
Abstract
Three new lower molars of Storchodon cingulatus and a fragmentary upper molar confirm its morganucodontan affinities. Cusps b and c are relatively higher than in Morganucodon, and cusp c is larger than cusp b. Cusp a is asymmetric, with slightly distally recurved tip. All known lower Storchodon molars have a pronounced furcation between the roots on the labial and lingual sides. The wear pattern of Storchodon corresponds to that observed in Morganucodon. Cusp A of the upper molar occludes into the valley between cusps a and c of the lower antagonist, causing large confluent wear facets. The smaller wear facets present on the mesiolabial flank of cusp b and the distolabial flank of cusp c were caused by cusp C of the upper antagonist that occluded into the embrasure between two lower molars. So far, the Morganucodon-like occlusal mode is restricted to the morganucodontans Morganucodon, Dinnetherium nezorum, Cifellilestes ciscoensis, and Storchodon cingulatus, whereas Megazostrodon and Erythrotherium exhibit embrasure occlusion. Although Storchodon belongs to the larger morganucodontans, it is questionable if it represents a case of island gigantism as suggested earlier.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
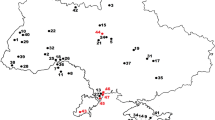
In the Langenberg Quarry (Fig. 1) near Goslar (Lower Saxony) at the northern rim of the Harz Mountains, marginal marine strata of the Süntel Formation of Late Jurassic (late Kimmeridgian) age are exposed. The Süntel Formation comprises impure limestones and marls and is well-dated by marine invertebrates (Lotze 1968; Pape 1970; Fischer 1991; Lallensack et al. 2015). The sediments were deposited in a near shore basin of a carbonate ramp system in the Lower Saxony Basin (Zuo et al. 2018) with influx of fluvial sediments, leading to temporary brackish conditions as indicated by the invertebrates (Lotze 1968; Pape 1970). The vertebrate assemblage (Wings 2015) comprises aquatic as well as terrestrial elements from the hinterland: chondrichthyan and osteichthyan fishes (Mudroch and Thies 1996), marine turtles (Jansen and Klein 2014), a paramacellodid lizard (Richter et al. 2013), the small atoposaurid crocodylomorph Knoetschkesuchus and other crocodyliforms (Thies et al. 1997; Karl et al. 2006; Schwarz et al. 2017), a pterosaur (Fastnacht 2005), the dwarfed sauropod dinosaur Europasaurus (Sander et al. 2006; Carballido and Sander 2014; Marpmann et al. 2015; Carballido et al. 2020), various theropod dinosaurs (Lubbe et al. 2009; Gerke and Wings 2016; Evers and Wings 2020), the morganucodontan mammaliaform Storchodon cingulatus, the docodontan Docodon langenbergensis, three multituberculate mammals of the paulchoffatiid line (Cimbriodon multituberculatus, the pinheirodontid Teutonodon langenbergensis, and Paulchoffatiidae indet.), as well as the dryolestid cladotherian Hercynodon germanicus (Martin et al. 2016, 2019, 2021a, b, 2024).

Location of the Langenberg Quarry (asterisk) at the northern border of the Harz Mountains in north central Germany. From Martin et al. (2021b)
After the description of the holotype (upper molar) of Storchodon cingulatus (Martin et al. 2019), additional molars of a large morganucodontan were recovered from the Langenberg Quarry. Based on morphology, size, and wear pattern these are assigned to Storchodon cingulatus. After Cifellilestes ciscoensis from the Tithonian of the Morrison Formation (Davis et al. 2022), Storchodon cingulatus is the second-youngest morganucodontan from the fossil record. The new sample from the Langenberg Quarry comprises three lower molars (one of them fragmentary), which were unknown so far, and an upper molar fragment which are described and analyzed here.
Material and methods
The Storchodon molars were recovered by screen-washing of about 12 tons of marly limestones, mostly from a dark grey lens within the Süntel Formation. The fossiliferous matrix was collected during several field operations (2012–2022) and brought to the preparation laboratory at the University of Bonn for further processing. In the laboratory, the dried matrix was disaggregated with 15% hydrogen peroxide solution and the tenside Rewoquat® (Evonik Industries AG, Essen, Germany), then subsequently screen-washed at a mesh size of 0.5 mm. The dried concentrate was picked under a stereomicroscope for microvertebrate remains.
The teeth and tooth fragments were scanned at a resolution of 1.66198 µm (NLMH 100023), 1.85986 µm (NLMH 100024), and 1.85988 (NLMH 1000025 and 100026), with the 180 kV x-ray tube of the v|tome|x s µCT device (GE Sensing & Inspection Technologies GmbH phoenix|x-ray) housed in the Bonn Institute of Organismic Biology, University of Bonn, Germany. Scan settings were 100 kV and a shutter speed of 500 ms per capture for all specimens, at 130 µA (NLMH 100026), 160 µA (NLMH 100024 and 100025), and 180 µA (NLMH 100023). The instrument produces isotropic voxels, and single image size is 2048 × 2048 pixels. VGSTUDIO MAX (Volume Graphics GmbH, Heidelberg, Germany) was used for generation of the image stacks, and Avizo Lite 2020.2 (Thermo Fisher Scientific, Waltham, Massachusetts, USA) was used for segmentation. The studied specimens are curated in the State Museum (Landesmuseum) Hannover (NLMH), Germany.
Measurements: H—crown height; L—crown length; W—crown width. All measurements are in millimeters (mm).
In systematics of Mesozoic mammals and dental terminology, we follow Kielan-Jaworowska et al. (2004) and Martin (2018). Upper case letter (M) refers to upper, lowercase letter (m) to lower molars. Cusp notation is based on that proposed in Crompton and Jenkins (1968: 432).
Systematic palaeontology
Mammaliaformes Rowe, 1988
Morganucodonta Kermack, Musset et Rigney, 1973
Storchodon Martin et al., 2019
Storchodon cingulatus Martin et al., 2019

Storchodon cingulatus NLMH 110023, right lower molar (m1) in a lingual, b, b’ occlusal (stereopair), c labial, and d ventral views. Langenberg Quarry, Lower Saxony, Germany; Süntel Formation, Late Jurassic (late Kimmeridgian) age. Scale bar equals 1 mm. ic interradical crest

Storchodon cingulatus NLMH 110024, right lower molar (m3) in a lingual, b, b’ occlusal (stereopair), c labial, and d ventral views. Langenberg Quarry, Lower Saxony, Germany; Süntel Formation, Late Jurassic (late Kimmeridgian) age. Scale bar equals 1 mm. ic interradical crest

Storchodon cingulatus NLMH 110025, left lower molar in a labial, b, b’ occlusal (stereopair), c lingual, and d ventral views. Langenberg Quarry, Lower Saxony, Germany; Süntel Formation, Late Jurassic (late Kimmeridgian) age. Scale bar equals 1 mm. ic interradical crest

Storchodon cingulatus NLMH 110026, right upper molar in a labial, b, b’ occlusal (stereopair), c lingual, and d dorsal views. Langenberg Quarry, Lower Saxony, Germany; Süntel Formation, Late Jurassic (late Kimmeridgian) age. Scale bar equals 1 mm. ic interradical crest
Description
Lower molars
Specimen NLMH 110023 (Fig. 2) is a right lower molar, probably m1, because the indentation between the tiny mesial cuspules e and f for accommodation of cusp d of the preceding tooth (premolar) is very shallow. The crown is labio-lingually narrow and mesio-distally elongated. The three main cusps are aligned antero-posteriorly, with cusps b and c slightly shifted lingually, causing a faint angulation. Highest cusp is a, second highest is c, and third highest is b. The height difference between cusp b and c is minor, but cusp a is nearly twice as high as cusp b. All three main cusps are somewhat labio-lingually compressed and have a convex labial side. The lingual side is flat (cusp a) and respectively slightly convex (cusps b and c). Cusp a has a high triangular, somewhat asymmetrical shape with the tip facing distally. The anterior crest of cusp a rises in the beginning almost vertically and recurves after having reached one third of the cusp height in distal direction; it is slightly worn in its upper half. The distal crest rises steeply (75°) and is strongly worn with dentine exposure. Apically, cusp a bears a rounded wear facet with dentine exposure that is facing distally.
Cusp b is adjacent to cusp a and separated from it by a narrow notch. The notch is not very deep and both cusps share a common base. The shape of cusp b is acute triangular and symmetric. Apically, cusp b bears a small rounded wear facet with dentine exposure occlusally.
Cusp c is separated from cusp a by a wide and deep embrasure enhanced by prominent wear. Cusp c has the shape of an acute angular isosceles triangle. Apically, it bears an oval wear facet with dentine exposure that continues onto the anterior crest. The embrasure between cusps a and c is deeply worn on the labial side and the wear facet extends to the base of the crown. The distal crest of cusp c is worn, and on its distolingual side, cusp c bears a large wear facet at the base.
The slightly abraded cusp d is very small and connected to cusp c by a faint crest. The labial flank of cusp d is worn.
On the lingual side of the crown there is a prominent cingulid with two large cingulid cusps. The larger and more distal of these is cusp g which has a conical shape and is situated lingually to the embrasure between cusps a and c. The smaller and more mesially situated, unnamed cingulid cusp is labio-lingually compressed and sits lingually of the notch between cusps a and b. Both lingual cingulid cusps bear apical wear facets with dentine exposure. Cusp d is connected to the distal cingulid cusp by a ledge-like cingulid that narrows when approaching the cusp. Between both lingual cingulid cuspules, the cingulid is interrupted and marked only by two small enamel risings. Mesially to the anterior lingual cuspule, the cingulid continues and forms a very small cusp e with tiny apical wear facet. The cingulid runs around the anterior end of the crown and fades when reaching the labial side. Labially of cusp e is a wear facet at the crown base that has partially removed the cingulid. A very small cuspule f can be hardly distinguished from cusp e. It bears a distinct oval wear facet on the labial side.
The mesial root is missing and the distal root is broken apically. Both roots have round cross sections and are well separated from each other; a thin interradical crest connects the bases of the roots which are equally strong. The partially preserved distal root suggests that the intact roots were slightly divergent in mesial and distal direction.
Measurements: L = 2.21, W = 1.07, H = 2.48.
Specimen NLMH 110024 (Fig. 3) is a right lower molar. It is wider (and somewhat shorter) than NLMH 100023, has a much smaller cusp b, and possesses an indentation for molar interlock at the mesial end, bordered by cusps e and f, of which the latter is hardly discernible. The proportions of the molar and presence of a mesial indentation suggest that it is an m3. Similar to NLMH 100023, cusps b and c are slightly shifted lingually resulting in a faint angulation. Highest cusp a has an asymmetric shape, similar to cusp a of NLMH 100023, with slight wear on the upper half of the anterior crest. Cusp c is half as high as cusp a and has the shape of an acute angular isosceles triangle. The wide embrasure between cusps a and c is deeply worn on the labial side. Both cusps, a and c, are apically worn. The apical wear facets are confluent with the wear on the distal crest of cusp a respectively on the mesial crest of cusp c. At its distolabial side, cusp c bears a large wear facet that is broad at the base and becomes narrower towards the tip. Cusp d is tiny and apically damaged. It is connected to cusp c by a faint crest and its labial side is affected by the aforementioned wear facet. The tip of cusp b is worn off and it was taller originally, but judged by its small base and the remaining part of the tip, cusp b was still lower than cusp c in unworn stage. As in NLMH 100023, cusp b is close to cusp a, separated by a narrow notch. On the mesiolabial side, cusp b bears a tongue-shaped wear facet with dentine exposure, and on the lingual side a short crest extends in a mesio-lingual direction.
On the lingual side of the crown there is a ledge-like cingulid extending between cusps e and d, which is stronger (especially between the large cingulid cusps) than in NLMH 100023. As in NLMH 100023, large cingulid cusp g is situated lingually to the embrasure between cusps a and c; its mesiolingual portion is chipped off. The cingulid continues mesial to the cusp as a ledge that is slightly crenulated. The smaller and more mesially situated, unnamed cingulid cusp is about one third the size of the larger one and sits half way between the tip of cusp a and the deepest point of the notch between cusps a and b (which is more distal than in NLMH 100023); it is slightly damaged. The ledge-like cingulid continues in a mesial direction and ends in small cusp e at the mesiolingual side of the crown. Labially to cusp e, a shallow indentation for accommodation of cusp d of the preceding molar is present which indicates molar interlock.
The roots are both broken off and have a round cross section. They are well separated from each other, but connected by a thin interradical crest.
Measurements: L = 1.92, W = 1.10, H = 1.82.
Specimen NLMH 110025 (Fig. 4) is a left lower molar, missing the distal part with cusps c and d. The mesial fragment corresponds in shape and proportions to NLMH 110024. Cusp b is less worn apically than in NLMH 110024, but bears a tongue-shaped wear facet with dentine exposure on its labial side. Cusp a bears an apical wear facet with dentine exposure that extends onto the distal crest. Cusp e is sitting in a mesiolingual position and has an oval apical wear facet. Labially to cusp e the crown margin is slightly indented for molar interlock; the indentation is bordered labially by cusp f of which the labial portion is worn away by the tongue-shaped wear facet running down from cusp b. Cingulid cusp g is slightly damaged and is, in comparison to NLMH 110024, sitting slightly more mesially. The unnamed smaller, mesial lingual cingulid cusp is also positioned more mesially, and sits lingually to cusp b; apically it bears a small wear facet. Between cusp g and the smaller cingulid cusp the cingulid is less pronounced than in NLMH 110024. The mesial root has a round cross section and it is evident that both roots were well separated; a thin interradical crest is present.
Measurements: L = –, W = 0.90 (widest part of mesial fragment), H = 1.59.
Upper molar
Specimen NLMH 110026 (Fig. 5) is a right upper molar fragment that preserves cusps A and B. Cusp A is conical and has double the height of cusp B. It bears a small apical wear facet with dentine exposure. Cusp B is tetrahedron-shaped and bears an apical wear facet with dentine exposure; it is connected to the mesial cusp E by a faint crest. The wide embrasure between cusps A and B is strongly worn on the lingual side. The large wear facet has removed part of the distal flank of cusp B and large parts of the lingual cingulum. The mesial flank of cusp A is worn along the crest A-B and at the base. The A-C crest is worn along its entire preserved length. NLMH 110026 is more heavily worn than the holotype specimen (NLMH 105654) where only the lingual cingulum is worn but the lingual sides of cusps A and B (and embrasure in between) are not yet affected by wear. Cusp C is broken off, and a large wear facet on the lingual side indicates that the embrasure between cusps A and C was considerably worn. The lingual cingulum is ledge-like, but large parts of it have been removed by the lingual embrasure wear facets. The lingual cingulum extends from the mesial cuspule in a distal direction and forms a cuspule at the level of cusp B. Immediately behind this cuspule the cingulum is worn away, and only a short portion at the level of cusp A is intact. Behind the intact portion the cingulum is worn off. The unworn labial cingulum is narrower than the lingual cingulum and bears two labio-lingually compressed cuspules.
The mesial root is incomplete apically. It has a round cross section and diverges slightly in a mesial direction. The distal root is broken off, but the partly preserved base indicates that both roots were well separated and connected by a low interradical crest.
Measurements: L = –, W = 0.99; H = 2.35.
Comparison with lower molars of other morganucodontans
Apart from a larger size, cusps b and c in the lower molars of Storchodon are relatively higher than in Morganucodon watsoni and M. oehleri. Cusp a is asymmetric in Storchodon, with a slightly distally recurved tip; in Morganucodon cusp a is symmetric (Mills 1971; Jäger et al. 2019). The e–f–d interlock of Storchodon differs from the e-b-d interlock seen in Morganucodon watsoni (Luo 1994). All available lower Storchodon molars have a pronounced furcation between the roots on the labial and lingual sides. In Morganucodon watsoni this is the case only in m1 (Jäger et al. 2019), but the furcation is still less pronounced than in the m1 (NLMH 110023) of Storchodon. The molars of Erythrotherium parringtoni, which are very similar to that of Morganucodon, are much smaller than those of Storchodon. The lower molars of Storchodon differ from the holotype specimen of Cherwellia leei Butler and Sigogneau-Russell (2016) from the late Bathonian of Kirtlington by a larger cusp g and a narrower lingual cingulid. The Cherwellia leei holotype specimen (M46811; L = 2.60 mm) is slightly larger than NLMH 110023 (m1; L = 2.21 mm). M46811 shows a mesial embayment for molar interlock (Butler and Sigogneau-Russell 2016: Fig. 1B3) which suggests that it is not an m1, but possibly an m2, which is the largest lower molar in Morganucodon watsoni (Jäger et al. 2019). Under this assumption, Storchodon and Cherwellia are in the same size class. In the lower molar holotype specimen of Stylidens hookeri Butler and Sigogneau-Russell (2016) from the late Bathonian of Watton Cliff (Dorset, England), cusp c is significantly larger than b, and cusp g is much smaller than in Storchodon, more a long swelling than a cusp. The lower molar (DORCM GS 734) tentatively attributed to the questionable morganucodontan Purbeckodon batei from the Lower Cretaceous Lulworth Formation of the Purbeck Limestone Group (Butler et al. 2012) is much smaller than the lower molars of Storchodon and differs greatly by its asymmetric crown with mesially shifted large cusp a, very small cusps b and c, and weak and interrupted lingual cingulid without cusp g. In contrast to Gondwanadon from the Late Triassic Tiki Formation in India (Datta and Das 1996), cusp a is distinctly larger than b and c in Storchodon. In Bridetherium dorisae from the Early Jurassic of Glamorgan (Clemens 2011), cusps b and c are relatively smaller than in Storchodon. Mesiolingual cusp e is much larger in Bridetherium which also has a distolingual cuspule that is absent in Storchodon. The holotype and only known specimen of Paceyodon davidi (NHMUK M45920) from the same deposit as Bridetherium found in (Pant Quarry, fissure 4) (Clemens 2011) is larger (150%) than the largest known lower molar of Storchodon (NLMH 100023). Cusp a is relatively lower in Paceyodon, the mesiolingual shelf is much larger, and cusp g is small, if present at all (Clemens 2011). Hallautherium schalchi from the Rhaetic bonebed (Late Triassic) of Hallau, Switzerland lacks the lingual cingulid and cusp g, and has relatively smaller cusps b and c (Clemens 1980). In the lower molars of the Early Jurassic megazostrodontid Dinnetherium nezorum from the Kayenta Formation in Arizona (Jenkins et al. 1983), cusp a is taller than in Storchodon. Megazostrodon rudnerae from the Early Jurassic of Lesotho (Crompton and Jenkins 1968) exhibits embrasure occlusion and has a hypertrophied kuehneocone (Parrington 1967; Crompton 1974). Storchodon has a similar molar interlock between cuspules e–f–d as seen in D. nezorum and M. rudnerae (Luo 1994). Storchodon lacks the wrinkled enamel surfaces of Brachyzostrodon, Wareolestes, and Helvetiodon (Clemens 1980; Hahn et al. 1991; Panciroli et al. 2017).
Discussion
Size of Storchodon and possible island gigantism
Martin et al. (2019) noticed the large size of the Storchodon holotype (upper) molar (length 3.07 mm) and interpreted this as an effect of insular isolation within the Langenberg fauna living on a palaeoisland in the Late Jurassic Lower Saxony Basin. On islands, large animals tend to become smaller due to a limited supply of foodstuffs (insular dwarfism) and small animals tend to grow larger due to a lack of predators (insular gigantism). In the Langenberg vertebrate assemblage both phenomena are represented. The first by the dwarfed sauropod dinosaur Europasaurus holgeri (Sander et al. 2006) and the second possibly by Storchodon and the large multituberculate mammal Teutonodon (Martin et al. 2016, 2019). Sizewise, the holotype molar of Storchodon is second only to Paceyodon davidi (lower molar length: 3.3 mm) and larger than Brachyzostrodon maior (lower molar length: 2.7 mm). Because the lower molars of Storchodon cingulatus were unknown previously, no direct size comparison with Paceyodon davidi and Brachyzostrodon maior was possible. Unfortunately, the phenomenon of island gigantism in the Langenberg vertebrate assemblage becomes less evident with the new lower molars of Storchodon. The available lower Storchodon molars are shorter than the (upper) holotype specimen. With a length of 2.21 mm and a width of 1.07 mm, NLMH 110023 is only 82% (length) of the known lower Brachyzostrodon maior molar (length 2.7 mm, width 1.2 mm; Hahn et al. 1991). According to Jäger et al. (2019), m2 is the longest molar in the lower molar row of Morganucodon watsoni (about 15% longer than m1 as evident from Fig. 2 in Jäger et al. 2019). Specimen NLMH 110023 is most likely an m1, as suggested by the faint mesial indentation for interlock (accommodation of cusp d of the preceding tooth), which is more pronounced in the other lower molars (NLMH 110024 and 110025) from the Langenberg Quarry. A similar gradient of molar interlock has been reported for other morganucodontans such as Megazostrodon rudnerae (Crompton 1974) and Dinnetherium nezorum (Crompton and Luo 1993). Specimen NLMH 110024 is shorter and narrower than NLMH 110023 and is probably an m3 (mesial fragment NLMH 100025 appears to be of similar size). The m2 of Storchodon cingulatus, which (in analogy to Morganucodon) is expected to be about 10–20% longer than the m1, is still unknown. With an estimated length of about 2.5–2.7 mm, the hypothetical m2 of Storchodon cingulatus would be of similar size to the known Brachyzostrodon maior lower molar (2.7 mm). If compared with the mean length of NLMH 110023 and 110024, the lower molars of Wareolestes rex (113%) and the putative morganucodontan Cherwellia leei (126%) are larger than those of Storchodon (measurements from Butler and Sigogneau-Russell 2016 and Panciroli et al. 2017). The lower molars of Morganucodon watsoni (60%), M. oehleri (86%), Bridetherium dorisae (73% of Storchodon for mesial lower molars and 55% for distal lower molars), Hallautherium schalchi (86%), Brachyzostrodon coupatezi (95%), and the putative morganucodontan Stylidens hookeri (90%) are smaller than those of Storchodon (percentages calculated from the mean values of measurements in Clemens 2011: tab. 1). No measurements of teeth are available for Erythrotherium, Megazostrodon, and Dinnetherium, but from the scale bars in the figures provided, it is evident that they are considerably smaller than those of Storchodon (Crompton 1964; Figs. 1 and 4; Jenkins et al. 1983; Fig. 1d, e). The size discrepancy between the single known upper and the lower molars of Storchodon can be explained by variation along the tooth row (longest lower molar of Storchodon not yet known) or just individual body size variation. While not supporting clear evidence for island gigantism, the new lower molars of Storchodon still document a large morganucodontan, surpassed only by Paceyodon, Wareolestes, and the putative morganucodontan Cherwellia.
Molar occlusion and dietary adaptation of Storchodon
The wear pattern in the lower Storchodon molars corresponds to that observed in Morganucodon watsoni (Jäger et al. 2019). Cusp A of the upper molar occludes into the valley between cusps a and c, causing the large confluent wear facets seen in this area. The smaller wear facets present on the mesiolabial flank of cusp b and the distolabial flank of cusp c were caused by cusp C of the upper antagonist that occluded into the embrasure between two lower molars. The occlusion of cusp a in the valley between cusps A and B caused the large wear facet seen on these cusps in NLMH 100026. This occlusal pattern differs from that seen in Megazostrodon, where cusp a occludes between two upper molars and cusp A occludes between two lower molars (= embrasure occlusion), and where cusp a is relatively lower and the valley between cusps a and c narrower (Crompton and Jenkins 1968; Crompton 1974; Jäger et al. 2019). Jäger et al. (2022) confirmed embrasure occlusion for the morganucodontid Erythrotherium parringtoni based on a virtual reconstruction of the chewing path. Jäger et al. (2020) suggested embrasure occlusion for Triconodontidae which previously were believed to have had a Morganucodon-like occlusion (Mills 1971). So far, the Morganucodon-like occlusal mode is restricted to the morganucodontans Morganucodon, Dinnetherium nezorum (Crompton and Luo 1993), the recently described Cifellilestes ciscoensis from the Upper Jurassic Morrison Formation (Davis et al. 2022), and Storchodon cingulatus, whereas Megazostrodon and Erythrotherium exhibit embrasure occlusion (Crompton 1964; Crompton and Jenkins 1968). The slight angulation of cusps observed in the upper molar of Storchodon cingulatus with lingual offset of cusp D resulting in a slight convex curvature of the line A-B-C-D (Martin et al. 2019: text-Fig. 2) is mirrored in the lower molars where cusps b and d are slightly shifted lingually.
Gill et al. (2014) suggested a dietary adaptation to hard-shelled arthropods for Morganucodon. Given the large size of Storchodon cingulatus and the shape of its molars with pointed cusps and sharp cutting edges, it probably preyed on small vertebrates as well as insects, and can be considered at least a second order consumer in the Langenberg insular palaeoecosystem as suggested by Martin et al. (2019).
Data availability
Specimens are deposited in the cited collections and institutions. Images of segmentation of specimens scanned by μCT for this study are presented in the text figures. Additional μCT scan data are available from the corresponding author upon reasonable request.
Abbreviations
- DORCM:
-
Dorset County Museum, England, UK
- M (Mammalia):
-
Specimens deposited at the Natural History Museum, London, UK
- NHMUK:
-
Natural History Museum, London, UK
- NLMH:
-
Landesmuseum Hannover, Germany
References
Butler, P.M., and D. Sigogneau-Russell. 2016. Diversity of triconodonts in the Middle Jurassic of Great Britain. Palaeontologia Polonica 65: 35–65. https://doi.org/10.4202/pp.2016.67_035.
Butler, P.M., D. Sigogneau-Russell, and P.C. Ensom. 2012. Possible persistence of the morganucodontans in the Lower Cretaceous Purbeck Limestone Group (Dorset, England). Cretaceous Research 33: 135–145.
Carballido, J.L., and P.M. Sander. 2014. Postcranial axial skeleton of Europasaurus holgeri (Dinosauria, Sauropoda) from the Upper Jurassic of Germany: Implications for sauropod ontogeny and phylogenetic relationships of basal Macronaria. Journal of Systematic Palaeontology 12: 335–387.
Carballido, J.L., M. Scheil, N. Knötschke, and P.M. Sander. 2020. The appendicular skeleton of the dwarf macronarian sauropod Europasaurus holgeri from the Late Jurassic of Germany and a re-evaluation of its systematic affinities. Journal of Systematic Palaeontology 18: 739–781. https://doi.org/10.1080/14772019.2019.1683770.
Clemens, W.A. 1980. Rhaeto-Liassic mammals from Switzerland and West Germany. Zitteliana 5: 51–92.
Clemens, W.A. 2011. New morganucodontans from an Early Jurassic fissure filling in Wales (united Kingdom). Palaeontology 54: 1139–1156.
Crompton, A.W. 1964. A preliminary description of a new mammal from the Upper Triassic of South Africa. Proceedings of the Zoological Society of London 142: 441–452.
Crompton, A.W. 1974. The dentitions and relationships of the southern African Triassic mammals Erythrotherium parringtoni and Megazostrodon rudnerae. Bulletin of the British Museum (Natural History). Geology 24: 397–437.
Crompton, A.W., and F.A. Jenkins Jr. 1968. Molar occlusion in Late Triassic mammals. Biological Reviews 43: 427–458.
Crompton, A.W., and Z.-X. Luo. 1993. Relationships of the Liassic mammals Sinoconodon, Morganucodon, and Dinnetherium. In Mammal Phylogeny: Mesozoic Differentiation, Multituberculates, Monotremes, Early Therians, and Marsupials, eds. F.S. Szalay, M.J. Novacek, and M.C. McKenna, 30–44. New York: Springer-Verlag.
Datta, P.M., and D.P. Das. 1996. Discovery of the oldest fossil mammal from India. India Minerals 50: 217–222.
Davis, B.M., K.R.K. Jäger, G.W. Rougier, K. Trujillo, and K. Chamberlain. 2022. A morganucodontan mammaliaform from the Upper Jurassic Morrison Formation, Utah, USA. Acta Palaeontologica Polonica 67: 77–93.
Evers, S.W., and O. Wings. 2020. Late Jurassic theropod dinosaur bones from the Langenberg Quarry (Lower Saxony, Germany) provide evidence for several theropod lineages in the central European archipelago. PeerJ 8: e8437. https://doi.org/10.7717/peerj.8437.
Fastnacht, M. 2005. The first dsungaripterid pterosaur from the Kimmeridgian of Germany and the biomechanics of pterosaur long bones. Acta Palaeontologica Polonica 50: 273–288.
Fischer, R. 1991. Die Oberjura-Schichtenfolge des Langenbergs bei Oker. Arbeitskreis Paläontologie Hannover 19: 21–36.
Gerke, O., and O. Wings. 2016. Multivariate and cladistic analyses of isolated teeth reveal sympatry of theropod dinosaurs in the Late Jurassic of Northern Germany. PLOS ONE 11: e0158334.
Gill, P.G., M.A. Purnell, N. Crumpton, K.R. Brown, N.J. Gostling, M. Stampanoni, and E.J. Rayfield. 2014. Dietary specializations and diversity in feeding ecology of the earliest stem mammals. Nature 512: 303–305.
Hahn, G., D. Sigogneau-Russell, and P. Godefroit. 1991. New data on Brachyzostrodon (Mammalia; Upper Triassic). Geologica et Palaeontologica 25: 237–249.
Jäger, K.R.K., P.G. Gill, I. Corfe, and T. Martin. 2019. Occlusion and dental function of Morganucodon and Megazostrodon. Journal of Vertebrate Paleontology 39: e1635135.
Jäger, K.R.K., R.L. Cifelli, and T. Martin. 2020. Molar occlusion and jaw roll in early crown mammals. Scientific Reports 10: 22378.
Jäger, K.R.K., P.G. Gill, T. Martin, and I. Corfe. 2022. Molar morphology and occlusion of the Early Jurassic mammaliaform Erythrotherium parringtoni. Acta Palaeontologica Polonica 67: 975–982.
Jansen, M., and N. Klein. 2014. A juvenile turtle (Testudines, Eucryptodira) from the Upper Jurassic of Langenberg Quarry, Oker, Northern Germany. Palaeontology 57: 743–756. https://doi.org/10.1111/pala.12085.
Jenkins, F.A., A.W. Crompton, and W.R. Downs. 1983. Mesozoic mammals from Arizona: New evidence on mammalian evolution. Science 222: 1233–1235.
Karl, H.-V., E. Gröning, C. Brauckmann, D. Schwarz, and N. Knötschke. 2006. The Late Jurassic crocodiles of the Langenberg near Oker, Lower Saxony (Germany), and description of related materials (with remarks on the history of quarrying the “Langenberg Limestone” and “Obernkirchen Sandstone”). Clausthaler Geowissenschaften 5: 59–77.
Kermack, K.A., F. Mussett, and H.W. Rigney. 1973. The lower jaw of Morganucodon. Zoological Journal of the Linnean Society 53: 87–175.
Kielan-Jaworowska, Z., R.L. Cifelli, and Z.-X. Luo. 2004. Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure, 630. New York: Columbia University Press.
Lallensack, J. N., P. M. Sander, N. Knötschke, and O. Wings. 2015. Dinosaur tracks from the Langenberg Quarry (Late Jurassic, Germany) reconstructed with historical photogrammetry: Evidence for large theropods soon after insular dwarfism. Palaeontologia Electronica 18.2.31A: 1–34.
Lotze, F. 1968. Zum Jura des Langenberges zwischen Oker und Bad Harzburg (nördl. Harzrand). Neues Jahrbuch Für Geologie Und Paläontologie, Monatshefte 1968: 730–732.
Luo, Z.-X. 1994. Sister-group relationships of mammals and transformations of diagnostic mammalian characters. In In the Shadow of the Dinosaurs: Early Mesozoic Tetrapods, eds. N.C. Fraser and H.-D. Sues, 98–128. Cambridge: Cambridge University Press.
Marpmann, J.S., J.L. Carballido, P.M. Sander, and N. Knötschke. 2015. Cranial anatomy of the Late Jurassic dwarf sauropod Europasaurus holgeri (Dinosauria, Camarasauromorpha): Ontogenetic changes and size dimorphism. Journal of Systematic Palaeontology 13: 221–263.
Martin, T. 2018. Mesozoic mammals—Early mammalian diversity and ecomorphological adaptations. In Handbook of Zoology, Mammalia. Mammalian Evolution, Diversity and Systematics, eds. F.E. Zachos and R.J. Asher, 199–299. Berlin: De Gruyter.
Martin, T., J.A. Schultz, A.H. Schwermann, and O. Wings. 2016. First Jurassic mammals of Germany: Multituberculate teeth from Langenberg Quarry (Lower Saxony). Palaeontologia Polonica 67: 171–179.
Martin, T., A.O. Averianov, K.R.K. Jäger, A.H. Schwermann, and O. Wings. 2019. A large morganucodontan mammaliaform from the Late Jurassic of Germany. Fossil Imprint 75: 504–509.
Martin, T., A.O. Averianov, J.A. Schultz, A.H. Schwermann, and O. Wings. 2021a. Late Jurassic multituberculate mammals from Langenberg Quarry (Lower Saxony, Germany) and palaeobiogeography of European Jurassic multituberculates. Historical Biology. https://doi.org/10.1080/08912963.2019.1650274.
Martin, T., A.O. Averianov, J.A. Schultz, A.H. Schwermann, and O. Wings. 2021b. A derived dryolestid mammal indicates possible insular endemism in the Late Jurassic of Germany. The Science of Nature 108: 23. https://doi.org/10.1007/s00114-021-01719-z.
Martin, T., A.O. Averianov, J.A. Schultz, A.J. Lang, and O. Wings. 2024. Docodontans (Mammaliaformes) from the Late Jurassic of Germany. Historical Biology. https://doi.org/10.1080/08912963.2023.2300635.
Mills, J. R. E. 1971. The dentition of Morganucodon. In Early Mammals, eds. D. M. Kermack, K. A. Kermack. Zoological Journal of the Linnean Society, London 50 (Supplement 1): 29–63.
Mudroch, A., and D.E.W. Thies. 1996. Knochenfischzähne (Osteichthyes, Actinopterygii) aus dem Oberjura (Kimmeridgium) des Langenbergs bei Oker (Norddeutschland). Geologica Et Palaeontologica 30: 239–265.
Panciroli, E., R.B.J. Benson, and S.A. Walsh. 2017. The dentary of Wareolestes rex (Megazostrodontidae): A new specimen from Scotland and implications for morganucodontan tooth replacement. Papers in Palaeontology 3: 373–386.
Pape, H. 1970. Die Malmschichtfolge vom Langenberg bei Oker (nördl. Harzvorland). Mitteilungen aus dem Geologischen Institut der Technischen Universität Hannover 9: 41–134.
Parrington, F.R. 1967. The origin of mammals. Advancements in Science 24: 165–173.
Richter, A., N. Knötschke, R. Kosma, G. Sobral, and O. Wings. 2013. The first Mesozoic lizard from northern Germany (Paramacellodidae, Late Jurassic, Langenberg Quarry) and its taphonomy. Program and Abstracts, Society of Vertebrate Paleontology 73rd Annual Meeting, October 30 —November 2, 2013, Los Angeles, USA. Supplement to the online Journal of Vertebrate Paleontology, October 2013: 198.
Rowe, T.B. 1988. Definition, diagnosis, and origin of Mammalia. Journal of Vertebrate Paleontology 8: 241–264.
Sander, P.M., O. Mateus, T. Laven, and N. Knötschke. 2006. Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur. Nature 441: 739–741.
Schwarz, D., M. Raddatz, and O. Wings. 2017. Knoetschkesuchus langenbergensis gen. nov. sp. nov., a new atoposaurid crocodyliform from the Upper Jurassic Langenberg Quarry (Lower Saxony, northwestern Germany), and its relationships to Theriosuchus. PLoS One 12: e0160617.
Thies, D.E.W., R. Windolf, and A. Mudroch. 1997. First record of Atoposauridae (Crocodylia: Metamesosuchia) in the Upper Jurassic (Kimmeridgian) of Northwest Germany. Neues Jahrbuch Für Geologie Und Paläontologie, Abhandlungen 205: 393–411.
van der Lubbe, T., U. Richter, and N. Knötschke. 2009. Velociraptorine dromaeosaurid teeth from the Kimmeridgian (Late Jurassic) of Germany. Acta Palaeontologica Polonica 54: 401–408. https://doi.org/10.4202/app.2008.0007.
Wings, O. 2015. The Langenberg Quarry near Goslar: Unique window into a terrestrial Late Jurassic ecosystem in Northern Germany. In Abstracts, 12th Symposium of Mesozoic Terrestrial Ecosystems August 16th—20th 2015, Shenyang, China. Eds. Y. Zhang, S. Z. Wu, and G. Sun, 99–100.
Zuo, F., U. Heimhofer, S. Huck, F.W. Luppold, O. Wings, and J. Erbacher. 2018. Sedimentology and depositional sequences of a Kimmeridgian carbonate ramp system, Lower Saxony Basin, Northern Germany. Facies 64: 1. https://doi.org/10.1007/s10347-017-0513-0.
Acknowledgements
Nils Knötschke (formerly Dinosaurier-Park Münchehagen), Olaf Dülfer, Rico Schellhorn, Julia Schultz (all University of Bonn), Achim Schwermann (LWL-Museum Münster), student helpers, and volunteers assisted in the field. Carolin Geißler, Josephine Jolie, Anna Kizner, Melody Kofranek, Jens Lallensack, Jan Matzker, Denis Theda, and Jannik Weidtke (all University of Bonn) sorted the screen washing concentrate for fossils. Janna von Pupka (Rohstoffbetriebe Oker) granted access to the Langenberg quarry and provided logistic support. We thank Kai Jäger (Bonn) for comments on an earlier draft and discussions. Referees Pam Gill (Bristol) and Zhe-Xi Luo (Chicago) provided constructive comments and suggestions. This research was funded by a grant (MA 1643/26-1) from the German Research Foundation (DFG) to TM. AA was supported by the Zoological Institute, Russian Academy of Sciences (project 122031100282-2) and a Humboldt Research Award by the Alexander von Humboldt Foundation. The Europasaurus-Project (grant no. 85 882) and OW were funded by the Volkswagen Foundation within the initiative “Research in Museums”.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Handling Editor: Hans-Dieter Sues.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Martin, T., Averianov, A.O., Lang, A.J. et al. Lower molars of the large morganucodontan Storchodon cingulatus from the Late Jurassic (Kimmeridgian) of Germany. PalZ (2024). https://doi.org/10.1007/s12542-024-00690-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12542-024-00690-0