
Abstract
The Carnivora (Mammalia) yielded in the coal mine Gračanica in Bosnia and Herzegovina are composed of the caniform families Amphicyonidae (Amphicyon giganteus), Ursidae (Hemicyon goeriachensis, Ursavus brevirhinus) and Mustelidae (indet.) and the feliform family Percrocutidae (Percrocuta miocenica). The site is of middle Miocene age and the biostratigraphical interpretation based on molluscs indicates Langhium, correlating Mammal Zone MN 5. The carnivore faunal assemblage suggests a possible assignement to MN 6 defined by the late occurrence of A. giganteus and the early occurrence of H. goeriachensis and P. miocenica. Despite the scarcity of remains belonging to the order Carnivora, the fossils suggest a diverse fauna including omnivores, mesocarnivores and hypercarnivores of a meat/bone diet as well as Carnivora of small (Mustelidae indet.) to large size (A. giganteus). Faunal similarities can be found with Prebreza (Serbia), Mordoğan, Çandır, Paşalar and Inönü (all Turkey), which are of comparable age. The absence of Felidae is worthy of remark, but could be explained by the general scarcity of carnivoran fossils. Gračanica records the most eastern European occurrence of H. goeriachensis and the first occurrence of A. giganteus outside central Europe except for Namibia (Africa). The Gračanica Carnivora fauna is mostly composed of European elements.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
New material belonging to the order Carnivora from the middle Miocene locality Gračanica (Bugojno Basin, Gornji Vakuf, Bosnia and Herzegovina) is presented herein. Gračanica is of great interest due to its geographic position and its middle Miocene age: carnivoran material of middle Miocene age is rare (e.g. Mayda et al. 2015) and Gračanica is located in the Balkans and in time between well-known other middle Miocene sites such as Sansan in France (Peigné 2012) in the West, Steieregg, Voitsberg and Göriach in Austria (Hofmann 1887, 1892; Thenius 1949a, b), Steinheim and Sandelzhausen in Germany (Heizmann 1973; Nagel et al. 2009) in the Northwest, Prebreza in Serbia (Marković et al. 2012), Mordoğan, Çandır, Paşalar, Inönü, Sinap Formation in Turkey (Kaya et al. 2003; Nagel 2003; Viranta and Andrews 1995; Viranta and Werdelin 2003) in the Southeast, Antonios, Chios Island in Greece in the South (Koufos 2008, 2011) and Belometcheskaya in the Northern Caucasus further East (Pickford et al. 2000). Material from Gračanica thus contributes to the knowledge of middle Miocene Carnivora in Southeastern Europe.
In this article, we discuss the assignment of the carnivoran specimens, the age and palaeoecological inferences based on the carnivoran association.
Locality and stratigraphy
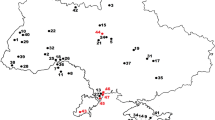
Gračanica is a still active opencast coal mine near the village Gračanica between the cities of Bugojno and Gornji Vakuf in western Bosnia and Herzegovina (Fig. 1). First coal exploitations at Gračanica started in 1939 as underground mining. The locality is situated in the Bugojno Basin, a large intra-mountain basin in the Dinarides, filled with lacustrine deposits (Mandic et al. 2016) (Fig. 1). The sediment exposed in the coal mine represents the lowermost of three lacustrine deposition cycles (Čičić 1976; Mandic et al. in prep., this issue). The lower half of the section in the Gračanica coal mine, of about 20 m thickness, contains the intercalations of lignite deposits, exploited by the mining company. The upper half of the section, also about 20 m thickness, is dominated by clear marly sediment, representing an open lake environment. The mammalian remains come from the lignite rich, lower half of the section, which is indicated by the predominantly dark brownish colour of the fossils. This lignite-bearing part of the section is interpreted as deriving from a swampy environment. The lacustrine deposits overlying the lignite deposits in the Gračanica mine contain a mollusc fauna typical of the early middle Miocene of the Dinaride Lake System (Mandic et al. 2016; Mandic et al. in prep., this issue); this age was confined to the early Langhian, correlating Mammal Zone MN 5 (Harzhauser et al. in press, this issue). The scanty small mammal fauna from the lignite-bearing deposits in the Gračanica section indicates an age ranging between the Mammal Zones MN4 and MN5 (Wessels et al. in press, this issue).

Geographic position of the locality Gračanica in Bosnia-Herzegovina (left) and within the Bugojno Basin (right), modified after Mandic et al. (2012) and Mandic et al. (2016)
Material and methods
All fossils presented in this article are housed in the Natural History Museum Vienna, in Austria (Figs. 2, 3, and 4). Dental measurements (given in mm) were taken with a caliper to the nearest 0.1 mm and are plotted for the carnassials in Fig. 5. Terminology and nomenclature of tooth morphology follow Van Valen (1994) and Smith and Dodson (2003), whereas definitions of dental measurements follow Peigné and Heizmann (2003). We refer herein to the greatest mesiodistal length of the crown simply as length (= L) and to the greatest labiolingual width as width (= W) of the tooth. Measurements given in quotes (“) indicate that the measurement is affected by strong wear or fractures. The abbreviation “frag.” indicates that the specimen is fragmented.

Teeth of Amphicyon giganteus from the middle Miocene of Gračanica, probably of one individual. a, d, g p4 dex. (NHMW 2015/0009/0001a). b, e, h m1 dex. (NHMW 2015/0009/0001b). c, f, i m2 dex. (NHMW 2015/0009/0001c). In lingual view (a–c), occlusal view (d–f) and labial view (g–i)

Teeth of the Ursidae Hemicyon goeriachensis and Ursavus brevirhinus from the middle Miocene of Gračanica. Hemicyon goeriachensis: (a–c) p4 dex. (NHMW 2013/0009/0001a); (d–f) m1 dex. (NHMW 2013/0009/0001b); (g–i) m2 dex. (NHMW 2013/0009/0001d); (j–l) m2 sin. (NHMW 2013/0009/0001c); (m) m3 frag. (NHMW 2013/0009/0001e); (n–p) m1 sin. (NHMW 2013/0009/0002a); (q, r) P4 frag. Sin. (NHMW 2013/0009/0002b); (s–u) P4 dex. (NHMW 2015/0008/0001); (v) M1 frag. sin. (NHMW 2015/0008/0002); (w) M2 sin. (NHMW 2015/0008/0003); (x–z) M2 dex. (NHMW 2015/0008/0004). Ursavus brevirhinus: (za–zc) m2 dex. (NHMW 2015/0010/0001). In occlusal (b, e, h, k, m, o, q, t, v, w, y, za), labial (a, d, g, l, p, s, x, zc) and lingual view (c, f, i, j, n, r, u, z, zb)

Fragmentary mandibles of a Mustelidae indet. and Percrocuta miocenica from the middle Miocene of Gračanica. a Mustelidae indet. mandible sin. with c+p1-3/p2-4? (NHMW 2015/0006/0001) in labial view. b–dPercrocuta miocenica mandible sin. with c+p2-m1 (NHMW 2015/0007/0001) in occlusal (b), labial (c) and lingual (d) view

Comparison of the dental measurements of the carnassials P4/m1. A. giganteus: m1 (diamonds), H. goeriachensis: m1 (circles), P4 (circles with borders), P. miocenica: m1 (squares). Material from Gračanica is in black, whereas comparative material is in grey (for references see Table 1)
Anatomical abbreviations: c, lower canine; P/p, upper/lower premolar, M/m, upper/lower molar
Institutional abbreviations: IPUW, Department for Paleontology, University of Vienna; NHMW, Fossil Vertebrate Collection, Natural History Museum Vienna, Austria
Systematic palaeontology
Order Carnivora Bowdich, 1821
Suborder Caniformia Kretzoi, 1943
Family Amphicyonidae Haeckel, 1866
Genus Amphicyon Blainville, 1841
Amphicyon giganteus Sz, 1825
Material: p4-m2 dex. with p4 slightly damaged (NHMW 2015/0009/0001 a–c); these three lower teeth were found together and most probable derive from the same individual.
Remarks: Amphicyonids radiated from southern North America and represent caniform carnivorans ranging in time from the Middle Eocene to the late Miocene (Tomiya and Tseng 2017). The genus Amphicyon is diverse and known from numerous species from Europe (e.g. Viranta 1996), Asia (e.g. Peigné et al. 2006a), Africa (e.g. Morales et al. 2016) and North America (e.g. Hunt 1998). The stratigraphic range of A. giganteus was assumed for MN 3–MN 5 (Viranta 1996), but shifted recently to MN 4–MN 6 with the youngest record in Europe in MN 6 in Arroyo del Val in Spain (Peigné et al. 2006b). Ginsburg (2000) included earlier forms, identified as A. lathanicus and A. laugnacensis (MN 3), but Peigné et al. (2008) commented on their similarity to A. giganteus. Furthermore, A. giganteus was mentioned from the late Miocene locality Kohfidisch (Austria, MN 11) in De Beaumont (1984), but this material was later identified by Viranta (1996) as A. gutmanni.
Description: The p4 is slightly damaged, preserving the main cuspid and the distal accessory cuspid. The root is broken. The possible presence of an anterior accessory cuspid cannot be assessed. The posterior accessory cuspid is about half as high as the main cusp and is separated by a deep notch. The cingulid is more developed lingually and reaches to the distal part of the p4. The cingulid is crenulated and forms a small cusplet aligned with the main cuspid and the posterior accessory cuspid at the mesial/distal part.
The m1 has an overall robust appearance with two strong roots. The paraconid-protoconid blade shows a shearing facet at the disto-labial side of the paraconid and the mesio-labial side of the protoconid. The protoconid is the highest cuspid of the trigonid. The metaconid is very close to the protoconid, subequal in height with the paraconid and leans slightly distally. Therefore, it is visible also in labial view. The talonid makes up about one third of the tooth. It carries a hypoconid, which is formed into a prominent cutting edge and a small entoconid. A second wear facet is present labially at the hypoconid between the talonid and the protoconid. The largest width can be measured in the centre of the m1 at the level of the protoconid and metaconid.
In occlusal view, the m2 is semicircular-shaped. The protoconid is the highest cuspid. A small notch is visible mesially between the protoconid and the paraconid. The metaconid is well developed, but much lower than the paraconid that approaches the protoconid in height. A low crest extends from the metaconid disto-lingually. The cingulid is most pronounced labially at the talonid. The enamel of p4-m2 is wrinkled where no wear facets are present.
Comparisons: Other contemporaneous genera considered for comparisons, such as Cynelos, Ysengrinia and Haplocyoninae, are at least 20% smaller, with Ysengrinia (m1 L:30) being closest to the m1 studied herein (Peigné and Heizmann 2003).
The genus Amphicyon is defined by the presence of diastemata between the premolars, the single-rooted p2, the enlarged M2 and the presence of an entoconid on m1 (Viranta 1996).
Several diagnostic features for A. giganteus mentioned by Viranta (1996) are verifiable in the present material, such as the presence of a posterior accessory cuspid on p4, the overall stout impression of m1, the small metaconid on m1, the massive talonid with the hypoconid as most prominent cusp and a m2 of subequal height as the talonid on m1. In addition, the lack of a talonid basin confirms the affiliation of the m1 to A. giganteus in contrast to A. major. Comparative material from Chilleurs-aux-Bois (France; MN 3 in Steininger 1999 but without absolute dating) housed in the Museum of Basel shows similar dental measurements (pers. comm. Loic Costeur; see Table 1; included in A. giganteus in Viranta 1996). The Gračanica specimen is of greater size (Fig. 5) than the specimen from Chilleurs-aux-Bois. Measurements given in Morales et al. (2003) for A. giganteus from the middle Miocene of Namibia are also similar to the material described herein. Therefore, we assign the Gračanica specimens to A. giganteus.
Family Ursidae Fischer von Waldheim, 1817
Genus Hemicyon Lartet, 1851
Hemicyon goeriachensis Toula, 1884
Material: (NHMW 2013/0009/0001 a–e) isolated p4 dex. (a), m1 dex. (b), m2 sin. (c) and dex. (d), m3 fragment (e); (NHMW 2013/0009/0002 a–e) isolated m1 sin. (a), P4 fragments sin. (b) and dext. (c), fragmented tooth root (d), fragmented maxillary (e) and bone fragments; (NHMW 2015/0008/0001-0004) isolated P4 dex. (0001), M1 sin. frag. (0002), M2 sin. (0003), M2 dex. (0004).
Remarks: The majority of the material could potentially belong to one individual with p4, m1-2 (NHMW 2013/0009/0001) and M1-2 (NHMW 2015/0008/00001-0004), showing a similar degree of wear. However, as the tooth position of P4 dex. is preserved two times, these remains represent at least two individuals.
The taxonomic affiliations of Hemicyon and its species were subject to revisions and are still discussed (see Ginsburg and Morales 1998; Nagel 2003; or Thenius 1979 for an overview). The genus Hemicyon includes besides others the European species H. gargan, H. stehlini, H. majorali, H. sansaniensis and H. goeriachensis and material of those taxa was included in different species over time (Ginsburg and Morales 1998). H. goeriachensis is known so far from the Mammal Zones MN 5–MN 8 and shows a hypocarnivore tendency, whereas H. sansaniensis is characterised by a hypercarnivore tendency (Ginsburg and Morales 1998).
Description: The mandibular material consists of p4 dex., m1 dex., m2 dex. and sin., m3 fragment (NHMW 2013/0009/0001 a–e), and m1 sin. (NHMW 2013/0009/0002 a). The morphology of the representatives of the same tooth position does not vary and the description is based on both teeth where applicable.
The p4 possessed two roots, but its mesial part is fragmented. The main cuspid is situated in the centre of the tooth, no accessory cuspids are present. A small crest runs across the mesial and distal margin of the main cuspid from the tip to the base. A lingual cingulid is present (Fig. 3(a–c)).
The m1 shows a slight wear facet on its paraconid-protoconid cutting blade. The protoconid is the highest cuspid. Paraconid and metaconid are subequal in height. The talonid bears a pronounced hypoconid, a small entoconid and entoconulid. The talonid shows a shallow basin (Fig. 3(d–f)).
The m2 trigonid consists of a small paraconid and a subequal metaconid and paraconid, which are connected by a crest. The metaconid is slightly lower than the paraconid. The talonid is elongated and a hypoconid is developed (Fig. 3(g–l)).
The m3 is only a small fragment showing a crest around the occlusal surface and another short crest running in the centre of the tooth crown (Fig. 3(m)).
The maxillary material consists of P4 dex., M1 sin. frag., M2 sin. and dex. (NHMW 2015/0008/0001-0004) and P4 sin. frag. and P4 dex. frag. (NHMW 2013/0009/0002 b + c). The morphology of the tooth positions is the same if not indicated differently.
The P4 is represented by a right complete one (2015/0008/0001) and two mesial halves of P4 sin. and dex (NHMW 2013/0009/0002 b + c); in the latter one, the protocone is broken (Fig. 3(q–u)). The P4 fragments show no wear facets. The complete P4 has the same degree of wear as seen in the mandibular material. Therefore, it might be possible that the latter two belong to one individual. The tip of the paracone is broken. Paracone and metastyle are subequal in height and length. A notch is present at the centre of the carnassial blade. There is no parastyle. The protocone is semicircular and there is a mesial indentation that gives it a detached appearance. The protocone is more posteriorly situated. A strong cingulum is present on P4 with the lingual cingulum being more pronounced.
The M1 is fragmented labio-distally and preserves only one root (Fig. 3(v, w)). A pronounced paracone is present (the metacone is broken). The M1 is V-shaped in occlusal view. The protocone is small, mesially shifted and accompanied by a cingulum. There is no paraconule or metaconule, instead a smooth crest is developed in their position.
The M2 is similar to the M1 and differs only in being slightly smaller, more rectangular in occlusal view (not V-shaped), the labial and the lingual borders are curved to each other (bowl-shaped) and the metacone is less developed than the paracone (Fig. 3(w)). The last feature cannot be directly compared to the fragmented M1, but is known from Thenius (1949b).
Comparisons: Phoberocyoninae including Plithocyon show a hypercarnivore tendency and morphological differences to Hemicyon (Ginsburg and Morales 1998), including a parastyle on P4, more developed entoconid and entoconulid on m1 and sharper cusps on m2. Zaragocyon included in Hemicyoninae differs from Hemicyon by a more rectangular M2 and smaller labial cusp on the lower molars (Ginsburg and Morales 1998). Dinocyon (Hemicyoninae) is characterised by an absent or weak parastyle on P4, a much shorter talonid on the lower molars and a higher hypoconid on m1 if compared to Hemicyon (Ginsburg and Morales 1998).
Within the genus Hemicyon, several species can be excluded for the present material from Gračanica: H. gargan due to its much smaller size, H. stehlini due to its slightly smaller size, its more mesially situated protocone on P4 and its m1 and m2 being of the same size (Ginsburg and Morales 1998) and H. mayorali due to its mesio-lingual crest on the paracone of P4, a more rectangular M2 and a more developed entoconid on m2 (Astibia et al. 2000).
H. goeriachensis is similar to H. sansaniensis especially in size (see Peigné 2012; Peigné et al. 2006b; Thenius 1949b; Zapfe 1950 for measurements of H. sansaniensis), but the former is characterised by the absence of paraconule and metaconule in the upper molars and by a weaker cingulum and thus more elongated appearance as well as by the absence of the parastyle on P4 (Nagel 2003). After Ginsburg and Morales (1998), H. goeriachensis further differs from H. sansaniensis as follows: paracone and metacone on M1–2 are lower, the lower premolars are lower, mesio-labially not concave, but mesio-labially narrower and concave disto-lingually, m1 with entoconid and entoconulid and a slightly higher hypoconid, m2 much lower and with a lower protoconid than metaconid. The M1 in H. sansaniensis shows a U-shaped M1, whereas the M1 is V-shaped in H. goeriachensis. Peigné et al. (2006b) added that H. goeriachensis possesses a less developed entoconid and entoconulid on m1 and a lower metaconid than protoconid on m2. The metrics of the specimens described herein match the measurements of the species’ material in Ginsburg and Morales (1998) and in Heizmann (1973). The carnassial data plot close to comparative material of H. goeriachensis (Fig. 5).
Genus Ursavus Schlosser, 1899
Ursavus brevirhinus Schlosser, 1899
Material: m2 dex. (NHMW 2015/0010/0001)
Remarks: The genus Ursavus comprises a couple of different species and its affiliations are discussed since a long time (e.g. Thenius 1979). Some species are meanwhile excluded (Abella et al. 2011; Qiu et al. 2014) such as U. elemenis and U. depereti (Kretzoi 1942; Ginsburg and Morales 1998) with the former one considered as Ballusia and the latter one considered as Agriarctos, although not all material of U. depereti is now referred to A. depereti (Kretzoi 1942). Ginsburg and Morales (1998) consider Ballusia as the earliest member of Ursinae which gave rise to the lineage of Ursavus and Ursus.
Currently, five species from Europe (U. isorei, U. brevirhinus, U. intermedius, U. primaevus, U. ehrenbergi), two species from Asia (U. sylvestris, U. tedfordi) and one species from North America (U. pawniensis) are considered as belonging to the genus of Ursavus (Qiu et al. 2014). The species U. brevirhinus is known from MN 4 to MN 9 (Qiu et al. 2014) and well known from the Austrian localities Steieregg and Voitsberg (MN 5) and Göriach (MN 5; Hofmann 1887, 1892; Thenius 1949a, b).
Description: The m2 is completely preserved with fragmented roots (Fig. 3(za–zc)). The trigonid consists of the protoconid and metaconid. The metaconid is stronger developed than the protoconid. Both are connected by a ridge. The mesial part of the trigonid is semi-circular (“shoe”-shaped) and about half the length of the distal part of the tooth. The talonid is characterised by a labio-lingual narrowing and a labial constriction. The hypoconid is situated distal to this constriction and developed as a low cuspid. A small entoconid is developed as well. The enamel of the occlusal surface is wrinkled.
Comparisons:Pseudarctos differs from the Ursavus specimen from Gračanica because its m2 is more elongated and misses the “shoe”-shaped trigonid besides its (Pseudarctos) smaller size (e.g. Thenius 1949b). The m2 of Adelpharctos shows a shorter trigonid (de Bonis 2011). Indarctos is larger and preserves two lingual cupids on the talonid (Koufos 2011; Peigné 2016).
The m2 of Ursavus brevirhinus was described by Schlosser (1899) as a molar with a low protoconid, a strong metaconid and no paraconid, a state preserved also in the specimen described herein. The morphology of the specimen also fits the description given by Thenius (1949b) for U. brevirhinus. The Gračanica specimen presented here (NHMW 2015/0010/0001) differs from those descriptions in a more structured talonid, preserving more cusps than the hypoconid. A similar talonid condition is described for U. elemensis, which is of smaller size, though (Dehm 1950). We owe this to the intraspecific morphological variation noted by Thenius (1949b) for U. brevirhinus. The systematic identification of the m2 further is supported by the tooth dimensions, which fit the published measurements for that species (Heizmann 1973; Qiu et al. 2014; Thenius 1949b; Zapfe 1950).
Family Mustelidae Swainson, 1835
Mustelidae indet.
Material: mandible sin. with c, p1-3/p2-4? partly covered in surrounding material (NHMW 2015/0006/0001).
Remarks: A more specific taxonomic assignment is omitted due to the fragmentary nature of the specimen.
Description: The mandible fragment preserves the canine and the first three premolars (Fig. 4a). The canine is about twice the height of the first premolar (p1/2?). There is a strong labial ridge that runs from the base of the tooth until the tip of the canine.
The most mesial premolar (p1/2?) possesses two roots and is a simple tooth consisting of a main cuspid and a cutting edge running mesially to the tip and distally from the tip. The main cuspid is shifted mesially. A labial cingulid is present. The canine and the first premolar are tilted mesially, whereas the following two premolars are tilted distally forming a right angle between cutting edges of the first and the second premolar position from a labial view.
The following premolar (p2/3?) is similar to the preceding tooth, but is larger and preserves a main cuspid situated in the middle of the tooth crown. A cutting edge is present as well. A faint lingual cingulid is developed in addition to the labial cingulid. The latter one is more pronounced.
The most distal preserved premolar (p3/4?) is significantly larger than the preceding tooth position, but of similar morphology. The cingulid is more pronounced than in the preceding premolars. No accessory cuspids are present.
There are no diastemata between the preserved teeth.
Comparisons: The missing carnassial and the combination of absent diastemata and accessory cuspids on the premolars complicate the affiliation of the specimen. In general, the Gračanica specimen resembles most the early “otter-like” genera (Grohé et al. 2010) and we discuss here rejected and possible affiliations.
The following taxa of Mustelidae can be excluded due to the following differences:
Martes (e.g. muncki/sansaniensis) shows accessory cuspids on its premolars (Ginsburg 1961; Ginsburg and Bulot 1982; Peigné 2012). Plesictis is different in more spaced premolars and accessory cuspids on the premolars (Dehm 1950). Ischyrictis zibethoides is larger and its premolars are more spaced (Ginsburg 1961; Peigné 2012). Proputorius possesses diastemata and is smaller (Ginsburg 1961; Peigné 2012). Trocharion albanense is of comparable size and shows a pronounced cingulid on its premolars, but possesses only one alveolus for the p1, a parastyle appears in outlines on p3 and is present on p4 (Zapfe 1950). Robles et al. (2010) describes the lower premolars as unicuspid and increasing in size from mesial to distal and note that the retained single-rooted p1 is a plesiomorph character. Taxodon (e.g. sansaniensis) shows wider premolars respective to their length and their height is not much increasing distally (Ginsburg 1961).
Possible affiliations within Mustelidae: Mustelidae of similar dental morphology and size are represented in the middle Miocene in Europe (Lartetictis dubia, Trochictis, Adroverictis), in North America (Mionictis) and in Southeastern Asia (Siamogale thailandica; Grohé et al. 2010). This refers especially to the former European “Mionictis” species, which are now assigned to Lartetictis, Trochictis and Adroverictis (Ginsburg and Morales 1996) and make them possible candidates:
Taxonomic position and affiliation of some specimens still await revision. This prevents a determination of the present fragmentary mandible, but a short overview shall be given here with the limitation that M1 is not preserved, but the most important element to distinguish the taxa discussed in the following: Thenius’ (1949b) “Mionictis dubia”, Lartetictis dubia in Ginsburg and Morales (1996), shows similarities to the specimen described herein. Its affiliation has still to be discussed (pers. obsv. DN). Lartetictis itself is a monospecific genus and was a widespread mustelid genus in the middle Miocene of Europe, but its taxonomical position within Mustelidae is debated (Heizmann and Morlo 1998; Salesa et al. 2013). The separation in size is figured for M1 in Ginsburg and Morales (1996), but no data for premolars is given. “Mionictis ginsburgi” (Alcalá et al. 1994), now assigned to Adroverictis (Ginsburg and Morales 1996), is most characteristic for its M1.
“Mionictis artenensis” (Ginsburg 1968), now Trochictis artenensis (Ginsburg and Morales 1996), is of the same size like the Gračanica mustelid and shows as well no diastemata, cingulids on the premolars and an asymmetrical p2-3 with a symmetrical p4. In addition, the right angle formed in labial view by the cutting edges of the first two premolars figured in Ginsburg (1968) should be noted. The metrics of a Trochictis artenensis specimen from Artenay given in Grohé et al. (2010; herein Table 1) are closest to the discussed specimen. It should be noted that Trochictis depereti is also of the same size, but is described as having more spaced premolars, a single alveolus for p1 and only a faintly developed cingulid in the distal part of the teeth (Pilgrim 1932; Thenius 1949b).
Another Miocene mustelid taxon from Europe (e.g. Steinheim in Germany (Helbing 1936), La-Grive-Saint-Alban in France (Ginsburg 1968) or Gargano in Italy (Villier et al. 2011)) worthy to be discussed is Paralutra. It differs from the Gračanica mustelid in the notably shifted position of the mesial alveolus of the first premolar (p2) and in more pronounced cingulids (Helbing 1936).
Due to the fragmentary nature of the mandible, we assign NHMW 2015/0006/0001 to Mustelidae indet. and only cautiously propose the possibility of the affiliation or proximity to these “otter-like” mustelids sensu Grohé et al. (2010).
Suborder Feliformia Kretzoi, 1945
Family Percrocutidae Werdelin and Solounias, 1991
Genus Percrocuta Kretzoi, 1938
Percrocuta miocenica Pavlovic and Thenius, 1965
Material: mandible sin. with c, p2-m1 with m1 strongly worn and canine fragmented (NHMW 2015/0007/0001).
Remarks: The genus Percrocuta was referred to Hyaenidae in former times until it was placed in its own family, the Percrocutidae (Chen and Schmidt-Kittler 1983; Werdelin and Solounias 1991). A short overview of the genus Percrocuta and synonyms can be found in Morales and Pickford (2006) and a more detailed review in Koufos (2008). The species P. miocenica is known earliest in Europe from MN 6 of Prebreza (Serbia; e.g. Stefanović 2004) and in the Yatağan Basin (Turkey) formerly dated as MN 7–8 (Alçiçek 2010), but now dated as MN 6 (Bouchal et al. 2017).
Description: The fragmented mandible holds the lower canine and p2-m1 (Fig. 4b–d). Two mental foramens are present, one below the anterior root of p2 and one below p3. The tip of the canine is broken off. A diastema is present between the c and the p2.
The p2 consists of a small anterior accessory cuspid, the main cuspid and a small posterior accessory cuspid. The distal part of the tooth displays a lingual extension. The main cuspid is strongly worn. The lingual cingulid is weak.
The p3 resembles the preceding tooth. The lingual extension of the distal part of the tooth is more pronounced besides the overall larger size. The main cuspid of the p3 is strongly worn. The posterior accessory cuspid is present as short cutting edge. The lingual cingulid is weak.
The p4 consists of an anterior accessory cuspid, the main cuspid and the posterior accessory cuspid formed to a short cutting blade. The anterior accessory cuspid was higher than the posterior accessory cuspid, because it is preserved despite heavy wear at a subequal height. The main cuspid is strongly worn and shows a pronounced buccal wear facet as well as on the posterior accessory cuspid as well. The lingual cingulid is more developed on the distal part of the tooth.
The m1 is heavily worn. A short talonid with a cutting edge and a lingual cingulid are present.
Comparisons: Metric data of different Percrocuta species show that the specimen fits to the smaller species such as P. miocenica in contrast to a larger species such as P. tungurensis (Ghaffar et al. 2011). The mandible of P. miocenica from Mordoğan (Turkey) described in Kaya et al. (2003) shows the same morphology and metrics although being slightly larger. Direct comparison with the holotype of P. miocenica (Pavlovic and Thenius 1965; IPUW 4031) shows a match with the present specimen with the only difference that NHMW 2015/0007/0001 belonged to an older individual and shows significant tooth wear which is reflected also in Fig. 5 in a similar length, but a lower width of the carnassial.
Discussion
The Carnivora remains from Gračanica are sparse and palaeoecological interpretations are thus limited. However, the known fossils indicate a diverse carnivoran guild composed of omnivores (U. brevirhinus; Viranta 1996), mesocarnivores (H. goeriachensis; Viranta 1996) and hypercarnivores of a meat/bone diet (P. miocenica; Viranta and Andrews 1995; A. giganteus; Viranta 1996). The taxa unraveled in Gračanica show a range of estimated body masses from 72 kg (P. miocenica; Viranta and Andrews 1995), 80 kg (U. brevirhinus; Viranta 1996), 120 kg (H. goeriachensis; Viranta 1996) up to 317 kg (for a male A. giganteus; 157 kg for a female; Viranta 1996). Small carnivores were present as well, although the scarcity of their remains does not allow further evaluations (Mustelidae indet.).
The absence of Felidae is noteworthy when compared to other (middle) Miocene sites. The felid Pseudaelurus is often part of the Miocene carnivoran fauna for example in La Romieu (Roman and Viret 1934) and La Grive St. Alban (Gaillard 1899) in France, Paşalar (Mayda et al. 2015; Viranta and Andrews 1995; Bernor and Tobien 1990) and Çandır (Nagel 2003) in Turkey or Belometchetskaya (Pickford et al. 2000) eastwards in the North Caucasus. However, the lack of felids might be attributed to the paucity of the Carnivora remains from Gračanica, since there are also records of felids from Greece during MN 4/5 (Koufos 2008). In addition, there is ongoing discussion on the age determination, e.g. if Çandır and Paşalar represent Mammal Zone MN 5 (Begun et al. 2003; Krijgsman 2003) or 6 (Van der Made 2003, 2005).
The deposits of Gračanica are currently assumed to be of Langhian age, correlating Mammal Zone MN 5. The carnivoran assemblage confirms the middle Miocene age: The latest confirmed occurrence of A. giganteus is known from MN 6 (Peigné et al. 2006b). The earliest occurrence of H. goeriachensis is based on the stratigraphical assignment of Göriach in late MN 5. While Percrocuta sp. and Percrocutidae were described for MN 4/5 from Greece before (Koufos 2008), P. miocenica from Prebreza is only known from MN 6 (Ginsburg and Morales 1998; Stefanović 2004). Thus, MN 6 would be the age estimation for Gračanica based on Carnivora remains, but would not contradict an age estimation of MN 5.
The following fossil sites, all of MN 6 age (Kaya et al. 2003), are considered to belong to one bioprovince and situated southeastern to Gračanica (Bosnia-Herzegovina): Prebreza in Serbia (Marković et al. 2012); Mordoğan (Kaya et al. 2003), Çandır (Nagel 2003), Paşalar (Viranta and Andrews 1995) and Inönü (Kaya et al. 2003) in Turkey; Belometcheskaya further East in the Northern Caucasus (Pickford et al. 2000). Gračanica (Bosnia-Herzegovina) shares with those sites the occurrence of Amphicyon, Hemicyon, Ursavus and Percrocuta miocenica, although there are differences on species-level, e.g. A. giganteus instead of A. major or H. goeriachensis instead of H. sansaniensis (see Mayda et al. 2015).
More differences become evident when the fauna of Gračanica is compared with Sandelzhausen (Germany; Nagel et al. 2009), a more northern middle Miocene site (MN 5): felids (Pseudaelurus), barbourofelids (Prosansanosmilus) and viverrids (Leptoplesictis) are known from there with percrocutids being absent.
The determined taxa of Gračanica allow certain geographic inferences: A. giganteus occurs in vastly separated regions of Europe (e.g. France) and in Africa (Namibia; Morales et al. 2003). To our knowledge, this is the first evidence of this taxon on a route between central Europe and Africa. The species H. goeriachensis was widespread in Europe from Spain, over France, Germany and Austria into the Czech Republic (Ginsburg and Morales 1998). The remains from Gračanica are to our knowledge the hitherto most eastern occurrence of the species. U. brevirhinus is a central European species (Qiu et al. 2014) and occurs around the same time quite close in Austrian sites (Hofmann 1887, 1892; Thenius 1949a, b). P. miocenica is known from Europe until Turkey (Alçiçek 2010; Stefanović 2004). Gračanica is the northernmost occurrence for P. miocenica and the southernmost for H. goeriachensis and U. brevirhinus so far. Similarities with taxa from Asia (e.g. Turkey) concern only the generic level. The enigmatic A. giganteus has only be described from Namibia so far (Morales et al. 2003), but remains are always fragmentary. So, the occurrence of a very large amphicyonid is confirmed, but more information about migration routes cannot be gained from that. Concluding, the central European influence is strong in the observed carnivore fauna of Gračanica with few elements from the South.
References
Abella, J., Montoya, P., & Morales, J. (2011). A new species of Agriarctos (Ailuropodinae, Ursidae, Carnivora) in the locality of Nombrevilla 2 (Zaragoza, Spain). Estudios Geológicos, 67(2), 187–191.
Alcalá, L., Montoya, P., & Morales, J. (1994). New large mustelids from the Late Miocene of the Teruel Basin (Spain). Comptes Rendus de Academie des Sciences Paris, série, 2(319), 1093–1100.
Alçiçek, H. (2010). Stratigraphic correlation of the Neogene basins in southwestern Anatolia: regional palaeogeographical, palaeoclimatic and tectonic implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 291, 297–318.
Astibia, H., Morales, J., & Ginsburg, L. (2000). Hemicyon mayorali nov. sp., Ursidae du Miocène moyen de Tarazona de Aragon (bassin de l’Ebre, Aragon, Espagne). Annales de Paléontologie, 86(1), 69–79.
Beaumont, G. De (1984). Des dents d’Amphicyon (Mammifère, Carnivore, Ursidé) du Turolien basal de Kohfidisch, Burgenland, Autriche. Archives des Sciences de Genève, 37, 77–83.
Begun, D., Geraads, D., & Gulec, E. (2003). The Candır hominoid locality: implications for the timing and pattern of hominoid dispersal events. Courier Forschungsinstitut Senckenberg, 240, 251–165.
Bernor, R. L., & Tobien, H. (1990). The mammalian geochronology and biogeography of Paşalar (middle Miocene, Turkey). Journal of Human Evolution, 19, 551–568.
Blainville, H. de (1841). Ostéographie et description iconographique des Mammifères récents et fossiles (Carnivores), Vols. 1 and 2 Paris.
Bonis, L. de (2011). A new species of Adelpharctos (Mammalia, Carnivora, Ursidae) from the late Oligocene of the “Phosphorites du Quercy” (France). Esutdios Geológicos, 67(2), 179–186.
Bowdich, T. E. (1821). An analysis of the natural classifications of Mammalia, for the use of students and travellers. J. Smith, pp. 115. Paris.
Bouchal, J. M., Mayda, S., Zetter, R., Grimsson, F., Akgün, F., & Denk, T. (2017). Miocene palynofloras of the Tınaz lignite mine, Muğla, Southwest Anatolia: taxonomy, palaeoecology and local vegetation change. Review of Palaeobotany and Palynology, 243, 1–36.
Chen, G., & Schmidt-Kittler, N. (1983). The deciduous dentition of Percrocuta Kretzoi and the diphyletic origin of hyaenas (Carnivora, Mammalia). Paläontologische Zeitschrift, 57, 159–169.
Čičić, S. (1976). Bugojanski basen. In R. Milojević (Ed.). Mineralne sirovina Bosne i Hercegovine. Knjiga I. Ležišta uglja. Geoinžinjering, Sarajevo, pp. 151–158.
Dehm, R. (1950). Die Raubtiere aus dem Mittel-Miocän (Burdigalium) von Wintershof-West bei Eichstätt in Bayern. Abhandlungen der Bayerischen Akademie der Wissenschaften. Mathematisch-naturwissenschaftliche Klasse, 58, 1–141.
Fischer von Waldheim, G. (1817). Adversaria zoologica. Mémories de la Societé Imperiale des Naturalistes des Moscou, 5, 368–428.
Fraas, O. (1885). Beiträge zur Fauna von Steinheim. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg, 41, 313–326.
Gaillard, C. (1899). Mammifères miocènes nouveaux ou peu connus de La Grive-St.-Alban. Archives du Museum d’Histoire naturelle Lyon, 7(2), 1–79.
Ghaffar, A., Akhtar, M., Butt, M. J., Khan, M. A., & Ikram, T. (2011). Late Miocene hyaenids from the Middle Siwaliks of Pakistan. Pakistan Journal of Zoology, 43(5), 855–862.
Ginsburg, L. (1961). La faune carnivores miocènes de Sansan. Mémoires du Muséum national d’Histoire Naturelle. Série C, Sciences de la terre, 9, 1–190.
Ginsburg, L. (1968). Les Mustélidés piscivores du Mioène francais. Bulletin du Muséum national d’Histoire naturelle, Paris, 2e série, 40(1), 228–238.
Ginsburg, L. (2000). Les especes du genre Amphicyon et un Amphicyonidae Mammalia, Carnivora nouveau des faluns miocenes de l’Anjou. Symbioses, 3, 35–40.
Ginsburg, L., & Bulot, C. (1982). Les carnivores du Miocène de Bézian près de la Romieu (Gers, France). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Series B Physical Sciences, 85(1), 53–76.
Ginsburg, L. & Morales, J. (1996). Lartetictis et Adroverictis, nouveaux genres de Melinae (Mustelidae, Carnivora, Mammalia) du Miocène de l’Ancien Monde. Bulletin du Muséum national d’Histoire naturelle, Paris, 4e série, 18, 663–671.
Ginsburg, L., & Morales, J. (1998). Les Hemicyoninae (Ursidae, Carnivora, Mammalia) et les forms apparentées du Miocène inférieur et moyen d’Europe occidentale. Annales de Paléontologie, 84(1), 71–123.
Grohé, C., Chaimanee, Y., de Bonis, L., Yamee, C., Blondel, C., & Jaeger, J.-J. (2010). New data on Mustelidae (Carnivora) from Southeast Asia: Siamogale thailandica, a peculiar otter-like mustelid from the late middle Miocene Mae Moh Basin, northern Thailand. Naturwissenschaften, 97(11), 1003–1015.
Haeckel, E. (1866). Generelle Morphologie der Organismen - Volume 2. Georg Reimer, pp. 462. In Berlin.
Harzhauser, M., Mandic, O., Nordsieck, H. & Neubauer, T.A. (in press). A new Helicidae (Gastropoda) from the middle Miocene of Bosnia and Herzegovina, with a revision of the genus Paradrobacia. In U.B. Göhlich & O. Mandic (Eds.) The drowning swamp of Gračanica (Bosnia-Herzegovina)—a diversity hotspot from the middle Miocene in the Bugojno Basin. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-018-0344-1 [this issue].
Heizmann, E. (1973). Die Carnivoren des Steinheimer Beckens. Palaeontographica, Supplement, 8(5B), 1–95.
Heizmann, E., & Morlo, M. (1998). Die semiaquatische Lartetictis dubia (Mustelinae, Carnivora, Mammalia) vom Goldberg/Ries (Baden-Württemberg). Mainzer naturwissenschaftliches Archiv/Beiheft, 21, 141–153.
Helbing, H. (1936). Die Carnivoren des Steinheimer Beckens. A. Mustelidae. Palaeontographica. Suppl., 8, 1–55.
Hofmann, A. (1887). Über einige Säugetierreste aus der Braunkohle von Voitsberg und Steieregg bei Wies. Jahrbuch der Geologischen Reichsanstalt, 37(2), 207–218.
Hofmann, A. (1892). Beiträge zur miocänen Säugetierfauna der Steiermark. Jahrbuch der Geologischen Reichsanstalt, 42(1), 63–76.
Hunt, R. M. (1998). Amphicyonidae. In Janis, C. M., Scott, K. M, Jacobs, L. L. (eds.): Evolution of tertiary mammals of North America. Vol. 1: Terrestrial carnivores, ungulates, and ungulate-like mammals (pp. 196–277): Cambridge: Cambridge University Press.
Kaya, T., Geraads, D., & Tuna, V. (2003). A new middle Miocene mammalian fauna from Mordoğan (Western Turkey). Paläontologische Zeitschrift, 77(2), 293–302.
Koufos, G. D. (2008). Carnivores from the early/middle Miocene locality of Antonios (Chalkidiki, Macedonia, Greece). Geobios, 41, 365–380.
Koufos, G. D. (2011). The Miocene carnivore assemblage of Greece. Estudios Geológicos, 67(2), 291–320.
Kretzoi, M. (1938). Die Raubtiere von Gombaszog nebst einer Übersicht der Gesamtfauna. Annals Museum Naturalis Hungarici, 31, 89–157.
Kretzoi, M. (1942). Zwei neue Agriotheriiden aus dem ungarischen Pannon. Földtani Közlöny, 72, 350–353.
Kretzoi, M. (1943). Kochictis centennii n. g. n. sp., ein altertümlicher Creodonte aus dem Oberoligozän Siebenbürgens. Földtani Közlöny, 73, 190–195.
Kretzoi, M. (1945). Bemerkungen über das Raubtiersystem. Annals Museum Naturalis Hungarici, 38(4), 59–83.
Krijgsman, W. (2003). Magnetostratigraphic dating of the Candır fossil locality (middle Miocene, Turkey). In E. Gülec, D. Begun, D. Geraads (Eds.), Geology and vertebrate paleontology of the middle Miocene hominoid locality Candır (Central Anatolia, Turkey). Courier Forschungsinstitut Senckenberg, 240, 41–49.
Lartet, E. (1851). Notice sur la colline de Sansan – suivie d’une récapitilation de diverses espèces d’animaux vertébrés fossils trouvés soit à Sansan, soit dans d’autres gisements du terrain tertiaire miocène dans le basin sous-pyrénéen. 45 pp, J.-A. Portes.
Made, J. van der (2003). Suoidea (pigs) from the Miocene hominoid locality Candır in Turkey. In E. Gülec, D. Begun, D. Geraads (Eds.), Geology and vertebrate paleontology of the middle Miocene hominoid locality Candır (Central Anatolia, Turkey). Courier Forschungsinstitut Senckenberg, 240, 149–179.
Made, J. van der (2005). Errate and reply to guest editor’s notes. Courier Forschungsinstitut Senckenberg, 254, 473–477.
Mandic, O., Pavelić, D., Kovačić, M., Sant, K., Andrić, N., & Hrvatović, H. (2016). Field trip guide book. Lake - Basin - Evolution, RCMNS Interim Colloquium 2016 & Croatian Geological Society Limnogeology Workshop, 19-24 May 2016, Zagreb, Croatia. Hrvatsko geološko društvo / Croatian Geological Society. 80 pp. ISBN 978-953-59036-0-4.
Marković, Z., Alaburić, S., Milivojević, M., & Lović, A. (2012). Geoconservation of the paleontological site “Prebreza”. Bulletin of the Natural History Museum, 5, 7–13.
Mayda, S., Koufos, G. D., Kaya, T., & Gul, A. (2015). New carnivore material from the middle Miocene of Turkey. Implications on biochronology and palaeoecology. Geobios, 48(1), 9–23.
Morales, J., & Pickford, M. (2006). A large Percrocutid Carnivore from the Late Miocene (ca. 10-9 Ma) of Nakali, Kenya. Annales de Paléontologie, 92, 359–366.
Morales, J., Pickford, M., Fraile, S., Salesa, M. J., & Soria, D. (2003). Creodonta and Carnivora from Arrisdrift, early middle Miocene of Southern Namibia. Memoirs of the Geological Survey of Namibia, 19, 177–194.
Morales, J., Pickford, M., & Valenciano, A. (2016). Systematics of African Amphicyonidae, with descriptions of new material from Napak (Uganda) and Grillental (Namibia). Journal of Iberian Geology, 42(2), 131–150.
Nagel, D. (2003). Carnivora from the middle Miocene hominoid locality of Candır. (Turkey). In E. Gülec, D. Begun, D. Geraads (Eds.), Geology and vertebrate palaeontology of the middle Miocene hominoid locality Candır (Central Anatolia, Turkey). Courier Forschungsinstitut Senckenberg, 240, 113–131.
Nagel, D., Stefen, C., & Morlo, M. (2009). The carnivoran community from the Miocene of Sandelzhausen (Germany). Paläontologische Zeitschrift, 83, 151–174.
Pavlovic, M. & Thenius, E. (1965). Eine neue Hyäne (Carnivora), Mammalia) aus dem Miozän Jugoslawiens und ihre phlyogenetische Stellung. Anzeiger der Österreichischen Akademie der Wissenschaften, Mathematisch-naturwissenschaftliche Klasse, 102(2), 177–185.
Peigné, S. (2012). Les Carnivora de Sansan. In S. Peigné & S. Sen (Eds.), Mammifères de Sansan. Mémoires du Muséum national d’Histoire naturelle. Paris, 203, 559–660.
Peigné, S. (2016). Carnivora. In S. Sen (Ed.), Late Miocene mammal locality of Küçükçekmece, European Turkey. Geodiversitas, 38(2), 197–224.
Peigné, S., & Heizmann, E. P. J. (2003). The Amphicyonidae (Mammalia, Carnivora) from Ulm-Westtangente (MN 2, Early Miocene), Baden-Württemberg, Germany—systematics and ecomorphology. Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie), 343, 1–133.
Peigné, S., Chaimanee, Y., Yamee, C., Pannipa, T., & Jaeger, J.-J. (2006a). A new amphicyonid (Mammalia, Carnivora, Amphicyonidae) from the late middle Miocene of northern Thailand and a review of the amphicyonine record in Asia. Journal of Asian Earth Sciences, 26(5), 519–532.
Peigné, S., Salesa, M. J., Antón, M., & Morales, J. (2006b). New data on carnivores from the middle Miocene (Upper Aragonian, MN 6) of Arroyo del Val area (Villafeliche, Zaragoza Province, Spain). Estudios Geológicos, 62(1), 359–374.
Peigné, S., Salesa, M. J., Antón, M., & Morales, J. (2008). A new Amphicyonine (Carnivora: Amphicyonidae) from the Upper Miocene of Batallones-1, Madrid, Spain. Paleontology, 51(4), 943–965.
Pickford, M., Gabunia, L., Mein, P., Morales, J., & Azanza, B. (2000). The middle Miocene mammalian site of Belometchetskaya, North Caucasus: an important biostratigraphic link between Europe and China. Geobios, 33(2), 257–267.
Pilgrim, G. E. (1932). The genera Trochictis, Enhydrictis, and Trocharion, with remarks on the taxonomy of the Mustelidae. Journal of Zoology, 102(4), 845–867.
Qiu, Z.-X., Deng, T., & Wang, B.-Y. (2014). A Late Miocene Ursavus skull from Guanghe, Gansu, China. Vertebrata PalAsiatica, 52(3), 265–302.
Robles, J. M., Alba, D. M., Moyà-Solà, S., Casanovas-Vilar, I., Galindo, J., Rotgers, C., Almécija, S., & Carmona, R. (2010). New craniodental remains of Trocharion albanense Major, 1903 (Carnivora, Mustelidae), from the Vallès-Penedès Basin (Middle to Late Miocene, Barcelona, Spain). Journal of Vertebrate Paleontology, 30(2), 547–562.
Roman, F., & Viret, J. (1934). La faune de mammifères du Burdigalien de la Romieu (Gers). Mémoires de la Société Géologique de France, nouvelle série, Paris, 21, 1–67.
Salesa, M. J., Antón, M., Siliceo, G., Pesquero, M. D., Morales, J., & Alcalá, L. (2013). A non-aquatic otter (Mammalia, Carnivora, Mustelidae) from the Late Miocene (Vallesian, MN 10) of La Roma 2 (Alfambra, Teruel, Spain): systematics and functional anatomy. Zoological Journal of the Linnean Society, 169, 448–482.
Schinz, R. H. (1825). Cuviers Thierreich. IV.
Schlosser, M. (1899). Über die Bären und bärenähnlichen Formen des europäischen Tertiärs. Palaeontographica, 46, 95–151.
Smith, J. B., & Dodson, P. (2003). A proposal for a standard terminology of anatomical notation and orientation in fossil vertebrate dentitions. Journal of Vertebrate Paleontology, 23(1), 1–12.
Steininger, F. F. (1999). Chronostratigraphy, geochronology and biochronology of the Miocene “European Land Mammal Mega-Zones” (ELMNZ) and the Miocene “Mammal-Zones (MN-Zones)”. In G. E. Rössner & K. Heissig (Eds.), The Miocene land mammals of Europe (pp. 9–24). München: Verlag Dr. Friedrich Pfeil.
Stefanović, I. (2004). The fauna of Prebreza (southern Serbia) and its position within the mammalian Neogene units. Annales Géologiques de la Péninsule Balkanique, 65, 77–84.
Swainson, W. (1835). On the natural history and classification of quadrupeds. In D. Lardner (Ed.), The cabinet cyclopaedia (pp. 397). London: Longman.
Thenius, E. (1949a). Die tortone Säugetierfauna von Neudorf an der March (ČSR) und ihre Bedeutung für die Helvet-Torton-Grenze. Anzeiger der Österreichischen Akademie der Wissenschaften. Mathematisch-naturwissenschaftliche Klasse, 7, 160–171.
Thenius, E. (1949b). Die Carnivoren von Göriach (Steiermark). Sitzungsberichte der Österreichischen Akademie der Wissenschaften, Mathematisch-naturwissenschaftliche Klasse, 1, 158(9–10), 695–762.
Thenius, E. (1979). Zur systematischen und phylogenetischen Stellung des Bambusbären: Ailuropoda melanoleuca David (Carnivora, Mammalia). Zeitschrift für Säugetierkunde, 44, 286–305.
Tomiya, S. & Tseng, Z. J. (2017). Whence the beardogs? Reappraisal of the Middle to Late Eocene “Miacis” from Texas, USA, and the origin of Amphicyonidae (Mammalia, Carnivora). Royal Society Open Science, https://doi.org/10.1098/rsos.160518
Toula, F. (1884). Über einige Säugethierreste von Göriach bei Turnau (Bruck a/M. Nord), Steiermark. Jahrbuch der Kaiserlich-königlichen geologischen Reichsanstalt, 34, 385–401.
Van Valen, L. M. (1994). Serial homology: the crests and cusps of mammalian teeth. Acta Palaeontologica Polonica, 38(3/4), 145–158.
Villier, B., Pavia, M., & Rook, L. (2011). New remains of Paralutra garganensis Willemsen, 1983 (Mustelidae, Lutrinae) from the Late Miocene “Terre Rosse” of Gargano (Apulia, Italy). Bolletino della Società Paleontologica Italiana, 50(2), 135–143.
Viranta, S. (1996). European Miocene Amphicyonidae—taxonomy, systematics and ecology. Acta Zoologica Fennica, 204, 1–61.
Viranta, S., & Andrews, P. (1995). Carnivore guild structure in the Paşalar Miocene fauna. Journal of Human Evolution, 28, 359–372.
Viranta, S. & Werdelin, L. (2003). Carnivora. In M. Fortelius, J. Kappelman, S. Sen, & R .L. Bernor (Eds.), Geology and paleontology of the Miocene Sinap Formation, Turkey (pp. 178–193). New York: Columbia University Press.
Werdelin, L., & Solounias, N. (1991). The Hyaenidae: taxonomy, systematics and evolution. Fossils and Strata, 30, 1–104.
Wessels, W., Bruijn, H. de, Marković, Z. & Milivojević, M. (in press). Small mammals from the opencast lignite mine Gračanica (Bugojno, middle Miocene), Bosnia and Herzegovina. In U.B. Göhlich & O. Mandic (Eds .) The drowning swamp of Gračanica (Bosnia-Herzegovina)—a diversity hotspot from the middle Miocene in the Bugojno Basin. Palaeobiodiversity and Palaeoenvironments Doi: https://doi.org/10.1007/s12549-018-0366-8 [this issue].
Zapfe, H. (1950). Die Fauna der miozänen Spaltenfüllung von Neudorf an der March (ČSR). Carnivora. Sitzungsberichte der Österreichischen Akademie der Wissenschaften, Mathematisch-naturwissenschaftliche Klasse, 1(159), 109–141.
Acknowledgements
Open access funding provided by Medical University of Vienna. We thank Serdar Mayda (Ege University, Izmir, Turkey) for fruitful discussions and providing literature as well as information on carnivoran material from other middle Miocene sites (e.g. Paşalar) and for his thorough review of the manuscript. In addition, we thank one anonymous reviewer for improving the manuscript. Many thanks are going to Loic Costeur (Natural History Museum Basel) who provided data and information on comparative material housed at the Natural History Museum in Basel. Lilith-Valerie Svaricek (University Vienna) assisted in picture editing.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the special issue “The drowning swamp of Gračanica (Bosnia-Herzegovina) - a diversity hotspot from the middle Miocene in the Bugojno Basin”
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Bastl, K., Nagel, D., Morlo, M. et al. The Carnivora (Mammalia) from the middle Miocene locality of Gračanica (Bugojno Basin, Gornji Vakuf, Bosnia and Herzegovina). Palaeobio Palaeoenv 100, 307–319 (2020). https://doi.org/10.1007/s12549-018-0353-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-018-0353-0