
Abstract
Re-examination of type material and abundant specimens enable the reinstatement of ‘Conocardium hainense Maurer, 1885’, a species erroneously synonymised with Babinicardia clathrata by Rogalla and Amler. Based on the completely reconstructed morphology, C. hainense can be assigned to the genus Bohemicardia within the Hippocardiidae of the Hippocardioidea. About 600 specimens from a single locality available for study including a wide range of ontogenetic stages allow documentation of allometric growth, with shell width increasing faster than shell height. Protoconchs are not preserved, leaving the development from larval stages to early juveniles unrecorded. The large number of specimens confirms a rather narrow range of morphological variation within the taxon. Based on the preservation of the rarely observed external shell layer, conclusions on shell shape, presence of a hood and external shell ornamentation of the taxon are possible. The material confirms that recognising different shell layer architecture in conocardiid rostroconchs is a prerequisite for specific identification and basic taxonomy.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rostroconchs generally played a minor role in Palaeozoic fossil assemblages and mostly occur in small numbers, if compared, e.g. with brachiopods. Hence, taxonomic concepts are often based on very few specimens, and many species are monotypic. This hampers comprehensive characterisation of intraspecific variability and meaningful morphometry. Moreover, characters vital for identification are often inadequately preserved (Rogalla and Amler 2007).
The present study is based on several hundred specimens collected in the early twentieth century from only a few localities of the ‘Stringocephalenschichten’ (upper Middle Devonian) of the Eifel Hills, the Bergisches Land and Hesse (Germany) (Fig. 1). The specimens had either been assigned to Conocardium hainense Maurer or were simply determined as Conocardium sp. and remained as yet unstudied. The taxon Bohemicardia hainense was established by Maurer (1885: 230) as ‘Conocardium hainense’ and first revised by Beushausen (1895: 390). Taxonomical errors caused by Rogalla and Amler (2006c) due to presumed missing type material are discussed below.

Sketch map of the Rhenish Massif, central Europe, showing outcrops of Middle Devonian reefs (black areas) and localities of the Bohemicardia samples studied herein. 1, localities near Bergisch Gladbach, Bergisches Land; 2, Girzenberg near Sötenich, Eifel Hills; 3, Villmar, near Limburg (Lahn), Lahn Syncline; 4, ‘Haina(u)’ mine near Waldgirmes, Gießen, Lahn Syncline, type locality of Bohemicardia hainense (Maurer). Based on Burchette (1981, modified)
For rostroconch studies, particularly on ontogeny and variability, abundant material from a single layer is particularly welcome. In this respect, the present contribution provides data not available from most other localities, as there is no previous study on rostroconchs that is based on such a lot of material. This dataset is one of the largest ones of a single rostroconch species and, thus, not only offers data for a comprehensive description of the taxon but also unique insights into intraspecific variability and post-larval growth trends. Our study aims at describing, characterising and interpreting the variability and the ontogeny of a single species of hippocardiid rostroconchs based on a reliable number of specimens in order to gain basic knowledge about the ontogenetic development of the family or even superfamily.
The history of research on the order Conocardiida Neumayr and the superfamilies Conocardioidea Miller and Hippocardioidea Pojeta and Runnegar was compiled and outlined in detail in Amler and Rogalla (2004: 318), and for an overview on the general morphology of conocardiids, we refer to Schröder-Rogalla (2005) as well as Amler and Rogalla (2007, 2013).
Geological background and stratigraphy
All specimens used for this study come from the Middle Devonian (middle to upper Givetian) of the Eifel Hills, the Bergisches Land and the Lahn Syncline in west-central Germany (Fig. 1).
The sedimentary succession of the upper Middle Devonian (Givetian) in the Eifel region mostly consists of limestones and marlstones. The majority of the studied specimens was collected from heaps of quarried material of the ‘Obere Stringocephalenschichten’ at the Girzenberg (Girzen Hill) near Sötenich, Sötenich Syncline, Eifel Hills, Germany. Schmidt (1936) and later authors were not able to place precisely the general locality name ‘Girzenberg’ cited by Beushausen (1895). Thus, the precise stratigraphic level that yielded the rostroconchs is unknown. Quiring (1914) described the marlstones and marly limestones of the Girzenberg as ‘Girzenbergschichten’, i.e. Girzenberg beds, and Ritz (1931) assigned the locality to the upper Middle Devonian. Schmidt (1936: 315) called this fossiliferous part of the middle to upper Givetian succession ‘Girzenberg fauna’ without definite stratigraphical position. Today, the rocks quarried at the Girzenberg are assigned to the ‘Rodert Schichten’ and ‘Kerpener Schichten’ or Rodert and Kerpen formations, respectively (Ribbert 1985; Struve et al. 2008; STD 2016; Litholex 2021) (Fig. 2).

Adopted from STD (2016)
Devonian stratigraphy of the Bohemicardia sample localities. The most likely age of the studied taxon is marked in red.
Further material was collected from the middle to upper Givetian ‘Obere Stringocephalen Kalk’, ‘Torringen Schichten’ and ‘Büchel Schichten’ of the Bergisches Land (Rhenish Massif). As yet, these strata have not been formalised (STD 2016; Litholex 2021; Fig. 2), but are regarded as equivalents of the above-mentioned limestones of the Sötenich Syncline (Jux 2008).
The original locality of Maurer (1885), now type locality of Bohemicardia hainense, was given as ‘Grube Haina(u)’ where limestones of Middle Devonian age (‘Kalke von Waldgirmes’) were quarried. Kegel (1933) placed the limestones from Haina in the lower part of the ‘Stringocephalen-Stufe’. Later, the term ‘Massenkalk’ was introduced for these rocks and is still in use as ‘Massenkalk Formation’ (Bender 2008; Clausen and Korn 2008; Flick and Nesbor 2021), which, however, is not a formal unit. The precise age of these limestones is still unknown, although these units are placed in the middle to upper Givetian (or even lowermost Frasnian) (Fig. 2).
From a palaeogeographical viewpoint, all localities were situated on the passive continental margin of southeastern Laurussia and adjacent submarine volcanic rises (Grabert 1998; Clausen and Korn 2008). While differing opinions exist on the precise latitude (Golonka 2002; Blakey 2021; Scotese 2021), the study region was certainly situated in the equatorial realm. Due to the rapid sea level rise during the Givetian (Johnson et al. 1985), the palaeogeography of central Europe was characterised by widespread carbonate shelf areas with local reef complexes (Fig. 1). The Eifel region in particular formed a patchwork of different tropical, shallow marine, reefal and lagoonal environments, which today are represented by a great variety of different rock facies (Grabert 1998; Struve et al. 2008). These ecological conditions favoured a flourishing development of rostroconch diversity and abundance (see also Rogalla and Amler 2006a, b, c).
Material and methods
Material
The present data pool consists of about 600 specimens; ca. 120 are embedded in the matrix and ca. 480 are isolated from the sediment. The size of the specimens at hand ranges from 2 to 15 mm (excluding the rostrum; Fig. 3), none of them has the larval shell preserved. Some 100 specimens show the outer shell layer altered to a whitish, soft, chalky matter and variable abrasion stages; in further ca. 120 specimens remains of the outer shell layer are still visible between the ribs of the middle shell layer or preserved only in patches (Figs. 4a, c, 7, 8, 11). In the remaining specimens, only the ornamentation of the middle shell layer is clearly visible (e.g. Figs. 8f–j, 11a–e). Roughly 300 specimens are damaged, 200 of which are variably crushed. In nearly 350 specimens, the base of the rostrum is preserved, rarely more (e.g. Figs. 7a, f, k, 8f, 11a, f, k). The fragile hood is preserved in very few specimens (Fig. 4a), but the lunulazone is well preserved in most of the specimens. Furthermore, the steinkern formerly filling the orifice is preserved in a few specimens, precisely indicating the former presence of a hood (Figs. 4b, 7f).


Morphological details of Bohemicardia hainense (Maurer, 1885), upper Middle Devonian of Girzenberg (Girzen Hill) near Sötenich, Eifel Hills, district Euskirchen, Germany (a, b) and building pit August Kierspel-Straße, (Auf der) Hand, Bergisch Gladbach, Germany (c). a Right lateral view; preserved white outer shell layer hiding radial ornamentation of the middle shell layer; cross section of the short hood clearly visible composed of white outer shell layer and situated on lunulazone; MB.Ro32.20, coll. Dohm. b Oblique posterior view showing ventral part of the rostral area surrounded by the lunulazone and steinkern filling the (ventral) orifice; MB.Ro32.3, coll. Dohm. c Left lateral view; preserved white outer shell layer with internal commarginal growth lines covering most of the radial ornamentation of the middle layer, which is visible in the umbonal region; MB.Ro48.1, coll. Ebbighausen. Scale bars 1 mm
In addition to the type material, for measurements, description and analyses, 50 well-preserved specimens were selected individually in order to cover different preservation modes and growth stages (Table 1). Originally, all specimens of our study had been determined as Conocardium sp. or Conocardium hainense, respectively. Some specimens labelled as Conocardium clathratum (Archiac and Verneuil, 1842) were used for comparison and turned out to be incorrectly determined.
The studied material originates from the following localities (Fig. 1):
-
Girzenberg near Sötenich, Kall, Eifel Hills, North Rhein-Westphalia (colls. Holzapfel 1899, Schießberg 1912, Dohm 1913); ca. N 50.522650 E 6.577500 (Fig. 1 [2]).
-
Temporary quarry ‘Rosenthaler Weg’, Herrenstrunden, Bergisch Gladbach, Bergisches Land, North Rhein-Westphalia (colls. Ebbighausen, H.M. Weber); ca. N 51.008300 E 7.180670 (Fig. 1 [1]).
-
Büchel, Herrenstrunden, Bergisch Gladbach, Bergisches Land, North Rhein-Westphalia (coll. Fliegel); ca. N 51.009450 E 7.173020 (Fig. 1 [1]).
-
Building pit August Kierspel-Straße, (Auf der) Hand, Bergisch Gladbach, North Rhein-Westphalia (coll. Ebbighausen); ca. N 50.993050 E 7.096600 (Fig. 1 [1]).
-
‘Haina(u)’ mine near Waldgirmes, Gießen, Hesse (colls. Maurer 1885; Beushausen 1895; Kayser 1900); N 50.616767 E 8.559490 (Fig. 1 [4]).
-
Villmar, near Limburg (Lahn), Hesse; ca. N 50.396090 E 8.184267 (Fig. 1 [3]).
Methods
The material was studied with conventional methods, i.e. analyses with microscope, measuring tools and basic statistics (Excel). Photographs were taken with the digital microscope VHX-500F (Keyence) and a Leica MC 170 HD, and all measurements by use of a measuring tool provided by the respective microscope software.
Based on Amler (1986), Richter and Amler (1994), Gierse (2003) and Schröder-Rogalla (2005), the following measurements were taken (Fig. 3): Lengthumbo (LU) refers to the length from the anterior end of the snout to the umbo parallel to the dorsal axis. The lengthposterior (LP) indicates the whole length of the rostroconch from the anterior margin to the posterior end of the rostral area (posterior margin) except the length of the rostrum. This is the basic measurement of the length for comparisons. The length of the whole specimen including the length of the rostrum (LR) is called total length (Ltotal). Due to mostly incomplete preservation of the rostrum, LR and Ltotal can rarely be measured. In well-preserved specimens, the diameter of the rostrum (DR) can be measured near the rostral area at the base of the rostrum. Furthermore, the angle α (alpha) can be measured between the vertical axis and the dorsoventral keel or ridge marking the attachment of the hood as indicated by the lunulazone, giving a ratio from the rostral area to the whole specimen. The width (W) indicates the thickness of the specimen. The height (H) is measured from the umbo to the most ventral point of the ventral margin perpendicular to the dorsal axis. The ratio between the total height (H) and the height from the umbo to the orifice (Hori), i.e. the rostral height, plays a major role for separation the superfamilies. In this study, this relation is used with reservation, as it is adulterated by abrasion and loss of the outer shell layer.
For details of the general palaeobiology and morphology of conocardiid rostroconchs used for the descriptions, we refer to Amler (1996, 2016, 2018), Rogalla et al. (2002, 2003), Schröder-Rogalla (2005), Amler and Rogalla (2013) and Amler et al. (2020).
Systematic palaeontology
Repository. Most of the specimens come from the Museum für Naturkunde der Humboldt-Universität zu Berlin, Germany (prefix MB.Ro.). A few specimens originate from the former collection of the Institut für Geologie und Paläontologie der Philipps-Universität Marburg and are now housed at the Senckenberg Forschungsinstitut und Naturmuseum, Frankfurt am Main, Germany (prefix SMF.Mbg.). The syntypes of the Maurer collection are housed at the Naturkundemuseum Darmstadt (prefix HLMD.MR.). The two presumed hypotypes of Beushausen (1895) are from the Beushausen collection of the Goldfuß Museum, Bonn, Germany (IPB beushausen 5, 6). A syntype of Bohemicardia rotunda (Paeckelmann) (SMF.Mbg.3362) and two specimens of Conocardium clathratum belong to the collections of the Senckenberg Forschungsinstitut und Naturmuseum Frankfurt (prefix SMF.).
Nomenclatural remark. In her unpublished PhD thesis (ICZN 1999, Article 8.5.), Schröder-Rogalla (2005) erected numerous new taxa, which were subsequently made available by Rogalla and Amler (2006a, b, c) and Rogalla (2007).
Class Rostroconchia Pojeta, Runnegar, Morris and Newell, 1972.
Order Conocardiida Neumayr, 1891
Superfamily Hippocardioidea Pojeta and Runnegar, 1976.
Family Hippocardiidae Pojeta and Runnegar, 1976.
Subfamily Bohemicardiinae Rogalla and Amler, 2006 [2006c].
Type genus. Bohemicardia Rogalla and Amler, 2006 [2006c].
Additionally included genera. Filicardia Rogalla and Amler, 2006 [2006c] and Globocardia Rogalla and Amler, 2006 [2006c].
Diagnosis. (emended from Rogalla and Amler 2006c, p. 354). Conch opisthocline, i.e. dorsoventral keel and rostrum meeting at acute angles; body carina not developed; rostral area occupying 75–90% of total height.
Genus Bohemicardia Rogalla and Amler, 2006 [2006c].
Type species. Conocardium bohemicum Barrande, 1881 (OD).
Additionally included species. Bohemicardia hainense (Maurer, 1885) and Bohemicardia rotunda (Paeckelmann, 1913).
Diagnosis (emended from Rogalla and Amler 2006c, p. 354). Conch bohemicardiine; snout triangular to nearly nasute in shape, snout rib narrow, snout gape drop-shaped in outline, marginal denticles developed, ventral snout margin diagonal to slightly concave; ventral margin of central body convex to narrowly convex, paralleling dorsal axis; rostral area gently concave, longitudinally ovate, cordate ovate or cordate circular with diagonal rostral keel; surface of conch ornamented with flattened radial costae.
Bohemicardia hainense (Maurer, 1885)
Figures 4, 5, 6, 7, 8, 10, 11, Table 1

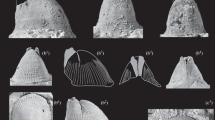
Bohemicardia hainense (Maurer, 1885). a, b Original illustrations of Maurer (1885, pl. 9, figure 21, 21a). c Presumed original of Fig. 5a, left lateral view, fragmented, HLMD.MR.11132, coll. Maurer. d–f Presumed original of Fig. 5b, slightly distorted and fragmented, in posterior (d), left lateral (e) and ventral (f) view, lectotype HLMD.MR.11135, coll. Maurer. Scale bar 1 mm

Bohemicardia hainense (Maurer, 1885). a–d Original illustrations of Beushausen (1895, pl. 29, fig. 2a–d). e–j Specimen IPB beushausen 5-1 from Sötenich, Eifel Hills, district Euskirchen, Germany, labelled as Conocardium hainense var. impressa Beushausen, 1895, but not matching the respective illustration (k–n) in right lateral (e), left lateral (f), ventral (g), dorsal (h), anterior (i) and posterior (j) view. k–n Original illustration of Conocardium hainense var. impressa Beushausen, 1895 (pl. 29, fig. 3a–d). Scale bar 1 mm

Bohemicardia hainense (Maurer, 1885) from the upper Middle Devonian of Girzenberg (Girzen Hill) near Sötenich, Eifel Hills, district Euskirchen, Germany, in different preservation. Note the variable outline caused by differing preservation of the outer shell layer. a–e MB.Ro32.2 in left lateral (a), dorsal (b), ventral (c), posterior (d) and anterior (e) view. f–j MB.Ro32.3 in left lateral (f), dorsal (g), ventral (h), posterior (i) and anterior (j) view. k–o MB.Ro32.6 in left lateral (k), dorsal (l), ventral (m), posterior (n) and anterior (o) view. Scale bars 1 mm

Bohemicardia hainense (Maurer, 1885) from the upper Middle Devonian of Girzenberg (Girzen Hill) near Sötenich, Eifel Hills, district Euskirchen, Germany, in different preservation. Note the variable outline caused by differing preservation of the outer shell layer and the different proportions in juvenile specimens. a–e MB.Ro32.7 in left lateral (a), dorsal (b), ventral (c), posterior (d) and anterior (e) view. f–j MB.Ro32.18 in left lateral (f), dorsal (g), ventral (h), posterior (i) and anterior (j) view. k–o SMF.Mbg.4999 in left lateral (k), dorsal (l), ventral (m), posterior (n) and anterior (o) view. Scale bars 1 mm
*1885 Conocardium Hainense Maurer: 229–230, pl. 9, fig. 21, 21a.
pt 1885 Conocardium Villmarense d’Arch. et de Vern.–Maurer: 226–227, pl. 9, fig. 19, 19a [additional material excluded].
1895 Conocardium hainense Maurer.–Holzapfel: 232.
pt 1895 Conocardium hainense Maur.–Beushausen: 389, pl. 29, fig. 2, 2a–e. [non pl. 29, fig. 1, 1a–c].
1895 Conocardium hainense var. impressa Beushausen: 389–390, pl. 29, fig. 3, 3a–e.
non 1895 Conocardium n. sp. aff. hainense Beushausen: 390–391, pl. 29, fig. 4, 4a–c.
pt 1896 Conocardium Bronn.–Beyer: 79 [all species of Maurer and Beushausen summarised].
2005 Conocardium hainense hainense Maurer, 1885.–Schröder-Rogalla: 105.
2005 Conocardium hainense var. impressa Beushausen, 1895.–Schröder-Rogalla: 106.
pt 2006c Babinicardia clathrata (Archiac and Verneuil, 1842).–Rogalla and Amler: 348.
2006d Conocardium hainense Maurer, 1885.–Rogalla and Amler: 37.
2006d Conocardium hainense var. impressa Beushausen, 1895.–Rogalla and Amler: 37.
Type material and lectotype. Maurer (1885) did not indicate the number of specimens he had available for his description. Four syntypes were located in the Hessisches Landesmuseum für Naturkunde, Darmstadt. HLMD.MR.11132 is a left semi-conch with rock matrix, presumably figured in Maurer (1885, pl. 9, fig. 21; here Fig. 5a, c). HLMD.MR.11135 is a fragmentary, distorted individual and here regarded as the specimen figured by Maurer (1885) on plate 9, figure 21a (Fig. 5b, d–f). HLMD.MR.11133 and HLMD.MR.11134 are plaster casts; the whereabouts of the original specimens are unknown. Here, we select specimen HLMD.MR.11135 as lectotype and HLMD.MR.11132 as the only paralectotype.
Studied material. Ca. 600 specimens (MB.Ro.31–34, Ro.47, Ro.48; SMF.Mbg.4999; IPB beushausen 5, 6).
Type locality, stratum and age. Haina(u) mine near Waldgirmes, Gießen, Hesse, Germany. ‘Stryngocephalus’ limestone (Maurer 1885) = Stringocephalus Limestone, ‘Massenkalk Formation’, middle to upper Givetian, Middle Devonian (Figs. 1, 2).
Occurrence. Upper Middle Devonian (upper Givetian); Lahn-Dill area (Hesse), Eifel Hills and Bergisches Land (North Rhine-Westphalia), Rhenish Massif, Germany (Figs. 1, 2).
Original description. See Maurer (1885, p. 229).
First revised description. See Beushausen (1895, p. 389).
Diagnosis. Bohemicardiine hippocardiid with ventral margin of snout running diagonally; ventral margin of central body convex; rostrum almost parallel to dorsal axis. Surface radially ornamented: middle shell layer of central body with 6–7 radial ribs and wide interspaces; outer shell layer with similar number of rounded radial ribs and coarse growth lines leading to reticulate ornamentation.
Revised description (Figs. 4, 5, 6, 7, 8, 10, 11). Conch bohemicardiine; body carina not developed; shape of snout triangular to slightly nasute; course of ventral margin of snout running diagonally.
Snout gape drop-shaped in outline, marginal denticles weakly developed; snout rib slender. Umbo small, pointed and protruding above dorsal margin; ventral margin of central body convex, running nearly parallel to dorsal axis (‘parallel convex’ after Rogalla and Amler 2006e); maximum curvature situated in the middle of the central body. Rostral area occupying about 80–85% of total height of the body, its surface covered with flat radial ribs forming a zigzag-shaped rostral keel (Fig. 7n). Juvenile specimens showing heart-shaped, longitudinally ovate rostral area increasing in width (laterally) during growth; rostral area of adult specimens circular heart-shaped (cordate) (Fig. 11). Rostral area surrounded by lunulazone (Fig. 7d, i), a row of crescent-shaped growth lines representing attachment line of hood. Ovate orifice located at ventralmost point of lunulazone; in a few specimens orifice preserved as tubular steinkern (Figs. 4b, 7f). Rostrum oriented parallel to dorsal axis and relatively large, occupying about 20–25% of total height (Figs. 7a, f, k, 8f, 11a, k); in most specimens rostrum broken off or preserved only about 2 mm of its length; rostral clefts present.
External shell layer, when preserved (Figs. 4a, c, 7f, 8a), ornamented with rounded radial plicae (ribs), 6–7 on the central body, 7–8 on the snout; additional 3–4 low ribs may be developed on snout rib. Ribs and interspaces covered with medium coarse growth lines (Fig. 4a, c) forming a reticulate pattern.
Middle shell layer (Figs. 5c, e, 6e, f, 7k, 8f, 11a, f, k) consisting of 6–7 radial plicae in the central body and 7–8 on the snout, all of which flat-topped. Surface of snout rib covered with fine growth lines (Figs. 6e, 8a), rarely visible in other parts of the conch. Due to interlocking at ventral margin (Figs. 6g, 8c, m), plicae and interspaces almost equal in width.
Measurements. See Table 1.
Remarks. A few specimens show a very slightly concave ventral margin of the snout due to variable preservation.
Discussion
Original material and nomenclature
Conocardium hainense was erected by Maurer (1885, p. 230) based on an unknown number of syntypes, he simply stated that he had several specimens available for study. Two of the syntypes plus two plaster casts of probable further syntypes were available for our revision. Comparison of these specimens with the original figures (Maurer 1885, pl. 9, fig. 21, 21a; here Fig. 5) leads to the conclusion that the illustrations are inaccurate and embellished, but most likely agree.
Beushausen (1895, p. 390), who regarded Maurer’s specimens as insufficient, revised the taxon based on some(?) original specimens and newly collected material (Fig. 6). Beushausen’s descriptions are much more detailed, but he did not re-illustrate the specimens figured by Maurer, nor did he mention the number of specimens available for his study. However, he considered Maurer’s specimens as ‘distorted and therefore not typical for the shape of the shell’ (Beushausen 1895, p. 390, translated from German; see Fig. 5f). Thus, in his opinion the syntypes do not show the correct proportions of the conch.
Most of the material described and illustrated by Beushausen (1895, pl. 29, figs. 1–4) is lost. The morphology of the specimen figured in pl. 29, fig. 1 (coll. Maurer; lost) is clearly different from that of typical C. hainense, and the rostral area extends over the entire conch height; this specimen is therefore excluded from the species concept. Apart from the absence of the first body carina, which is probably abraded, this specimen rather conforms to the description of Bohemicardia clathrata. The specimen illustrated in pl. 29, fig. 2 of Beushausen (1895) from Sötenich (Fig. 6a–d) represents a well-preserved individual of Bo. hainense and perfectly fits the revised species concept. The specimen was kept in the collections of the ‘Breslau Museum’ (today Muzeum Uniwersytetu Wrocławskiego = Museum of the University of Wrocław, Poland), but is also considered lost (pers. comm. Paweł Raczyński, University of Wroclaw).
Four specimens of the Beushausen collection kept in the Goldfuß Museum, University of Bonn (IPB beushausen 5–1, 5–2, beushausen 6–1, 6–2) were available for study and comparison. Specimen IPB beushausen 5–1, although labelled as ‘Conocardium hainense var. impressa’ (Fig. 6e–j), rather matches figure 2 in pl. 29 (Conocardium hainense) except for the length of the rostrum. The two specimens IPB beushausen 6 labelled as Conocardium n. sp. aff. hainense do not match the figures (Beushausen 1895, pl. 29, fig. 4) and are here assigned to Babinicardia clathrata (Archiac and Verneuil, 1842) based on the presence of the first body carina.
Beushausen (1895) also erected the variety C. hainense var. impressa (Fig. 6k–n) and mentioned a depressed rostral area as a characteristic feature. Furthermore, he described and figured one specimen provisionally named Conocardium n. sp. aff. hainense. However, the two specimens from the Beushausen collection that are labelled accordingly are not the illustrated ones and may have been mixed up.
Schröder-Rogalla (2005) and Rogalla and Amler (2006c) erroneously regarded Conocardium hainense as a subjective junior synonym of Babinicardia clathrata (Archiac and Verneuil, 1842), but excluded the variety impressa from the species concept of Ba. clathrata. This error is based on the fact that the type material was considered lost and, thus, not revisited.
Preservation and taxonomy
Most rostroconchs are preserved with both semi-conchs in their original position and contact as the continuous dorsal shell layer(s) prevent(s) a gaping of the ‘valves’ and disarticulation after death in contrast to bivalves (see Rogalla et al. 2003; Schröder-Rogalla 2005; Amler and Rogalla 2007 for details). Fragile and distal parts of the shell, such as the hood, the most anterior portion of the snout and the rostrum, are often damaged or broken off owing to post-mortem taphonomic processes. The ample material studied, however, allows almost complete reconstruction of the morphology of Bohemicardia hainense including indication of the hood deduced from the specimens embedded in matrix (Fig. 4a) and the presence of the lunulazone (Figs. 3, 4b, 7d, i).
As exemplified by the figured specimens (Figs. 4, 7, 8, 11), for the determination of rostroconchs preservation of the different shell layers is proven fundamentally important. Rostroconchs possess three architectural shell layers, not mirrored by microstructure (Rogalla et al. 2002, 2003; Amler and Rogalla 2007). The inner layer is delicate and rarely exposed, and the middle layer is relatively thick, mostly consisting of strong radial ribs (Figs. 5c, e, 6e, f, 7k, 8f, 11a, f, k). This is the layer exposed and hitherto described in the majority of rostroconch taxa. The outer layer fills the interspaces of these ribs and smooths the external surface or shows reticulate patterns (Figs. 4a, c, 8a). If the outer shell layer and parts of the middle layer are abraded, ill preserved or removed, the outline of the conch is modified significantly, leading to incorrect reconstruction or description of the outline (Amler and Rogalla 2013 and references therein). Many specimens show different modes of preservation, sometimes even in a single specimen (Figs. 7, 8; Amler 1996; Schröder-Rogalla 2005; Rogalla and Amler 2006b, 2007).
In contrast to Schröder-Rogalla (2005) and Rogalla and Amler (2006c, d), it could be proven by the presence of a lunulazone that Bo. hainense possesses a hood and, thus, is a member of the superfamily Hippocardioidea Pojeta and Runnegar and the family Hippocardiidae Pojeta and Runnegar (Rogalla and Amler 2006c). The six subfamilies of the Hippocardiidae differ in the ratio of rostral height to total height. Whereas the Barrandeicardiinae is characterised by a rostral height of only 50% of the total height, the Bohemicardiinae distinguishes a rostral area of about 75–90% of the total height. In all other subfamilies, the rostral area is as high as the entire height of the conch (100%). Consequently, due to the development of the rostral area and the lack of body carinae, Bo. hainense is a member of the Bohemicardiinae, even though the rostral area occupies a greater height than the original subfamily definition requires (for further details on differences within the family Hippocardiidae, see Schröder-Rogalla 2005, p. 133; Rogalla and Amler 2006c, p. 347). Therefore, the subfamily diagnosis was emended here.
Within the Bohemicardiinae, the genera mostly differ in the shape of the snout and the ventral margin. The triangular snout and the convex, curved ventral margin prove an assignment to the genus Bohemicardia Rogalla and Amler, 2006a, b, c, d, e. Consequently, the taxon Bo. hainense is not to be included in Babinicardia clathrata, as erroneously considered by Rogalla and Amler (2006c, d).
The specimens at hand labelled as Conocardium hainense var. impressa Beushausen 1895 (Fig. 6e–j) do not correspond to the description and the illustration of the variety presented by Beushausen. The most important difference from the type subspecies is the development of the rostral area. Beushausen described and illustrated the rostral area as depressed, concave, but both specimens at hand do not show this feature. This seems to have been either an observation error by Beushausen or the specimens are not the figured syntypes. It can be stated, however, due to the fact that the specimens of var. impressa show a different size and, thus, a different growth stage that there is a change in the shape of the rostral area from cordate longitudinally oval to cordate circular, just as in Bo. hainense. Based on these facts and supported by the large number of specimens from Sötenich, the two present specimens of Co. hainense var. impressa are included in the species concept of Bohemicardia hainense.
Furthermore, Beushausen (1895) erected a new species in open nomenclature, Conocardium n. sp. aff. hainense, and figured one specimen. We have two specimens at hand, neither of which is the illustrated one. One specimen is badly preserved, slightly deformed and shows different proportions compared with Bo. hainense, with an elongated snout. The other specimen is better preserved, partly showing the outer shell layer. As a difference from Bo. hainense, Beushausen stated a more elongated shape and more but weaker ribs. Furthermore, both specimens show a distinct and regular commarginal striation crossing the radial ribs, especially visible on the middle shell layer, but also with a fine cover by the outer shell layer. We exclude both specimens from the species concept of B. hainense based on the characters mentioned above and presume affinities with Babinicardia clathrata.
Species-level comparison
Three species are currently assigned to the genus Bohemicardia Rogalla and Amler 2006c, i.e. Bo. bohemica (Barrande, 1881), Bo. rotunda (Paeckelmann, 1913) and Bo. hainense (Maurer, 1885), which mostly differ in conch proportions. For both Bo. bohemica and Bo. hainense abundant material exists in different preservation and also with almost completely preserved outer shell layer. In Bo. rotunda, however, there is only the lectotype available, without the outer shell layer preserved (SMF.Mbg.3362; Fig. 9), all further syntypes are lost and additional material is not (yet) known. The species was described and illustrated by Paeckelmann (1913, p. 258) and Rogalla and Amler (2006c, p. 360).

Bohemicardia rotunda (Paeckelmann, 1913). a–d Original illustrations of Paeckelmann (1913, pl. 6, fig. 1, 1a–c). e–h lectotype SMF.Mbg.3362 from the Dorp Limestone (Knappertsbusch quarry) of Beek near Wuppertal-Elberfeld, Germany, in right lateral (e), dorsal (f), ventral (g) and posterior (h) view. Scale bar 1 mm
In comparison, the ventral margin of Bo. hainense is curved, slightly convex, whereas Bo. rotunda shows a tighter convex ventral margin, slightly pointed in the central part. In contrast, Bo. bohemica shows a regular, evenly convex ventral margin. Bo. bohemica, particularly, differs from the other two species in general shape of the central body. The central body is much larger and regularly rounded, the snout is very small, similar to the rostral area and Bo. bohemica has a small rostral field, which is about 75% of the total height only. Bo. bohemica shows a moderately wide variability (see illustrations in Rogalla and Amler 2006c). Specimens of Bo. bohemica are easily distinguishable from Bo. hainense and Bo. rotunda as long as the thick outer shell layer is preserved. In abraded specimens, however, the ventral margin of the central body in Bo. bohemica becomes narrower and resembles specimens of Bo. hainense.
Moreover, the orientation of the rostrum relative to the dorsal axis differs among the species compared. In Bo. hainense, the rostrum and the dorsal axis are almost parallel, whereas in Bo. bohemica rostrum and dorsal axis diverge distinctly dorsally. In Bo. rotunda no details on the position of rostrum can be observed because the rostrum of the lectotype is not preserved (see Rogalla and Amler 2006c for more information).
Based on specimens found at the same locality in the Eifel region, the Wachtberg near Sötenich, Rogalla (2007) erected Rheiacardia lerami Rogalla, 2007. This taxon differs from Bo. hainense in the height of the rostral area (100%), a nasute snout with a slightly sinuate ventral margin and the orientation of the ventral margin of the central body. Additionally, Bieberiana pugnans (Whidborne, 1892) occurs in Middle Devonian strata of central and western Europe. This taxon was described under several names, e.g. Conocardium confusum Beushausen, 1895, Conocardium eifeliense Beushausen, 1895 and Conocardium retusum Maurer, 1885, and later revised by Rogalla and Amler (2006c). Bi. pugnans differs from Bo. hainense mainly by the significantly higher rostral area, which accounts for the entire height of the conch. In addition, the snout of Bi. pugnans is straight alaeform and, thus, elongated. Although some specimens figured as Bi. pugnans may be slightly tectonically deformed, its overall appearance is more elongate than Bo. hainense.
Contemporaneously, Babinicardia clathrata (Archiac and Verneuil, 1842) occurs in Middle Devonian strata of the Eifel Hills. It is the only genus and species within the subfamily Babinicardiinae of the family Hippocardiidae, and it is the only species with a body carina, which gives this species a triangular appearance of the central body. This characteristic morphology, which is well recognised both with and without the outer shell layer preserved, differs in a few subtleties from Bohemicardia hainense. Ba. clathrata shows a characteristic, triangular box-shaped central body, delimited from the snout by a body carina, and a comparably straight or very slightly convex ventral margin. The height of the rostral area approximates nearly 100% of the total height, placing the orifice at the posteroventral corner. In well-preserved condition, these differences are easily visible, but if the body carina in Bo. clathrata is abraded, the box-like shape of the central body gets lost and the ventral margin appears more rounded. In contrast, if the ventral margin of the central body in Bo. hainense is abraded, it runs almost parallel to the dorsal axis and the rostral surface occupies almost the entire height of the specimen, mimicking the shape of Ba. clathrata. Thus, careful study of preservation patterns is necessary for proper identification of the specimens.
Variability and post-larval ontogeny
The large number of specimens of Bohemicardia hainense allows to extend the species description with some details on growth stages from post-larval juvenile to adult specimens. Furthermore, based on the 50 well-preserved specimens selected individually for measurements (Table 1), we received enough data on the variability of Bo. hainense concerning the basic dimensions of the conch (Fig. 10). This has been impossible so far for most other rostroconch taxa as usually there is not enough well-preserved material available. In this study, the largest specimen has a total length of about 15 mm and the smallest 2.5 mm, but not all of them were included in the measurements due to incomplete preservation. To estimate the range of variation and for comparison of the morphological development, three dimensions were determined in all specimens measured: length (LP), height (H) and width (W). Although really basic the three ratios, LP to H, LP to W and H to W, clearly show very low dispersion of values (Fig. 10). The mean value \(\overline{{\varvec{x}} }\) for LP to H is 1.22, for LP to W 1.36 and for H to W 1.11. The standard deviation in all three relations is 0.09, and the standard error of mean is 0.01. These data confirm that the taxon is relatively well circumscribed and characterised.

During post-larval ontogeny, the length of the specimens grows faster in comparison to the height and width. The width is small at first and equalises with the height values in the course of growth. The increase in width and height at different rates causes a change, particularly, in the shape of the rostral area. The smallest specimen shows a narrow and high rostral area (cordate longitudinally oval), whereas the larger specimens show a much rounder (cordate circular) rostral face (Fig. 11) being the result of increasing width relative to the height.

Bohemicardia hainense (Maurer, 1885) from the upper Middle Devonian of Girzenberg (Girzen Hill) near Sötenich, Eifel Hills, district Euskirchen, Germany, in ontogenetic growth stages. Note the different proportions from juvenile to adult specimens. a–e MB.Ro32.5 in left lateral (a), dorsal (b), ventral (c), posterior (d) and anterior (e) view. f–j MB.Ro32.4 in left lateral (f), dorsal (g), ventral (h), posterior (i) and anterior (j) view. k–o MB.Ro32.1 in left lateral (k), dorsal (l), ventral (m), posterior (n) and anterior (o) view. Scale bars 1 mm
The number of radial ribs of the middle shell layer in the central body of the conch does not increase when the shell grows. Instead, both ribs and interspaces become broader ventrally. With increasing conch width, the number of ribs covering the rostral area increases slightly. A third morphological change concerns the diameter of the rostrum. In small specimens, the size of the rostrum is relatively larger than in bigger individuals. Its relative diameter decreases from 2/5 of the entire conch height to 1/4 (Fig. 11a, k).
The ontogenetic growth from the larval to the early juvenile stage is not preserved in the studied material. Deduced from earlier observations (Schröder-Rogalla 2005), rather differing relations of the measured dimensions are expected compared with our analysis (Fig. 10). Growth of the two-lobed, pseudo-bivalved larval shell in anterior and lateral, but slowly in posterior direction terminates at about 1 mm length (Schröder-Rogalla 2005) and the early juvenile shell is formed below the protoconch. Thus, specimens of 1–2 mm in length are required to report on this growth stage.
Palaeoecology and possible life habits
The palaeoecological distribution of conocardiid rostroconchs seems to have been controlled by various environmental factors such as nutrient supply, sediment grain size and stability, salinity, oxygen content, and water temperature. The wide range of basic morphologies in rostroconchs as well as palaeobiological constraints derived from the basic molluscan bauplan lead to the assumption that several lifestyles were realised for the occupation of different niches, e.g. reef margin, soupy mud or unstable sand (Richter and Amler 1995; Rogalla and Amler 2005a, b; Amler and Rogalla 2013; Amler 2016). Thus, the morphology of many Conocardiida reflects their life habits, such as mobile epibenthic crawling, immobile semi-endobenthic sediment sticking, and the loss of the hood in advanced conocardiids indicates a rather mobile, almost endobenthic life position (Runnegar 1978; Pojeta 1987; Richter and Amler 1995; Amler and Richter 1997; Schröder-Rogalla 2005; Amler and Rogalla 2013). The related trophic perspective, particularly an alternative interpretation of the life habit of hippocardioid rostroconchs, was discussed elsewhere (Schröder-Rogalla 2005; Rogalla and Amler 2005a, b; Amler and Rogalla 2013; Amler 2016). Accordingly, hippocardiids most probably obtained a specific life position relative to the sediment surface. With respect to mobility, stable position within the substrate and efficient orientation of the posterior surface of the hood, only one life position is able to meet each of the demands properly, a rather horizontal orientation of the rostral area parallel with the surface of the substrate (Amler 2016).
The morphology of Bohemicardia hainense supports these assumptions except for the large conch size required for photosymbiosis and the relatively large diameter of the rostrum. The short, compact conch and the relatively large diameter of the posterior rostral area plus the hood (compared with height and length) were hardly suitable for rapid or even moderate locomotion, even if the grain size of the sediment offered sufficient pore space for grain movement (Amler 2016). The small size of the taxon, the relatively small rostral area surrounded by a narrow hood if compared with Devonian taxa from North America [e.g. Hoareicardia cunea (Conrad, 1840)] and Mississippian taxa [e.g. Hippocardia hibernica (Sowerby, 1812)], as well as the relatively wide diameter of the rostrum, indicate an early evolutionary stage of the mixed feeding habit.
Conclusions
-
The study of a relatively large number of specimens and the re-examination of preserved type material enabled the reinstatement of ‘Conocardium hainense Maurer, 1885’.
-
The completely reconstructed and redescribed morphology of C. hainense allows the assignment to the genus Bohemicardia within the Hippocardiidae of the Hippocardioidea.
-
The large number of specimens shows a rather narrow range of morphological variation within the taxon.
-
The abundant material available, including a wide size range of specimens, allows the description of the ontogenetic development. A change of shell dimensions during growth, particularly a distinct increase in width versus height is documented. Thus, the shape of the rostral area changes from narrow and high (cordate longitudinally oval) to roundish (cordate circular) in larger specimens.
-
Protoconchs are not preserved leaving the development from larval stages to early juveniles unrecorded.
-
Based on ample preservation of the rarely observed external shell layer information on the proper shell shape, presence of a hood and external shell ornamentation of Bo. hainense and on hippocardiids in general are possible.
-
The studied material confirms the difficulties in recognising different shell layer architecture in conocardiid rostroconchs, which is a prerequisite for specific determination.
-
Bohemicardia hainense appears to have been an abundant taxon in late Middle Devonian shallow shelf environments at the southern margin of Laurussia.
-
From the stratigraphic viewpoint, an evolutionary development from Bohemicardia bohemica (late Lower Devonian) to Bo. hainense in the Middle Devonian and Bo. rotunda in the early Upper Devonian may be plausible. Unfortunately, several strata between the recorded occurrences are void of rostroconchs and the precise age of the latter species is unknown, leaving the postulated lineage unproven.
-
The morphology of Bohemicardia hainense supports the assumption of an early evolutionary stage of the mixed feeding habit in hippocardiids for this taxon.
References
Amler, M.R.W. 1986. Revision der Rostroconchien aus dem Unter-Karbon von Königsberg bei Gießen (nördliche Lahnmulde). Geologica et Palaeontologica 20: 73–85.
Amler, M.R.W. 1996. Giant hippocardiids from the Lower Carboniferous of Western Europe. Irish Journal of Earth Sciences 15: 113–122.
Amler, M.R.W. 2016. Shallow marine Rostroconchia (Mollusca) from the latest Devonian (Strunian) and their significance for rostroconch life style and evolution. Geologica Belgica 19: 111–120.
Amler, M.R.W. 2018. Late Early Devonian Rostroconchia (Mollusca) from Hamar Laghdad (Morocco). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 290: 183–189.
Amler, M.R.W., and E. Richter. 1997. Conocardium und Hippocardia − Ernährungs- und Habitatspezialisten unter den Rostroconchien? Programm der 67. Jahrestagung der Paläontologischen Gesellschaft, Daun. Terra Nostra 97/6: 18–19.
Amler, M.R.W., and N.S. Rogalla. 2004. History and nomenclature of the Conocardioidea (Mollusca: Rostroconchia). Paläontologische Zeitschrift 78: 307–322.
Amler, M.R.W., and N.S. Rogalla. 2007. “Moderne” Rostroconchien: Conocardium und Hippocardia. Fossilien 2007: 149–154.
Amler, M.R.W., and N.S. Rogalla. 2013. Chapter 17. Biogeographical distribution patterns in Early Palaeozoic Rostroconchia (Mollusca). In Early Palaeozoic Biogeography and Palaeogeography, eds. D.A.T. Harper, and T. Servais, Geological Society of London Memoirs 38: 235–255.
Amler, M.R.W., B. Balik, and J.C. Friedel. 2020. Neue alte Rostroconchia (Mollusca) aus dem Mitteldevon der Eifel. Archäologie im Rheinland 2019: 41–43; Oppenheim (Nünnerich-Asmus).
Archiac, E.JAd.’, and P.E.P. de Verneuil. 1842. On the fossils of the older deposits in the Rhenish Provinces; preceded by a general survey of the fauna of the Palaeozoic rocks, and followed by a tabular list of the organic remains of the Devonian system in Europe. Transactions of the Geological Society of London, Series 2(6): 303–408.
Balik, B. 2018. Rostroconchia (Mollusca) aus dem Mitteldevon der Eifel. Unpublished BSc thesis, Universität zu Köln.
Barrande, J. 1881. Systême Silurien du centre de la Bohême. 1. Recherches Paléontologiques, 6: Classe des Mollusques. Ordre des Acéphalés. Prag, Paris: published by the author.
Bender, P. 2008. Chapter 4.1.5 Lahn- und Dill-Mulde. In Stratigraphie von Deutschland VIII. Devon, ed. Deutsche Stratigraphische Kommission. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften 52: 221–246.
Beushausen, L. 1895. Die Lamellibranchiaten des rheinischen Devon mit Ausschluss der Aviculiden. Abhandlungen der Königlich Preussischen geologischen Landesanstalt, Neue Folge 17: 1–514.
Beyer, E. 1896. Beitrag zur Kenntniss der Fauna des Kalkes von Haina bei Waldgirmes (Wetzlar). Verhandlungen des Naturhistorischen Vereines der Preussischen Rheinlande 53: 55–102.
Blakey. https://deeptimemaps.com. Access 21 July 2021
Burchette, T.P. 1981. European Devonian reefs: a review of current concepts and models. In European Fossil Reef Models, ed. D.F. Toomey, SEPM Special Publications 30: 85–142.
Clausen, C.-D., and D. Korn. 2008. Chapter 4.1.15 Höheres Mitteldevon und Oberdevon des nördlichen Rheinischen Schiefergebirges (mit Velberter Sattel und Kellerwald). In Stratigraphie von Deutschland VIII. Devon, ed. Deutsche Stratigraphische Kommission. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften 52: 439–481.
Conrad, T.A. 1840. Third annual report on the Palaeontological Department of the Survey [of New York]. Fourth Annual Report, State of New York [Geological Survey]. Assembly Document 50: 199–207.
Flick, H., and H.-D. Nesbor. 2021. Chapter 3.3 Lahn-Dill-Gebiet. In Geologie von Hessen, ed. R. Becker and T. Reischmann, 49–77. Schweizerbart.
Gierse, D. 2003. Anwendung statistischer Methoden in der Paläobiologie jungpaläozoischer Rostroconchien. Unpublished Diploma thesis, Philipps-Universität Marburg.
Golonka, J. 2002. Plate-tectonic maps of the Phanerozoic. In Phanerozoic reef pattern, vol. 72, ed. W. Kiessling, E. Flügel, and J. Golonka, 21–75. Society for Sedimentary Geology, Special Publications.
Grabert, H. 1998. Abriß der Geologie von Nordrhein-Westfalen. E. Schweizerbart’sche Verlagsbuchhandlung.
Holzapfel, E. 1895. Das Obere Mitteldevon (Schichten mit Stringocephalus burtini und Maeneceras terebratum) im Rheinischen Gebirge. Abhandlungen der Königlich Preussischen geologischen Landesanstalt, Neue Folge 16: 1–459.
ICZN (International Commission on Zoological Nomenclature). 1999. International code of zoological nomenclature. 4th edition, xxix + 306 p. London: International Trust for Zoological Nomenclature.
Johnson, J.G., G. Klapper, and C.A. Sandberg. 1985. Devonian eustatic fluctuations in Euramerica. Geological Society of America, Bulletin 96: 567–587.
Jux, U. 2008. Chapter 4.1.13 Bergisch Gladbach-Paffrather Mulde. In Stratigraphie von Deutschland VIII. Devon, ed. Deutsche Stratigraphische Kommission. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften 52: 402–416.
Kegel, W. 1933. Erläuterungen zur geologischen Karte von Preußen und benachbarten deutschen Ländern, Lief. 317 Blatt Rodheim. Berlin: Preußische Geologische Landesanstalt.
Litholex. 2021. https://www.bgr.bund.de/DE/Themen/GG-Stratigraphie/LithoLex/litholex_node.html. Access 03 Oct 2021
Maurer, F. 1885. Die Fauna der Kalke von Waldgirmes bei Giessen. Abhandlungen der Grossherzoglichen Hessischen Geologischen Landesanstalt zu Darmstadt 1(2): 61–340.
Neumayr, M. 1891. Beiträge zu einer morphologischen Eintheilung der Bivalven. Denkschriften der Mathematisch Naturwissenschaftlichen Classe der Kaiserlichen Akademie der Wissenschaften 58: 708–801.
Paeckelmann, W. 1913. Das Oberdevon des Bergischen Landes. Abhandlungen der Königlich Preußischen Geologischen Landesanstalt, Neue Folge 70: 1–356.
Pojeta, J., III. 1987. Class Rostroconchia. In Fossil invertebrates, ed. R.S. Boardman, A.H. Cheetham, and A.J. Rowell, 358–380. Blackwell Science Publications.
Pojeta, J., Jr., and B. Runnegar. 1976. The palaeontology of rostroconch molluscs and the early history of the phylum Mollusca. United States Geological Survey, Professional Paper 968: 1–88.
Pojeta, J., Jr., B. Runnegar, N.J. Morris, and N.D. Newell. 1972. Rostroconchia: A new class of bivalved molluscs. Science 177: 264–267.
Quiring, H. 1914. Zur Stratigraphie und Tektonik der Eifelkalkmulde von Sötenich. Jahrbuch der Königlich Preußischen Geologischen Landesanstalt [für 1913] 34(2): 81–223.
Ribbert, K.-H. 1985. Geologische Karte von Nordrhein-Westfalen 1 : 25000. Erläuterungen zu Blatt 5405 Mechernich. Krefeld: Geologisches Landesamt Nordrhein-Westfalen.
Richter, E., and M.R.W. Amler. 1994. Bivalven und Rostroconchien aus dem Velberter Kalk von Velbert (Unter-Karbon; Bergisches Land). Geologica et Palaeontologica 28: 103–139.
Richter, E., and M.R.W. Amler. 1995. Wie lebte Conocardium? In Programm der 65. Jahrestagung der Paläontologischen Gesellschaft, Hildesheim. Terra Nostra 4/95: 71.
Ritz, H. 1931. Die Südwesthälfte der Sötenicher Mitteldevon-Mulde. Jahrbuch der Preußischen Geologischen Landesanstalt 51: 565–574.
Rogalla, N.S. 2007. Rheiacardia lerami n. gen. n. sp., ein hippocardiider Rostroconch aus dem Devon Laurussias. Paläontologische Zeitschrift 81(1): 93–96.
Rogalla, N.S., and M.R.W. Amler. 2005a. Paläophotosymbiose – der Schlüssel zur Lebensweise hippocardioider Rostroconchia (Mollusca). In Beitragskurzfassungen zur 75. Jahrestagung der Paläontologischen Gesellschaft 2005 in Graz. Berichte des Instituts für Erdwissenschaften der K-F-Universität Graz 10: 106–107.
Rogalla, N.S., and M.R.W. Amler. 2005b. Palaeophotosymbiosis – the key to understanding the life habit of hippocardioid Rostroconchia (Mollusca). In 49th Annual Meeting 2005b at Oxford, Abstracts. Palaeontological Association Newsletters 60 (Abstracts): 55.
Rogalla, N.S., and M.R.W. Amler. 2006a. Revision der Familie Pseudobigaleaidae Hoare, Mapes and Yancey 2002 (Mollusca; Rostroconchia). Paläontologische Zeitschrift 80(2): 167–208.
Rogalla, N.S., and M.R.W. Amler. 2006b. Revision der Familie Hippocardiidae Pojeta and Runnegar, 1976 (Mollusca; Rostroconchia). Paläontologische Zeitschrift 80(3): 238–276.
Rogalla, N.S., and M.R.W. Amler. 2006c. Taxonomie und Systematik der Hippocardioidea Pojeta and Runnegar, 1976 [n. superfam.] (Mollusca; Rostroconchia). Paläontologische Zeitschrift 80(4): 344–383.
Rogalla, N.S., and M.R.W. Amler. 2006d. Index of conocardioid and hippocardioid rostroconch taxa (Mollusca: Rostroconchia). Geologica et Palaeontologica 40: 27–61.
Rogalla, N.S., and M.R.W. Amler. 2006e. New terminology of rostroconch morphology. In Bivalvia 2006. International Congress on Bivalvia July 22–27, 2006. Abstracts, eds. N. Malchus, and J.M. Pons, Organisms, Diversity and Evolution 6, Electronic Supplement 16(1): 64–65 plus poster. Bellaterra: Universitat Autònoma de Barcelona.
Rogalla, N.S., and M.R.W. Amler. 2007. Nomina dubia der Hippocardioidea (Mollusca; Rostroconchia). Paläontologische Zeitschrift 81(1): 29–70.
Rogalla, N.S., M.R.W. Amler, J.G. Carter, and J. Pojeta Jr. 2002. Schalenarchitektur und -mikrostruktur bei Conocardioidea (Rostroconchia, Mollusca). In GEO 2002 – Planet Erde: Vergangenheit, Entwicklung, Zukunft. Programm und Kurzfassungen. Schriftenreihe der Deutschen Geologischen Gesellschaft 21: 280.
Rogalla, N.S., J.G. Carter, and J. Pojeta Jr. 2003. Shell microstructure of the Late Carboniferous rostroconch mollusc Apotocardium lanterna (Branson, 1965). Journal of Paleontology 77: 655–673.
Runnegar, B. 1978. Origin and evolution of the class Rostroconchia. Philosophical Transactions of the Royal Society of London, B 284: 319–333.
Schmidt, W.E. 1936. Die Schichtenfolge des Devons bei Sötenich in der Eifel. Jahrbuch der Preußischen Geologischen Landesanstalt 56: 292–323.
Schröder-Rogalla, N. 2005. Paläobiologie und Systematik der neuen Überfamilie Hippocardioidea Pojeta and Runnegar, 1976 (Rostroconchia, Mollusca). PhD thesis, Philipps Universität Marburg. http://archiv.ub.uni-marburg.de/diss/z2005/0515/pdf/dnsr.pdf
Scotese. http://www.scotese.com/earth.htm, http://www.scotese.com/newpage4.htm and http://www.scotese.com/late.htm. Access 21 July 2021
Sowerby, J. 1812–15. The mineral conchology of Great Britain; or coloured figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the earth, 1. 234 p. + index, pls. 1–102. London: published by the author [1–32, pls. 1–9 (1812), 33–96, pls. 10–44 (1813), 97–178, pls. 45–78 (1814), 170–234, pls. 79–102, Index (1815)].
STD 2016, ed. German Stratigraphic Commission. 2016. Stratigraphic Table of Germany 2016. Potsdam: German Research Centre for Geosciences.
Struve, W., M. Basse, and K. Weddige. 2008. Chapter 4.1.10 Prädevon, Ober-Emsium und Mitteldevon der Eifeler Kalkmulden-Zone. In Stratigraphie von Deutschland VIII. Devon, ed. Deutsche Stratigraphische Kommission. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften 52: 297–374.
Whidborne, G.F. 1892–1907. A monograph of the Devonian fauna of the South of England, 2. The fauna of the limestones of Lummaton, Wolborough, Chircombe Bridge, and Chudleigh, 2(1). Palaeontographical Society Monograph 45(216): 1–56, pls. 1–5 [1892a]; 2(2). Palaeontographical Society Monograph 46(221): 57–88, pls. 6–10 [1892b]; 2(3). Palaeontographical Society Monograph 47(225): 89–160, pls. 11–17 [1893]; 2(4). Palaeontographical Society Monograph 49(233): 161–214, pls. 18–24 [1895]; 2(5). Title, Index [1907].
Acknowledgements
It is a pleasure to acknowledge D. Korn and A. Abele-Rassuly (Museum für Naturkunde Berlin), A. Ehling (Bundesanstalt für Geowissenschaften und Rohstoffe, Berlin), T. Wappler (Hessisches Landesmuseum Darmstadt), G. Heumann (Goldfuß Museum, University of Bonn) and H.M. Weber (Ruhrmuseum Essen), who provided the specimens for study. We thank J.-O. Ebbestad (Evolutionsmuseet, University of Uppsala) and Simon Schneider (CASP, Cambridge) for their detailed and constructive reviews, which improved the manuscript significantly.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Markus Aretz.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Friedel, J.C., Amler, M.R.W. & Balik, B. Bohemicardia hainense (Maurer) revisted: insights into ontogeny and variability of a Devonian hippocardiid rostroconch (Conocardiida: Hippocardiidae). PalZ 97, 245–263 (2023). https://doi.org/10.1007/s12542-022-00645-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-022-00645-3