
Abstract
The Jebel Begaa region of the eastern Anti-Atlas in SE Morocco yielded a single, very well-preserved goniatite that is closely related to Helicocyclus divergens Riley, 1996 from the lower Viséan (Chadian) of northern England. Both are placed in a new genus, Tectocyclus n. gen., with the Moroccan T. herbigi n. sp. as the type species. Characteristic are its subtriangular suture elements, which distinguish it strongly from Helicocyclus gracillimus Schindewolf, 1951, which is revised based on the re-examination of its types. The type-species of Helicocyclus is characterized by wide, bell-shaped lateral lobes, unlike as in described helicocyclids from the upper Tournaisian and Viséan of North Africa, Russia, and Central Asia. These are assigned, partly with reservation, to Serpenticyclus n. gen., with the oldest species, Helicocyclus fuscus Korn, Bockwinkel, Ebbighausen, and Klug, 2003, as the type-species. All three genera form the Helicocyclinae n. subfam., which first appeared in North Africa, migrated at the end of the Tournaisian across the Variscan Sea to the southern shelf of Laurussia, to Central Asia, and to the Urals seaway, where it survived into Arundian levels of the Viséan.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The eastern Anti-Atlas of southern Morocco is world famous for its abundant and diverse Devonian ammonoid faunas but a range of studies from roughly the last two decades (e.g. Korn et al. 1999, 2003, 2007; Bockwinkel and Ebbighausen 2006; Klug et al. 2006, 2016; Ebbighausen and Bockwinkel 2007; Becker et al. 2018a, 2018b) showed that Lower Carboniferous (Mississippian) faunas can also be rich in terms of specimen and species numbers. They are of high significance for phylogeny, regional and international biostratigraphy, palaeobiogeography, and palaeodiversity analyses. Their investigation is far from complete (see Becker 2019); various genera and species are still undescribed. This includes a pericyclid specimen purchased in the field from boys of the Begaa village (= Hassi El Beg`a) in the southeastern Tafilalt (Beg in Fig. 1), which is unique in terms of preservation and closely related to the English Helicocyclus divergens Riley, 1996. Its description and the revision of the type-species of Helicocyclus enable progress in pericyclid systematics and some palaeobiogeographic conclusions based on the current, certainly still incomplete knowledge.

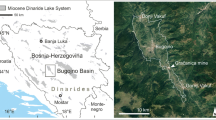
Simplified geological map of the eastern Anti-Atlas showing the position of known pericyclid localities. BE = Bordj Erfoud (top Tournaisian), KB = Kzar Bouhamed (lower part of upper Tournaisian), Beg = Begaa (reworked higher upper Tournaisian and probable lower Viséan), EA = El Atrous and lateral collecting spots (middle Tournaisian), OZ = Oued Ziz, 7 km north of Ouzina (lower part of upper Tournaisian), Jd = south of Jdaid (lower part of upper Tournaisian), Rh = Rharrhiz (middle Tournaisian)
Chronostratigraphic terminology
Substages used in this paper are informal chronostratigraphic units without ICS approval, which definitions are potentially subjective and, therefore, briefly explained (for more details see Aretz et al. 2020). The base of the middle Tournaisian follows its traditional definition starting with the Belgian Tn 2 (= base upper Hastarian) and the base of the German Pericyclus Stufe (cu II). The base of the upper Tournaisian agrees with the base of the Belgian Ivorian (Hance et al. 2006; Tn 3) and is correlated with the base of the North American Osagean (Lane and Brenckle 2005; Boardman et al. 2013). Following the proposal of Waters et al. (2011) and Aretz et al. (2020), the base of the Viséan coincides with the base of the British Chadian substage (= upper Chadian sensu Riley 1996). The base of the second British Viséan substage, the Arundian, predates the base of the Belgian middle Viséan, defined at the base of the Livian (Poty and Hance 2006; Aretz et al. 2020, p. 825; roughly = base of British Holkerian). The base of the upper Viséan in the sense of the Belgian Warnantian predates the base of the English Asbian (Aretz et al. 2020) but is roughly correlated with the base of the Goniatites Stufe (cu III) of the pelagic ammonoid succession (e.g. Korn 2006).
Locality, regional geology and age
The studied specimen is preserved as an internal phragmocone mould in shiny, middle- to dark-brown goethite, without any distortion. It was recognized ca. 20 years ago among a large collection of similarly preserved upper Givetian goniatites that was assembled by boys from the Begaa village in the Hassi Nebech region of the eastern Tafilalt (see Givetian Hassi Nebech monograph by Bockwinkel et al. 2013). It was the only Carboniferous ammonoid among more than 1.000 Middle Devonian specimens. It was collected loose, either around their home village or on the way from Begaa to Hassi Nebech. This area belongs on the geological map of Hollard (1971, fig. 4) to the “Schistes gris somber et calcaires du Viséen inferieur”. Hollard recognized a lower thick unit of shales with nodules, covered by the crinoidal “calcaire récifaux” that forms the widely visible Jebel Begaa cliff (= Jebel El Beg`a; Hollard 1971; Fig. 2). In the revised regional lithostratigraphy of geological sheet 1:50.000 Tawz (= Taouz, Alvaro et al. 2014), this succession belongs to the up to 800 m thick Merdani Formation. In the area at the western and southwestern foot of the Jebel Begaa (Fig. 3), there is a thick, poorly exposed lower shale unit, followed by an alternation of shales and light-grey crinoidal limestones that pass laterally into a chaotically bedded or conglomeratic debris flow and olistolite succession, followed by a package of greenish, silty shales with siderite nodules, and eventually by the crinoidal to biostromal Begaa Member that contains colonial rugose corals. A younger very thick package of shales follows towards the north and east, interrupted by several levels of mudmounds (Wendt et al. 2001). As outlined by Alvaro et al. (2014, fig. 40), the lower shaly member wedges out towards Hassi Nebech, where the upper shales are widely covered. Since the preservation is typical for hypoxic shale, the specimen came probably not from the Hassi Nebech region.

Field photo of the Jebel Begaa from the west, showing the rugged weathering of biostromal and crinoidal limestones of the Begaa Member at the top, partly covered greenish shales with Arundian pelagic fauna of the lower Merdani Formation in the slope and light-grey crinoidal limestones and the chaotic conglomeratic unit at the base and to the left. The small Begaa village lies outside the photo, across a small wadi less than a km to the right

Geological map of the Jebel Begaa (= Jebel El Beg`a) and Hassi Nebech region, showing the significant local extent of the Merdani Formation (dark bluish-green, hv1) as the likely source of Tectocyclus herbigi n. gen. n. sp. (extracted from geological sheet, 1: 50 000, Taouz, Alvaro et al. 2014); x coordinates at the top = 1 km, d3b = upper Emsian/basal Eifelian Amerboh Formation, d5 = Givetian part of Boutchrafine Formation, d6 = upper Frasnian part of Achguig Formation (Kellwasser facies), d7c = uppermost Famennian to lower Tournaisian Aoufilal Formation, hT = middle/upper Tournaisian Oued Znaïgui Formation, q1-4, qd, Af, and AI = various Quaternary deposits, δ = dolerite; Hassi El Beg`a = Begaa village
All units of the Merdani Formation were originally placed in the lower Viséan by Delepine (1941) and Hollard (1970). Support came from conodont dating by Wendt et al. (2001: records of the Viséan index species Gnathodus texanus) and by the age of a coral fauna from the conglomeratic limestones at the western foot (Aretz et al. 2013). The latter includes reworked, yellowish weathering micritic pebbles with crinoids, goniatites (e.g. Ammonellipsites) and trilobites. The fauna resembles that of the famous, top-Tournaisian Erdbach Limestone of Germany, but there are also subordinate typical Viséan forms, such as Michiganites (e.g. Riley 1996; Work et al. 2000; Becker 2019). Despite numerous search attempts in the last 20 years, no Carboniferous goniatite with similar preservation as the here described specimen has been found in the Jebel Begaa-Hassi Nebech region. Laterally to the conglomerate mentioned above, the lower Merdani Formation yielded rare Merocanites from light-grey crinoidal limestone. Small, squashed, sideritic goniatites, including ?Rotopericyclus, occur in the lower part of the overlying shales, rare siderite nodules with Bollandoceras and Bollandites in the greenish silty shales just below the Jebel Begaa cliff. The latter two genera are indicative of the Arundian/Holkerian, which supports the assumed younger, basal upper Viséan age of corals from the overlying Begaa Member (Aretz et al. 2013; Aretz in Alvaro et al. 2014). In the absence of similarly preserved specimens, it is not possible to judge whether the new pericyclid came from lower Viséan (Chadian) shales below or from Arundian/Holkerian level shales above the conglomerate level. The shales above the Begaa Member can be excluded since on the global scale, no pericyclid has ever been found in the upper Viséan.
The comparison with the morphologically very close Helicocyclus divergens from the Craven Basin of northwest England supports a lower Viséan (Chadian) age for the Moroccan form. The British species was described from the Leagram Mudstone Member of the Hodder Mudstone Formation, where it is associated with Viséan index genera, such as Michiganites and Eonomismoceras. In his refined zonation, Riley (1996) assigned H. divergens to the Viséan (“Upper Chadian”) part of his Fascipericyclus-Ammonellipsites Zone that straddles the Tournaisian/Viséan boundary. Tilsley and Korn (2009) described a second population of H. divergens from a locality (Wetton Hill East) of the Peak District, which lies east of the Craven Basin. Its association with Eonomismoceras, but without Bollandites/Bollandoceras, suggests the same age as in the latter and lends further support for a lower Viséan (Chadian) age of the Tafilalt specimen.
Previous records of Anti-Atlas pericyclids
The first Anti-Atlas Pericyclus was discovered by Clariond (1934), together with muensteroceratids, at the Butte or Bordj Erfoud (= Djebel Erfoud) of the central Tafilalt (BE in Fig. 1). Additional collecting produced a richer fauna (Delepine 1939), which was then described in a monograph by Delepine (1941). Based on a partial re-examination of originals and on limited new material, a revision of this important top-Tournaisian fauna has begun. Preliminary data (Becker 2019) confirmed the presence of three pericyclid genera, Rotopericyclus, Ammonellipsites, and Hammatocyclus.
A second pericyclid locality of the Tafilalt was announced by Clariond (1954), who noted that O.H. Schindewolf had identified Pericyclus and Muensteroceras from the shales underlying the rugged crinoidal limestones of the Jebel Begaa (= Begaa Member of Merdani Formation, Beg in Fig. 1). Additional details for this locality were provided by Hollard (1971). As outlined above, re-sampling yielded a single poorly preserved pericyclid but the associated Bollandites-Bollandoceras fauna contradicts an upper Tournaisian age, as considered by Wendt et al. (2001, p. 220). It is likely that their quoted conodonts from the Scaliognathus anchoralis Zone derived from reworked limestone in the mentioned conglomerate.
According to Hollard & Jaquemont (1956), pyritic Eoglyphioceras and Fascipericyclus occur in a transgressive unit just above the Djebel Tazout in the western Dra Valley south of Assa (DZ in Fig. 1). Later, Hollard (1971) added that relatives of Pericyclus hauchecornei Holzapfel, 1889 enter at a higher level in the Betaina plain.
A third Tafilalt fauna with pericyclids was discovered by Hollard (1960, 1971), in company of J. Agard and B. Owodenko, at El Atrous in the Amessoui Syncline (EA in Fig. 1). Korn et al. (2002) showed that this occurrence in the Oued Znaïgui Formation refers to a middle Tournaisian Goniocyclus fauna. There are several collection spots along the syncline. Rich collections from El Atrous belong to successive G. ammari and G. elatrous faunas (Becker et al. 2006; Kaiser et al. 2013, 2018) but are not yet fully documented. Ebbighausen and Bockwinkel (2007) documented the Goniocyclus fauna also from the eastern margin of the Aguelmous Syncline in the central Maïder, at Rharrhiz (Rh in Fig. 1).
Korn et al. (2003) described four genera/species of pericyclids, including the oldest known helicocyclid, from the lower part of the upper Tournaisian at the eastern end of the Amessoui Syncline, just east of the Kzar of the Bouhamed family (KB in Fig. 1). The same fauna occurs 4 km south of Jdaid in the southeast corner of the Maïder (Korn et al. 2007, Jd in Fig. 1). “Pericyclus” mercatorius was also found within a “Bouhamed Fauna” collected between the winding piste from Taouz to Ouzina and the Oued Ziz, ca. 9.5 km NNE of Ouzina, well before the piste makes a final sharp turn to the south (OZ in Fig. 1).
Systematic palaeontology
Superfamily Pericycloidea Hyatt, 1900
Family Pericyclidae Hyatt, 1900
Subfamily Helicocyclinae n. subfam.
Type-genus. Helicocyclus Schindewolf, 1951
Diagnosis. Small-sized, early stages evolute and depressed (serpenticonic), adult stages (sub)evolute, rarely subinvolute, discoidal to extremely discoidal (depressed or compressed), ornament sublinear or concave, ventral sinus reduced, ribbing weak to absent, with regular, linear or concave shell constrictions. Sutures with weakly to strongly divergent E-lobe, low to moderately high median saddle (up to 50% of external saddle), rounded or subtriangular flank saddles, pointed A-lobe, short, funnel-shaped, subdivided, or bell-shaped L-lobe, deep internal U- and I-lobes. Suture formula: (E1E2E1)AL:UI to (E2E1E2)ALe:LiUI.
Included genera. Helicocyclus, Serpenticyclus n. gen., Tectocyclus n. gen., and possibly Gen. nov. A sensu Kusina and Konovalova (2004).
Discussion. The Pericycloidea have traditionally been placed in the suborder Goniatitina. They originated in the basal middle Tournaisian earlier and independently from the Muensteroceratidae, which did not appear before the top of the middle Tournaisian. This is confirmed by the examination of a juvenile topotype of one of the oldest muensteroceratids, Xinjiangites medius (Miller and Collinson, 1951), which bears no similarity in terms of shell form, sutures, and ornament with the strongly ribbed, globally widespread middle Tournaisian goniocyclids. All known top-middle Tournaisian Xinjiangites species, such as X. medius, X. brevis (Ruan, 1995), X. applanatus (Ruan, 1995), and X. modestus (Popov and Kusina, 1997), share smooth, involute shells resembling the middle Tournaisian gattendorfiid Zadelsdorfia (sensu its typical group around Gattendorfia asiatica Librovitch, 1940). Possible ancestors of the Pericyclidae are the extremely rare and poorly known lower Tournaisian Gattenpleura species of Bartzsch and Weyer (1988) that lack spiral furrows. Clarification of the precise origin of the Pericycloidea requires further discoveries but a different origin of oldest Pericyclidae (Goniocyclus) and Muensteroceratidae (Xinjiangites) is beyond any doubt. Because of this polyphyly, both families cannot be assigned to a single superfamily, as adopted in the Carboniferous/Permian Treatise (Furnish et al. 2009) and later papers (e.g. Korn et al. 2010). The Muensteroceratidae, including the descendent Maxigoniatitidae Korn et al., 1999, are, therefore, excluded from the Pericycloidea. The phylogenetic relationships between the Muensteroceratidae and the oldest (middle upper Tournaisian) Goniatitoidea (the Habadraitinae) are also poorly resolved, but it is unlikely that the latter derived from upper Tournaisian pericyclids. When this is further constrained, the Pericycloidea cannot remain in a monophyletic suborder Goniatitina. This applies to other groups with subdivided external lobes, such as the Karagandoceratidae Librovitch, 1957 and Prodromitidae Arthaber, 1911 (see Becker and Weyer 2004).
The Helicocyclinae n. subfam. comprise three genera that are distinguished by their sutures, especially by the shape of the flank saddles and L-lobes. In terms of shell size, weak and mostly simple ribbing, early serpenticonic, later mostly (sub)evolute and thinly discoidal shell form, and by the reduction of the ventral sinus of the ornament, the subfamily is rather homogeneous.
By comparison, the Pericyclinae Hyatt, 1900 comprise thicker forms with narrow umbilicus of early stages that opens gradually, shallow to deep ventral sinus, and narrow, subparallel E-lobes; bifurcating or intercalated ribbing is more common. The Ammonellipsitinae Riley, 1996 are similar to the Pericyclinae in terms of shell form but differ in their divergent E-lobes and subangular ventrolateral saddles. The poorly defined, large-sized Rotopericyclinae Korn et al., 2010 are also similar to the Pericyclinae. They are characterized by reduced ribbing, low median saddles, and marked ontogenetic opening of the umbilicus. The Hammatocyclinae Ebbighausen et al., 2010 comprise cadiconic species with weak ribbing at the umbilicus; they share with the new subfamily a trend to reduce the ventral sinus. The Temertassetiidae Korn et al., 2010 are characterized by thickly discoidal to pachyconic, weakly ribbed shells with rather narrowly umbilicate early whorls. Their wide, strongly divergent and flexured (lyrate) E-lobes are especially characteristic but there is some homoeomorphy with the shape of the E-lobe in the cadiconic Parahammatocyclus Riley, 1996 (Hammatocyclinae).
Geographic distribution. Northern England (Craven Basin and Peak District), Germany (Harz Mountains), southern Morocco (Tafilalt), southern Algeria (Mouydir), and Russia (northern Urals/Timan).
Stratigraphic range. Lower part of upper Tournaisian (Progoniatites Genozone, Korn et al. 2003) to lower Viséan (Arundian, lower part of Bollandoceras Genozone, Kusina and Konovalova 2004).
Helicocyclus Schindewolf, 1951 (emend.)
Type-species. H. gracillimus Schindewolf, 1951 (monospecific genus).
Diagnosis (emend.). Very small-sized, evolute, depressed (serpenticonic), flanks and venter rounded, with moderately fast expanding whorls, slightly concave, rarely bifurcating, dense ribbing, and regular, concave shell constrictions. Sutures with narrow, subparallel, slightly flexured (lyrate) E-lobe, moderately high median saddle (up to 50% of narrow external saddle), bell-shaped A-lobe, very narrow lateral saddle, short, wide, bell-shaped L-lobe (with incipient tripartition), deep internal U- and I-lobes. Suture formula: (E2E1E2)AL:UI.
Discussion. The suture of H. gracillimus, especially the wide, bell-shaped L-lobe, with an incipient trend to become trifid, and inner flank saddle, differ from sutures in all other named species that have previously been assigned to Helicocyclus. Especially the older Tournaisian forms are, therefore, placed in Serpenticyclus n. gen. The group of H. divergens with distinctively subtriangular suture elements falls in Tectocyclus n. gen.
Geographic distribution. Only known from the Harz Mountains of Germany.
Stratigraphic range. Either top-Tournaisian or lower Viséan (see below).
Helicocyclus gracillimus Schindewolf, 1951 (emend.)
Figures 4d–h, 5a–b, 6b, Table 1
v* 1951 Pericyclus (Helicocyclus) gracillimus Schindewolf, p. 79–81, fig. 22, pl. VI, figs. 2, 3.

Representative Helicocyclinae n. subfam. a–c Holotype of Tectocyclus herbigi n. gen. n. sp., GMM B6C.60.1 (lateral, adoral, and ventral views), × 3. d–f Holotype SW 19 of Helicocyclus gracillimus Schindewolf, 1951 (lateral, adoral, and ventral views), × 3. g –h Paratype SW 18 of Helicocyclus gracillimus Schindewolf, 1951 (lateral and adoral views), × 3

Detailed views of characteristics in Helicocyclinae n. subfam. a Early whorls of holotype of Helicocyclus gracillimus Schindewolf, 1951, SW 19, showing a slightly bulbous (inflated) initial part (protoconch/parts of first whorl). b Early whorls of paratype of Helicocyclus gracillimus Schindewolf, 1951, SW 18, showing the same bulbous initial part. c Early whorls of Tectocyclus herbigi n. gen. n. sp., GMM B6C.60.1, showing a possible primary septum, weakly inflated first whorl, and the position (see arrow) of a secondary varix marking the ammonitella end. d Umbilical region of last septum in Tectocyclus herbigi n. gen. n. sp., GMM B6C.60.1, showing the subangular outer Le-lobe, flat saddle around the umbilical seam, and shallow, internal Li-lobe (see arrow)

Sutures of various Helicocyclinae n. subfam.; scale bars = 1 mm. a Holotype of Serpenticyclus fuscus (Korn et al., 2003) n. gen., at 6.8 mm dm (after Korn et al. 2003, Fig. 24). b Holotype of Helicocyclus gracillimus Schindewolf, 1951, at 11.5 mm dm. c Holotype of Tectocyclus herbigi n. gen. n. sp., GMM B6C.60.1, at 17 mm dm. d Holotype of Tectocyclus divergens (Riley, 1996), at ca. 15 mm dm (after Riley 1996, text-Fig. 45A)
Material. Holotype SW 19 (Fig. 4d–f) and the smaller paratype SW 18 (Fig. 4g–h), collection of originals in the Institute for Geology and Palaeontology, TU Clausthal (very recently transferred to the collection at the Geoscience Centre, Georg-August University Göttingen).
Diagnosis (emend.). Whorls well-rounded, with largest width on the lower flank, evolute (uw/dm = ca. 0.48), discoidal (ww/dm = 0.37), depressed (ww/wh 1.30–1.25 at 8–12 mm dm), moderately fast expanding (WER ca. 1.75), covered by weakly concave, dense, rounded ribs that are wider than the interspaces (ca. 40/half whorl), with two or three constrictions/whorl and associated minor anteriorly lips. M-lobe very short and small, median saddle ca. 37% as high as the narrowly rounded external saddle, total E-lobe ca. half as wide as deep, slightly lanceolate, A-lobe wide, asymmetrically bell-shaped, as deep as E-lobe, lateral saddle narrow, lower than external saddle, L-lobe short, asymmetrically bell-shaped, with incipient tripartition, central part forming a narrow, pointed tip, deep, narrow inner U- and I-lobes.
Description. Schindewolf (1951) gave a good characterization of the species. The conch parameters are refined (Table 1). As noted originally, bifurcation of the dense, rounded ribs is very rare and occurs ca. on the mid-flank. The holotype possesses three constrictions on the last whorl, one of which is poorly visible in Fig. 4d due to abrasion by preparation. They follow the course of the ribbing, which crosses the venter without a sinus (Fig. 4e). The second last constriction, which is the best preserved, shows a characteristic fine lip at the anterior margin. A bulbous (inflated) initial part (protoconch to first whorl) has a diameter of 0.55 mm, the ammonitella size is estimated as ca. 0.85 mm. Sutures have been prepared at the end of the last whorl (Fig. 6b). The suture diagram of Schindewolf (1951) missed the small, short M-lobe and the slightly lanceolate shape of the E-lobe. The median saddle reaches 36.5% of the height of the narrow external saddle. The width/depth ratio of the E-lobe is 57% measured at the narrowest point and ca. 85% measured at the tip of the external saddle. The lateral saddle reaches ca. 80% of the height of the latter. The dorsal suture is visible on the terminal septum of the paratype (Fig. 4h), which proves that the internal U- and I-lobes are narrow and deep. There is no evidence for an additional shallow lobe just inside the umbilical seam. Due to corrosion, the constrictions are only weakly visible on the paratype (Fig. 4g). Its bulbous part of the initial whorl has a diameter of 0.5–0.6 mm (Fig. 5b). Sutures are not visible.
Discussion. The wide, markedly bell-shaped L-lobe, with an incipient trend towards lobe subdivision, distinguishes the type-species of Helicocyclus from all other named helicocyclids. Popov (1968, figs. 21d-e) illustrated an incipient concavity of the inner prong of the lateral saddle in a specimen identified as Fascipericyclus tianchanicus, a species that is here excluded from the Helicocyclinae n. subfam. (see discussion below).
Geographic distribution. Restricted to the type locality at the Iberg, Oberharz region (Germany).
Stratigraphic range. The faunal list of Schindewolf (1951) includes taxa that are either typical for the top-Tournaisian, such as Ammonellipsites kochi (Holzapfel, 1889) and A. hauchecornei (Holzapfel, 1889), or lower Viséan (e.g. nomismoceratid species), and some much younger, upper Viséan taxa (species of Goniatites and Pronorites; compare Mottequin and Weyer 2019). The exposed equivalent of the Erdbach Limestone was 10–12 m thick and probably included the Tournaisian/Viséan boundary. It is currently not possible to specify the type level within this interval.
Serpenticyclus n. gen.
Type-species. Helicocyclus fuscus Korn et al., 2003.
Derivation of name. Based on the serpenticonic shell form in combination with the family name.
Diagnosis. Small-sized, evolute and depressed (serpenticonic), thinly or thickly discoidal, with very weak to moderately strong, sublinear to concave, dense, sometimes bifurcating ribs and regular constrictions. Sutures with low to moderately high median saddle (25–40% of narrowly rounded external saddle), moderately narrow, slightly divergent E-lobe, asymmetric v- or bell-shaped A-lobe that is shorter or as deep as the E-lobe, asymmetrically arched lateral saddle, and wide, short, funnel-shaped L-lobe, sometimes with a weak flexure in the low saddle just outside the umbilical seam; internal U- and I-lobes narrow and deep. Suture formula: (E1E2E1)AL:UI.
Species included.
Helicocyclus formosus Korn et al., 2010: lower part of upper Tournaisian (regional Helicocyclus-Ouaoufilalites Assemblage), Mouydir, southern Algeria
Helicocyclus fuscus Korn et al., 2003: lower part of upper Tournaisian (Progoniatites Genozone), Tafilalt, southern Morocco
Helicocyclus inornatus Korn et al., 2010: lower part of upper Tournaisian (regional Helicocyclus-Ouaoufilalites Assemblage), Mouydir, southern Algeria
Helicocyclus laxaris Korn et al., 2010: lower part of upper Tournaisian (regional Helicocyclus-Ouaoufilalites Assemblage), Mouydir, southern Algeria
Species included with reservation.
Helicocyclus involutus Kusina in Shimankiy and Kusina, 1977: lower Viséan (Chadian, fauna with Michiganites rhipaeus, Kusina and Konovalova 2004), northern Timan
Helicocyclus aberratus Kusina and Konovalova, 2004: lower Viséan (Arundian, Bollandoceras Genozone), northern Timan
Discussion. The new genus differs from Helicocyclus in its simpler, divergent and funnel-shaped, not bell-shaped (incipiently trifid) L-lobe and in the broader, asymmetrically arched lateral saddle (Fig. 6a). Such suture differences are used to separate other Lower Carboniferous genera, e.g. in the Prolecanitidae or in the transition from the Goniatitidae to early Delepinoceratidae (see Furnish et al. 2009).
The North African species form a relatively homogeneous group that is distinctively older than the German Helicocyclus. Viséan questionable Serpenticyclus n. gen. probably fall in different or new genera. For example, ?S. involutus differs in its subinvolute and compressed adult shell form. Its minor flexure of the low subumbilical LU-saddle is also known in S. inornatus (see Korn et al. 2010, fig. 30C). A specimen from high in the Bollandoceras Genozone of the Kozhim River section of the Timan, identified by Kusina and Konovalova (2004) as Gen. nov. A, shows a similar shell form but lacks fine ribbing; its sutures are unknown. The Viséan ?S. aberratus, also from the Kozhim River, represents a very different late side-branch of helicocyclid evolution characterized by an evolute and strongly compressed (planorbiconic) adult shell with markedly concave ribbing and minor ventral sinus. The change from depressed to compressed whorls occurs before 16 mm dm. The introduction of a new genus is currently not advisable since the suture of this species is unknown.
Pericyclus tianshanicus Librovitch, 1927 (= P. evolutus Librovitch, 1927; lectotype of that form selected as specimen II illustrated on pl. IV, figs. 3a-b) from the lower Viséan of Kirgisia (Tien Shan) is another problematical species. Schindewolf (1951) included it together with the co-occuring P. djaprakensis Librovitch, 1927 and P. sonkulensis Librovitch, 1927 in his new subgenus Helicocyclus because of their wide umbilici. However, this assignment was rejected by Riley (1996). Pericyclus tianshanicus reaches much larger size than other helicocyclids (at least 66 mm dm, see Popov 1968), with convex ribbing and a shallow ventral sinus of its mature stage. We exclude from it two forms, the subevolute specimen IV figured by Librovitch, (1927, pl. V, figs. 4a-b) as P. tianchanicus var., and the widely evolute specimen 827/87 with strongly bifurcating ribs figured by Popov (1968, pl. VII, figs. 2a–b). There seems to be considerable variability of the adult cross-section in the typical form, ranging from compressed at ca. 35 mm dm in the lectotype selected by Librovitch (1941, “holotype” re-illustrated on pl. XXXVIII, figs. 3a–b), to weakly depressed at ca. 55 m dm in the largest specimen of Popov (1968, pl. VII, figs. 4a–b) to markedly depressed in the largest evolutus syntype of Librovitch (1927, pl. IV, figs. 2a–b) at ca. 50 mm dm (ww/wh = ca. 1.6). The latter specimen, which also shows convex ribbing, is somewhat intermediate towards the even thicker P. djaprakensis and P. sonkulensis, which Popov (1968) regarded as conspecific (see also Ebbighausen et al. 2010, p. 166). Based on its subacute external saddles, documented by Popov (1968), P. djaprakensis belongs to Ammonellipsites. Popov (1968, Fig. 21c) showed Ammonellipsites-type sutures with low median saddle also in an adult P. tianshanicus. Therefore, we regard P. tianshanicus as an evolute species of Ammonellipsites that is somewhat homoeomorphic to the Helicocyclinae n. subfam. Ammonellipsites is anyway based on an evolute type-species, A. funatus (Sowerby, 1814) (see re-illustration of the holotype in Tilsley and Korn 2009). For the better-known involute A. kayseri-A. kochi Group, the name Kaypericyclus Turner, 1948 is available as subgenus name, which has priority to Eurycyclus Schindewolf, 1951.
Small-sized upper Tournaisian specimens from the Chatkal River of Uzbekistan identified by Konovalova (2015) as H. tianchanicus are not conspecific with the Kirgisian type population. They show relatively strong whorl overlap, weakly biconvex to sublinear ribbing, and convex constrictions, combined with a Serpenticyclus-type suture. They represent a new form that could belong to the Helicocyclinae n. subfam.
Geographic distribution. Southern Morocco (Tafilalt), southern Algeria (Mouydir), and ?Russia (northern Urals/Timan).
Stratigraphic range. Upper Tournaisian, questionably lower Viséan (Chadian/Arundian).
Tectocyclus n. gen.
Type-species. T. herbigi n. sp.
Derivation of name. After the tectiform (subtriangular) shape of the three outer saddles.
Diagnosis. Small-sized, early stages evolute and depressed (serpenticonic), smooth, adult stages > ca. 10 mm dm subevolute, compressed, tegoid, with slowly expanding whorls (WER 1.5–1.6), smooth or with very weak, concave, widely spaced ribs and regular, concave constrictions. Sutures with subtriangular (tectiform) lobes and saddles on the flanks and venter; median lobe very small and short, median saddle roof-shaped, ca. half as high as the narrowly triangular external saddle, E-lobe widely divergent, ca. 1.3 to 1.6 times as wide as the deep, widely v-shaped A-lobe, subtriangular lateral saddle lower than external saddle, short outer L-lobe (sub)angular, divided by low saddle at the umbilical seam, internal L-lobe very shallow, U- and I-lobes narrow and deep. Suture formula: (E2E1E2)ALe:LiUI.
Species included.
Tectocyclus herbigi n. gen. n. sp., probably lower Viséan, southeast Morocco.
Helicocyclus divergens Riley, 1996, lower Viséan (Chadian, faunas with Eonomismoceras and juvenile Michiganites), northern England.
Discussion. The new genus differs by its subtriangular outer lobes and saddles markedly from the two other genera of the Helicocyclinae n. subfam. In addition, it can be distinguished by the very shallow, internal Li-lobe, which occurs at least in the type-species. The subangular external saddle is an iteration to the Ammonellipsitinae, which have narrower E-lobes and thicker shell forms.
Geographic distribution. Northern England (Craven Basin and Peak District), eastern Anti-Atlas (Tafilalt).
Stratigraphic range. Probably restricted to the lower Viséan (Chadian).
Tectocyclus herbigi n. sp.
v 2012 “Helicocyclus” n. sp. aff. divergens Hahn et al., fig. 2.
Derivation of name. In honor of Prof. Dr. H.-G. Herbig (Cologne) for his major contributions to Carboniferous stratigraphy, including research in Morocco.
Material. Only holotype GMM B6C.60.1 (Geomuseum Münster, Westfälische Wilhelms-University).
Type locality. Jebel Begaa region (Figs. 2, 3), southeastern Tafilalt, Morocco.
Type level. Merdani Formation, probably lower Viséan (Chadian).
Diagnosis. Adult whorl (at 17.5 mm dm) subevolute (uw/dm = 0.44), thinly discoidal (ww/dm < 0.25), compressed (ww/wh = ca. 0.75), and slowly expanding (WER = ca. 1.5), tegoid, smooth, with two concave constrictions/whorl. Sutures with very small and shallow M-lobe, tectiform median saddle reaching ca. 55% of the height of the asymmetric, subtriangular external saddle, strongly divergent E-lobe that is 1.6 times wider than deep at the top, wide, v-shaped A-lobe that is slightly shorter than the E-lobe, moderately high, subtriangular lateral saddle, short, subangular Le-lobe, very low, flat secondary lateral saddle at the umbilical seam, very shallow, rounded internal Li-lobe, and deep, narrow U- and I-lobes.
Description. The holotype is a well-preserved, fully septate phragmocone mould that shows well all shell parameters (Fig. 4a–b, Table 1) and the sutures (Fig. 6c), including a terminal septal face (Fig. 4c) that enables a view on the very shallow, internal Li-lobe (Fig. 5d). The median lobe is poorly developed in some of the septa. Measured at the base, the median saddle is 1/3 wider as high. The deep E-lobe is slightly inflexed, with weakly concave flanks of the median saddle, arched lower flanks of the external saddle changing to straight to slightly concave in the upper half. The lateral saddle reaches ca. 73% of the height of the external saddle. Two constrictions are separated by half a whorl distance and do not reach the umbilical wall (Fig. 4a) This may explain why almost no constrictions can be discerned on the exposed parts of the eight inner whorls. The maximum whorl width lies on the lower flanks, giving a tegoid cross-section with narrowly rounded venter (Fig. 4c). Even under oblique light, there are only very weak indications of undulose ribbing (Fig. 4a). Based on the assumed preservation of the laterally concave primary septum, the bowl-shaped protoconch would have a diameter of less than 0.2 mm, which is very small for a goniatite. The subsequent bulbous (inflated) part of the first whorl reaches ca. 0.55 mm dm. As indicated by the position of a secondary varix, the ammonitella diameter is ca. 0.65 mm (Fig. 5c).
Discussion. The English T. divergens is generally similar but can be clearly distinguished based on several features. At comparable size, it is less evolute (uw/dm ca. 0.40), somewhat thicker (ww/dm ca. 0.30), weakly ribbed, and has three constrictions/whorl. The E-lobe is even more asymmetric, with a somewhat stronger concavity of the flanks of the median saddle, the A-lobe becomes deeper than the E-lobe during ontogeny, and the saddle at the umbilical seam is better developed (Fig. 6d). The description of Riley (1996) did not comment on the inner suture, the terminal septal face of the holotype was not illustrated. The high number of whorls at small size and whorl expansion rates are similar in both taxa. Only the divergens cross-section of Riley (1996), not that of Tilsley and Korn (2009), shows a minor inflation of the protoconch/first whorl.
Stratigraphic range. Restricted to the type level.
Geographic distribution. Restricted to the type region.
Palaeobiogeographic aspects
The review of helicocyclid distribution shows that geographical groups correlate with morphological characteristics. In the lower part of the upper Tournaisian, Serpenticyclus n. gen. is currently restricted to the northwestern margin of Gondwana (Morocco, Algeria). From there, a migration towards the northern Variscan Sea must have occurred in the top-Tournaisian, leading to the German Helicocyclus. Another branch may have led to Central Asia (H. tianchanicus sensu Konovalova 2005, non Librovitch). In the lower Viséan (Chadian), the Armorican Terrain Assemblage and southern European terrains within the Variscan Sea obviously did not form an obstacle for faunal exchange, as evidenced by the “pan-Variscan” Tectocyclus n. gen. distribution. Based on the peculiar suture specialization of HelicocycIus, Tectocyclus n. gen. cannot have been derived from the latter. The distribution suggests that the pre-upper Viséan Eovariscan tectonic movements in the Moroccan Meseta had not yet created a palaeogeographic barrier in the lower Viséan. At the same time, the subfamily had migrated into the Urals-Timan seaway, where it survived far in the north into Arundian levels (questionable Serpenticyclus n. gen. taxa) but it had already disappeared from the partly very rich and diverse faunas of North Africa and Europe. The Kozhim River area seems to have been part of a final refugium. The subfamily never managed to migrate through the narrow Afro-Appalachian Seaway into eastern North America, where Polaricyclus and Rotopericyclus are the only upper Tournaisian/lower Viséan pericyclids (Work and Mason 2003, 2004, 2018). It is expected that future records will alter this palaeobiogeographic scenario.
Conclusions
The Pericyclidae originated independently from the Muensteroceratidae. In order to avoid polyphyly of the Pericycloidea, the latter and their descendents are removed from the superfamily, which questions its position within a monophyletic suborder Goniatitina. Helicocyclus and two related new genera comprise the distinctive Helicocyclinae n. subfam. It is characterized by small adult size (a high number of whorls at small size), typically evolute and markedly compressed shells, especially by serpenticonic early whorls, weak ribbing, and a trend to widen and subdivide the L-lobe. Within the new subfamily, Serpenticyclus n. gen. differs from Helicocyclus, which is revised, by very different shapes of the L-lobe and flank saddles, a genus-level criterion in other Carboniferous goniatites. Tectocyclus n. gen. differs from both genera by its strictly subtriangular lobe and saddle shapes and subdivided L-lobe. According to the present record, the subfamily first appeared early in the upper Tournaisian in North Africa, expanded then across the Variscan Sea to the southern shelf of Laurussia, and possibly to Central Asia, and includes youngest Chadian/Arundian representatives of still unsettled generic affinity in the northern Urals-Timan seaway.
Availability of data and materials
All data are provided in the text and figures.
Abbreviations
- dm:
-
diameter
- wh:
-
whorl height
- ww:
-
whorl width
- uw:
-
umbilical width
- ah:
-
apertural height
- WER:
-
whorl expansion rate
- M-lobe:
-
median lobe on the mid-venter
- E-lobe:
-
external or ventral lobe on the outer flanks
- A-lobe:
-
adventitious lobe on the mid-flank
- L-lobes:
-
lateral lobes around the umbilical seam
- U-lobe:
-
umbilical lobe
- I-lobe:
-
inner or dorsal lobe
- median saddle:
-
saddle between the E-lobes on the mid-venter
- external saddle:
-
ventrolateral saddle between E- and A-lobes
- lateral saddle:
-
saddle between A- and L-lobes
- A.:
-
Ammonellipsites
- G. :
-
Goniocyclus
- H. :
-
Helicocyclus
- P. :
-
Pericyclus
- S. :
-
Serpenticyclus n. gen.
- T. :
-
Tectocyclus n. gen
References
Alvaro, J.-J., M. Aretz, M. Benharref, M. Hibti, A. Pouclet, H. El Hadi, A. Koukaya, E.M. Ettachfini, and L. Boudad. 2014. Carte géologique du Maroc au 1/50 000, Feuille Tawz, Mémoire explicativ. Notes et Mémoires du Service Géologique 551bis: 1–123.
Aretz, M., J. Denayer, and B. Mottequin. 2013. Preliminary data on Viséan (Carboniferous) corals and brachiopods from the strata between the Djebel Begaa and the Gara el Itima (Eastern Tafilalt, Morocco). In International Field Symposium “The Devonian and Lower Carboniferous of northern Gondwana”, Field Guidebook, eds. R.T. Becker, A. El Hassani, and A. Tahiri. Document de l´Institut Scientifique Rabat 27: 87–94.
Aretz, M., H.-G. Herbig, X.-D. Wang, F.M. Gradstein, F.P. Agterberg, and J.G. Ogg. 2020. The Carboniferous Period. In Geological Time Scale 2020, Volume 2, pp. 811–874, eds. F.M. Gradstein, J.G. Ogg, M.D. Schmitz, and G.M. Ogg. Amsterdam: Elsevier.
Bartzsch, K., and D. Weyer. 1988. Neue Gattenpleura-Funde aus dem Untertournai des Saxothuringikums (Ammonoidea, Unterkarbon). Hallesches Jahrbuch für Geowissenschaften 13: 37–48.
Becker, R.T., A. El Hassani, Z.S. Aboussalam, S. Hartenfels, and L. Baidder. 2018a. The Devonian and Lower Carboniferous of the eastern Anti-Atlas: Introduction to a “cephalopod paradise”. Münstersche Forschungen zur Geologie und Paläontologie 110: 145–157.
Becker, R.T., S. Hartenfels, C. Klug, Z.S. Aboussalam, and L. Afhüppe. 2018b. The cephalopod-rich Famennian and Tournaisian of the Aguelmous Syncline (southern Maïder). Münstersche Forschungen zur Geologie und Paläontologie 110: 273–306.
Becker, R.T., and D. Weyer. 2004. Bartzschiceras n. gen. (Ammonoidea) from the Lower Tournaisian of Southern France. Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 88: 11–36.
Becker, R.T., S.I. Kaiser, and Z.S. Aboussalam. 2006. The Lower Alum Shale Event (Middle Tournaisian) in Morocco – facies and faunal changes. In Carboniferous Conference Cologne, From Platform to Basin, September 4–10, 2006, Program and Abstracts, eds. M. Aretz and H.-G. Herbig. Kölner Forum für Geologie und Paläontologie 15: 7–8.
Becker, R.T. 2007. The revised global Middle Tournaisian to Middle Viséan ammonoid biostratigraphy. In XVI International Congress on the Carboniferous and Permian, Abstracts, eds. Wang, Y., Zhang, H., and Wang, X. Journal of Stratigraphy, 31 (Suppl. 1): 45–46.
Becker, R.T. 2019. New Lower Carboniferous ammonoid faunas from the eastern Anti-Atlas, Southern Morocco. In 19th International Congress on the Carboniferous and Permian, Cologne, July 29-August, 2. 2019, Abstracts, eds. S. Hartenfels, H.-G. Herbig, M.R.W. Amler, and M. Aretz. Kölner Forum für Geologie und Paläontologie 23: 40–41.
Boardman, D.R. II, T.L. Thompson, C. Godwin, S.J. Mazzullo, B.W. White, and B.T. Morris. 2013. High-Resolution Conodont Zonation for Kinderhookian (Middle Tournaisian) and Osagean (Upper Tournaisian-Lower Viséan) Strata of the Western Edge of the Ozark Plateau, North America. Shale Shaker, September-October 2013: 98-141.
Bockwinkel, J., and V. Ebbighausen. 2006. A new ammonoid fauna from the Gattendorfia-Eocanites Genozone of the Anti-Atlas (Early Carboniferous; Morocco). Fossil Record 9 (1): 87–129.
Bockwinkel, J., R.T. Becker, and V. Ebbighausen. 2013. Late Givetian ammonoids from Hassi Nebech (Tafilalt Basin, Anti-Atlas, southern Morocco). Fossil Record 16 (1): 5–65 and online supplement 1–58.
Clariond, L. 1934. Sur le Dévonien du Tafilalelt et du Maïder. Compte Rendu Sommaire des Séances de la Société Géologique de France 1934 (13): 189–190.
Clariond, L. 1954. Resume des resultats scientifiques de l´excursion C36 apport fourni par les discussions. In Compte rendu des excursions au Maroc, ed. P. Morin. Congrès Géologique International, XIXe Session, Alger 1952, Série Maroc 16: 62.
Delepine, G. 1939. La discordance Viséan sur Famennien près Taouz (Confins algéro-marocaina du Sud). Bulletin de la Société Géologique de France 9 (5): 97–104, pl. 7.
Delepine, G. 1941. Les goniatites du Carbonifère du Maroc et des confins Algéro-Marocains du sud (Dinantien-Westphalien). Notes et Mémoires, Service Géologique, Division des Mines et de la Géologie 56: 1–111, pls. 1–8.
Ebbighausen, V., and J. Bockwinkel. 2007. Tournaisian (early Carboniferous/Mississippian) ammonoids from the Ma`der Basin (Anti-Atlas, Morocco). Fossil Record 10 (2): 125–163.
Ebbighausen, V., D. Korn, and J. Bockwinkel. 2010. The ammonoids from the Dalle à Merocanites of Timimoun (Late Tournaisian-Early Viséan; Gourara, Algeria). Fossil Record 13 (1): 153–202.
Furnish, W.M., B.F. Glenister, J. Kullmann, and Z. Zhou 2009. Carboniferous and Permian Ammonoidea (Goniatitida and Prolecanitida). Treatise on Invertebrate Paleontology, Part L, Mollusca 4 (Revised): 1–258. Lawrence: The University of Kansas Paleontological Institute.
Hahn, G., P. Müller, and R.T. Becker. 2012. Unter-karbonische Trilobiten aus dem Anti-Atlas (S-Marokko). Geologica et Palaeontologica 44: 37–74.
Hance, L., E. Poty, and F.-X. Devuyst. 2006. Ivorian. Geologica Belgica 9 (1/2): 117–122.
Hollard, H., and P. Jaquemont. 1956. Note sur l´age de la serie de la Betaïna (vallée du Dra, Sud Marocain). Comptes rendus des séances de l´Academie des Sciences 242: 2651–2654.
Hollard, H. 1960. Extension du Tournaisien dans la région de Taouz et dans le Sud du Tafilalt (Maroc). Comptes rendus des séances de l´Academie des Sciences 250 (4): 733–735.
Hollard, H. 1970. Sur la transgression dinantienne au Maroc présaharien. Compte Rendu 6e Congrès International de Stratigraphie et Géologie du Carbonifère, Sheffield 1967 III: 923–936.
Holzapfel, E. 1889. Die Cephalopoden-führenden Kalke des unteren Carbon von Erdbach-Breitscheid bei Herborn. Paläontologische Abhandlungen, Neue Folge 1 (1): 1–74.
Hyatt, A. 1900. Tetrabranchiate Cephalopoda. In Text-book of Palaeontology, volume 1, pp. 502–604, ed. K.B. Zittel. New York: McMillan.
Kaiser, S.I., R.T. Becker, S. Hartenfels, and Z.S. Aboussalam. 2013. Middle Famennian to middle Tournaisian stratigraphy at El Atrous (Amessoui Syncline, southern Tafilalt). In International Field Symposium “The Devonian and Lower Carboniferous of northern Gondwana”, Field Guidebook, eds. R.T. Becker, A. El Hassani, and A. Tahiri. Document de l´Institut Scientifique Rabat 27: 77–86.
Kaiser, S.I., R.T. Becker, S. Hartenfels, and Z.S. Aboussalam. 2018. The global Hangenberg Crisis and Lower Alum Shale Event at El Atrous (southern Tafilalt Platform). Münstersche Forschungen zur Geologie und Paläontologie 110(online): 327–338.
Klug, C., S. Döring, D. Korn, and V. Ebbighausen. 2006. The Viséan sedimentary succession at the Gara el Itima (Anti-Atlas, Morocco) and its ammonoid faunas. Fossil Record 9 (1): 3–60.
Klug, C., K. de Baets, and D. Korn. 2016. Exploring the limits of morphospace: ontogeny and ecology of late Viséan ammonoids from the Tafilalt, Morocco. Acta Palaeontologica Polonica 61 (1): 1–14.
Konovalova, V. 2015. Early Carboniferous Ammonoids from the Chatkal river Basin middle Tien Shan Uzbekistan. Paleontologicheskiy Zhurnal 2015 (4): 33–46 (in Russian with English summary).
Korn, D. 2006. Ammonoideen. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften 41: 147–170.
Korn, D., C. Klug, and R. Mapes. 1999. Viséan and Early Namurian Ammonoids from the Tafilalt (Eastern Anti-Atlas, Morocco). Abhandlungen der Geologischen Bundesanstalt 54: 345–375.
Korn, D., C. Klug, V. Ebbighausen, and J. Bockwinkel. 2002. Palaeogeographical meaning of a Middle Tournaisian ammonoid fauna from Morocco. Geologica et Palaeontologica 36: 79–86.
Korn, D., J. Bockwinkel, V. Ebbighausen, and C. Klug. 2003. Palaeobiogeographic and evolutionary meaning of an early Late Tournaisian ammonoid fauna from the Tafilalt of Morocco. Acta Palaeontologica Polonica 48 (1): 71–92.
Korn, D., J. Bockwinkel, and V. Ebbighausen. 2007. Tournaisian and Viséan ammonoid stratigraphy in North Africa. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 243 (2): 127–148.
Korn, D., J. Bockwinkel, and V. Ebbighausen. 2010. The ammonoids from the Argiles de Tegentour of Oued Temertasset (early Late Tournaisian; Mouydir, Algeria). Fossil Record 13 (1): 35–152.
Kusina, L., and V. Konovalova. 2004. Novye ammonoidei iz pogranichnykh Turneysko-Viseyskikh otlozhenii na reke Kozhim (Pripolyanyi Ural). Paleontologicheskiy Zhurnal 2004 (1): 10–18.
Lane, H.R., and P.L. Brenckle. 2005. Type Mississippian subdivisions and biostratigraphic succession. In Stratigraphy and Biostratigraphy of the Mississippian Subsystem (Carboniferous System) in Its Type Region, the Mississippi River Valley of Illinois, Missouri, and Iowa, ed. P.H. Heckel. Guidebook for Field Conference, Subcommission on Carboniferous Stratigraphy, September 8–13, 2001: 83–107.
Librovitch, L.S. 1927. Nizhnekamennougolnye golovonogie iz rayona ozera Son-kul (Tyan Shan). Materialy po obschey i prikladnoy geologii, Geologichesky Komitet 74: 1–55, pls. 1–7.
Librovitch, L.S. 1940. Carboniferous ammonoids of North Kazakhstan. Paleontology of USSR 4 (9.1): 1–395.
Librovitch, L.S. (ed.) 1941. The Atlas of the guide forms of the fossil faunas of the USSR, Volume IV, Lower Carboniferous. Moscow, Leningrad: State Editorial Office for Geological Literature, Committee for Geology at the Sovnarkom of the USSR [in Russian].
Librovitch, L.S. 1957. O nekotorykh novykh gruppakh goniatitov iz kamennougol´nykh otlozheniy SSSR. Ezhegodnik vesiuznogo paleontologicheskogo obshchestva 16: 246–273, 4 pls.
Miller, A.K., and C. Collinson. 1951 Lower Mississippian ammonoids of Missouri. Journal of Paleontology 25 (4): 454–487, pls. 68–71.
Mottequin, B., and D. Weyer. 2019. On some Mississippian (Carboniferous) brachiopods from Neptunian dykes of the Hart Mountains (central Germany). Palaeobiodiversity and Palaeoenvironments
Popov, A.V., and L.F. Kusina. 1997. The Earliest Goniatitina (Ammonoidea) from the South Urals. Paleontological Journal 31 (1): 28–34.
Popov, A.V. 1968. Vizeyskie ammonoidei cevernogo Tyan-Shanya i ikh stratigraficheskoe zhachenie. Trudy Geologicheskiy Instituta, Kirgizkiy Filal Academiya Nauk SSSR: 1–116, pls. 1–8.
Poty, E., and L. Hance. 2006. Livian. Geologica Belgica 9 (1/2): 133–138.
Riley, N.J. 1996. Mid-Dinantian ammonoids from the Craven Basin, north-west England. Special Papers in Palaeontology 53: 1–87.
Ruan, Y.-P. 1995. Tournaisian ammonoids of Northern Xinjiang, China. Palaeontologica Cathayana 6: 407–430.
Schindewolf, O.H. 1951. Über ein neues Vorkommen unterkarbonischer Pericyclus-Schichten im Oberharz. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 93 (1): 23–116, pls. 3–7.
Shimansky, V.N., and L. Kusina. 1977. Rannekamennougolnye golovonogie pripolyatnogo Urala. Byulleten ´Moskovskogo Obshchestva Ispytatelei Prirody, Otdel Geologicheskiy 52 (6): 79–90.
Sowerby, J. 1812–1814. The mineral conchology of Great Britain; or coloured figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the earth, 1: 1–234. London.
Tilsley, J.W., and D. Korn. 2009. Chadian (Tournaisian – Viséan, Carboniferous) ammonoids from the Milldale Limestone Formation of the southern Peak District, England. Proceedings of the Yorkshire Geological Society 57 (3/4): 217–234.
Turner, S. 1948. Mid-Dinantian reef limestones of Dublin and Cork. Transactions of the Leeds Geological Association 6 (2): 44–56.
von Arthaber, G. 1911. Die Trias von Albanien. Beiträge zur Paläontologie und Geologie Österreich und Ungarns 24: 169–277.
Waters, C.N., I.D. Somerville, N.S. Jones, C.J. Cleal, J.D. Collinson, R.A. Waters, B.M. Besley, M.T. Dean, M.H. Stephenson, J.R. Davies, E.C. Freshney, D.I. Jackson, W.I. Mitchell, J.H. Powell, W.J. Barclay, M.A.E. Browne, B.E. Leveridge, S.L. Long, and D. McLean. 2011. A revised correlation of Carboniferous rocks in the British Isles. Geological Society Special Report 26: 1–186.
Wendt, J., B. Kaufmann, and Z. Belka. 2001. An exhumed Palaeozoic underwater scenery: The Visean mud mounds of the eastern Anti-Atlas (Morocco). Sedimentary Geology 145: 215–233.
Work, D.M., and C.E. Mason. 2003. Mississippian (middle Osagean) ammonoids from the Nada Member of the Borden Formation Kentucky. Journal of Paleontology 77 (3): 593–596.
Work, D.M., and C.E. Mason. 2004. Mississippian (late Osagean) ammonoids from the New Providence Shale Member of the Borden Formation, north-central Kentucky. Journal of Paleontology 78 (6): 1128–1137.
Work, D.M., and C.E. Mason. 2018. The Mississippian (late Tournaisian) ammonoid Rotopericyclus from the Borden Formation, Kentucky. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 289 (3): 311–323.
Acknowledgements
This contribution is dedicated to the life-time scientific achievements of Hans-Georg Herbig (University of Cologne). Zhor Sarah Aboussalam (Münster), Jürgen Bockwinkel (Leverkusen), the late Volker Ebbighausen, Till Söte, and Phillip Herbers (both Münster) accompanied the author in the Begaa/Hassi Nebech region of the Anti-Atlas. Traudel Fährenkemper (Münster) assisted in the production of the figures. Svetlana Nikolaeva (London) helped to translate some Russian literature. I am indebted to the helpful reviews by Dieter Weyer (Berlin) and by an anonymous reviewer.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Sven Hartenfels.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Becker, R. A unique pericyclid from the Viséan of the eastern Anti-Atlas (Morocco) and other Helicocyclinae n. subfam. (Goniatitida). PalZ 97, 807–819 (2023). https://doi.org/10.1007/s12542-023-00655-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-023-00655-9