
Abstract
The treefall pit of a large tree at the base of the middle Eocene Mittelkohle has been studied in the former open-cast mine Neumark Nord of the Geiseltal Mining District. Above a light clastic soil, the sedimentation in the pit started with backfall and downwashed material grading upwards into pond deposits that filled the pit. This represents a high-resolution section equivalent in time to the initial peat-forming environment. Palynology revealed a striking difference between a fern-dominated herbaceous plant community below the pit and a pulse of washed-in fagaceous pollen (Tricolpopollenites liblarensis) at the base of the pit. This is overlain by an assemblage of woody taxa typical of a peat swamp forest associated with a number of tropical elements. Comparison with a nearby but undisturbed transition from underlying sediments to the seam confirms the ecotonal character of the parent plant of T. liblarensis, and suggests that Cupressaceae s.l., Nyssaceae and Myricaceae especially benefited from moisture and light in the clearing of the treefall pit. The transition from the underlying sediment to the lignite represents a sharp break in the sedimentary regime from clastic to purely organic material with a bounding surface colonised by large trees, most of them preserved as individual stumps, but including some fallen logs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
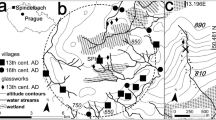
The valley of the Geisel creek (Geiseltal) is located about 20 km south of the city of Halle an der Saale (Saxony-Anhalt, Central Germany). Until the 1980s, the Paleogene lignites of the Geiseltal Mining District, which are limited to an area of about 5 by 9 km (Fig. 1), were one of Germany’s most productive coal resources. Locally, they even consisted of an almost continuous seam of more than 100 m in thickness. In the scientific community the lignites of the Geiseltal are especially well-known for the wealth of exceptionally well-preserved plant and vertebrate fossils which were recovered from a number of individual sites (Krumbiegel et al. 1983). After most of the lignites had been removed by surface mining, the open pits were abandoned in 1993, leaving marginal exposures and stripped planar surfaces. Prior to flooding of the area, they offered the last opportunity to document geological and palaeontological information during several field campaigns between 2000 and 2006 (Hellmund and Wilde 2001; Wilde and Hellmund 2006). None of the outcrops most recently studied occurs in the known distribution of vertebrate sites as summarised by Krumbiegel (1977). Thus, it is not surprising that skeletal and other animal remains have not been recovered, and palaeontological information is limited to some plant remains and palynomorphs.

The Geiseltal Mining District at the time of the field campaigns of 2001 (modified from Wilde and Riegel 2010)
Treefall as a result of external (e.g. wind, weight of ice/snow cover) or internal factors (e.g. infestation of a tree by fungi or insects), or a combination of both (e.g. Maser et al. 1988; Schaetzl et al. 1989), has a vital influence on forest structure and succession, especially by causing gaps in the forest, which permit light to reach the forest floor (e.g. Čuchta 2020; Gastaldo and Staub 1999; Schupp et al. 1989; Ulanova 2000). Furthermore, nutrient cycling is influenced by uprooted soil material and accumulation of plant litter. Pits and mounds resulting from treefall can result in lasting effects on microtopography (e.g. Harrison-Day and Kirkpatrick 2019) and microclimate directly influencing the character and regeneration of the local vegetation (e.g. Khanina et al. 2017; Muscolo et al. 2014; Schaetzl et al. 1989; Schupp et al. 1989; Ulanova 2000). If the groundwater table is near to the surface, treefall pits may be temporally or permanently filled with water (e.g. Bobrovsky and Loyko 2016; Gastaldo and Staub 1999; Langohr 1993; Verb and Rubino 2012), leading to a succession of aquatic and wet ground communities (e.g. Bobrovsky and Loyko 2016; Verb and Rubino 2012).
Mining activities at the southern edge of the former open-cast mine “Neumark Nord” had removed the lignite down to the base of the seam or slightly below, irregularly exposing the surface of the underlying clays and silts with basal cross sections of stems, stumps and root stocks of trees (combined as “stumps”). These mark the onset of peat accumulation and seam formation. In early autumn of 2001, the distribution of “stumps” was recorded in an area of about 75 × 75 m (Fig. 2). One of the stumps and the cross section of a fallen tree just at the transition from the underlying sediments to the coal has already been figured by Wilde and Hellmund (2006). Simoneit et al. (2020) document resin that has been recovered from three of the “stumps”, suggesting an origin from Cupressaceae s.l. (Taxodium, stump 13), Burseraceae (stump 35) and Dipterocarpaceae (stump 46), respectively. The subject of the present palynological study is the treefall pit of a fallen tree and its infill (section G VIII, stump 40). Remnants of the basal part of the seam which were left intact at the margin of the area occur within a relatively short distance to the root pit and have been studied for comparison (section G VII F).

Sketch of the stripped surface (G VIII) with numerous remnants of tree bases (“stumps”) and the studied fallen tree (stump 40); stumps 13, 35 and 46 have been studied geochemically by Simoneit et al. (2020)
Geology and stratigraphy
The middle to upper Eocene coal-bearing sequence of the Geiseltal formed in a semi-isolated basin which was largely caused by subsurface solution of Lower Triassic (Röt) evaporites (Küstermann et al. 2008). Based on a continuous record of vertebrate faunas, the Geiseltal lignites are correlated with European standard Mammal Paleogene zones (MP zones). The lower and middle lignites (“Unterkohle” and “Mittelkohle”, respectively) are thus regarded as middle Eocene (MP 11 to MP 13) and the upper lignite (“Oberkohle”) as late Eocene (mainly MP 14) in age (Haubold and Hellmund 1994; Haubold and Thomae 1990). According to field observation, the surface with the treefall pit most probably represents the base of the Mittelkohle, which is placed within the regional pollen zone 15, subzone B (Krutzsch 1976) (Fig. 3), which covers an approximate age between 43 and 45 million years (Standke 2008). In spite of the fact that the exact correlation of the Geiseltal faunas to the European mammal chronology has recently been questioned (Ring et al. 2020), our sections can still be regarded as middle Eocene in age.

Methods
Two trunks of fallen trees prostrate in different directions have been encountered between the many “stumps” exposed on the planar surface at the base of the Mittelkohle (Fig. 2). When turning over, the largest of them (stump 40) had produced a more or less circular treefall pit that showed a clayey infill on the exposed surface. To study the connection between the fallen tree and the pit, the depth of the pit and its filling in detail, a trench was dug by hand following the direction of the fallen tree in the middle. The side wall including a section of the base of the tree and the pit was cleaned and smoothed with palette knives and documented by photographs and field drawing (Fig. 4). Above the underlying soil, three layers have been distinguished in the centre of the pit, and seven samples were taken from the whole section for palynological study at less than 6-cm intervals over a vertical distance of 40 cm. In addition, the undisturbed transition from interbed to seam was measured, and 11 samples were taken in a short section ca. 50 m away (section G VII F). Both series of samples were numbered independently from bottom to top according to individual layers.

Trench face (section G VIII, stump 40) with position of samples indicated by numbers. a Sketch. b Photo
Palynological preparation followed standard procedures for siliciclastic sediments, i.e. brief boiling with 10% H2O2 for particle disaggregation, several days of treatment with 30% HF for removal of silica and clay minerals, removal of HF by several steps of decanting and final sieving through a 10-μm mesh screen. Quantitative determination of assemblage composition is based on counts between 400 and 600 grains from three slides per sample and represented as percentages of total pollen and spores. In addition, fungal remains and resin particles were counted and calculated as additional percentages based on 100% of total pollen and spores. The use of additional percentages based on a certain basic pollen sum of 100% as proposed, e.g. by Straka (1975), is common in Quaternary palynology (e.g. Urban et al. 2011; Van Geel 1978). Because of their great abundance in several samples, an exponential scale was used for representation in the respective figures.
Conventional light microscopy (LM) images were obtained by a LEITZ Metallux 3 microscope equipped with interference contrast and a LEICA DFC490 digital camera. For scanning electron microscopy (SEM), individual grains were selected and picked from uncovered spreads of suspended residue in glycerol under LM control, washed twice in a drop of distilled water, and transferred to the emulsion-free side of a piece of developed photographic film that was mounted on a standard SEM-stub. The stubs were sputtered with gold and studied in a JEOL FM6490 LV SEM by using a 20 kV beam in secondary electron mode. Pieces of apparently well-preserved wood were examined by SEM. They were removed from the middle of the preserved tree length and broken in the lab by using tweezers and a scalpel to reveal the three planes needed (transverse, radial and tangential) for potential systematic determination. The fragments were mounted on standard SEM stubs by a commercial carbon adhesive, sputtered with gold, and also studied in the JEOL FM6490 LV SEM (20 kV, secondary electron mode).
The material for the present publication is deposited in the collections of the Palaeobotanical Section of the Senckenberg Forschungsinstitut und Naturmuseum in Frankfurt.
Results
Figure 2 shows an area of about 5600 m2 at the base of the middle Eocene Mittelkohle with basal remnants of 81 trees of various size (“stumps”), including the studied example of an uprooted tree that left a treefall pit at its base (stump 40). The exposed surface in the area of the fallen log represented a level slightly below the very base of the lignite. This is indicated by the fact that no lignite remained on the surface, not even in the centre of the filling of the treefall pit and that only the lower part of the trunk was preserved. The diameter of the treefall pit at the exposed surface was abou 4 m, and the remaining maximum depth in the trench was about 40 cm (Fig. 4). Remnants of the fallen trunk could be traced from the treefall pit for a length of about 23 m on the weathered surface, including gaps which were caused by differential stripping due to mining activities and subsequent weathering. Therefore the trunk was not preserved as a singular unit and had mostly fallen apart into a series of longitudinal sections. No obvious signs of branching were observed, but branches were likely stripped and weathered away together with the upper part of the stem. The remains of the stem body are preserved as compressed lignitic mummification in the sense of Mustoe (2018) that does not show much anatomical detail and therefore precludes systematic assignment (Supplementary Material 1). An undisturbed transition from the underlying sediment to the lignite seam could be studied for comparison at a nearby short section (G VII F, distance about 50 m), where it was not removed by mining.
Lithology
The root pit (Figs. 4 and 5) is underlain by a light grey sandy clay to clayey sand (sample 1), which is densely rooted (root diameter < 1 cm) and shows occasional brown patches and nodules. Interspersed are several thick, mainly flat lying lignitic root branches (diameter up to about 10 cm). The following layer (sample 2) represents the base of the pit fill and consists of brownish sandy material probably mainly washed down from the underlying sediment which initially remained between the roots of the fallen tree. The succeeding layer consists of a light grey silty clay (samples 3 and 4) that grades into a carbonaceous and darker silty clay (samples 5 and 6). The top layer (sample 7), which is only preserved in the centre of the depression, is dark grey to almost black, highly carbonaceous and brittle probably due to recent surface weathering. Higher parts of the filling and the contact to the overlying coal are missing because it had been removed by mining. In general, the filling of the pit becomes more fine grained and organic-rich to the top, but macroscopic plant material has not been observed.

Palynological diagram of the treefall pit section G VIII, stump 40, with position of samples (for description of the section see text and Fig. 4)
The undisturbed nearby section (Fig. 6) does not include the mottled light grey sandy clay that we observed in the underlying soil of the treefall pit. Instead, the top of the rooted underlying sediment up to the very base of the seam is dominated by coarser grain size including fine gravel, but appears slightly more transitional with respect to colour due to an increasing content of organic matter. This section further continues into the succeeding lignite for about 40 cm.

Nearby section G VII F with palynological diagram
Palynology
During the study eight species of spores and 67 species of pollen were identified in the sections (list of taxa and quantitative data see Supplementary Material 2-4). The palynological diagram from the site of the treefall pit (Fig. 5) shows some striking vertical trends in assemblage composition. Palynomorphs are rather rare and often poorly preserved in the basal sample (sample 1), which represents the soil of the initial mire forest (seat earth). There, monolete fern spores (Laevigatosporites, Verrucatosporites) are highly dominant, although somewhat masked by the abundance of resin particles. Laevigatosporites type spores (Fig. 7d, e) are in most cases rather small with more than 80% in the range of 25-39 μm (size distribution see Supplementary Material 5). According to Krutzsch (1967), most of them can either be assigned to Laevigatosporites gracilis Webster and Wilson (size below 30 μm; Krutzsch 1967) or to Laevigatosporites haardtii (R. Potonié and Venitz) Thomson and Pflug 1953 (size range about 35–40 μm; Krutzsch 1967). Since the size distribution of 83 individual spores shows a distinct peak between 25 and 40 μm, which covers both species, priority should here be given to the older name, L. haardtii. The size distribution in Laevigatosporites in sample 5 (Supplementary Material 5) is more even and extends well into dimensions beyond 60 μm that conforms to the range of L. discordatus Thomson and Pflug 1953 (Krutzsch 1967). Slightly verrucate monoletes are also quite common and are identified as Verrucatosporites microfavus Thiele-Pfeiffer 1988 (Fig. 7f). In addition to monolete spores, there is a variety of tricolpate and tricolporate pollen, which have in most cases not been more closely identified due to poor preservation or insecure taxonomic assignment of rare or single occurrences. Unique for the section, though rare, is the occurrence in this sample of alete verrucate spores fitting to the genus Corrusporites Krutzsch, 1967 (Fig. 7a–c) which has been assigned to bryophytes. These spores appear similar to those of several extant bryophyte taxa (compare, e.g. Boros and Járai-Komlódi 1975; Zhang et al. 2005).

Spores and pollen from the treefall pit section G VIII, stump 40. a–c Corrusporites sp. d–e Laevigatosporites haardti. f Verrucatosporites microfavus. g–i Inaperturate pollen. j–l Punctilongisulcites microechinatus. (a, d, g LM, interference contrast. j LM, interference contrast multifocus image. b, c, e, f, h, i, k, l SEM, secondary electron mode)
The abundance of monolete spores drops dramatically in the bottom layer of the pit fill (sample 2). Most noteworthy is the singular and isolated sharp peak of Tricolpopollenites liblarensis reaching 16% against a background of less than 3% in subsequent samples. Many of the more important pollen taxa shown in the diagram begin to rise in abundance or appear here for the first time. A marked increase from 3.7 to 12.5% is shown by inaperturates (“Taxodiaceae”, now with Cupressaceae s.l.) (Fig. 7g–i) and from 2 to 15.2% by Tricolporopollenites cingulum (Fagaceae). Both taxa rise in abundance almost continuously in the succeeding five samples. Juglandaceous pollen, especially Momipites, Platycaryapollenites and Plicatopollis plicatus, rises moderately to 5.2%, roughly maintaining this level throughout the remainder of the section (maximum 7.9%).
A distinct change occurs in sample 3 with the sudden rise to dominance of myricaceous pollen totalling 24% of the pollen assemblage. Although most of the myricaceous type pollen in our sections can generally be assigned to Triatriopollenites bituitus, similar triporate taxa (e.g. T. rurensis, Triporopollenites robustus, T. rhenanus) were lumped with the species since preservation and variability limited consistent distinction. The loss of about two-thirds of myricaceous pollen in the succeeding samples 4 and 5 is largely compensated by gradual increases of inaperturates, T. cingulum and Nyssapollenites kruschii.
Sample 6 stands out by the absence of N. kruschii and a singular show of palm pollen (Monocolpopollenites tranquillus; 6.3%). Finally, sample 7 from the highly carbonaceous top layer is dominated by inaperturates and T. cingulum, which together make up more than 50% of the assemblage.
Nyssapollenites kruschii (Fig. 8g–i) seems to parallel the trends of inaperturates and Tricolporopollenites cingulum initially reaching nearly 10% in sample 5, but almost disappears in the top two samples. A number of typically middle Eocene taxa, e.g. Porocolpopollenites vestibulum (Fig. 8d–f), Tricolporopollenites vancampoae (Fig. 9a–c), Tricolporopollenites marcodurensis and Spinaepollis spinosus (Fig. 9j–l), are rare, although P. vestibulum and Tricolp(or)opollenites parmularius are somewhat more common in the lower and S. spinosus and T. marcodurensis more common in the upper part. The occurrence of Tricolporopollenites willrathae (Fig. 9 g–i) is typical for pollen zone 15 in the region (Krutzsch, 1970, 1992) and substantiates an assignment of the treefall pit to this zone. The purported affiliation of T. mansfeldensis (Fig. 8a–c) to the mangrove genus Rhizophora by Krutzsch (1969), however, is in conflict with all other environmental indices of the site and suggests a different affinity, e.g. Euphorbiaceae (Gruas-Cavagnetto and Köhler 1992; Thiele-Pfeiffer 1988).

Pollen from the treefall pit section G VIII, stump 40. a–c cf. Tricolporopollenites mansfeldensis. d–f Porocolpopollenites vestibulum. g-i: Nyssapollenites kruschi. j–l Tetracolporopollenites sp. (a, d, g, j LM, interference contrast. b, c, e, f, h, i, k, l SEM, secondary electron mode)

Pollen from the treefall pit section G VIII, stump 40. a–c Tricolporopollenites vancampoae. d–f Tricolporopollenites pseudointergranulatus. g–i Tricolporopollenites willrathae. j–l Spinaepollis spinosus. (a, d, j LM, interference contrast. g LM, interference contrast multifocus image. b, c, e, f, h, i, k, l SEM, secondary electron mode)
Similar to juglandaceous pollen, no vertical trends can be observed from samples 2 to 7 for both, the T. retiformis group, in which various tricolpate reticulate taxa are combined, and the genus Tetracolporopollenites (mostly Sapotaceae) (Fig. 8j–l, compare Harley 1991), which also includes more than a single species as well.
Figure 10 illustrates the high proportion of resin particles in the two basal samples, particularly in sample 2. The resin particles are deep orange in colour, and mostly rectangular in shape, and have probably been released from decomposing roots of the fallen tree. A change in shape and colour of resin particles occurs in the upper two samples, in which they are pale, roundish droplike and, in part, vacuolated. In view of the massive occurrence of resin particles, their quantitative assessment in palynological preparations can only be considered as an approximation, which, nevertheless, appears highly significant. In the two lowermost samples, resin particles overwhelmingly mask the palynological residues. Although they remain very abundant in succeeding samples, a sharp drop occurs in sample 3 followed by a gradual decrease to the top as a normal forest mire appears to become established.

Light microscopical image of resin particles in palynological residue from section G VIII, stump 40, sample 2
The apparent abundance of fungal remains, mainly spores, in the underclay (sample 1) may be an artefact rather than really due to the absolute scarcity and poor preservation of pollen and spores, on the basis of which their percentage frequency is calculated. But in samples higher in the fill, their frequency increases almost consistently to the top of the pit infill.
Discussion
The Geiseltal lignites are punctuated by sinkholes, which developed concurrently with peat formation due to karstification in the subsurface (Krumbiegel 1959, 1977). They are commonly associated with the formation of small pools in the order of 10 m in diameter, which are one of the major traps for the famous Geiseltal fossils (Krumbiegel 1977) and shown on various reconstructions of a lush tropical vegetation and highly diverse middle Eocene biota. They are also sufficiently large to reveal a clear hydrosere from limnic to telmatic environments within the original Geiseltal mire (Krumbiegel et al. 1983). Instead, treefall pits differ greatly in dimension, duration, and ecologic effect and have not been described from the Geiseltal Eocene, thus far. They nevertheless leave an ecologic fingerprint in closed forests in consequence of canopy gaps and depressions on the floor as studies of recent sites (e.g. Čuchta 2020; Schupp et al. 1989; Ulanova 2000) and rare fossil examples suggest (Gastaldo and Staub 1999).
Figure 2 shows the studied area of about 5600 m2 at the base of the middle Eocene Mittelkohle with 81 “stumps” of various sizes, including two trees that appear to have fallen in opposite directions. If windthrow was the cause, rather than the death of an individual tree, it therefore only affected individual trees with rather limited damage to the forest, leaving an areally restricted clearing and a mound-and-pit structure. Brown patches and nodules in the underlying clayey sand to sandy clay of the studied pit indicate a wet soil (gley; Bobrovsky and Loyko 2016) which may have favoured uprooting and the succeeding formation of a pond in the pit (Langohr 1993). Root failure (Gastaldo 1990; Piirto et al. 1984) affected even roots of considerable thickness which remained in the sediment below the pit. Downwash from sediment captured between the overturned roots may have started quickly (Gastaldo 1990). But, how small a clearing may be, it will be the site for secondary recovery within the damaged forest (e.g. Langohr 1993). Our study attempts to trace such a recovery in the middle Eocene by means of palynology. The depth of our pit is well in accordance with the fact that a depth of 1.5 m is rarely exceeded by uprooting in modern trees (e.g. Bobrovsky and Loyko 2016; Clinton and Baker 2000; Putz 1983). The filling appears similar to the situation outlined by Langohr (1993), his fig. 2) with a mixture of uprooted soil material on one side and a pond fill on the other. Mound-and-pit structures of fallen trees can last from decades (e.g. Lutz 1940; Putz 1983) to hundreds or more than thousand years (e.g. Harrison-Day and Kirkpatrick 2019; Šamonil et al. 2013; Schaetzl and Follmer 1990); however, a more exact time frame for the filling of our pit can hardly be inferred from the data at hand. It is interesting to note that no plant litter has been observed in the studied part of the fill. This may be explained either by rapid degradation as observed even under the tropical climate of Panama (Putz 1983), or accumulation of litter as discussed by Gastaldo and Staub (1999) for treefall pits on Borneo started in succeeding layers which had already been removed by mining and/or erosion.
As indicated in Figs. 4 and 5, the basal sample is taken from well within the underlying mineral soil, in which the initial mire forest rooted, at a depth of about 40 cm below seam base. The great abundance of small smooth monolete spores (Laevigatosporites hardtii; Fig. 7d,e) and occasional slightly verrucate monolete spores (Verrucatosporites microfavus; Fig. 7f) suggests that a low-growing herbaceous vegetation, in which a variety of polypodiaceous ferns played a dominant role, preceded the initial forest mire vegetation. The scarcity and relatively poor preservation of pollen and the occasional occurrence of moss spores (Corrusporites sp.; Fig. 7a–c) are signs of fairly deep weathering, respectively, nutrient depletion which could have supported an open herbaceous community. Abundant resin particles do not belong to the sediment but have likely been released from root tissue of the succeeding vegetation which was included in the sample.
Similarly, decomposition of roots in the uprooted stump and the side walls of the pit, together with outwash concentration at the base of the pit, may have led to the dominance of resin particles in sample 2 (Fig. 10). This sample is representative for the initial phase of deposition within the pit. Palynologically, this is demonstrated by a sharp decrease of polypodiaceous fern spores and the onset of pollen of a mire forest vegetation, which shows a more or less continuous distribution in overlying samples. The only exception is the unique abundance of Tricolpopollenites liblarensis, which is commonly attributed to a more open landscape (Lenz 2005; Lenz and Riegel 2001; Pflug 1952; Riegel and Wilde 2016), and the high proportion of this pollen may indicate a stage of forest development prior to the closure of the peat-forming mire forest. T. liblarensis is also abundant at the very base of the seam in nearby localities, thus, apparently representing an ecotonal element at the mire margin. At the same level, the typical mire elements of the Paleogene, such as inaperturate pollen of some Cupressaceae s.l. (formerly Taxodiaceae) (Fig. 7g–i), Tricolporopollenites cingulum (Fagaceae), and, in part, Nyssapollenites kruschii (Nyssaceae) (Fig. 8g–i), become important and show a clear trend to dominance towards the top of the section.
The main phase of pit infill (samples 3 to 6) is characterised by the dominance of Triatriopollenites bituitus with frequencies up to 40% of the total assemblage (Fig. 6). This may include a minor proportion of T. rurensis, which is not consistently distinguishable in routine counts (Wilde et al. in press). Both are generally attributed to the Myricaceae. Significantly, Lenz (2005) mentions that T. bituitus more commonly occurs in transitions between lignite seams and siliciclastic interbeds in the middle Eocene of Helmstedt, suggesting a preference of Myricaceae for marginal or initial ecotonal communities of peat-forming environments.
Several rare taxa are restricted to the lower and middle part of the section and mostly attaining less than 1% in the counts. Nervertheless, these taxa are important as climatic and environmental indicators. The morphologically unique Porocolpopollenites vestibulum (Fig. 8d–f), f.i., has been attributed to the mainly tropical family Symplocaceae (Thiele-Pfeiffer 1980; Thomson and Pflug 1953). Although the botanical affinity of Tricolporopollenites abbreviatus (Fig. 8a–c) is still not known, its occurrence in Central Europe is largely restricted to, and typical for, the middle Eocene, which is considered to support the most tropical vegetation in this region (e.g. Krutzsch 1992; Lenz 2005; Pflug 1952; Riegel et al. 2012). Conspicuous by their decorative sculpture are specimens of Reticulataepollis intergranulatus with a potential affinity to Nyctaginaceae (Bougainvillea; Wilde and Schaarschmidt 1993). The similarly ornamented, but distinctly tricolporate, Tricolporopollenites pseudointergranulatus (Fig. 9d–f) has been compared to pollen of Menispermaceae (Thiele-Pfeiffer 1980) or Viburnum (Adoxaceae; Hofmann and Gregor 2018). In addition, the common occurrence of Compositoipollenites rhizophorus, a putative pollen of Icacinaceae, and the occasional appearance of Matanomadhyasulcites sp., affiliated with the Annonaceae (Venkatachala et al. 1988), together with various forms of Tetracolporopollenites (Sapotaceae) (Fig. 8j–l), strongly imply that a (para)tropical forest surrounded the site.
Rare occurrences of Punctilongisulcites microechinatus (Fig. 7j–l) may represent pollen of a species of Hydrocharitaceae as suggested by Thiele-Pfeiffer (1988) and could be evidence that open water occupied the treefall pit at least temporarily. However, some palms have similar pollen (Hofmann et al. 2015). Even the most careful search has not revealed any freshwater plankton such as, e.g. freshwater dinoflagellates of the genus Geiselodinium, which have been described as locally frequent in sinkhole fills in the Geiseltal lignite (Krutzsch 1962). The absence of aquatic elements may possibly be explained by nutrient depleted acidic water in the pit between a highly depleted soil and the initial peat swamp.
The complete absence of charcoal particles in combination with pollen of mostly tropical families, such as Sapotaceae (Tetracolporopollenites spp.) or Symplocaceae (Porocolpopollenites vestibulum) among others, is in line with the previously postulated perhumid paratropical climate of the middle Eocene in Central Europe (Lenz 2005; Riegel et al. 1999; Riegel et al. 2012).
A soil of the kind with the low-diversity Laevigatosporites-dominated assemblage underlying the treefall pit is not included in the undisturbed neighbouring section G VII F (Fig. 6). Instead, the rooted clastic sediment beneath the coal is transitional with the base of the seam and dominated by Tricolpopollenites liblarensis. It is, therefore, conceivable that the singular peak of this species in sample 2 of the treefall pit has been sourced by downwash from this layer, which was merely stripped off by mining at the treefall site together with the contact to the seam. Since there is no coal preserved on top of the treefall pit, it is possible that the very top of the underlying sediments was stripped away here.
It remains conjectural about what proportion of palynological assemblages in the higher samples 3 to 7 of the treefall pit have been influenced by downwash from the base of the seam, but some differences to the undisturbed section are evident. The treefall pit possibly includes a protracted record of the onset of the peat swamp when compared to the undisturbed nearby site. Inaperturate pollen (former Taxodiaceae, now included in Cupressaceae s.l.) and Nyssapollenites are more common in parts of the treefall pit but rare in the undisturbed section. This may indicate that Taxodium and Nyssa may have benefited somewhat from a wetter substrate and more light around the pit. The same may be true for Myricaceae because their pollen is common in the treefall pit but rare in the other locality. These three taxa may be considered as gap-colonising and temporarily replacing the surrounding flora after a treefall (Gastaldo and Staub 1999). On the other hand, palm pollen (Monocolpopollenites tranquillus) is rare in the pit, but slightly more common in the other section, especially in its upper part. There is a striking opposite trend between the two sample sites in different fagaceous pollen (Tricolpopollenites liblarensis and Tricolporopollenites cingulum, respectively), with juglandaceous pollen remaining a common background element without notable vertical trends in both sections. However, it remains unclear if the differences can fully be explained by a local clearing caused by treefall or simply by patchiness of the vegetation in general.
Summary and conclusions
Mining in the former open-cast mine Neumark Nord of the Geiseltal Mining District exposed a planar surface at the immediate base of the middle Eocene Mittelkohle. Among numerous remnants of tree bases, it revealed a large diameter fallen tree with a treefall pit measuring about 4 m in diameter at its base. The filling of the pit represents a high-resolution section equivalent to the initial time of forestation and peat accumulation. Palynology revealed a striking difference between a fern-dominated herbaceous plant community in the sediments underlying the pit, a pulse of washed-in fagaceous pollen (Tricolpopollenites liblarensis) at the base of the pit, followed by an increase of woody taxa pollen typical of a peat swamp forest including a number of tropical elements. Comparison with pollen from a nearby undisturbed transition from the underlying sediment to the seam confirms the ecotonal character of the parent plant of T. liblarensis, and suggests that Cupressaceae s.l., Nyssaceae and Myricaceae may have benefited from moisture and light in the clearing of the treefall pit. It is remarkable that there is no unequivocal evidence of aquatic elements in the fill. The transition from the underlying sediment to the lignite represents a sharp break in the sedimentary regime from clastic to purely organic material with a bounding surface colonised by large trees, mostly preserved as individual “stumps”, but including several fallen logs just at the transition from the underlying sediment to the coal.
Dedication
The paper is dedicated to the memory of the two succeeding curators of the Geiseltal-collections at the Geiseltalmuseum of the Martin Luther Universität Halle-Wittenberg in Halle an der Saale, Günter Krumbiegel (1926-2014) and Meinolf Hellmund (1960-2016). Already retired, Günter Krumbiegel supported our field and research activities in the area of the former Geiseltal Mining District with great interest. He generously supplied his special knowledge and experience. We are indebted to his successor and our friend Meinolf Hellmund who initially inspired, supported, and accompanied our activities in Geiseltal fieldwork and research until his untimely death.
References
Bobrovsky, M. V., & Loyko, S. V. (2016). Patterns of pedoturbation by tree uprooting in forest soils. Russian Journal of Ecosystem Ecology, 1(1). https://doi.org/10.21685/2500-0578-2016-1-3.
Boros, A., & Járai-Komlódi, M. (1975). An Atlas of recent European moss spores. Budapest: Akadémiai Kiadó.
Clinton, B. D., & Baker, C. R. (2000). Catastrophic windthrow in the Appalachians: Characteristics of pits and mounds and initial vegetation responses. Forest Ecology and Management, 126(1), 51–60.
Čuchta, P. (2020). Natural disturbances (with a special reference to windthrow): A literature review. Advances in Environmental Research, 72, 153–172.
Gastaldo, R. A. (1990). The paleobotanical character of log assemblages necessary to differentiate blow-downs resulting from cyclonic winds. Palaios, 5(5), 472–478.
Gastaldo, R. A., & Staub, J. R. (1999). A mechanism to explain the preservation of leaf litter lenses in coals derived from raised mires. Palaeogeography, Palaeoclimatology, Palaeoecology, 149(1-4), 1–14.
Gruas-Cavagnetto, C., & Köhler, E. (1992). Pollens fossiles d’Euphorbiacées de l‘Eocene francais. Grana, 31, 291–304.
Harley, M. M. (1991). The pollen morphology of the Sapotaceae. Kew Βulletin, 46, 379–491.
Harrison-Day, V., & Kirkpatrick, J. (2019). Pits, mounds, and vernal ponds in a Tasmanian subalpine grassland. Geographical Research, 2019, 230–237. https://doi.org/10.1111/1745-5871.12328.
Haubold, H., & Hellmund, M. (1994). Das Geiseltalmuseum am Institut für Geologische Wissenschaften. Veröffentlichungen der Akademischen Sammlungen und Museen der Martin-Luther-Universität Halle-Wittenberg, 1, 11–39.
Haubold, H., & Thomae, M. (1990). Stratigraphische Revision der Wirbeltierfundstellen des Geiseltaleozäns. Hallesches Jahrbuch für Geowissenschaften, 15, 3–20.
Hellmund, M., & Wilde, V. (2001). Das Geiseltal-Projekt 2000 - erste wissenschaftliche Ergebnisse. Hallesches Jahrbuch für Geowissenschaften. Reihe B: Geologie, Paläontologie, Mineralogie Beiheft, 13, 1–99.
Hofmann, C. C., & Gregor, H.-J. (2018). Scanning electron microscope and light microscope investigations of pollen from an atypical mid-Eocene coal facies in Stolzenbach mine (PreußenElektra) near Borken (Kassel, Lower Hesse, Germany). Review of Palaeobotany and Palynology, 252, 41–63.
Hofmann, C. C., Egger, H., & King, C. (2015). LM and SEM investigations of pollen from PETM and EECO localities of Austria and Great Britain: New findings of Atherospermataceae, Annonaceae, Araceae and Arecaceae from the Lower Eocene. Plant Systematics and Evolution, 301, 773–793.
Khanina, L. G., Bobrovsky, M. V., & Zhmaylov, I. V. (2017). Vegetation diversity on the microsites caused by tree uprooting during a catastrophic windthrow in temperate broadleved forests. Russian Journal of Ecosystem Ecology, 4(3). https://doi.org/10.21685/2500-0578-2019-3-1.
Krumbiegel, G. (1959). Die tertiäre Pflanzen- und Tierwelt der Braunkohle des Geiseltales. Wittenberg: A. Ziemsen Verlag.
Krumbiegel, G. (1977). Genese, Palökologie und Biostratigraphie der Fossilfundstellen im Eozän des Geiseltales/1/. Wissenschaftliche Beiträge, Martin-Luther-Universität Halle-Wittenberg, 1977(2)(P5), 113-138.
Krumbiegel, G., Rüffle, L., & Haubold, H. (1983). Das eozäne Geiseltal, ein mitteleuropäisches Braunkohlenvorkommen und seine Pflanzen- und Tierwelt. Wittenberg: A. Ziemsen Verlag.
Krutzsch, W. (1962). Die Mikroflora der Geiseltalbraunkohle. Teil III: Süßwasserdinoflagellaten aus subaquatisch gebildeten Blätterkohlenlagen des mittleren Geiseltales. Hallesches Jahrbuch für Mitteldeutsche Erdgeschichte, 4, 40–45.
Krutzsch, W. (1967). Atlas der mittel- und jungtertiären dispersen Sporen- und Pollen- sowie Mikroplanktonformen des nördlichen Mitteleuropas. Lieferung IV und V: Weitere azonotrilete (apiculate, murornate), zonotrilete, monolete und alete Sporenformen. Jena: VEB Gustav Fischer Verlag.
Krutzsch, W. (1969). Über einige stratigraphisch wichtige neue Longaxoner-Pollen aus dem mitteleuropäischen Alttertiär. Geologie, 18, 472–487.
Krutzsch, W. (1970). Die stratigraphisch verwertbaren Sporen-und Pollenformen des mitteleuropäischen Alttertiärs. Jahrbuch für Geologie, 3, 309–379.
Krutzsch, W. (1976). Die Mikroflora des Geiseltales. Teil IV: die stratigraphische Stellung des Geiseltalprofils im Eozän und die sporenstratigraphische Untergliederung des mittleren Eozäns. Abhandlungen des Zentralen Geologischen Instituts, 26, 47–92.
Krutzsch, W. (1992). Paläobotanische Klimagliederung des Alttertiärs (Mitteleozän bis Oberoligozän) in Mitteldeutschland und das Problem der Verknüpfung mariner und kontinentaler Gliederungen (klassische Biostratigraphien – paläobotanisch-ökologische Klimastratigraphie – Evolutions-Stratigraphie der Vertebraten). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 186(1-2), 137–253.
Küstermann, W., Rappsilber, I., & Thomae, M. (2008). Regionale und lokale tektonische Voraussetzungen zur Bildung der Geiseltal-Senke. Exkursionsführer und Veröffentlichungen der Deutschen Gesellschaft für Geowissenschaften, 235, 12–21.
Langohr, R. (1993). Types of tree windthrow, their impact on the environment and their importance for the understanding of archaeological excavation data. Helinium, 33(1), 36–49.
Lenz, O. K. (2005). Palynologie und Paläoökologie eines Küstenmoores aus dem mittleren Eozän Mitteleuropas - Die Wulfersdorfer Flözgruppe aus dem Tagebau Helmstedt, Niedersachsen. Palaeontographica B, 271(1-6), 1–157.
Lenz, O. K., & Riegel, W. (2001). Isopollen maps as a tool for the reconstruction of a coastal swamp from the Middle Eocene of Helmstedt (Northern Germany). Facies, 45(1), 177–194.
Lutz, H. J. (1940). Disturbance of forest soil resulting from the uprooting of trees. Yale University School of Forestry Bulletin, 45, 1–37.
Maser, C., Cline, S. P., Cromack Jr., K., Trappe, J. M., & Hansen, E. (1988). What we know about large trees that fall to the forest floor. In C. Maser, R. F. Tarrant, J. M. Trappe, & J. F. Franklin (Eds.), From the forest to the sea: a story of fallen trees (pp. 25–45). Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station/U.S. Department of the Interior, Bureau of Land Management.
Muscolo, S. A., Bagnato, S., Sidari, M., & Mercurio, R. (2014). A review of the roles of forest canopy gaps. Journal of Forestry Research, 25(4), 725–736.
Mustoe, G. E. (2018). Non-mineralized fossil wood. Geosciences, 2018(8), 223. https://doi.org/10.3390/geosciences8060223.
Pflug, H. (1952). Palynologie und Stratigraphie der eozänen Braunkohle von Helmstedt. Paläontologische Zeitschrift, 26(1/2), 112–137.
Piirto, D. D., Wilcox, W. W., Parmeter, J. R., & Wood, D. L. (1984). Causes of uprooting and breakage of specimen giant Sequoia trees. University of California Division of Agriculture and Natural Resources Bulletin, 1909, 1–14.
Putz, F. E. (1983). Treefall pits and mounds, buried seeds, and the importance of soil disturbance to pioneer trees on Barro Colorado Island, Panama. Ecology, 64(5), 1069–1074.
Riegel, W., & Wilde, V. (2016). An early Eocene Sphagnum bog at Schöningen, northern Germany. International Journal of Coal Geology, 159, 57–70.
Riegel, W., Bode, T., Hammer, J., Hammer-Schiemann, G., Lenz, O., & Wilde, V. (1999). The paleoecology of the Lower and Middle Eocene at Helmstedt, northern Germany - A study in contrasts. Acta Palaeobotanica Supplement, 2, 349–358.
Riegel, W., Wilde, V., & Lenz, O. K. (2012). The early Eocene of Schöningen (N-Germany) - An interim report. Austrian Journal of Earth Sciences, 105(1), 82–101.
Ring, S. J., Bocherens, H., Wings, O., & Rabi, M. (2020). Divergent mammalian body size in a stable Eocene greenhouse climate. Scientific Reports, 10, 3987. https://doi.org/10.1038/s41598-020-60379-7.
Šamonil, P., Schaetzl, R. J., Valtera, M., Goliáš, V., Baldrian, I., Vašíčková, D., Adam, D., Janik, D., & Hort, L. (2013). Crossdating of disturbances by tree uprooting: Can treethrow microtopography persist for 6000 years? Forest Ecology and Management, 307, 123–135.
Schaetzl, R. J., & Follmer, L. R. (1990). Longevity of treethrow: Implications for mass wasting. Geomorphology, 3(2), 113–123.
Schaetzl, R. J., Burns, S. F., Johnson, D. L., & Small, T. W. (1989). Tree uprooting: Review of impacts on forest ecology. Vegetatio, 79(3), 165–176.
Schupp, E. W., Howe, H. F., & Augspurger, C. K. (1989). Arrival and survival in tropical treefall gaps. Ecology, 70(3), 562–564.
Simoneit, B. R. T., Otto, A., Menor-Salván, C., Oros, D. R., Wilde, V., & Riegel, W. (2020). Composition of resinites from the Eocene Geiseltal brown coal basin, Saxony-Anhalt, Germany, and comparison to their possible botanical analogues. Organic Geochemistry, 152, 104138. https://doi.org/10.1016/j.orggeochem.2020.104138.
Standke, G. (2008). Paläogeografie des älteren Tertiärs (Paleozän bis Untermiozän) im mitteldeutschen Raum. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften, 159(1), 81–103.
Straka, H. (1975). Pollen- und Sporenkunde. Eine Einführung in die Palynologie. Stuttgart: Gustav Fischer Verlag.
Thiele-Pfeiffer, H. (1980). Die miozäne Mikroflora aus dem Braunkohletagebau Oder bei Wackersdorf/Oberpfalz. Palaeontographica B, 174(4-6), 95–224.
Thiele-Pfeiffer, H. (1988). Die Mikroflora aus dem mitteleozänen Ölschiefer von Messel bei Darmstadt. Palaeontographica B, 211(1-3), 1–86.
Thomson, P. W., & Pflug, H. (1953). Pollen und Sporen des mitteleuropäischen Tertiärs. Palaeontographica B, 94(1-4), 1–138.
Ulanova, N. G. (2000). The effect of windthrow on forests on different spatial scales: A review. Forest Ecology and Management, 135(1-3), 155–167.
Urban, B., Sierralta, M., & Frechen, M. (2011). New evidence for vegetation development and timing of Upper Middle Pleistocene interglacials in Northern Germany and tentative correlations. Quaternary International, 241, 125–142.
Van Geel, B. (1978). A palaeoecological study of Holocene peat bog sections in Germany and the Netherlands. Review of Palaeobotany and Palynology, 25, 1–120.
Venkatachala, B. S., Caratini, C., Tissot, C., & Kar, R. K. (1988). Paleocene-Eocene marker pollen from India and tropical Africa. Palaeobotanist, 37(1), 1–25.
Verb, R. G., & Rubino, D. L. (2012). A preliminary survey of diatom taxa from old-growth forest tip-up pools in a southeastern Indiana flatwoods. Proceedings of the Indiana Academy of Sciences, 121(1), 1–7.
Wilde, V., & Hellmund, M. (2006). Neue Geländearbeiten im ehemaligen Braunkohlenrevier Geiseltal. Kooperation zwischen dem Forschungsinstitut Senckenberg in Frankfurt (Main) und dem Geiseltalmuseum am Institut für Geologische Wissenschaften in Halle (Saale). Natur und Museum, 136(7/8), 162–173.
Wilde, V., & Riegel, W. (2010). "Affenhaar" revisited - Facies context of in situ preserved latex from the Middle Eocene of Central Germany. International Journal of Coal Geology, 83(2-3), 182–194.
Wilde, V., & Schaarschmidt, F. (1993). High resolution CLSM-studies of in-situ preserved pollen from the Messel oil shale. Scientific and Technical Information, 10(6), 209–214.
Wilde, V., Frankenhäuser, H., & Lenz, O. K. (in press). A myricaceous male inflorescence with pollen in situ from the middle Eocene of Europe. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-020-00479-4.
Zhang, Y.-L., Wu, P.-C., Wang, M.-Z., Luo, J.-X., & Lin, B.-J. (Eds.) (2005). Spore morphology of Chinese bryophytes. Qingdao: Qingdao Publishing House.
Acknowledgements
Funds from the Lotto Toto GmbH Sachsen-Anhalt and from the Ungerer Foundation of the Senckenberg Forschungsinstitut und Naturmuseum for fieldwork in the Geiseltal area are gratefully acknowledged. Access to the sites in the abandoned open-cast mines was possible due to the friendly permission of the LMBV, the company managing the area at the time of our fieldwork. Fieldwork in the Geiseltal was done in close cooperation with the Geiseltalmuseum of the Martin Luther Universität Halle-Wittenberg in Halle an der Saale (M. Hellmund, Ch. Köhn) and benefited much from the participation of B.R.T. Simoneit and A. Otto (Corvallis, OR, USA). Special thanks are due to Karin Schmidt (Senckenberg Forschungsinstitut und Naturmuseum, Frankfurt am Main) for her constructive and dedicated support in the field. The palynological samples were thoroughly prepared by Simone Huwe. Finally, a language check by Alan Lord (London/Frankfurt) and the helpful comments of Steven R. Manchester, Robert A. Gastaldo and an anonymous reviewer are gratefully acknowledged.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary material
Supplementary Material 1:
Wood from the middle of the log belonging to the treefall pit (SEM, secondary electron mode). a Transverse section showing almost complete destruction of original structures. b Detail from a. c Radial section with faintly visible original alignment of conducting tissue. d Detail from c exceptionally showing some badly preserved pitting. e Transverse section showing longitudinal arrangement of tissues. f Detail from e showing longitudinally arranged fibrous elements with remnants of a ray in between (PDF 785 kb)
Supplementary Material 2
List of taxa encountered during the study (DOCX 21 kb)
Supplementary Material 3
Quantitative data of palynological counts from the treefall pit section G VIII (XLS 32 kb)
Supplementary Material 4
Quantitative data of palynological counts from nearby section G VII F (XLS 35 kb)
Supplementary Material 5
Size distribution of Laevigatosporites in section G VIII, samples 1 and 5 (PDF 1349 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wilde, V., Riegel, W. A middle Eocene treefall pit and its filling: a microenvironmental study from the onset of a forest mire in the Geiseltal (Germany). Palaeobio Palaeoenv 102, 237–251 (2022). https://doi.org/10.1007/s12549-021-00501-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-021-00501-3