
Abstract
A new periotic (MuMAB 240508) is described from the Tortonian of the Stirone River, Northern Italy. The new specimen is described and compared to all the known periotics of extant and extinct balaenopterid species. The new specimen shows balaenopterid characters, including the triangular anterior process, the transverse and anteroposterior elongations of the pars cochlearis, as well as in the arrangement of the endocranial foramina of the periotic. The presence of a medial promontorial groove, a lack of separation between the round window and the perilymphatic foramen together with the presence of a protruding set of medial crests suggest that this periotic may belong to an archaic and undescribed balaenopterid species. A phylogenetic analysis was performed to understand the relationships of this specimen within the broader context of balaenopterid phylogeny; this analysis showed that MuMAB 240508 belongs to a basal balaenopterid taxon that is the sister group to all the other Balaenopteridae with the exception of ‘Balaenoptera’ ryani. An ordinary least squares regression analysis showed that a relationship exists linking the length of the anterior process of the periotic and the total body length in Balaenopteridae. By means of such a relationship, the total body length of the individual to which MuMAB 240508 belonged to was reconstructed and resulted in c. 14.4 m. This individual was longer than all the other contemporaneous balaenopterid mysticetes confiming the hypothesis that the origin of large size occurred in this family earlier than previously thought.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In 1985 Avio Martini and the late Raffaele Quarantelli discovered the holotype skeleton of Plesiobalaenoptera quarantellii Bisconti, 2010 in the Tortonian outcrop of the Stirone River near to the La Bocca locality (Fig. 1). This associated skeleton included a well preserved rostrum, two mandibular rami, two periotics, two tympanic bullae and a series of vertebrae and ribs showing various degrees of articulation (Fig. 2). In the original description of this specimen, Bisconti (2010) published a diagram showing the arrangement of the bones, including an additional element labeled as ‘alien periotic’. No additional words were spent about this bone.

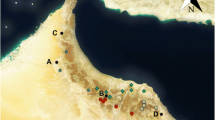
Locality of the discovery of MuMAB 240508. a Italy with the discovery locality indicated by an arrow and the whale icon in the Emilia Romagna region (in green); b Salsomaggiore Terme area with the discovery locality indicated by an arrow and the whale icon

The holotype skeleton of Plesiobalaenoptera quarantellii (MuMAB 240505) in dorsal view with the position of the ‘alien periotic’ (MuMAB 240508) described in the present paper. Excerpts from the photogrammetry of the specimen. Scale bar equals 50 cm
The discovery of this allochtonous periotic among the skeletal remains of Plesiobalaenoptera quarantellii took place during the study of the specimen by one of us (MB). The periotic was located posteriorly to the rostrum on the left side of the skeleton (Fig. 2); it represented the third periotic found among the bones of P. quarantellii and differed from those described by Bisconti (2010) in a number of characters. The two periotics assigned to P. quarantellii were found in close association to the poorly preserved skull remains and they clearly belonged to the holotype skeleton due to their relative position and morphological homogeneity.
It is difficult to explain how the allochtonous periotic reached its final position but it must be said that in the Tortonian outcrop of the Stirone River, fossils are abundant and include whales, shark teeth, corals, mollusks, brachiopods and crustaceans (Marasti 1973). It is therefore reasonable to hypothesise that this periotic was detached from another specimen and moved until it was buried together with the skeleton of P. quarantellii. The fact that in the allochtonous periotic the posterior process and the tympanic bulla are absent and the attachment sites for these two structures are rounded suggests that this specimen was subject to a prolonged transport on the bottom from its original depositional site.
The allochtonous periotic received the accession number MuMAB 240508 at the Museo Mare Antico e Biodiversità in Salsomaggiore Terme and is described and compared herein to the known species of balaenopterid whales. The phylogenetic relationships of this specimen were investigated as far as the original body size of the individual in order to understand the biotic implications of this find.
Even though MuMAB 240508 is different from the periotics of all the other currently known balaenopterid species (see Comparisons section), we prefer to postpone the establishment of a new taxon until a better preserved and more complete specimen will be available for study.
Geological setting
The Northwestern Apennine is a fold-and-thrust belt composed by two distinct structural levels, the upper of which, named Ligurian unit, is formed by "Mesozoic oceanic and forearc deposits derived from the closure of the Ligure-Piemontese ocean and its continental margin" (Artoni et al. 2004).
On the top of the Ligurian unit lies the Epiligurian succession, which is primarily constituted by "tectonically controlled turbidite and slope deposits" which settled between the middle Eocene and the early Messinian in a piggy-back, wedge-top basin (Artoni et al. 2004).
The Tortonian-Messinian sediments, encompassed in the Epiligurian succession, outcrop along the banks of the Stirone river, near La Bocca locality and are attributed to the “Termina Formation” (upper Serravallian-lower Messinian) represented here by the “Member of Vigoleno” and the “Member of Scipione”, respectively related to the shelf environments (Member of Vigoleno) and to the transition between the internal and the external shelf (Member of Scipione).
The Upper Miocene outcrop of the Stirone river is strongly faulted and highly fossiliferous and bioturbated on a local scale; it is characterised by glauconitic-silty sands interposed within fossiliferous sandy silts. A biocalcarenitic level formed by brachiopods, bivalves, gastropods and corals is recognised in the lower part. The macrofauna is more abundant above the calcarenitic level and in correspondence of major concentration of glauconite. The Messinian ipohaline deposits occur at the top of this succession.
Molluscs represent the dominant component of the benthic fauna; however, corals, brachiopods and crustaceans are present. Pectinidae and Terebratulidae show the best preservation while fossil specimens assigned to different families show highly different degrees of preservation and, sometimes, they are found as internal molds.
The unavailability of data on facies and macrobenthonic associations do not allow a careful palaeoenvironmental reconstruction. Moreover, the qualitative studies on molluscs (Marasti 1973) show that the most common species are Ancillaria glandiformis (Lamarck), Thracia convexa (Wood), Clavilithes klipsteini (Michelotti), Clavatula semimarginata (Lamarck), Pecten revolutus Michelotti, P. fraterculus (Sowerby), Amusium cristatum (Bronn), beside the presence of solitary corals and brachiopods (Marasti 1973). These data suggest a reference to distal circalittoral environment characterised by low energy in accordance with the Plancton/Benthos ratio that indicates an average depth between 80 and 100 m (Zermani 2001).
Based on biostratigraphic data, this succession has been assigned to the Tortonian (Zermani 2001; Monegatti and Raffi 2010; Lin et al. 2017) even though there are discrepancies on the precise age of these deposits. Zermani (2001), based on planktonic foraminifera, has attributed the succession to MMi9 and MMi10 biozones (lower Tortonian) calibrated on the emended Standard Mediterranean planktonic foraminiferal biozonation (Lirer et al. 2019) but the examined specimens have been probably reworked (Zermani 2001). Recently, Lin et al. (2017) has assigned the Miocene succession to the N17 foraminiferal biozone (upper Tortonian/lower Messinian) and probably to the upper Tortonian NN10 calcareous nannofossil zone of Martini (1971).
The diversity of the malacofauna of the Stirone River is typical of the upper Tortonian in terms of both biogeographic and stratigraphic distributions (Monegatti and Raffi 2010). In fact, 14.3% of the species belongs to the Miocene Atlantic–Proto-Mediterranean species (MAM group of Monegatti and Raffi 2010) which became definitively extinct in the Atlantic realm during the Messinian. 12.5% of the species belongs to endemic Proto-Mediterranean taxa: 5.4% of the species are already cited since Early and/or Middle Miocene (i.e. Pecten vigolenensis Simonelli and Clavatula ditissima (Mayer) and 7.1% of the species are neoendemics taxa of the Tortonian-Messinian (as Tectonatica dertomammilla (Sacco) and Semicassis miolaevigata (Sacco)). Proto-Mediterranean Messinian species (MEM species) are absent in the assemblage while present in molluscan assemblages collected from other Messinian localities (Monegatti and Raffi, 2010). Taken together, these data confirm that the Stirone succession can be referred to the late Tortonian (c. 9.0-7.2 Ma).
Materials and methods
Institutional abbreviations
AMNH, American Museum of Natural History, New York, USA; CASG, California Academy of Science Geology, San Francisco, USA; IRSNB, Institut Royal des Sciences Naturelles de Belgique, Bruxelles, Belgium; IZIKO, Iziko South African Museum, Cape Town, South Africa; MSNM, Museo di Storia Naturale del Mediterraneo, Livorno, Italy; MSNUP, Museo di Storia Naturale dell’Università di Pisa, Calci, Italy; MuMAB, Museo Mare Antico e Biodiversità, Salsomaggiore Terme, Italy; MUSE, Museo delle Scienze, Trento, Italy; SDNHM, San Diego Natural History Museum, San Diego, USA; USNM, United States National Museum, Smithsonian Institution, Washington, USA.
Studied material
Left periotic MuMAB 240508 in MuMAB collection. The specimen was found together with the holotype skeleton of Plesiobalaenoptera quarantellii (MuMAB 240505) by Raffaele Quarantelli and Avio Martini. Data about the discovery were provided by the late Raffaele Quarantelli by two written documents; one of these documents is dated 10 September 1987 and includes a detailed map of the locality with the precise indication of the discovery site. The discovery of MuMAB 240505 was done in the Autumn 1985 and the excavation ended in August 1987. The geographic co-ordinates of the discovery site are 44°50’23.1” N, 2°29’43.6” W; the locality is known as La Bocca, 3.7 km northwest of Salsomaggiore Terme (Fig. 1). The sediment at the outcrop consisted in massive and not stratified, clayey-sandy marls. The specimen immerged in the sediment in SW direction with an inclination of c. 25°.
Anatomy and measurements
Anatomical terminology follows Mead and Fordyce (2009) and Ekdale et al. (2011). Measurements were taken with digital calipers Sourcingmap (300 mm) and Tacklife D02 (150 mm) both with error margin to the nearest 0.01 mm. A few additional measurements were taken by the digital instruments available in Adobe Photoshop 2021.
Photography
The periotic MuMAB 240508 and the holotype of Plesio-balaenoptera quarantellii were photographed by a full-frame, mirrorless Nikon Z7 camera with a Nikkor Z 24-70 mm f/4 lens. Light was provided by a Nikon SB700 speedlight flash. The photogrammetry of Plesiobalaenoptera quarantellii was assembled from 201 pictures by Agisoft Photoscan (www.agisoft.com) software.
Phylogenetic analysis
A phylogenetic analysis was performed to understand the phylogenetic relationships of MuMAB 240508 within Mysticeti. The morphological characters of MuMAB 240508 were coded and added to the dataset published by Bisconti et al. (2021a) that is a slightly modified version of the datasets published by Bisconti and Bosselaers (2020) and Bisconti et al. (2019, 2020, 2021b). After the addition of MuMAB 240508, this dataset includes 355 characters scored for 89 taxa. The search for the most parsimonious cladograms was performed by TNT (Goloboff and Catalano 2016) through 2000 runs of the tree-bisection-reconnection (hereinafter: TBR) algorithm retaining 10 trees per replication. Supporting analyses included: (a) 1000 bootstrap replicates (bootstrap support value, hereinafter: BSV); (b) symmetric resampling with 1000 replicates and 33 change probability (absolute frequencies); (c) Bremer support (collapsing trees with support below 50) using TBR from suboptimal trees. Tree visualization and colour addition were performed by MESQUITE 3.61 (Maddison and Maddison 2021) and Adobe Photoshop 2021. The calculus of the Consistency Index (CI) and of the Retention Index (RI) was performed by a dedicated subroutine in TNT; the calculus of the Homoplasy Index (HI) was perfomed using the following formula: HI=1-CI.
Reconstruction of body size
We used the morphometric data of MuMAB 240508 to infer the total body length of the individual that it belonged to. In particular, we assessed the relationship between the length of the anterior process of the periotic (in mm) and the total length of the individual (in m) by an ordinary least square regression analysis performed by PAST (Hammer et al. 2001). We used the dataset provided in Table 1 including specimens assigned to extant Balaenoptera and Megaptera species. We calculated the regression equation by PAST and used the value of the R2 to assess the reliability of the inference. We then compared our body size inference to the body sizes of the balaenopterid species (Table 2) that were existent in the stratigraphic interval corresponding to the age of MuMAB 240508 to understand the implications of our result.
Systematic Palaeontology
Class Mammalia Linnaeus, 1758
Order Cetacea Brisson, 1762
Suborder Mysticeti Flower, 1864
Infraorder Chaeomysticeti Mitchell, 1989
Parvorder Balaenomorpha Geisler and Sanders, 2003
Superfamily Thalassotherii Bisconti, Lambert and Bosselaers, 2013
Epifamily Balaenopteroidea Gray, 1868
Family Balaenopteridae Gray, 1864
gen. et sp. indet.
Description
The periotic MuMAB 240508 is relatively large measuring 154.4 mm in anteroposterior length (Figs. 3, 4 and 5). The posterior process is absent as it is broken at its base. The anterior process is long (Table 3) and triangular in outline. In dorsal view, both the medial and lateral borders of the anterior process are concave. The anterior apex of the anterior process is pointed in dorsal view. Ventrally, the anterior process widens posteriorly and shows a small and rounded ventrolateral tuberosity. The anterior pedicle of the tympanic bulla is located along the posterior portion of the lateroventral border of the anterior process; this structure is blade-like, with a length of 31.8 mm and a width of about 5 mm. Ventrally, the anterior process is slightly convex and is posteriorly delimited by an anteriorly convex groove for the tensor tympani muscle.

The periotic MuMAB 240508. a ventral view; b lateroventral view; c dorsal view; d posterior view; e anterior view; f laterodorsal view

The periotic MuMAB 240508. a lateral view; b medial view

Anatomy of the periotic MuMAB 240508. a dorsal view; b ventral view; c anterior view; d lateral view; e medial view; f posterior view
The dorsal surface of the anterior process is dorsally convex and the process itself is pyramidal in shape. The dorsal surface of the body of the periotic, above the pars cochlearis, is rough and dorsally convex in medial view. An elongated process protrudes medially from the postero-medial edge of the anterior process; the length of this protrusion is 37 mm and the diameter is 22 mm at its base. The apex of the protrusion is pointed resulting in a triangular shape of the protrusion in dorsal view. Anteroventrally to the protrusion, a wide concavity develops anteriorly, ventrally and dorsally (Fig. 5e). This concavity is 31 mm in anteroposterior length, 31 mm in dorsoventral heigth and its depth is 35 mm. Plesiobalaenoptera quarantellii is characterised by a similar concavity in the same position. Bisconti (2010) suggested that the concavity could result from the loss of the spongy bone located in this portion of the periotic due to taphonomic processes.
The suprameatal area gently descends from a raised dorsomedial border of the central portion of the periotic to the surface where the internal acoustic meatus opens. The suprameatal area is flat-to-slightly concave and shows presence of spongy bone that is still partly immersed in the matrix (Fig. 4). The internal acoustic meatus is a large concavity (Table 3) that comprises two smaller concavities: a concavity including the tractus spiralis foraminosus and the foramen singulare posteriorly, and the endocranial opening of the facial canal. These two concavities are separated by a thin crista transversa that is medially concave and does not reach the rim of the internal acoustic meatus. The depth of the posterior concavity is 34 mm; the tractus spiralis foraminosus is not completely exposed and appears as an elliptical structure located posteriorly on the floor of the concavity (Fig. 6). A small and almost circular foramen singulare is located close to the anterior wall of the posterior concavity. The dorsal edge of the internal acoustic meatus is circular in outline; posteroventrally, the outline forms a right angle with the ventral edge of the internal acoustic meatus that is straight.

Close-up view of the central portion of the periotic MuMAB 240508. a photographic representation; b anatomical interpretation
The endocranial opening of the facial canal is wide (Table 3) and its outline is uniformly convex giving the foramen an elliptical shape. On the opposite side of the periotic, the lateral opening of the facial canal is a small foramen (Table 3) that opens on the floor of a groove that widens posteriorly being well delimited by a ventral crest running above the oval window (Fig. 5d).
In medial view, a wide medial promontorial groove is present below the ventral rim of the internal acoustic meatus resembling that observed in the periotics of Plesio-balaenoptera quarantellii and Plesiobalaenoptera hubachi described by Bisconti (2010, 2011) and Bisconti et al. (2021b). The pars cochlearis as a whole is ventrally inflated and elongated both anteroposteriorly and transversely. Its anterior border, in ventral view, is an anteriorly convex groove for the tensor tympani muscle. The ventral surface of the pars cochlearis is almost completely flat with some grooves running anteroposteriorly that are probably due to post mortem damages.
In lateral view, a small and elliptical fossa for the head of the malleus is evident dorsally to the oval window; this fossa is dorsally delimited in part by the flange of the ventrolateral tuberosity. A small fossa incudis is located anteroventrally with respect to the fossa for the head of the malleus (Figs 4a and 5d). The oval window is elliptical in shape and relatively large (Table 3). Posterior to the oval window, a dorsoventrally running crest forms the posterior border of the fossa for the stapedial muscle that is concave and almost squared in outline. The caudal tympanic process of the pars cochlearis is rounded and slightly protruding. It forms the roof for the round window that is drop-like and large (Table 3). The round window is not separated from the perilymphatic foramen as a small, tube-like opening is present that begins from the medial border of the round window and widens medially, as seen in posterior view (Fig. 5f). In medial view, the perilymphatic foramen opens on the floor of a cavity with triangular outline in medial view. A crest separates this cavity from the cavity where the endolymphatic foramen opens; the latter is located more closely to the posterior edge of the internal acoustic meatus than the perilymphatic foramen (Figs. 5e and 6). Both the endolymphatic and perilymphatic foramina are small (Table 3) and elliptical in shape.
Posteriorly, the anterior portion of the stylomastoid fossa is observed on the posterior wall of the pars cochlearis. The stylomastoid fossa is shallow and concave and is sur-mounted by a protruding roof similar to a crest with rounded edge.
Reconstruction of body size
The ordinary least squares regression analysis resulted in the plot shown in Fig. 7a. The data were located around a regression line whose equation is the following:
-
(1)
(anterior process length) = 4.7192(total body length) + 25.009

Relationship between length of the anterior process and total body length in Balaenopteridae. a ordinary least squares regression analysis; b distribution of total body lengths in late Tortonian balaenopterid species; c distribution of total body lengths in late Tortonian balaenopterid specimens including individuals not still assigned to species or genera
Resolving the equation (1) by using 93 mm as anterior process length, provides a total body length of 14.40 m that corresponds to the length of the individual MuMAB 240508 belonged to. According to the operational definitions provided by Bisconti et al. (2021c) and literature therein), such a body length corresponds to a large-sized individual (body length included between 14 and 20 m). Despite the paucity of morphometric data, the value of R2 is relatively high (0.81775) suggesting that the relationship is well supported by the available data.
Table 2 shows that there are only ten balaenopterid species known from the stratigraphic interval between 9 and 7 Ma and a number of undetermined balaenopterid specimens mainly from Peru (Collareta et al. 2021; Bianucci et al. 2019; Esperante et al. 2015; Pilleri 1989). Our body size inference supports the fact that MuMAB 240508 is the largest in terms of body length (and mass). However, we must stress that the morphometric data are few and additional data would be desirable to better support the equation (1).
Comparisons
Compared to other Miocene balaenopterids, the periotic MuMAB 240508 shows peculiar characters that are not observed elsewhere. Comparisons are done here with taxa represented by specimens that share with MuMAB 240508 a comparable ontogenetic age (subadult to adult individuals). The two periotics of the holotype of the Tortonian Plesiobalaenoptera quarantellii are markedly different from MuMAB 240508 because they show a strongly raised, dome-like dorsal surface of the body of the periotic. Plesio-balaenoptera quarantellii shows a comparatively shorter and wider anterior process and a stockier pars cochlearis. The latter, in particular, appears to be comparatively shorter and more inflated than MuMAB 240508.
The periotic of the late Tortonian Norrisanima miocaena shows an anterior process characterised by a convex and rounded lateral border that is comparatively shorter and wider than that of MuMAB 240508 (Kellogg 1924; Leslie et al. 2019). Moreover, the anterior edge of the pars cochlearis of N. miocaena is protruding and crest-like, a character observed in the extant Megaptera novaeangliae. In medial view, the internal acoustic meatus of N. miocaena is small and the crista transversa is deeply recessed. The perilymphatic and endolymphatic foramina open in a common concavity (Leslie et al. 2019) but in MuMAB 240508 and in most of the other balaenopterid species they open in concavities separated from each other by a crest.
The periotic of the Tortonian Nehalaennia devossi is known only in ventral view because it is still articulated with the skull (Bisconti et al. 2019). The most striking differences between the periotic of Nehalaennia devossi and MuMAB 240508 is that, in the former, the pars cochlearis is enormous with respect to the size of the anterior process. Moreover, the flange of the ventrolateral tuberosity is triangular and protruding in N. devossi and slightly developed in MuMAB 20508. Finally, in N. devossi, the anterior attachment site for the tympanic bulla is short and located close to the ventrolateral tuberosity while in MuMAB 240508 it is developed along a large part of the lateroventral edge of the anterior process and is located more anteriorly.
The Messinian Miobalaenoptera numataensis shows a periotic characterised by a protruding pars cochlearis with posteriorly-located internal acoustic meatus (Tanaka and Watanabe 2019). The lateral tuberosity is indistinct in ventral view and the fossa for the head of the malleus is small.
The basal-most balaenopterid ‘Balaenoptera’ ryani shows a peculiar periotic with a strong posteromedial, triangular protrusion of the anterior process and a transversely narrow pars cochlearis (Hanna and McLellan 1924; MB pers. obs. on the holotype specimen CASG 1733). This periotic is highly unusual for a balaenopterid and shows a complex morphological pattern at the interface between the anterior process and the body of the periotic; there, a fissure marks the posterior border of the anterior process and the position of a sulcus that crosses the periotic transversely and continues on the suprameatal area. We hypothesise that this sulcus is homologous to the vascular groove described by Luo and Gingerich (1999) and Geisler and Luo (1998) in archaeocete periotics that allows the passage for the superior ramus of the stapedial artery.
MuMAB 240508 shares with Fragilicetus velponi the gently descending suprameatal area (Bisconti and Bosselaers 2016) and the confluence of the round window in the perilymphatic fossa. However, F. velponi differs from MuMAB 240508 in several respects: (1) narrower anterior process, (2) lower fossa for the head of the malleus, (3) crista transversa reaching the rim of the internal acoustic meatus, (4) thick crista transversa, (5) lack of medial protrusion from the anteromedial corner of the suprameatal area. The confluence of the round window in the perilymphatic fossa is observed in the extant Balaenoptera borealis (USNM 504699), B. musculus (AMNH 34869) and in Eschrichtius robustus (USNM 571931, SDNHM S-274). Among accepted fossil species, the confluence is observed in Protororqualus wilfriedneesi (Bisconti and Bosselaers 2020), Miobalaenoptera numataensis (Tanaka and Watanabe 2019), and Marzanoptera tersillae (Bisconti et al. 2021b).
In MuMAB 240508 both the groove for the tensor tympani muscle and the medial promontorial groove are present. These characters are plesiomorphic in Balaenopteridae as they are present in almost all the fossil species for which the periotic is known. Both characters are absent in the extant balaenopterid genera (Megaptera and Balaenoptera) in which the ventral border of the pars cochlearis approaches the ventral rim of the internal acoustic meatus in medial view (thus leaving no space for the development of the medial promontorial groove), and the anteroventral surface of the pars cochlearis gradually melts with the ventral surface of the anterior process without the interposition of structures related to the attachment of the tensor tympani muscle.
Peters et al. (2021) described the periotic of an unnamed balaenopterid whale from the late Tortonian (8.7-8.1 Ma) of Holland that received the accessory number Li 2153 at the Museum Klok & Peel in Asten (Holland); they supposed it could belong to a species similar to Archaebalaenoptera liesselensis described from the same sandpit from where this periotic comes. This periotic differs from MuMAB 240505 in having a thick and wide crista transversa and well spaced internal acoustic meatus and endocranial opening of the facial canal. Interestingly, both MuMAB 240508 and Li 2153 share the absence of a raised, dome-like central portion of the periotic, a dorsoventrally low anterior process and a deep and well-developed medial promontorial groove. However, the anterior process of MuMAB 240508 is proportionally more elongated and narrow and the distribution of the endocranial foramina differs enough to warrant that these periotics belonged to different balaenopterid species and, very likely, genera.
The internal acoustic meatus of MuMAB 240508 is similar to that of the extant Balaenopteridae in that it includes both the cavity for the tractus spiralis foraminosus and foramen singulare, and the endocranial opening of the facial canal that are separated from each other by a thin and recessed crista transversa. This pattern is observed in several extant and fossil balaenopterid species (e.g. Bisconti and Bosselaers 2020; Ekdale et al. 2011). Some fossil balaenopterid species show different patterns: (1) in Marzanoptera tersillae the crista transversa is thick and the endocranial opening of the facial canal is widely separated from the internal acoustic meatus; (2) in Fragilicetus velponi an intermediate pattern is observed in which the crista transversa is thick although not as in M. tersillae and the endocranial opening of the facial canal is located more closely to the internal acoustic meatus; (3) the current reconstruction of the endocranial foramina in Miobalaenoptera numataensis (Tanaka and Watanabe 2019) suggests that the internal acoustic meatus includes tractus spiralis foraminosus, foramen singulare and endocranial opening of the facial canal with the interposition of a very thin and recessed crista transversa however the presence of a foramen near to the anterior border of the medial surface of the pars cochlearis suggests that the endocranial opening of the facial canal could be located outside the internal acoustic meatus resembling the pattern described in M. tersillae. Alterna-tively, that foramen could represent a hiatus Falloppii. A CT scan of the periotic of Miobalaenoptera numataensis should allow to better describe the channels crossing the pars cochlearis in this species (in the same way as it was done for M. tersillae; Bisconti et al. 2021b) and solve the problem.
Phylogenetic analysis
The TBR search resulted in 32 equally parsimonious cladograms with CI=0.274, RI=0.748 and HI=0.726. The strict consensus (Nelsen) tree is shown in Fig. 8. The major mysticete radiations were recovered including Mysticeti, Chaeomysticeti, Balaenomorpha, Balaenoidea, Thalassotherii, Balaenopteroidea. Family-rank taxa were recovered as follows: Eomysticetidae, Balaenidae, Neobalaenidae, Cetotheriidae, Eschrichtiidae, Balaenopteridae and an unnamed clade including basal thalassotherian taxa. The bootstrap, symmetric resampling and Bremer support values were usually low (Fig. 8: numbers in bold represent BSV, numbers in regular font represent symmetric resampling supporting values, numbers in italics font represent Bremer supporting values) with the exception of the following clades within Mysticeti: Mysticeti, Chaeomysticeti, Balaenomorpha, Balaenoidea, Balaenidae. Additional relationships receiving high support values were the following: the genus Atlanticetus, Eschrichtius+Eschrichtioides, the genus Protororqualus, ‘Balaenoptera’ cortesii var. portisi, and Incakujira. The low support values found in this analysis may be explained by the large amount of homoplasy in the dataset. As previously discussed by Bisconti (2007), the problem of the homoplasy in the morphological datasets used in mysticete relationships is a major issue that will be difficult to overcome.

Strict consensus cladogram representing the phylogenetic relationships of baleen whales including MuMAB 240508. Numbers in bold represent bootstrap supporting values; numbers in regular font represent symmetric resampling supporting values; number in italics font represent Bremer supporting values
Eschrichitiidae is the sister goup of Balaenopteridae thus confirming the monophyly of Balaenopteroidea. The basal-most balaenopterid taxon is ‘Balaenoptera’ ryani. The next branch is represented by MuMAB 240508 that is the sister group of all the other balaenopterid species but ‘B.’ ryani. The basal position of MuMAB 240508 is supported by the lack of several synapomorphies, as reconstructed by using MESQUITE. In particular, it shows: (1) presence of medial promontorial groove that is absent in more crownward balaenopterids like Balaenoptera, Megaptera and Marzanoptera; (2) presence of a groove for the tensor tympani muscle that is absent in the extant balaenopterids Balaenoptera and Megaptera; (3) confluence of round window into the fossa for the perilymphatic foramen that is present in some archaic balaenopterids like Marzanoptera, Miobalaenoptera and Fragilicetus; (4) reduced size of the lateral tuberosity; and (5) rounded outline of the lateral tuberosity in dorsal view (Ekdale et al. 2011).
The relationships of the other balaenopterid species found with the present analysis confirm those already published by Bisconti et al. (2022) with a few exceptions. Probably the most important difference consists in the position of Plesiobalaenoptera that, in the present paper, belongs into a clade including Parabalaenoptera, Fragilicetus and Plesiobalaenoptera (confirming earlier results by Bisconti et al. 2021a, 2020, 2019; Bisconti and Bosselaers 2021) but, in Bisconti et al. (2022), it belongs to a separate clade that is sister group to a more crownward clade including Diunatans, Balaenoptera, their common ancestor and all the descendants of this ancestor. Additional differences include the relationships of Marzanoptera in that Marzanoptera bertae is not monophyletic with Marzanoptera tersillae in Bisconti et al. (2022) but it is in the present paper. Small changes in the relative positions of a number of taxa are expected because of the large amount of homoplasy in the dataset.
Discussion
The presence of Balaenopteridae in Mediterranean is ascertained in Tortonian sediments of northern Italy. In particular, the Salsomaggiore area provided fossils of three potentially different genera of fossil rorquals from localities along the Stirone River: MuMAB 240508, Plesiobalaenoptera quarantellii and a still unpublished associated skeleton of an archaic balaenopterid (M.B., pers. obs.). The stratigraphic analysis performed in the present paper supports a late Tortonian (i.e. from c. 9 to c. 7 Ma) age for both Plesiobalaenoptera quarantellii and MuMAB 240505. Nine balaenopterid species are currently accepted to be existent during the same stratigraphic interval worldwide. These include the species listed in Table 2. We did not include the periotic Li 2153 (Peters et al. 2021) in the table because its anterior process is incomplete and it was not possible to infer the total body length of the individual it belonged to. As shown in Table 2 and Fig. 7b, the total body length of eight species from a total of 10 falls in the range between 4 and 10 m. Norrisanima miocaena and MuMAB 240508 represent the largest specimens. If also undetermined balaenopterid specimens (see Table 2; Collareta et al. 2021; Bianucci et al. 2019; Esperante et al. 2015; Pilleri 1989) from the same stratigraphic range are added to the plot, it is even more evident that most of the specimens’ total body length falls in the range between 4 and 10 m (16 from a total of 19, representing the 84.21% of the total). Even in this case, MuMAB 240508 represents the largest specimen based on our inference of total body length. Based on the available data, MuMAB 240508 represents the largest balaenopterid specimen known from the late Tortonian and its total body length is almost twice than that exhibited by most of the coeval balaenopterid species.
Works on the evolution of mysticete body size published in the last decade suggested that the origin of gigantism in these whales occurred recently as a consequence of temperature decline in the latest Pliocene and Pleistocene (Slater et al. 2017). Statistical analyses on an extended dataset corroborated this hypothesis showing a clear trend towards large size only in the recent-most part of the mysticete evolution. However, an early study by Tsai and Kohno (2016) on Oligocene mysticetes suggested that the evolution of large body size (> 5 m in length in their work) was best interpreted as the result of independent selective processes acting on different rami of the phylogeny of early mysticetes. Bianucci et al. (2019) reinforced this hypothesis by adding evidence regarding the presence of a 13.7 m balaenopterid and similarly-sized, putative Pelocetus specimens in field works in Miocene Peruvian formations; these new data suggested the origin of mysticete gigantism as Early as Middle Miocene. Furthermore, Fordyce and Marx (2018) suggested that gigantic mysticetes were present already in the late Eocene as testified by the 8-m-long holotype of Llanocetus denticrentatus about 34 Ma.
It is evident that an operational definition of gigantism in mysticetes is necessary to proceed with this kind of research. Goldbogen et al. (2019) provided experimental and field evidence that only few species of extant mysticetes attain body lengths exceedingly large (20+ m) and they found that this may be due to the quantity and quality of food that they are able to exploit. Bisconti et al. (2021c) used such an approach and provided the operational definition of gigantism in mysticetes adopted in the present paper. By means of that approach, gigantic mysticetes are those whose total body length is more than 20 m. Following this line of reasoning, MuMAB 240508 should not represent a gigantic whale but can be considered a large-sized whale. All the other mysticetes from the late Tortonian are considered small-to-medium sized mysticetes and Fig. 7c shows that most of the specimens from this period have body lengths included between 8 and 10 m.
The disparity of body size observed in Tortonian mysticetes represents a possible case of niche partitioning following the suggestions provided by Tsai and Ando (2016) but much more work is necessary to better understand niche partitioning patterns and processes in extinct mysticete assemblages.
We suggest that the length of the anterior process of the periotic may be useful to get an estimate of the total body length in the absence of more reliable measurements (e.g. bizygomatic width, supraoccipital length, linear length of dentary; see Slater et al. 2017; Bisconti et al. 2021c; Pyenson and Sponberg 2011). Apparently, even with a limited amount of data, the relationship between the length of the anterior process and the total body length is more robust than that between the length of the tympanic bulla and the total body length (R2 = 0.4635 published by Groves et al., 2021) and is thus worth further investigations.
Conclusions
The periotic MuMAB 240508 was collected from the Tortonian of the Stirone River where also the holotype of Plesiobalaenoptera quarantellii and a still undescribed balaenopterid skeleton have been found. Morphologically, this periotic belongs to Balaenopteridae because it shows a triangular and pointed anterior process and an elongated pars cochlearis along both the transverse and the anteroposterior axes. The internal acoustic meatus is very close to the endocranial opening of the facial canal as the crista transversa is very thin and recessed. An analysis of the relationship between the length of the anterior process and the total body length resulted in the inference of 14.4 m for the individual documented by the periotic MuMAB 240508. This body length supports the hypothesis that the individual was large-sized and a comparison with all the known records of Tortonian balaenopterids revealed that it represents the largest balaenopterid individual known up to now from that stratigraphic interval. A phylogenetic analysis resulted in a basal position for MuMAB 240508 which is the sister group to all the other balaenopterids except for ‘Balaenoptera’ ryani.
Data availability
All the necessary data are published in the present paper.
References
Artoni, A., Papani, G., Rizzini, F., Calderoni, M., Bernini, M., Argnani, A., Roveri, M., Rossi, M., Rogledi, S., & Gennari, R. (2004). The Salsomaggiore structure (Northwestern Apennine foothills, Italy): a Messinian mountain front shaped by mass-wasting products. Geoacta, 3, 107–127.
Bianucci, G., Marx, F. G., Collareta, A., Di Stefano, A., Landini, W., Morigi, C., Varola, A. (2019). Rise of the titans: baleen whales became giants earlier than thought. Biology Letters, 15, 20190175. https://doi.org/10.1098/rsbl.2019.0175
Bisconti, M. (2001). Morphology and postnatal growth trajectory of rorqual petrosal. Italian Journal of Zoology, 68, 87–93.
Bisconti, M. (2007). A new basal balaenopterid from the Early Pliocene of northern Italy. Palaeontology, 50, 1103–1122.
Bisconti, M. (2010). A new balaenopterid whale from the Late Miocene of the Stirone River, northern Italy (Mammalia, Cetacea, Mysticeti). Journal of Vertebrate Paleontology, 30, 943–958.
Bisconti, M. (2011). New description of ‘Megaptera’ hubachi Dathe, 1983 based on the holotype skeleton held in the Museum für Naturkunde, Berlin. In M. Bisconti, A. Roselli, A. Borzatti de Loewenstern (Eds.), Climatic Change, Biodiversity, Evolution: Natural History Museum and Scientific Research. Proceedings of the Meeting, Quaderni del Museo di Storia Naturale di Livorno 23: 37–68.
Bisconti, M., & Bosselaers, M. (2016). Fragilicetus velponi: a new mysticete genus and species and its implications for the origin of Balaenopteridae (Mammalia, Cetacea, Mysticeti). Zoological Journal of the Linnean Society, 177, 450–474.
Bisconti, M., & Bosselaers, M. (2020). A new balaenopterid species from the southern North Sea Basin informs about phylogeny and taxonomy of Burtinopsis and Protororqualus (Cetacea, Mysticeti, Balaenopteridae). PeerJ, 8, e9570.
Bisconti, M., Lambert, O., & Bosselaers, M. (2013). Taxonomic revision of Isocetus depawi (Mammalia, Cetacea, Mysticeti) and the phylogenetic relationships of archaic ‘cetothere’ mysticetes. Palaeontology, 56, 95–127.
Bisconti, M., Munsterman, D.K., & Post, K. (2019). A new balaenopterid whale from the late Miocene of the Southern North Sea Basin and the evolution of balaenopterid diversity (Cetacea, Mysticeti). PeerJ, 7, e6915.
Bisconti, M., Munsterman, D.K., Fraajie, R.H.B., Bosselaers, M.E.J., & Post, K. (2020). A new species of rorqual whale (Cetacea, Mysticeti, Balaenopteridae) from the late Miocene of the southern North Sea Basin and the role of North Atlantic in the paleobiogeography of Archaebalaenoptera. PeerJ, 8, e8315.
Bisconti, M., Damarco, P., Mao, S., Pavia, M., & Carnevale, G. (2021a). The earliest baleen whale from the Mediterranean: large-scale implications of an early Miocene thalassotherian mysticete from Piedmont, Italy. Papers in Palaeontology, 2020, 1–20.
Bisconti, M., Damarco, P., Pavia, M., Sorce, B., & Carnevale, G. (2021b). Marzanoptera tersillae, a new balaenopterid genus and species from the Pliocene of Piedmont, north-west Italy. Zoological Journal of the Linnean Society, 192, 1253–1292.
Bisconti, M., Ochoa, D., Urbina, M., & Salas-Gismondi, R. (2022). Archaebalaenoptera eusebioi, a new rorqual from the late Miocene of Peru (Cetacea, Mysticeti, Balaenopteridae) and its impact in reconstructing body size, evolution, ecomorphology and paleobiogeography of Balaenopteridae. Journal of Systematic Palaeontology, 19, 1129–1160.
Bisconti, M., Pellegrino, L., & Carnevale, G. (2021c). Evolution of gigantism in right and bowhead whales (Cetacea, Mysticeti, Balaenidae). Biological Journal of the Linnean Society, 134, 498–524.
Boessenecker, R. W. (2013). A new marine vertebrate assemblage from the Late Neogene Purisima Formation in Central California, part II: pinnipeds and cetaceans. Geodiversitas, 35, 815–940.
Bosio, G., Collareta, A., Di Celma, C., Lambert, O., Marx, F. G., de Muizon, C., Gioncada, A., Gariboldi, K., Malinverno, E., Varas Malca, R., Urbina, M., Bianucci, G. (2021). Taphonomy of marine vertebrates of the Pisco Formation (Miocene, Peru): Insights into the origin of an outstanding Fossil-Lagerstätte. PLoS ONE, 16(7), e0254395. https://doi.org/10.1371/journal.pone.0254395
Brisson, A.D. (1762). Regnum animale in classes IX Distributum, sive synopsis methodica. Lugdum Batarorum, apud. Leiden: Theodorum Haak.
Ekdale, E.G., Berta, A., & Deméré, T. A. (2011). The comparative osteology of the petrotympanic complex (ear region) of extant baleen whales (Cetacea: Mysticeti). PLoS ONE, 6, e21311. https://doi.org/10.1371/journal.pone.0021311
Esperante, R., Brandt, L. R., Chadwick, A. V., Poma, O. (2015). Taphonomy and paleoenvironmental conditions of deposition of fossil whales in the diatomaceous sediments of the Miocene/Pliocene Pisco Formation, southern Peru—A new fossil-lagerstätte. Palaeogeography, Palaeoclimatology, Palaeoecology, 417, 337–370.
Flower, W.H. (1864). Notes on the skeletons of whales in the principal museums of Holland and Belgium, with descriptions of two species apparently new to science. Proceedings of the Zoological Society of London, 1864, 384–420.
Fordyce, R. E., & Marx, F. G. (2018). Gigantism precedes filter feeding in baleen whale evolution. Current Biology, 28, 1670–1676.
Geisler, J. H., & Luo, Z. (1998). Relationships of Cetacea to terrestrial Ungulates and the evolution of cranial vasculature in Cete. In J. G. M. Thewissen (Ed.), The emergence of whales.Evolutionary patterns in the origin of Cetacea (pp.163-212). New York: PlenumPress.
Geisler, J.H., & Sanders, A.E. (2003). Morphological evidence for the phylogeny of Cetacea. Journal of Mammalian Evolution, 10, 23–129.
Goldbogen, J. A., Cade, D. E., Wisniewska, D. M., Potvin, J., Segre, P. S., Savoca, M. S., Hazen, E. L., Czapanskiy, M. F., Kahane-Rapport, S. R., Deruiter, S. L., Gero, S., Tønnesen, P., Gough, W. T., Hanson, M. B., Holt, M. M., Jensen, F. H., Simon, M., Stimpert, A. K., Arranz, P., Johnston, D. W., Nowacek, D. P., Parks, S. E., Visser, F., Friedlaender, A. S., Tyack, P. L., Madsen, P. T., & Pyenson, N. D. (2019). Why whales are big but not bigger: physiological drivers and ecological limits in the age of ocean giants. Science, 366, 1367–1372.
Goloboff, P. A., & Catalano, S. A. (2016). TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics, 32, 231–238.
Gray, J.E. 1864. On the Cetacea which have been observed in the seas surrounding the British Islands. Proceedings of the Scientific Meetings of the Zoological Society of London, 1864, 195–248.
Gray, J.E. (1868). Synopsis of the Species of Whales and Dolphins in the Collection of the British Museum. London: Bernard Quaritch.
Groves, S. L., Peredo, C. M., & Pyenson, N. D. (2021). What are the limits on whale ear bone size? Non-isometric scaling of the cetacean bulla. PeerJ 9, e10882 https://doi.org/10.7717/peerj.10882
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4: 4.
Hanna, G. D., & McLellan, M. E. (1924). A new species of fin whale from the type locality of the Monterey Group. Proceedings of the California Academy of Sciences, 13, 237–241.
Kellogg, R. (1924). Description of a new genus and species of whalebone whale from the Calvert Cliffs, Maryland. Proceedings of the United States National Museum, 63, 1–14.
Leslie, M. S., Peredo, C. M., & Pyenson, N. D. (2019). Norrisanima miocaena, a new generic name and redescription of a stem balaenopteroid mysticete (Mammalia, Cetacea) from the Miocene of California. PeerJ, 7, e7629.
Lin, C. H., Brzobohaty, R., Nolf, D., & Girone, A. (2017). Tortonian teleost otoliths from northern Italy: taxonomic synthesis and stratigraphic significance. European Journal of Taxonomy, 322, 1–44.
Linnaeus, C. (1758). Systema naturae. Holmiae (Stockholm), Sweden: Salvii.
Lirer, F., Foresi, L. M., Iaccarino, S. M., Salvatorini, G., Turco, E., Cosentino, C., Sierro, F. J., & Caruso, A. (2019). Mediterranean Neogene planktonic foraminifer biozonation and biochronology. Earth-Science Reviews, 196, 1–36.
Luo, Z., & Gingerich, P.D. (1999). Terrestrial Mesonychia to aquatic Cetacea: transformation of the basicranium and evolution of hearing in whales. University of Michigan, Papers in Paleontology, 31, 1–98.
Maddison, W. P., & Maddison, D. R. (2021). Mesquite: a modular system for evolutionary analysis. Version 3.70 http://www.mesquiteproject.org
Marasti, R., (1973). La fauna tortoniana del Torrente Stirone (Limite Parmense - Piacentino). Bollettino Società Paleontologica Italiana, 12, 76–120.
Martini, E. (1971). Standard Tertiary and Quaternary Calcareous Nannoplankton Zonation. In A. Farinacci (Ed.), Proceedings of the 2nd Planktonic Conference (pp. 739–785). Edizioni Tecnoscienza.
Marx, F. G., & Kohno, N. (2016). A new Miocene baleen whale from the Peruvian desert. Royal Society Open Science, 3(10), 160542. https://doi.org/10.1098/rsos.160542.
Mead, J.G., & Fordyce, R.E. (2009). The therian skull. A lexicon with emphasis on the odontocetes. Smithsonian Contributions to Zoology, 627, 1–248.
Mitchell, E.D. (1989). A new cetacean from the late Eocene La Meseta Formation, Seymour Island, Antarctic Peninsula. Canadian Journal of Fisheries & Aquatic Sciences, 46, 2219–2235.
Monegatti, P., & Raffi, S. (2010). The Messinian marine molluscs record and the dawn of the eastern Atlantic biogeography. Palaeogeography, Palaeoclimatology, Palaeoecology, 297, 1–11.
Negra O., & Lanzinger M. (2014). Lo scheletro di balenottera comune, Balaenoptera physalus, esposto al MUSE (Museo delle ScienzE di Trento): un caso insolito di cetaeo in un museo alpino. Museologia Scientifica Memorie, 12, 106–109.
Nicolosi, P., Braschi, S., Cagnolari, L., & Zuffi, M.A.L. (2014). Il patrimonio di Cetacei attuali del Museo di Storia naturale dell’Università di Pisa (Certosa di Calci). Profilo storico e catalogo della collezione. Museologia Scientifica Memorie, 12, 215–238.
Peters, N., Munsterman, D., & Post, K. (2021). A late Miocene balaenopterid petrotympanic from Liessel (The Netherlands). Cainozoic Research, 21, 165–172.
Pilleri, G. (1989). Beiträge zur Paläontologie der Cetaceen Perus. Vammala: Hirnanatomisches Institut Ostermundigen.
Pyenson, N. D., Sponberg, S. N. (2011). Reconstructing body size in extinct crown cetacea (Neoceti) using allometry, phylogenetic methods and tests from the fossil record. Journal of Mammalian Evolution, 18, 269–289.
Slater, G. J., Goldbogen, J. A., & Pyenson, N. D. (2017). Independent evolution of baleen whale gigantism linked to Plio-Pleistocene ocean dynamics. Proceedings of the Royal Society B: Biological Sciences, 284(1855), 20170546. https://doi.org/10.1098/rspb.2017.0546
Tanaka, Y., & Watanabe, M. (2019). An early and new member of Balaenopteridae from the upper Miocene of Hokkaido, Japan. Journal of Systematic Palaeontology, 17, 1417–1431.
Tsai C.-H. and Ando, T. (2016). Niche partitioning in Oligocene toothed mysticetes (Mysticeti: Aetiocetidae). Journal of Mammalian Evolution, 23, 33–41.
Tsai C.-H. and Kohno, N. (2016). Multiple origins of gigantism in stem baleen whales. The Science of Nature, 103, 89. https://doi.org/10.1007/s00114-016-1417-5
Zermani, A. (2001). Biostratigrafia a Foraminiferi planctonici della successione Burdigaliano – Serravalliano del Torrente Stirone. Università degli Studi di Parma, unpublished thesis.
Zeigler, C. V., Chan, G. L., & Barnes, L. G. (1997). A new late Miocene balaenopterid whale (Cetacea: Mysticeti), Parabalaenoptera baulinensis, (new genus and species) from the Santa Cruz Mudstone, Point Reyes Peninsula, California. Proceedings of the California Academy of Sciences, 50, 115–138.
Acknowledgements
Many thanks are due to Graham and Margareth Avery and to Leonard Compagno (IZIKO), Annelise Folie, Alain Dreze and Olivier Lambert (IRSNB), Nancy Simmons, Richard Monk, Eric Brothers and Eileen Westwig (AMNH), and Dave Bohaska (USNM) for granting access to the specimens under their care. Toshiyuki Kimura and Cheng-Hsiu Tchai provided detailed reviews that greatly improved the paper and we want to thank them very much. We want to thank Peter Koenigshof (Editor-in-Chief of Palaeobiodiversity and Palaeo-environments) and Sinje Weber (Managing Editor) for the detailed work done that improved the quality of this paper. Different parts of this work were financed by ex-60% funds of the University of Torino, the Natuurmuseum Rotterdam (2007), a Collection Study Grant from AMNH (2005) and two Synthesys grants (Project BE-TAF 305 and BE-TAF 3057) which were financed in 2009 and 2013 to MB by the European Community Research Infrastructure Action under the FP 7.
Funding
Open access funding provided by Università degli Studi di Torino within the CRUI-CARE Agreement. This research was funded by grants (ex-60%, 2021) from the University of Turin, the Natuurmuseum Rotterdam (2007), a Collection Study Grant from AMNH (2005) and two Synthesys grants (Project BE-TAF 305 and BE-TAF 3057) which were financed in 2009 and 2013 to MB by the European Community Research Infrastructure Action under the FP 7.
Author information
Authors and Affiliations
Contributions
P. M. and G. R. provided the geological setting; G. R. curated the specimen; G. T. realised the photogrammetry of Plesiobalaenoptera quarantellii; M. B. studied the specimen, performed the analyses, prepared the illustrations and wrote the paper; G. C. coordinated the research group and contributed to the discussion.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix 1
Appendix 1
Morphological coding of MuMAB 240508 for phylogenetic analysis.—Character list and taxon x character matrix provided by Bisconti et al. (2020a).
????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? 1110121111 000--001-0 112?0?00?0 11110?1011 01???????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ??????
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bisconti, M., Raineri, G., Tartarelli, G. et al. The periotic of a basal balaenopterid from the Tortonian of the Stirone River, northern Italy (Cetacea, Mysticeti, Balaenopteridae). Palaeobio Palaeoenv 103, 663–679 (2023). https://doi.org/10.1007/s12549-022-00550-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-022-00550-2