Extraordinario no 1. En honor al dr. Remmert Daams, 47–76.
Bruijn, H. de, Marković, Z., Wessels, W., Milivojević, M., & Weerd, A. A. van de (2017). Rodent faunas from the Paleogene of south-east Serbia. Palaeobiodiversity and Palaeoenvironments. (2018) 98(3), 441-458. https://doi.org/10.1007/s12549-017-0305-0.
Bruijn H. de , Marković Z., Wessels W., & Weerd A. A. van de (2018). Pappocricetodontinae (Rodentia, Muroidea) from the Paleogene of south-east Serbia. Palaeobiodiversity and Palaeoenvironments. (2019) 99(3), 511–526. https://doi.org/10.1007/s12549-018-0343-2.
Cater, J. M. L., Hanna, S. S., Ries, A. C., & Turner, P. (1991). Tertiary evolution of the Sivas Basin, Central Turkey. Tectonophysics, 195, 29–46, https://doi.org/10.1016/0040-1951(91)90142-F
Crochet, J.-Y., (1974). Les insectivores des Phosphorites du Quercy. Palaeovertebrata 6 (1/2), 109-159.
Crochet, J.-Y., (1995). Le Gorouillas et las sites contemporains (Oligocene, MP 25) des phosphorites du Quercy (Lot, Tarn-et-Garonne, France) et leurs faunes de vertébrés. 4. Marsupiaux et insectivores. Palaeontographica (A) 236, 39-75.
Daxner-Höck, G. (1999). Family Zapodidae. In G. E. Rössner & K.Hessig (Eds.), The Miocene land mammals of Europe (pp. 337-342). München: Verlag Dr. Friedrich Pfeil.
Daxner-Höck, G. (2001). New zapodids from the Oligocene-Miocene deposits in Mongolia. Senckenbergiana lethaea, 81, 359–389.
Daxner-Höck, G., Badamgarav, D., & Maridet, O. (2014). Dipodidae (Rodentia, Mammalia) from the Oligocene and Early Miocene of Mongolia. Annalen des Naturhistorischen Museums in Wien, Serie A, 116, 131–214.
D’Elía, G., Fabre, P.-H., & Lessa, E.P. (2019). Rodent systematics in an age of discovery: recent advances and prospects. Journal Mammalogy. 100(3), 852–871. https://doi.org/10.1093/jmammal/gyy179.
Doukas, C. S., & Theocharopoulos, C. D. (1999). Smaller mammals from the Oligocene of Kyprinos (Thrace, N. Greece). In J. W. F. Reumer, & J. de Vos (Eds.), Elephants have a snorkel! Papers in honour of Paul Y. Sondaar. Deinsea, 7, 133–145.
Engesser, B. (1987). New Eomyidae, Dipodidae and Cricetidae (Rodentia, Mammalia) of the Lower Freshwater Molasse of Switzerland and Savoy. Eclogae Geologicae Helvetiae, 80(3), 945–993.
Filhol, H. (1877). Recherches sur les Phosphorites du Quercy: Étude des fossiles qu’on y rencontre et spécialement des Mammifères (pp. 1-561). Paris: G. Masson.
Fischer, G. (1817). Adversaria Zoologica. Mémoires de la Société Impériale des Naturalistes de Moscou, 5, 357–446.
Flynn L. J., & Cheema I. U. (1994). Baluchimyine rodents from the Zinda Pir Dome, western Pakistan: systematic and biochronologic implications. In Y. Tomida, C. Li, & T. Setoguchi (Eds.), Rodent and Lagomorph Families of Asian Origins and Diversification. National Science Museum Monographs. Kyoto, 115-129.
Flynn, L. J., Jacobs, L. L., & Cheema, I. U. (1986). Baluchimyinae, a new ctenodactyloid rodent subfamily from the Miocene of Baluchistan. American Museum Novitates, 2841, 1–58.
Freudenthal, M. (1988). Redescription of Eucricetodon atavus (Misonne, 1957) (Rodentia) from Hoogbutsel (Belgium). Bulletin van het Koninklijk Belgisch Instituut voor Natuurwetenschappen, Aardwetenschappen, 58, 199-207.
Freudenthal, M. (1996). The Early Oligocene rodent fauna from Olalla 4A (Teruel, Spain). Scripta Geologica. 112,1–67.
Freudenthal, M., & Martín-Suárez E. (2016). A review of Oligocene and early Miocene European Cricetidae (Mammalia). Spanish Journal of Palaeontology, 31 (2), 341-352.
Gomes Rodrigues, H., Marivaux, L., & Vianey-Liaud, M. (2012). The Cricetidae (Rodentia, Mammalia) from Ulantatal area (Inner Mongolia, China): new data concerning the evolution of Asian cricetids during the Oligocene. Journal of Asian Earth Sciences, 56, 160–179. https://doi.org/10.1016/j.jseaes.2012.05.007.
Gündogan, I., Onal, M., & Depçi, T. (2005). Sedimentology, petrography and diagenesis of Eocene-Oligocene evaporites: the Tuzhisar formation, SW Sivas Basin, Turkey. Journal of Asian Earth Sciences. 25, 791-803. https://doi.org/10.1016/j.jseaes.2004.08.002
Hoek Ostende, L. W. van den (1992). Insectivore faunas from the lower Miocene of Anatolia. Part 1: Erinaceidae. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 94(4), 437–467.
Hooker, J. J., Collinson, M., and Sille, N. P. (2004). Eocene–Oligocene mammalian faunal turnover in the Hampshire Basin, UK: calibration to the global time scale and the major cooling event. Journal of the Geological Society, London, 161, 161–172. http://jgs.lyellcollection.org.
Huang, X. (1992). Zapodidae (Rodentia, Mammalia) from the middle Oligocene of Ulantatal, Nei Mongol. Vertebrata PalAsiatica, 30(4), 249–286.
Hugueney, M. & Adrover, R. (2003). Tetracus daamsi, une nouvelle espèce de Galericinae (Erinaceidae, Mammalia) dans l’Oligocène de Majorque (Espagne). Coloquios de Paleontología, Vol. Volumen Extraordinario no 1. En honor al dr. Remmert Daams. 311-324.
Illiger, C. (1811). Prodromus systematis mammalium et avium additis terminis zoographicis utriusque classis, eorumque versione germanica (pp. 1–301). Berolini: Sumptibus C. Salfeld.
Kalthoff, D. C. (2000). Die Schmelzmikrostructur in den Incisiven der hamsterartigen Nagetiere und anderer Myomorpha (Rodentia, Mammalia). Palaeontographica (A), 259, 1–193.
Kergaravat, C., Ribes, C., Legeay, E., Callot, J. P., Kavak, K. S., & Ringenbach, J.-C. (2016). Minibasins and salt canopy in foreland fold-and-thrust belts: the central Sivas Basin, Turkey. Tectonics, 35(6), 1342–1366.
Kergaravat, C., Ribes, C., Callot, J.-P., & Ringenbach, J.-C. (2017). Tectono-stratigraphic evolution of salt-controlled minibasins in a fold and thrust belt, the Oligo-Miocene central Sivas Basin. Journal of Structural Geology, 102: 75-97.
Krijgsman W., Duermeijer, C. E., Langereis, C. G., Bruijn H. de, Saraç G., & Andriessen, P. A. M. (1996). Magnetic polarity stratigraphy of Late Oligocene to Middle Miocene mammal-bearing continental deposits in Central Anatolia (Turkey). Newsletters on Stratigraphy, 34(1), 13-29.
Legeay, E., Pichat, A., Kergaravat, C., Ribes, C., Callot, J.-P., Ringenbac, J.-C., Bonnel, C. Guilhem, H., Poisson, A., Mohn, G., Crumeyrolle, P., Sevki Kavak, K. & Temiz, H. (2019). Geology of the Central Sivas Basin (Turkey). Journal of Maps, 15 (2), 406–417. https://doi.org/10.1080/17445647.2018.1514539
Li, Q., Meng, J., & Wang, Y. (2016). New Cricetid Rodents from Strata near the Eocene-Oligocene Boundary in Erden Obo Section (Nei Mongol, China). PLoS ONE 11(5), e0156233. https://doi.org/10.1371/journal.pone.0156233
Licht, A., Métais, G., Coster, P., Ibilioğlu, D., Ocakoğlu, F., Westerweel, J., Mueller, M., Campbell, C., Mattingly, S., Wood, M. C., and Beard, K. C., (2022). Balkanatolia: The insular mammalian biogeographic province that partly paved the way to the Grande Coupure. Earth-Science Reviews 226 (2022) 103929. https://doi.org/10.1016/j.earscirev.2022.103929
Lopatin, A. V. (1999). Oligocene and Early Miocene Insectivores (Mammalia) from Mongolia. Paleontological Journal, 33 (2), 182-183.
Maridet, O. Hugueney, M., & Costeur, L. (2019). Aubenas-les-Alpes (S-E France). Part III – Last and final part of the mammalian assemblage with some comments on the palaeoenvironment and palaeobiogeography. Annales de Paléontologie. 105(2), 139–153. https://doi.org/10.1016/j.annpal.2019.03.001
Marivaux, L., & Boivin, M. (2019). Emergence of hystricognathous rodents: Palaeogene fossil record, phylogeny, dental evolution and historical biogeography. Zoological Journal of the Linnean Society. 20, 1-36. https://doi.org/10.1093/zoolinnean/zlz048
Marivaux, L., & Welcomme, J.-L. (2003). New diatomyid and baluchimyine rodents from the Oligocene of Pakistan (Bugti Hills, Baluchistan): systematic and paleobiogeographic implications. Journal of Vertebrate Paleontology. 23, 420–434. https://doi.org/10.1671/0272-4634(2003)023[0420:NDABRF]2.0.CO;2
Marivaux, L., Vianey-Liaud, M., & Welcomme, J. L. (1999). Premiere découverte de Cricetidae (Rodentia, Mammalia) Oligocènes dans le synclinal sud de Gandoe (Bugti Hills, Balouchistan, Pakistan). Comptes Rendus de l’Académie des Sciences, 329, 839–844. https://doi.org/10.1016/S1251-8050(00)88640-1
Marivaux, L., Benammi, M., Ducrocq, S., Jaeger, J.-J., & Chaimanee Y. (2000). A new baluchimyine rodent from the Late Eocene of the Krabi Basin (Thailand): paleobiogeographic and biochronologic implications. Comptes rendus de l'Academie des Sciences. 331, 427-433.
Marivaux, L. Vianey-Liaud, M. Welcomme, J-L. & Jaeger, J.-J. (2002), The role of Asia in the origin and diversification of hystricognathous rodents. Zoologica Scripta, 31(3), 225–239.
Marković, Z., Wessels, W., Weerd, A. A. van de, & Bruijn, H. de (2017). On a new Diatomyid (Rodentia, Mammalia) from the Paleogene of S. E. Serbia, the first record of the family in Europe. Palaeobiodiversity and Palaeoenvironments. (2018) 98(3), 459–469. https://doi.org/10.1007/s12549-017-0301-4.
Marković, Z., Wessels, W., Weerd, A. A. van de, & Bruijn, H. de (2019). Pseudocricetodontinae (Mammalia, Rodentia) from the Paleogene of south-east Serbia. Palaeobiodiversity and Palaeoenvironments. (2020) 100(1), 251–267. https://doi.org/10.1007/s12549-019-00373-8.
Mein, P., & Freudenthal, M. (1971). Une nouvelle classification des Cricetidae (Mammalia, Rodentia) du Tertiaire de l’Europe. Scripta Geologica. 2, 1–35.
Métais, G., Antoine, P.-O., Hassan Baqri, S. R., Crochet, J.-Y., De Franceschi, D., Marivaux, L., & Welcomme, J.-L. (2009). Lithofacies, depositional environments, regional biostratigraphy and age of the Chitarwata Formation in the Bugti Hills, Balochistan, Pakistan. Journal of Asian Earth Sciences, 34, 154–167. https://doi.org/10.1016/j.jseaes.2008.04.006
Meulenkamp, J. E., & Sissingh W. (2003). Tertiary palaeogeography and tectonostratigraphic evolution of the Northern and Southern Peri-Tethys platforms and the intermediate domains of the African–Eurasian convergent plate boundary zone. Palaeogeography, Palaeoclimatolology, Palaeoecolology. 196, 209–228. https://doi.org/10.1016/S0031-0182(03)00319-5
Ozsvárt, P., Kocsis, L., Nyerges, A., Győri, O., & Pálfy, J. (2016). The Eocene-Oligocene climate transition in the Central Paratethys. Palaeogeography, Palaeoclimatology, Palaeoecology, 459, 471–487, https://doi.org/10.1016/j.palaeo.2016.07.034.
Palcu, D. V., & Krijgsman, W. (2021). The dire straits of Paratethys: Gateways to the anoxic giant of Eurasia. Geological Society, London, Special Publications 523, https://doi.org/10.1144/SP523-2021-73.
Pichat, A., Hoareau, G., Callot, J.-P., & Ringenbach, J.-C. (2016). Diagenesis of Oligocene continental sandstones in salt-walled mini-basins Sivas Basin, Turkey. Sedimentary Geology, 339, 13-31. https://doi.org/10.1016/j.sedgeo.2016.03.025.
Poisson, A., Guezou, J., Ozturk, A., Inan, S., Temiz, H., Gürsöy, H., Kavak, K., & Özden, S. (1996). Tectonic setting and evolution of the Sivas Basin, Central Anatolia, Turkey. International Geology Review, 38, 838-853. https://doi.org/10.1080/00206819709465366
Poisson, A., Vrielynck, B., Wernli, R., Negri, A., Bassetti, M.-A., Büyükmeriç, Y., Ozer, S., Guillou, H., Kavak, K. S., & Temiz, H. (2016). Miocene transgression in the central and eastern parts of the Sivas Basin (Central Anatolia, Turkey) and the Cenozoic palaeogeographical evolution. International Journal Earth Sciences, 105, 339-368. https://doi.org/10.1007/s00531-015-1248-1.
Pomel, N. A. (1848). Études sur les carnassiers insectivores. I. Insectivores fossils. II. Classification des insectivores. Archives des sciences physiques et naturelles. 9, 159–165, 244–251.
Popov, S. V., Rögl, F., Rozanov, A. Y., Steiniger, F. F., Shcherba, I. G., & Kovac, M. (Eds.). (2004). Lithological-Paleogeographic maps of Paratethys, 10 Maps, Late Eocene to Pliocene. Courier Forschungs-institut Senckenberg, 250, 1–46.
Ribes, C., Kergaravat, C., Bonnel, C., Crumeyrolle, P., Callot, J.-P., Poisson, A., Temiz, H. & Ringenbach, J.-C., (2015). Fluvial sedimentation in a salt-controlled mini-basin: stratal patterns and facies assemblages, Sivas Basin, Turkey. Sedimentology, 62, 1513-1545. https://doi.org/10.1111/sed.12195.
Rich, T. H. V., Zhang, Y.-P. & Hand, S. J., (1983). Insectivores and a bat from the early Oligocene Caijiachong Formation of Yunnan, China. Australian Mammalogy, 6, 61-75.
Rögl, F. (1999). Mediterranean and Paratethys. Facts and hypotheses of an Oligocene to Miocene Paleogeography (short overview). Geologica Carpathatica, 50 (4). 339-349
Russel, D. E. & Zhai, R., (1987). The Paleogene of Asia: mammals and stratigraphy. Mémoires du Muséum National d’Histoire Naturelle, serie C, 52, 1–488.
Schaub, S. (1925). Die hamsterartigen Nagetiere des Tertiärs und ihre lebenden Verwandten. Abhandlungen Schweizerischen Paläontologische Gesellschaft. 45, 1–114.
Schaub, S. (1958). Simplicidentata (Rodentia). Traité de paléontologie, 6(2), 659–818.
Sümengen, M., Ünay, E., Saraç, G., Bruijn, H. de, Terlemez, I., & Gürbüz, M. (1990). New Neogene rodent assemblages from Anatolia (Turkey). In E. H. Lindsay, V. Fahlbusch, and P. Mein (Eds.), European Neogene mammal chronology (pp. 61-72). New York: Plenum Press, Nato serie A 180.
Thaler, L. (1966). Les rongeurs fossiles du Bas-Languedoc dans leur rapports avec l’histoire des faunes et la stratigraphie d’Europe. Mémoires Musée Histoire Naturelles., nouvelle série C. 17, 1–295.
Thaler, L. (1969). Rongeurs nouveaux de l'Oligocène moyen d'Espagne. Palaeovertebrata, 2, 191–207.
Tong, Y. (1997). Middle Eocene small mammals from Liguanqiao basinof Henan province and Yuanqu basin of Shanxi province, Central China. Palaeontologica Sinica, 18, New Series C(26), 1–256.
Tullberg, T. (1899). Über das System der Nagetiere: eine phylogenetische Studie. Nova Acta Regiae Societatis Scientiarum Upsaliensis. Upsala, 3(18), 1-514.
Ünay-Bayraktar, E. (1989). Rodents from the Middle Oligocene of Turkish Thrace. Utrecht Micropaleontological Bulletins Special Publication, 5, 1–119.
Ünay, E., & Bruijn, H. de, (1987). Middle Oligocene to early Miocene Rodent assemblages from Turkey, a preliminary report. Münchner Geowissenshaftlicher Abhandlungen (A) 10, 303-210.
Ünay, E., Bruijn, H. de, & Saraç, G. (2003). The Oligocene rodent record of Anatolia: a review. In W. F. Reumer, & W. Wessels (Eds.), Distribution and migration of Tertiary mammals in Eurasia. Deinsea, Annual of the Natural History Museum, Rotterdam, 531–537.
Vandenberghe, N., Hilgen, F.J., & Speijer, R. P. (2012). The Paleogene Period. In F. M. Gradstein, J. G. Ogg, M. Schmitz, & G. Ogg (Eds.), The Geologic Time Scale 2012 (pp. 855-921). Elsevier Publication.
Wang, B-Y. (1984). Dianomys gen. nov. (Rodentia, Mammalia) from the lower Oligocene of Qujing, Yunnan, China. Mainzer geowissen-schaftliche Mitteilungen, 13, 37-48.
Wang, B.-Y. (1985). Zapodidae (Rodentia, Mammalia) from the lower Oligocene of Qujing, Yunnan, China. Mainzer geowissenschaftliche Mitteilungen, 14, 345–367.
Wang, B.-Y. (2001). Late Eocene Ctenodactyloids (Rodentia, Mammalia) from Quijing, Yunnan, China. Vertebrata PalAsiatica, 39, 24-42. http://www.vertpala.ac.cn/EN/Y2001/V39/I01/24
Wang, B.-Y. (2008). First record of late Eocene Insectivores and Chiropteres from Nei Mongol, China. Vertebrata PalAsiatica, 46(4), 249-264. [In Chinese with English summary] 1. http://www.vertpala.ac.cn/EN/Y2008/V46/I4/249
Wang, B.-Y. (2009). On Tatalsminthus (Dipodidae, Rodentia). Vertebrata PalAsiatica, 47/1: 81–84. [in Chinese with English summary]. http://www.vertpala.ac.cn/EN/Y2009/V47/I1/81
Wang, B. & Meng, J. (1986). Eucricetodon (Rodentia, Mammalia) from the lower Oligocene of Qujing, Yunnan, China. Vertebrata PalAsiatica, 24(2), 110-120. [in Chinese with English summary]
Wang, Y.-Q., Li. Q., Bai, B., Jin, X., Mao, F. Y., & Meng, J. (2019). Paleogene integrative stratigraphy and timescale of China. Science China Earth Sciences, 62. https://doi.org/10.1007/s11430-018-9305-y.
Weerd, A. A. van de, Bruijn, H. de, & Wessels, W. (2021a). New rodents from the late Oligocene site of Gözükızıllı in Anatolia (Turkey). Historical Biology, https://doi.org/10.1080/08912963.2020.1800682.
Weerd, A. A. van de, Bruijn, H. de, Wessels, W., & Marković, Z. (2021b). New late Oligocene rodent faunas from the Pannonian basin. Palaeobiodiversity and Palaeoenvironments (2022) 102(2), 465-492. https://doi.org/10.1007/s12549-021-00487
Wessels, W., Weerd, A. A. van de, Bruijn, H. de, & Marković, Z. (2019). Dipodidae (Mammalia, Rodentia) from the Paleogene of south-east Serbia. Palaeobiodiversity and Palaeoenvironments, (2020), 100(3), 841–848. https://doi.org/10.1007/s12549-019-00392-5.
Ziegler, R., Dahlmann, T., & Storch, G. (2007). Oligocene-Miocene Vertebrates from the Valley of Lakes (Central Mongolia): Morphology, phylogenetic and stratigraphic implications. Annales Naturhistoriches Museum. Wien, 108A, 53-164.
Acknowledgements
We gratefully acknowledge the facilities offered by the MTA (Turkish Geological Survey) during the field parties many years ago, in particular Gerçek Saraç (now retired) is thanked for his support. The SEM pictures of the cheek teeth were made by Tilly Bouten (Utrecht University) and retouched by Jaap Luteijn (formerly at Utrecht University). The map of Figure 1 was prepared by Margot Stoete (Utrecht University); Leonard Bik (Utrecht University, Earth Sciences Geolab) assisted in the preparation of the incisor samples. The SEM photographs of incisor microstructure were made by Hans Meeldijk (Utrecht University, Electron Microscopy Department). A discussion with Daniela Kalthoff (Swedish Museum of Natural History, Stockholm) sharpened our interpretation of Eucricetodon incisor structure. Jan van Dam (Utrecht University) critically read the typescript. We thank Gudrun Höck and Lars van den Hoek Ostende (Naturalis, Leiden) for their careful and detailed reviews.
Funding
The field work has been supported financially by the Department of Earth Sciences (Utrecht University).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
†Hans de Bruijn passed away 11 September 2021
This article is registered in Zoobank under: http://zoobank.org/urn:lsid:zoobank.org:pub:37F90FE0-7270-4A94-AFCC-E253420D68DC
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
van de Weerd, A.A., de Bruijn, H. & Wessels, W. A small assemblage of early Oligocene rodents and insectivores from the Sivas basin, Turkey. Palaeobio Palaeoenv 103, 609–632 (2023). https://doi.org/10.1007/s12549-022-00563-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-022-00563-x