
Abstract
The hadrosauroid Telmatosaurus and the rhabdodontid Zalmoxes were the first and second dinosaur taxa that were described in detail from the famous Upper Cretaceous continental deposits of the Haţeg Basin by Franz Baron Nopcsa at the beginning of the twentieth century. Although they are among the most common and best-known dinosaurs discovered from these deposits, there are still many open questions as to their taxonomy and anatomy. Here, we re-describe two partial braincases from the uppermost Cretaceous of the Haţeg Basin that have been recently referred to the rhabdodontid Zalmoxes and re-assign them to hadrosauroids, possibly to Telmatosaurus. These specimens both exhibit basicranial features that are characteristic of derived hadrosauroids but are absent in more basal iguanodontians. These include an antero-posteriorly short basioccipital lacking a distinct neck, the presence of two well-developed sphenoccipital tubercles on the ventral aspect of the braincase and that are directly positioned anterior to the basioccipital, as well as a deep depression on the ventral aspect of the braincase between the sphenoccipital tubercles. The comparison provided herein demonstrates several important differences between the basicranium of hadrosauroids and that of rhabdodontids, which allows for the confident identification of even isolated and incomplete specimens. Moreover, the removal of the only basicranium that has been referred to Zalmoxes shqiperorum prompts a revised diagnosis of that species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The hadrosauroid dinosaur Telmatosaurus transsylvanicus was the first vertebrate taxon named from the famous Upper Cretaceous vertebrate-yielding continental deposits of the Haţeg Basin in western Romania. It was described in detail by the renowned palaeontologist Franz Nopcsa based on a largely complete but crushed skull (Nopcsa 1900). In the following years, Nopcsa described a diverse vertebrate fauna from the Haţeg Basin including turtles, crocodyliforms, and dinosaurs (e.g., Nopcsa 1902a, b, 1904, 1923, 1929), which now is one of the best-known Late Cretaceous vertebrate assemblages of Europe (for an overview, see Weishampel et al. 1991; Grigorescu 2010; Csiki-Sava et al. 2015, 2016). A number of additional specimens referred to Telmatosaurus have been discovered and reported since the initial description by Nopcsa, both by himself as well as by other researchers more recently, making it one of the better-known taxa from the Haţeg Basin and the whole Upper Cretaceous of Europe (Weishampel et al. 1993; Dalla Vecchia 2006, 2009a); it is conceivable, however, that hadrosauroid fossils from the Transylvanian area that were customarily assigned to Telmatosaurus transsylvanicus may represent instead several, possibly closely related taxa (e.g., Dalla Vecchia 2009a). Despite being hailed as the basal-most hadrosaurid dinosaur (e.g., Weishampel et al. 1993), Telmatosaurus is now generally regarded as a basal hadrosauroid, lying outside the radiation of derived hadrosaurids (Sues and Averianov 2009; Prieto-Márquez and Norell 2010; Godefroit et al. 2012a; McDonald et al. 2012). During the last decades, additional hadrosauroid taxa have been discovered in several different Upper Cretaceous units across Europe, revealing that the group was very successful and widespread on the Late Cretaceous European Archipelago (for an overview, see Dalla Vecchia 2006, 2009a, 2014; Prieto-Márquez et al. 2013). Most of these hadrosauroids, however, represent different hadrosaurid subclades more derived than the position reconstructed for Telmatosaurus (e.g., Pereda-Suberbiola et al. 2009; Cruzado-Caballero et al. 2010; Prieto-Márquez et al. 2013, 2019), with the notable exception of Tethyshadros from northeastern Italy (Dalla Vecchia 2009b) and Fylax from northeastern Spain (Prieto-Márquez and Carrera Farias 2021).
Despite being a relatively well-known taxon, there has been considerable debate over the referral of isolated and disarticulated remains to Telmatosaurus, primarily involving the other ornithopod dinosaur known from the Haţeg Basin, the rhabdodontid Zalmoxes (Weishampel et al. 1993, 2003; Brusatte et al. 2017). In fact, Zalmoxes was the second taxon that was described in detail by Nopcsa, although he originally referred the rhabdodontid material from the Haţeg Basin to Mochlodon and subsequently to Rhabdodon (Nopcsa 1902a, 1904, 1915, 1928). Only later, Weishampel et al. (2003) erected the new genus Zalmoxes for the rhabdodontid material from the Transylvanian area with the type species Z. robustus and its newly recognized sister-species Z. shqiperorum. The Rhabdodontidae is an endemic clade of Cretaceous European basal iguanodontians that are among the most common and abundant terrestrial vertebrates known to have inhabited the Late Cretaceous European Archipelago (e.g., Buffetaut and Le Loeuff 1991; Pereda-Suberbiola 1999; Weishampel et al. 2003; Ősi et al. 2012; Csiki-Sava et al. 2015; Dieudonné et al. 2016; Godefroit et al. 2017; Párraga and Prieto-Márquez 2019).
A recent re-investigation of the rhabdodontid cranial material from the Haţeg Basin also led to the re-examination of two partial ornithopod braincases that have been referred to Zalmoxes in the past. The first of these, UBB NVZ1-42 has been discovered from the Râul Mare River section near Nălaţ-Vad and was referred to Zalmoxes shqiperorum (Godefroit et al. 2009). The second, NHMUK R.3401A, is a historical Nopcsa specimen that was collected at the beginning of the twentieth century from the Sibişel Valley section of the Sînpetru Formation near Sânpetru. The latter specimen was, together with most of the original Nopcsa collection, later purchased by the Natural History Museum of London, where it is still deposited today. We here re-describe these specimens and, based on a through comparison with hadrosauroids as well as basal iguanodontians, we reject their rhabdodontid affinities and re-assign them tentatively to the hadrosauroid dinosaur Telmatosaurus.
Geological setting
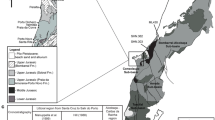
The intramontane Haţeg Basin is located in the southwestern Carpathians, western Romania (Fig. 1a). Upper Cretaceous sedimentary rocks crop out mainly in the southcentral, central, southeastern and northwestern parts of the basin (Fig. 1b). The strata cropping out along the Sibişel Valley in the south-central parts of the Haţeg Basin represent the stratotype section of the Sînpetru Formation and yielded the quasi-totality of the original finds of Franz Nopcsa. The deposits in the northwestern parts of the basin have been referred to the Densuş-Ciula Formation, which differs from the Sînpetru Formation mainly in its higher content of volcanoclastic sediments mixed with the siliciclastics. Although other uppermost Cretaceous continental deposits in the central and southeastern parts of the basin are of a roughly similar age and yielded an overall similar vertebrate fauna with these two formally defined units, their precise correlation with either of the abovementioned formations has been rather contentious and they are often simply referred to as the ‘Râul Mare River section’ and the ‘Pui Beds’, respectively (see Csiki-Sava et al. 2016).

Geological setting of the specimens UBB NVZ1-42 and NHMUK R.3401A. a Position of the study area within Romania. b Simplified geological map of the Haţeg Basin, highlighting the distribution of the uppermost Cretaceous continental deposits. Stars mark the origin of the braincases re-identified here as of hadrosauroid affinities: 1—approximate location of Nopcsa's Quarry 1, yielding specimen NHMUK R.3401A, 2—location of the fossiliferous pocket yielding specimen UBB NVZ1-42 (site 5 in Smith et al. 2002). 1—Pre-Alpine crystalline basement units surrounding the Haţeg Basin; 2—outcropping areas of pre-Quaternary sedimentary rocks of the basin, with 3–5 highlighting the distribution of the uppermost Cretaceous continental beds: 3—Sînpetru Formation; 4—deposits tentatively correlated with the Sînpetru Formation (see Csiki-Sava et al. 2016); 5—Densuş-Ciula Formation (v—volcanoclastic subunit); 6—Quaternary deposits; 7—stars mark the origin of the braincases re-identified here as of hadrosauroid affinities (see above). c View of the right-side flank of the Sibişel Valley south of Sânpetru, with exposures of the stratotype Sînpetru Formation. Approximate location of Nopcsa’s Quarry 1 can be constrained to somewhere in the middle part of the section represented in the photo. d Overview of the Nălaţ-Vad locality, looking towards north. The approximate position of site (fossiliferous lens) NV5 (see Smith et al. 2002) pointed by arrow
The first specimen re-described and re-assigned here (UBB NVZ1-42) was discovered in the Râul Mare River section near Nălaţ-Vad in the central part of the Haţeg Basin (Fig. 1b). The rocks of the Râul Mare River section crop out in, and along, the river Râul Mare and are strongly tilted, dipping almost vertically. The succession comprises siliciclastic sedimentary rocks that were likely deposited in a meandering river system (Therrien 2004; Săsăran et al. 2011). In the past, these deposits have been correlated with the upper part of the type Sînpetru Formation (Codrea et al. 2002; Therrien 2006; Therrien et al. 2009), or else were assigned tentatively to the Densuş-Ciula Formation (Panaiotu et al. 2011), or to a separate lithostratigraphic unit (Csiki-Sava et al. 2016). The age of the succession has been estimated as ‘middle’ (i.e., close to the early/late Maastrichtian boundary; Van Itterbeeck et al. 2005) or late Maastrichtian (Panaiotu et al. 2011; Ciobanete et al. 2011). Only relatively recently have systematic excavations along the Râul Mare revealed rich and diverse vertebrate assemblages comprising amphibians, turtles, squamates, pterosaurs, different dinosaurs including birds and multituberculate mammals as well as dinosaur eggs (Smith et al. 2002; Codrea et al. 2002; Godefroit et al. 2009; Csiki et al. 2010a; Wang et al. 2011; Csiki-Sava et al. 2016; Brusatte et al. 2017; Mayr et al. 2020). Some of the most important finds occurred in so-called fossil pockets, local accumulations of vertebrate fossils that are sometimes associated or even partially articulated such as the holotype of the titanosaur Paludititan (Csiki et al. 2010a). The braincase described here, UBB NVZ1-42, was recovered from a fossil pocket that almost exclusively yielded disarticulated rhabdodontid remains assigned to Zalmoxes shqiperorum and assumed to represent a single individual (Godefroit et al. 2009). Notably, however, a left quadrate was found in the same fossil pocket that apparently does not belong to a rhabdodontid and instead is probably assignable to the hadrosauroid Telmatosaurus (Godefroit et al. 2009), thus indicating the presence of at least one more taxon in this fossil pocket. The presence of more than one taxon within the commonly occurring fossiliferous pockets from the uppermost Cretaceous of the Haţeg Basin is a widespread phenomenon (e.g., Nopcsa 1902b; Csiki et al. 2010b), and the case for such a multitaxic composition would be strengthened now for the NVZ1 fossil pocket as well by our re-assignment of the basicranium UBB NVZ1-42 to Telmatosaurus as argued below.
The second specimen that is re-examined here (NHMUK R.3401A) was collected by Franz Nopcsa from the Sibişel Valley section near Sânpetru in the south-central part of the Haţeg Basin (Fig. 1b). This sedimentary succession is exposed along a 2.5 km long natural valley carved by the Sibişel River and exposing an 860 m thick sequence of terrestrial Upper Cretaceous sediments. The sedimentary rocks comprise coarse-grained and fine-grained layers arranged in multiple fining-upward sequences, which were laid down on a poorly channelized alluvial plain drained by braided river systems (Grigorescu 1983; Therrien 2006; Therrien et al. 2009). Stratigraphically, the Sibişel Valley section represents the stratotype section of the Sînpetru Formation and has been estimated to be early to early late Maastrichtian in age (Therrien 2004; Panaiotu and Panaiotu 2010). The Sibişel Valley section includes many classical Nopcsa localities and has ever since yielded rich and diverse vertebrate assemblages comprising fishes, amphibians, turtles, squamates, crocodyliforms, pterosaurs, dinosaurs and multituberculate mammals (Nopcsa 1900, 1902a, b, 1904, 1923, 1929; Andrews 1913; Grigorescu 1983; Csiki and Grigorescu 1998; Csiki et al. 2010b; Martin et al. 2014; Csiki-Sava et al. 2016; Augustin et al. 2021). The fossil vertebrate remains often show complex taphonomic histories and can occur isolated, associated and partly articulated or in local multitaxic concentrations, so-called fossil pockets (Nopcsa 1902b; Grigorescu 1983; Csiki et al. 2010b; Augustin et al. 2019). The second specimen re-described here, NHMUK R.3401A, was discovered in such a multitaxic fossil concentration, identified as Quarry 1 by Nopcsa, together with different skull and postcranial bones of both rhabdodontids and hadrosauroids among others (Nopcsa 1904, and comments below).
Materials and methods
Two specimens are re-described herein. The first, UBB NVZ1-42, is a partial basicranium that was recovered together with other vertebrate remains (see below) in 2002 from the Râul Mare River section, near Nălaţ-Vad (Godefroit et al. 2009); the specimen is currently housed at the Palaeontological Collection of the Babeș-Bolyai University. The second specimen, NHMUK R.3401A, consists of a partial braincase and was collected by Franz Nopcsa from his ‘Quarry 1’ (see below), which he discovered in 1895 on the eastern side of the Sibişel Valley section near Sânpetru (Nopcsa 1902a, b; Grigorescu 2010); the specimen is currently stored at the Natural History Museum London. Both specimens were digitalized using the photogrammetry technique detailed by Mallison and Wings (2014) as well as the software Agisoft Photoscan Professional, in order to create surface models. In addition, 3D prints were produced based on the surface models, which are deposited in the Palaeontological Collection of the University of Tübingen.
Institutional abbreviations. LPB (FGGUB)—Laboratory of Paleontology, Faculty of Geology and Geophysics, University of Bucharest, Bucharest, Romania; NHMUK—Natural History Museum, London, UK; UBB—Babeș-Bolyai University, Cluj-Napoca, Romania.
Systematic Palaeontology
Dinosauria Owen (1842)
Ornithischia Seeley (1888)
Ornithopoda Marsh (1881)
Iguanodontia Baur (1891)
Hadrosauroidea Cope (1869)
Telmatosaurus transsylvanicus Nopcsa (1900)
Holotype. NHMUK R.3386, an almost complete but crushed skull.
Referred material. UBB NVZ1-42, a partial basicranium composed of the basioccipital, the partial exoccipital–opisthotic complexes, and the basisphenoid–parasphenoid complex from the Râul Mare River section, near Nălaţ-Vad, central Haţeg Basin, as well as NHMUK R.3401A, the posterior part of a skull composed of the basioccipital, both exoccipital–opisthotic complexes, the posterior part of the basisphenoid, and the supraoccipital from the Sibişel Valley section near Sânpetru, southern Haţeg Basin.
Remarks on UBB NVZ1-42. Specimen UBB NVZ1-42 has been previously referred to the rhabdodontid ornithopod dinosaur Zalmoxes shqiperorum (Godefroit et al. 2009). This referral was primarily based on the association of UBB NVZ 1-42 with other disarticulated bones in the ‘third fossil pocket’ at the Nălaţ-Vad locality, remains that almost exclusively belong to Zalmoxes shqiperorum and apparently represent a single individual (Godefroit et al. 2009). Recent investigations of the rhabdodontid fossils from the Haţeg Basin also led to a re-examination of UBB NVZ 1-42 and, based on comparison with several hadrosauroid and rhabdodontid basicrania from the Haţeg Basin as outlined below, we here confidently identify UBB NVZ1-42 as a hadrosauroid basicranium and refer it to the only known local hadrosauroid taxon, Telmatosaurus. Interestingly, Godefroit et al. (2009) also reported the presence of a second type of quadrate from the same fossil pocket, which they did not refer to Zalmoxes shqiperorum but instead noted that it closely resembles in morphology that of Telmatosaurus. In the original description, Godefroit et al. (2009) referred to the braincase as UBB NVZ1-40, although this number actually identifies a rhabdodontid left quadrate from this fossil site. The assignment of UBB NVZ 1-42 to Zalmoxes shqiperorum has previously been questioned by Brusatte et al. (2017), who also noted that it might instead belong to Telmatosaurus.
Remarks on NHMUK R.3401A. The braincase specimen NHMUK R.3401A is part of the original Nopcsa Collection that was later purchased by the Natural History Museum in London (e.g., Dalla Vecchia 2009a). As already pointed out by Dalla Vecchia (2009a), and as with most registry entries of the Nopcsa collection, the inventory number NHMUK R.3401 identifies a large number of different specimens, belonging to several individuals that actually represent more than one taxon. Furthermore, skeletal remains included under this specimen number are assumedly originating from individuals whose other (usually postcranial) remains are now registered under different entry numbers (for a more detailed discussion of this issue, see Dalla Vecchia 2009a: pp. 6–7).
Intriguingly, the same fossil pocket, from which braincase NHMUK R.3401A has been recovered, also yielded disarticulated, but fairly complete and definitively diagnostic skull bones, including dentaries, of both Telmatosaurus transsylvanicus and Zalmoxes robustus. These specimens had been lumped together under the same original specimen number NHMUK 3401, from which at one point certain specimens, some showing definitive hadrosauroid characters and also including the partial braincase discussed here, have been separated as NHMUK 3401A. It is worth noting that the fossil pocket that yielded this mixed material (mentioned as Quarry 1 in the NHMUK registry book) contains a similar mix of rhabdodontid and hadrosauroid remains just as the one reported for the (smaller) Zalmoxes-lens from Nălaţ-Vad from where the partial braincase UBB NVZ1-42 originates. This mixture of disarticulated skeletal remains belonging to different individuals and representing different taxa is a hallmark feature of the so-called ‘fossiliferous pockets’ that represent one of the main types of fossil occurrences in the Haţeg Basin (Nopcsa 1914; Csiki et al. 2010b), one for which Quarry 1 of Nopcsa is among the first and best examples.
These complications, together with the absence of original field notes or quarry maps to document the discovery conditions of the different specimens excavated by Nopcsa at Sânpetru, seriously hindered the understanding of the taxonomic identity and potential skeletal association of the specimens that make up this mixed fossil assemblage, including that of the partial braincase NHMUK R.3401A discussed here. Indeed, this specimen has been referred to Telmatosaurus transsylvanicus by Weishampel et al. (1993), who also used it to reconstruct the braincase anatomy of that taxon (Weishampel et al. 1993: Fig. 2). This taxonomic choice appears to be in accordance with Nopcsa’s original taxonomic identification of this specimen (see Nopcsa 1904, Table 1) who reported it as belonging to a second individual (Individual B) of Telmatosaurus (see also Dalla Vecchia 2009a). Subsequently, however, the specimen was re-assigned to Zalmoxes robustus by Weishampel et al. (2003), albeit without providing any arguments in favor of this view. Similar to UBB NVZ1-42, recent investigations of the rhabdodontid material from the Haţeg Basin also led to a re-examination of NHMUK R.3401A, and this resulted in the re-interpretation of this specimen as belonging to Telmatosaurus transsylvanicus, as originally proposed by Nopcsa (1904), respectively by Weishampel et al. (1993).
Description of UBB NVZ1-42
The specimen UBB NVZ1-42 includes the complete basioccipital, the partial exoccipital–opisthotic complexes, and most of the basisphenoid–parasphenoid complex (Fig. 2). Aside from the missing parts, the basicranium is well-preserved with delicate processes and small foramina still present. The occipital condyle is u-shaped to kidney-shaped in posterior view and composed of the basioccipital ventrally and the exoccipital–opisthotic complexes dorso-laterally (Fig. 2a). The suture between the basioccipital and the exoccipital–opisthotic complexes is not discernible. In ventral view, the occipital condyle is somewhat rectangular in outline with a convex surface (Fig. 2b). In lateral view, the occipital condyle is ellipsoidal and exhibits three large foramina, two openings next to each other for cranial nerve XII (hypoglossal nerve) and cranial nerves X (vagus nerve) and XI (accessory nerve), respectively, as well as one foramen located antero-ventrally to the latter, possibly for the exit of the jugular vein (Fig. 2c). Immediately above the opening for cranial nerve XII, the braincase is broken and the paroccipital process (mainly composed of the opisthotic) is largely missing, although the bone gets markedly thinner just above the opening for cranial nerve XII, likely indicating the ventral part of the paroccipital process.

Partial ornithopod basicranium, UBB NVZ1-42, here tentatively referred to the hadrosauroid dinosaur Telmatosaurus transsylvanicus, from the Upper Cretaceous of Nălaţ-Vad, Haţeg Basin. a Posterior view. b Ventral view. c Left lateral view. d Right lateral view. e Anterior view. f Dorsal view. All figures to the same scale. ap, alar process; bo, basioccipital; bg, groove on the lateral aspect of the basisphenoid; bp, basisphenoid platform on the ventral aspect of the basisphenoid; bs, basisphenoid–parasphenoid complex; ca, carotid artery; de, depression on the ventral aspect of the basicranium between the sphenoccipital tubercles; ds, dorsum sellae; ef, endocranial floor; ex, exoccipital; fc, foramen in the pituitary fossa connecting the carotid artery; fm, foramen magnum; jv, jugular vein; pf, pituitary fossa; pp, paroccipital process formed by the exoccipital–opisthotic complex; st, sphenoccipital tubercle; vi, cranial nerve six (abducens nerve); x, cranial nerve ten (vagus nerve); xi, cranial nerve eleven (accessory nerve); xii, cranial nerve twelve (hypoglossal nerve)
On the ventral aspect of the braincase, directly adjacent to the basioccipital condyle, there are two round processes lying anteriorly and slightly laterally to the condyle (Fig. 2b). These processes, the sphenoccipital tubercles or basal tubera, are directed ventro-laterally and have a deep depression between them that is roughly triangular in outline. The suture between the basioccipital and the basisphenoid is hardly visible on the ventral aspect of the braincase but is probably present as a shallow indentation running across the sphenoccipital tubercles, as this is also the position of the suture in other hadrosauroids (e.g., Sues and Averianov 2009: p. 2551; Prieto-Márquez 2010: p. 847; McDonald et al. 2012: pp. 16–17; Xing et al. 2017: pp. 24–25). The basisphenoid, in turn, is completely fused to the parasphenoid anteriorly, forming the basisphenoid–parasphenoid complex. The sphenoccipital tubercles are connected to the antero-ventral part of basisphenoid through a ridge rising anteriorly and eventually merging with the antero-ventral part of basisphenoid. The antero-ventral part of the basisphenoid forms a transverse surface that mainly extends ventrally, approximately perpendicular to the long axis of the braincase. In ventral view, this antero-ventral part of the basisphenoid forms a slightly concave platform that is inclined antero-dorsally and would be connected to the missing basipterygoid processes.
In lateral view, there is a well-developed groove anterior to the sphenoccipital tubercles that extends antero-dorsally, is bordered by the alar process of the basisphenoid anteriorly and accommodates the latero-ventral entrance of the carotid artery in its lower half (Fig. 2c–d). The alar process is well preserved and forms a thin ridge extending mainly latero-posteriorly from the lateral side of the basisphenoid. Anterior to the alar process and near its upper margin is a small foramen, representing the opening for cranial nerve VI (abducens nerve). The anterior portion of the basisphenoid is incompletely preserved and wedge-shaped. In anterior view, a dorso-ventrally elongated pituitary fossa is visible (Fig. 2e), which exhibits two small foramina in its postero-ventral part that are likely connected to the latero-ventral entrance of the carotid artery (Xing et al. 2017: p. 25). The endocranial floor is completely visible in dorsal view (Fig. 2f) and is formed by the basioccipital and exoccipitals posteriorly and by the basisphenoid–parasphenoid complex anteriorly. The endocranial floor is relatively straight and has a constant width from the foramen magnum posteriorly to the anterior part of the preserved element. The dorsum sellae is located in the anterior part, more or less dorsally to the latero-ventral entrance of the carotid artery, and exhibits two foramina for the cranial nerve VI (abducens nerve). The lateral walls of the endocranial floor exhibit the suture between the basisphenoid–parasphenoid complex and the (missing) prootic.
Description of NHMUK R.3401A
The specimen NHMUK R.3401A comprises the basioccipital, both exoccipital–opisthotic complexes, the supraoccipital, and the anterior part of the basisphenoid–parasphenoid complex (Fig. 3). Aside from the missing parts, the specimen is relatively well-preserved. In posterior view, the occipital condyle is kidney-shaped and consists of the basioccipital ventrally and the exoccipitals dorso-laterally (Fig. 3a). On the right side of the occipital condyle, the suture between the basioccipital and the exoccipital is visible but slightly damaged. On the left side, the occipital condyle is weathered, obliterating the sutural contact between the basioccipital and the exoccipital. Both the basioccipital and the exoccipitals take part in the formation of the endocranial floor. The exoccipitals are completely fused to the opisthotics, forming the exoccipital–opisthotic complex (or paroccipital processes), which continues as an elongated and flat element from the triangular and bulbous ventral part that is forming the dorso-lateral corner of the occipital condyle.

Posterior part of an ornithopod skull, NHMUK R.3401A, here tentatively referred to the hadrosauroid dinosaur Telmatosaurus transsylvanicus, from the Upper Cretaceous Sînpetru Formation near Sânpetru, Haţeg Basin. a Posterior view. b Ventral view. c Left lateral view. d Right lateral view. e Anterior view. f Dorsal view. All figures to the same scale. bo, basioccipital; bg, groove on the lateral aspect of the basisphenoid; bs, basisphenoid–parasphenoid complex; de, depression on the ventral aspect of the basicranium between the sphenoccipital tubercles; ef, endocranial floor; ex, exoccipital; fm, foramen magnum; pp, paroccipital process formed by the exoccipital–opisthotic complex; st, sphenoccipital tubercle
The exoccipital–opisthotic complex is extending mainly dorsally and, to a much lesser degree, laterally, forming the lateral walls of the foramen magnum (Fig. 3a, b). In posterior view, the foramen magnum has a dorso-ventrally elongated, ellipsoidal shape. Dorsally, the exoccipital–opisthotic complex broadens and makes a postero-lateral curve (Fig. 3c–e). Medially, the dorsal part of the exoccipital–opisthotic complex is fused to the supraoccipital, the latter forming the dorsal margin of the foramen magnum. The supraoccipital is a roughly triangular to trapezoidal element in posterior view, wedged between the exoccipital–opisthotic complexes laterally. The dorsal part of the supraoccipital is relatively flat and extends antero-dorsally (Fig. 3f). The dorso-lateral and anterior parts of the exoccipital–opisthotic complexes as well as the anterior part of the supraoccipital are missing. However, the paroccipital processes clearly extend posteriorly beyond the level of the basioccipital and the exoccipitals in dorsal view, while the supraoccipital is lying anteriorly to the occipital condyle.
In ventral view, the basioccipital is rectangular in outline with a convex surface, although the left part of the basioccipital is missing and thus its morphology cannot be completely assessed (Fig. 3b). Directly anterior to the basioccipital, there are two bulbous processes on the ventral aspect of the braincase, the sphenoccipital tubercles (= basal tubera), which extend mainly ventro-laterally (Fig. 3e). The suture between the basioccipital and the basisphenoid is hardly visible but it probably runs across the sphenoccipital tubercles as indicated by a shallow, sinuous indentation on the right sphenoccipital tubercle; this is also the position of the suture between basioccipital and basisphenoid in other hadrosauroids (e.g., Sues and Averianov 2009: p. 2551; Prieto-Márquez 2010: p. 847; McDonald et al. 2012: pp. 16–17; Xing et al. 2017: pp. 24–25), as well as in NVZ1-42 discussed previously. Between the sphenoccipital tubercles, there is a deep depression. Anterior to the sphenoccipital tubercles, the basisphenoid rises markedly ventrally but the antero-ventral part of the basisphenoid is broken. The sphenoccipital tubercles are connected to the anterior and ventrally rising part of the basisphenoid through narrow ridges.
In lateral view, the occipital condyle formed by the basioccipital and the exoccipitals is slightly convex (Fig. 3c, d). A shallow depression is present anterior to the sphenoccipital tubercles that likely represents the dorsal part of the groove on the lateral side of the basisphenoid that is bordered by the alar process anteriorly (see above). The openings for the cranial nerves that are usually visible in lateral view (see above) are not discernible because the bone surface is imperfectly preserved and some missing parts, just dorsal of the exoccipitals, have been restored with plaster. The anterior part of the braincase is not preserved.
Comparison
Both UBB NVZ1-42 and NHMUK R.3401A have been referred to the rhabdodontid dinosaur Zalmoxes in the past (Weishampel et al. 2003; Godefroit et al. 2009), although the latter specimen has originally been assigned to the hadrosauroid Telmatosaurus (Nopcsa 1904: tab. 1; Weishampel et al. 1993). Therefore, the following discussion focuses primarily on comparisons of these two specimens with other hadrosauroid and rhabdodontid basicrania from the Haţeg Basin. In addition, a short comparison to other hadrosauroids and basal iguanodontians is also provided.
Comparison with Telmatosaurus and other hadrosauroids
For Telmatosaurus, two undisputed basicrania are known, both of which come from the Sînpetru Formation of the Sibişel Valley section near Sânpetru, southern Haţeg Basin. One is part of the largely complete but crushed holotype skull NHMUK R.3386 (Fig. 4a) that was described and figured by Nopcsa (1900); it is noteworthy that this skull was explicitly mentioned as the basis for the erection of the genus (Nopcsa 1900: 559; contra Dalla Vecchia 2009a). The second, NHMUK R.3387 (Fig. 4b), is an isolated basicranium, comprising the basioccipital and the basisphenoid that was described and figured in the same publication as the holotype skull (Nopcsa 1900). Both of these basicrania described by Nopcsa (1900) are extremely similar to the specimens described here (UBB NVZ1-42 and NHMUK R.3401A)—in fact, they are nearly indistinguishable, aside from the different preservational states of the skulls and the different sizes. These similarities are most apparent in ventral view, also because this is the best-preserved part in all four specimens (Fig. 4). Both the holotype of Telmatosaurus (NHMUK R.3386) and the referred basicranium (NHMUK R.3387) described by Nopcsa (1900), strongly resemble the specimens described here (UBB NVZ1-42 and NHMUK R.3401A), as they have a short, convex basioccipital that is slightly rectangular in ventral view and directly connected to a pair of sphenoccipital tubercles (= basal tubera) anteriorly, which have a well-developed depression between them (Fig. 4; Nopcsa 1900: pl. 3). In addition, all four specimens share a medio-laterally narrow basisphenoid, which lies immediately anterior to the depression, is steeply rising ventrally, and is connected to the sphenoccipital tubercles through a narrow, antero-medially extending ridge. In the holotype of Telmatosaurus (NHMUK R.3386), the sphenoccipital tubercles are slightly more splayed and farther apart than in the other basicrania referred to Telmatosaurus, in which they are closer to each other. Moreover, the central depression between the sphenoccipital tubercles is narrower in NHMUK R.3387 as compared to the other specimens. In all specimens, the foramen magnum opens into a relatively straight endocranial floor.

Comparison of the ornithopod basicrania described herein (UBB NVZ1-42 and NHMUK R.3401A) with the holotype skull (NHMUK R.3386) and the referred isolated basicranium (NHMUK R.3387) of Telmatosaurus transsylvanicus, all from the Upper Cretaceous of the Haţeg Basin. All specimens are displayed in ventral view but are not to the same scale. a Surface model of the holotype skull of Telmatosaurus, NHMUK R.3386 from the Sînpetru Formation near Sânpetru, south-central Haţeg Basin. a’ The same specimen as in A with texture. b Surface model of the isolated Telmatosaurus basicranium NHMUK R.3387 from the Sînpetru Formation near Sânpetru, south-central Haţeg Basin. b’ The same specimen as in B with texture. c Surface model of the basicranium UBB NVZ1-42 from the Râul Mare River section, near Nălaţ-Vad, central Haţeg Basin. c’ The same specimen as in C with texture. d Surface model of the posterior skull NHMUK R.3401A from the Sînpetru Formation near Sânpetru, south-central Haţeg Basin. d’ The same specimen as in D with texture. Scales equal 1 cm. bo, basioccipital de, depression on the ventral aspect of the basicranium between the sphenoccipital tubercles; st, sphenoccipital tubercle
The basicranium anatomy of hadrosauroids is well known and has been described for many different taxa. Overall, an antero-posteriorly short basioccipital, bulbous sphenoccipital tubercles lying immediately anterior to the basioccipital, and a depression between the sphenoccipital tubercles are typically present in basal hadrosauroids, including Bactrosaurus johnsoni (Gilmore 1933: Fig. 25), Batyrosaurus rozhdestvenskyi (Godefroit et al. 2012a: Fig. 20.2), Eolambia caroljonesa (McDonald et al. 2012: Fig. 20), Gobihadros mongoliensis (Tsogtbaatar et al. 2019: Fig. 8), Levnesovia transoxiana (Sues and Averianov 2009: Fig. 1), Ouranosaurus nigeriensis (Taquet 1976: Fig. 12), Plesiohadros djadokhtaensis (Tsogtbaatar et al. 2014: Fig. 7.8), Proa valdearinnoensis (pers. com. Fabien Knoll), Sirindhorna khoratensis (Shibata et al. 2015: Fig. 2), Tanius sinensis (Wiman 1929: pl. 5), and Tethyshadros insularis (Chiarenza et al. 2021: Fig. 2, p. 5). Additionally, the same general bauplan of the braincase is also seen in more derived hadrosaurids such as Acristavus gagslarsoni (Gates et al. 2011: Fig. 9), Amurosaurus riabinini (Godefroit et al. 2004: Fig. 7), Arenysaurus ardevoli (Pereda-Suberbiola et al. 2009; Fig. 3; Cruzado-Caballero et al. 2015: suppl. 1), Brachylophosaurus canadensis (Prieto-Márquez 2005: Fig. 6), Edmontosaurus regalis (Lull and Wright 1942: Fig. 4; Xing et al. 2017: Fig. 10), Eotrachodon orientalis (Prieto-Márquez et al. 2016: Fig. 16), Gryposaurus notabilis (Ostrom 1961: Fig. 11; Prieto-Márquez 2010: Fig. 4), Gryposaurus monumentensis (Gates and Sampson 2007: Fig. 11), Hypacrosaurus altispinus (Evans 2010: Figs. 10, 11), Maiasaura peeblesorum (McFeeters et al. 2021: Fig. 13), Olorotitan arharensis (Godefroit et al. 2012b: Fig. 4), Ornatops incantatus (McDonald et al. 2021: Fig. 7), Parasaurolophus cyrtocristatus (Gates et al. 2021: figs. 5, 9, 13), Parasaurolophus tubicen (Sullivan and Williamson 1999: Fig. 8), Sahaliyania elunchunorum (Godefroit et al. 2008: Fig. 3), and Secernosaurus koerneri (Prieto-Márquez and Salinas 2010: figs. 6, 7). Therefore, the basicranial morphology of these different hadrosauroids closely resembles that of the holotype and referred basicrania of Telmatosaurus (NHMUK R.3386 and NHMUK R.3387), but also that seen in the taxonomically contentious Romanian braincase specimens described above (UBB NVZ1-42 and NHMUK R.3401A).
Comparison with rhabdodontids and other basal iguanodontians
Four definitively identified rhabdodontid basicrania are currently reported from the Upper Cretaceous of the Haţeg Basin. The first two of these, NHMUK R.3408 and NHMUK R.3409, were excavated, apparently isolated according to Nopcsa (1904), more than a century ago in the stratotype Sînpetru Formation along the Sibişel Valley, near Sânpetru. These specimens were described and figured by Nopcsa (1904) who referred them tentatively to the rhabdodontid Mochlodon (= Zalmoxes). NHMUK R.3408 (Fig. 5a) comprises the basioccipital and most of the basisphenoid, whereas NHMUK R.3409 (Fig. 5b) only preserves the region around the basal tubera. A third, relatively complete basicranium, LPB (FGGUB) R.1723, was recovered much later (2000) from the middle part of the Densuş-Ciula Formation at the Tuştea-Oltoane nesting site, in the northwestern part of the Haţeg Basin (see Botfalvai et al. 2017, for a recent review of the locality). It comprises the basioccipital and most of the basisphenoid and has been described and figured by Weishampel et al. (2003: Fig. 11). The last rhabdodontid basicranium specimen that was referred previously to the Rhabdodontidae, LPB (FGGUB) R.1629 (Fig. 5c), is also from the Tuştea locality, where it was found in 1998. The specimen was briefly mentioned by Weishampel et al. (2003: p. 78) and then was preliminarily illustrated in articulation with the matching left exoccipital LPB (FGGUB) R.1591 by Botfalvai et al. (2017: Fig. 8H), but it has never been thoroughly figured before nor described in detail.

Rhabdodontid basicrania from the Upper Cretaceous of the Haţeg Basin in ventral view. a Surface model of the isolated rhabdodontid basicranium NHMUK R.3408 from the Sînpetru Formation near Sânpetru, south-central Haţeg Basin. a’ The same specimen as in A with texture. b Surface model of the isolated fragmentary rhabdodontid basicranium NHMUK R.3409 from the Sînpetru Formation near Sânpetru, south-central Haţeg Basin. b’ The same specimen as in B with texture. c Surface model of the isolated rhabdodontid basicranium LPB (FGGUB) R.1629 from the Densuş-Ciula Formation near Tuştea, northwestern Haţeg Basin. c’ The same specimen as in C with texture. Scales equal 1 cm. bn, basioccipital neck; bo, basioccipital; bt, basal tubera (= sphenoccipital tubercles); mr, midline ridge on the posterior part of the basisphenoid
All three rhabdodontid specimens comprising the complete basioccipital (NHMUK R.3408, LPB (FGGUB) R.1723 and R.1629), have a distinct, elongated and well-developed neck connecting the occipital condyle with the basal tubera anteriorly (Fig. 5a, c). Moreover, all the rhabdodontid basicrania from the Haţeg Basin lack bulbous sphenoccipital tubercles (= basal tubera) and a median depression on the ventral aspect of the braincase (Figs. 5, 6). Furthermore, the basal tubera join together medially to form a wide, medio-laterally extending ridge that has a wrinkled appearance on the posterior side with a prominent transverse midline ridge (Fig. 5). Therefore, these rhabdodontid basicrania from the Haţeg Basin differ markedly from the corresponding elements of Telmatosaurus described by Nopcsa (1900) (NHMUK R.3386 and NHMUK R.3387), as well as from the basicranial specimens described herein (UBB NVZ1-42 and NHMUK R.3401A). In fact, as already noted, specimens UBB NVZ1-42 and NHMUK R.3401, resemble the Telmatosaurus basicrania described by Nopcsa (1900), including the holotype of that genus, much more closely than those of rhabdodontids.

Simplified drawing comparing the two basicranial specimens described here (UBB NVZ1-42 and NHMUK R.3401A) with the basicranium of the holotype skull of Telmatosaurus transsylvanicus (NHMUK R.3386) and the referred basicranium of Zalmoxes robustus (NHMUK R.3408) in ventral view. a Basicranium of the holotype skull of Telmatosaurus transsylvanicus, NHMUK R.3386. b Basicranium UBB NVZ1-42. c Basicranium of the partial skull NHMUK R.3401A. d Referred basicranium NHMUK R.3408 of Zalmoxes robustus. The drawings are not to the same scale. bn, basioccipital neck; bt, basal tubera (= sphenoccipital tubercles); de, depression between the sphenoccipital tubercles
In contrast, the morphology of these rhabdodontid basicrania from the Haţeg Basin is quite similar to that of the rhabdodontid Rhabdodon from France (Chanthasit 2010: figs. 4.2, 4.3, the only other rhabdodontid for which some information is currently available, in features such as the possession of a distinct basioccipital neck supporting the occipital condyle, the lack of sphenoccipital tubercles, the lack a ventral median depression between these tubercles, and the presence of wide transversal ridge formed by the medially confluent basal tubera. In addition, these same features can be found in a wide variety of other basal iguanodontians, including Anabisetia saldiviai (Coria and Calvo 2002: Fig. 4), Camptosaurus dispar (Gilmore 1909: figs. 4, 5; Carpenter and Lamanna 2015: figs. 7, 8), Cumnoria prestwichii (Hulke 1880; Fig. 1; Galton and Powell 1980; Fig. 1), Dakotadon lakotaensis (Weishampel and Bjork 1989: figs. 4, 5; Boyd and Pagnac 2015: Fig. 8), Dryosaurus altus (Galton 1983: figs. 2, 3, 1989: pl. 2), Dryosaurus elderae (Carpenter and Galton 2018: Fig. 28), Dysalotosaurus lettowvorbecki (Galton 1989: pl. 1, 3; Hübner and Rauhut 2010: Fig. 7), Iguanodon bernissartensis (Norman 1980: figs. 5, 9), Mantellisaurus atherfieldensis (Norman 1986: figs. 7, 10, 18), Owenodon sp. (Galton 2009: Fig. 19), Tenontosaurus tilletti (Galton 1989: Fig. 4), Tenontosaurus dossi (Winkler et al. 1997: figs. 6, 7, 8), and an indeterminate iguanodontian from Australia (Bell et al. 2018: Fig. 8). The wide distribution of this type of basicranial morphology in basal iguanodontians, markedly different from that seen in both more basal and derived (i.e., hadrosaurid) hadrosauroids, suggests that it represents a plesiomorphic feature within the clade—as is also indicated by the presence of this type of basicranium morphology in basal ornithopods such as Hypsilophodon foxii (Galton 2001: figs. 7, 9, 17, 19), and Convolosaurus marri (Andrzejewski et al. 2019: Fig. 10), in basal neornithischians such as Changchunsaurus parvus (Liyong et al. 2010: Fig. 6), and Thescelosaurus neglectus (Galton 1989: pl. 4; Boyd 2014: Fig. 11), as well as in basal ornithischians such as Heterodontosaurus tucki, (Norman et al. 2011: figs. 3, 6, 13, 15), and Lesothosaurus diagnosticus (Porro et al. 2015: Fig. 9).
Discussion and Conclusion
Based on the comparison presented above, we confidently identify both ornithopod basicrania from the Haţeg Basin described herein, UBB NVZ1-42 and NHMUK R.3401A, as hadrosauroid basicrania, and tentatively refer them to Telmatosaurus transsylvanicus, the only currently recognized hadrosauroid taxon from the Transylvanian area. In fact, both specimens are nearly indistinguishable from the holotype of that species (Fig. 4a, Nopcsa 1900: pl. 3). In addition, both UBB NVZ1-42 and NHMUK R.3401A exhibit several features that are only present in hadrosauroids and absent in more basal iguanodontian ornithopods. These features include: (i) an antero-posteriorly short basioccipital lacking a distinct neck; (ii) the presence of two well-developed bulbous processes, the sphenoccipital tubercles (= basal tubera) on the ventral aspect of the braincase that are directly positioned anterior to the basioccipital; and (iii) a deep depression on the ventral aspect of the braincase between the sphenoccipital tubercles. Conversely, characters that are typically present in rhabdodontids and other basal iguanodontians, including a distinct and elongated basioccipital neck and the basal tubera forming a medio-laterally extending wide ridge, are absent in UBB NVZ1-42 and NHMUK R.3401A.
The re-assignment of UBB NVZ1-42 to Telmatosaurus as advocated herein also requires a revision of the most recent diagnosis of Zalmoxes shqiperorum proposed by Godefroit et al. (2009) because it was partly based on features of UBB NVZ1-42—the only basicranium referred to this species to date. More specifically, one autapomorphy that has been proposed for Zalmoxes shqiperorum—i.e., an ‘occipital condyle not separated from the sphenooccipital tubercles (= basal tubera) by a distinct neck’ (Godefroit et al. 2009: p. 528)—has to be removed from the species diagnosis. Remarkably, these authors also noted that this character is not seen in any other basal iguanodontian (Godefroit et al. 2009: pp. 546–548) and that is in fact more similar to the morphology usually seen in hadrosaurids (Godefroit et al. 2009: p. 534), which would make this character highly derived within Rhabdodontidae. With the taxonomic re-interpretation of the basicranium UBB NVZ1-42 as proposed here, the diagnosis of Z. shqiperorum can be accordingly emended, and the autapomorphically derived basicranial morphology of this taxon that was previously suggested to differentiate it from its sympatric sister-species Z. robustus, can be rejected. As a consequence, the basicranial morphology of the genus Zalmoxes (currently only documented in Z. robustus) seems to be rather conservative, reminiscent to that seen in basal iguanodontians in general, and this plesiomorphic morphology appears to be typical of rhabdodontids as it also occurs in the closely related Rhabdodon.
Interestingly, several of the differences noted here between the hadrosauroid type of braincase morphology and that of more primitive iguanodontians were already pointed out by Nopcsa (1904), in his description of additional skull remains attributable to Mochlodon (= Zalmoxes). There, he compared the basicranium of ‘Mochlodon’ with that of Telmatosaurus, and even figured basicrania referred to these genera (NHMUK R.3408 and NHMUK R.3387, respectively) in ventral view (Nopcsa 1904: figs. 1, 2), noting the much more elongated basioccipital of the rhabdodontid basicranium and the different morphology of the basal tubera, among other aspects (Nopcsa 1904: pp. 244–245).
Our re-assignment of the specimens UBB NVZ1-42 and NHMUK R.3401A to Telmatosaurus transsylvanicus doubles the number of known partial braincases for that genus but significantly reduces the number of previously reported rhabdodontid basicrania from the Haţeg Basin from six to only four. The comparisons provided herein also demonstrate the presence of several important morphological differences between the basicranium of hadrosauroids and that of rhabdodontids, which allows for the confident assignment of even isolated specimens in the future.
References
Andrews, C.W. 1913. On some bird remains from the Upper Cretaceous of Transsylvania. Geological Magazine 10: 193–196. https://doi.org/10.1017/S0016756800126196.
Andrzejewski, K.A., D.A. Winkler, and L.L. Jacobs. 2019. A new basal ornithopod (Dinosauria: Ornithischia) from the Early Cretaceous of Texas. PLoS ONE 14: e0207935. https://doi.org/10.1371/journal.pone.0207935.
Augustin, F.J., A.T. Matzke, Z. Csiki-Sava, and H.-U. Pfretzschner. 2019. Bioerosion on vertebrate remains from the Upper Cretaceous of the Haţeg Basin, Romania and its taphonomic implications. Palaeogeography, Palaeoclimatology, Palaeoecology 534: 109318. https://doi.org/10.1016/j.palaeo.2019.109318.
Augustin, F.J., Z. Csiki-Sava, A.T. Matzke, G. Botfalvai, and M. Rabi. 2021. A new latest Cretaceous pleurodiran turtle (Testudinata: Dortokidae) from the Haţeg Basin (Romania) documents end-Cretaceous faunal provinciality and selective survival during the K-Pg extinction. Journal of Systematic Palaeontology 19: 1059–1081. https://doi.org/10.1080/14772019.2021.2009583.
Baur, G. 1891. Remarks on the reptiles generally called Dinosauria. American Naturalist 25: 434–454.
Bell, P.R., M.C. Herne, T. Brougham, and E.T. Smith. 2018. Ornithopod diversity in the Griman Creek Formation (Cenomanian), New South Wales, Australia. PeerJ 6: e6008. https://doi.org/10.7717/peerj.6008.
Botfalvai, G., Z. Csiki-Sava, D. Grigorescu, and Ş Vasile. 2017. Taphonomical and palaeoecological investigation of the Late Cretaceous (Maastrichtian) Tuştea vertebrate assemblage (Romania; Haţeg Basin) - insights into a unique dinosaur nesting locality. Palaeogeography, Palaeoclimatology, Palaeoecology 468: 228–262. https://doi.org/10.1016/j.palaeo.2016.12.003.
Boyd, C.A. 2014. The cranial anatomy of the neornithischian dinosaur Thescelosaurus neglectus. PeerJ 2: e669. https://doi.org/10.7717/peerj.669.
Boyd, C.A., and D.C. Pagnac. 2015. Insight on the anatomy, systematic relationships, and age of the Early Cretaceous ankylopollexian dinosaur Dakotadon lakotaensis. PeerJ 3: e1263. https://doi.org/10.7717/peerj.1263.
Brusatte, S.L., M. Dumbravă, M. Vremir, Z. Csiki-Sava, R. Totoianu, and M.A. Norell. 2017. A catalog of Zalmoxes (Dinosauria: Ornithopoda) specimens from the Upper Cretaceous Nălaţ-Vad locality, Haţeg Basin, Romania. American Museum Novitates 3884: 1–36. https://doi.org/10.1206/3884.1.
Buffetaut, E., and J. Le Loeuff. 1991. Une nouvelle espèce de Rhabdodon (Dinosauria, Ornithischia) du Crétacé supérieur de l’Hérault (Sud de la France). Comptes Rendus De L’académie Des Sciences - Series IIA - Earth and Planetary Science 312: 943–948.
Carpenter, K., and P. Galton. 2018. A photo documentation of bipedal ornithischian dinosaurs from the Upper Jurassic Morrison Formation, USA. Geology of the Intermountain West 5: 167–207. https://doi.org/10.31711/giw.v5.pp167-207.
Carpenter, K., and M.C. Lamanna. 2015. The braincase assigned to the ornithopod dinosaur Uteodon McDonald, 2011, reassigned to Dryosaurus Marsh, 1894: Implications for iguanodontian morphology and taxonomy. Annals of Carnegie Museum 83: 149–165. https://doi.org/10.2992/007.083.0201.
Chanthasit, P. 2010. The ornithopod dinosaur Rhabdodon from the Late Cretaceous of France: anatomy, systematics and paleobiology. Unpublished PhD Thesis. Université Claude Bernard Lyon: 195 pp.
Chiarenza, A.A., M. Fabbri, L. Consorti, M. Muscioni, D.C. Evans, J.L. Cantalapiedra, and F. Fanti. 2021. An Italian dinosaur Lagerstätte reveals the tempo and mode of hadrosauriform body size evolution. Scientific Reports 11: 23295. https://doi.org/10.1038/s41598-021-02490-x.
Ciobanete, D., A. Panaiotu, C.G. Panaiotu, and C.E. Panaiotu. 2011. Geochemical and palaeomagnetic constrains on the evolution of Haţeg Basin (Romania) during Late Cretaceous. 73rd EAGE Conference and Exhibition incorporating SPE EUROPEC 2011: 3. https://doi.org/10.3997/2214-4609.20149693.
Codrea, V.A., T. Smith, P. Dica, A. Folie, G. Garcia, P. Godefroit, and J. Van Itterbeeck. 2002. Dinosaur egg nests, mammals and other vertebrates from a new Maastrichtian site of the Haţeg Basin (Romania). Comptes Rendus Palevol 1: 173–180. https://doi.org/10.1016/S1631-0683(02)00021-0.
Cope, E.D. 1869. Synopsis of the extinct Batrachia, Reptilia, and Aves of North America. Transactions of the American Philosophical Society 14: 1–252.
Coria, R.A., and J.O. Calvo. 2002. A new iguanodontian ornithopod from Neuquen Basin, Patagonia, Argentina. Journal of Vertebrate Paleontology 22: 503–509. https://doi.org/10.1671/0272-4634(2002)022[0503:ANIOFN]2.0.CO;2.
Cruzado-Caballero, P., X. Pereda-Suberbiola, and J.I. Ruiz-Omeñaca. 2010. Blasisaurus canudoi gen. et sp. nov., a new lambeosaurine dinosaur (Hadrosauridae) from the Latest Cretaceous of Arén (Huesca, Spain). Canadian Journal of Earth Sciences 47: 1507–1517. https://doi.org/10.1139/E10-081.
Cruzado-Caballero, P., J. Fortuny, S. Llacer, and J.I. Canudo. 2015. Paleoneuroanatomy of the European lambeosaurine dinosaur Arenysaurus ardevoli. PeerJ 3: e802. https://doi.org/10.7717/peerj.802.
Csiki, Z., and D. Grigorescu. 1998. Small theropods from the Late Cretaceous of the Haţeg Basin—an unexpected diversity at the top of the food chain. Oryctos 1: 87–104.
Csiki, Z., V.A. Codrea, C. Jipa-Murzea, and P. Godefroit. 2010a. A partial titanosaur (Sauropoda, Dinosauria) skeleton from the Maastrichtian of Nălaţ-Vad, Haţeg Basin, Romania. Neues Jahrbuch Für Geologie Und Paläontologie - Abhandlungen 258: 297–324. https://doi.org/10.1127/0077-7749/2010/0098.
Csiki, Z., D. Grigorescu, V.A. Codrea, and F. Therrien. 2010b. Taphonomic modes in the Maastrichtian continental deposits of the Haţeg Basin, Romania – palaeoecological and palaeobiological inferences. Palaeogeography, Palaeoclimatology, Palaeoecology 293: 375–390. https://doi.org/10.1016/j.palaeo.2009.10.013.
Csiki-Sava, Z., E. Buffetaut, A. Ősi, X. Pereda-Suberbiola, and S.L. Brusatte. 2015. Island life in the Cretaceous—faunal composition, biogeography, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago. ZooKeys 469: 1–161. https://doi.org/10.3897/zookeys.469.8439.
Csiki-Sava, Z., M. Vremir, Ş Vasile, S.L. Brusatte, G. Dyke, D. Naish, M.A. Norell, and R. Totoianu. 2016. The east side story—the Transylvanian latest Cretaceous continental vertebrate record and its implications for understanding Cretaceous-Paleogene boundary events. Cretaceous Research 57: 662–698. https://doi.org/10.1016/j.cretres.2015.09.003.
Dalla Vecchia, F.M. 2006. Telmatosaurus and the other hadrosauroids of the Cretaceous European Archipelago. An Overview. Natura Nacosta 32: 1–55.
Dalla Vecchia, F.M. 2009a. Telmatosaurus and the other hadrosauroids of the Cretaceous European Archipelago. An Update. Natura Nacosta 39: 1–18.
Dalla Vecchia, F.M. 2009b. Tethyshadros insularis, a new hadrosauroid dinosaur (Ornithischia) from the Upper Cretaceous of Italy. Journal of Vertebrate Paleontology 29: 1100–1116. https://doi.org/10.1671/039.029.0428.
Dalla Vecchia, F.M. 2014. An overview of the latest Cretaceous hadrosauroid record in Europe. In Hadrosaurs, ed. D.A. Eberth, D.C. Evans, and P.E. Ralrick, 268–297. Life of the Past. Bloomington: Indiana University Press.
Dieudonné, P.-E., T. Tortosa, F.F.B. Torcida, J.I. Canudo, and I. Díaz-Martínez. 2016. An unexpected early rhabdodontid from Europe (Lower Cretaceous of Salas de los Infantes, Burgos Province, Spain) and a re-examination of basal iguanodontian relationships. PLoS ONE 11: e0156251. https://doi.org/10.1371/journal.pone.0156251.
Evans, D.C. 2010. Cranial anatomy and systematics of Hypacrosaurus altispinus, and a comparative analysis of skull growth in lambeosaurine hadrosaurids (Dinosauria: Ornithischia). Zoological Journal of the Linnean Society 159: 398–434. https://doi.org/10.1111/j.1096-3642.2009.00611.x.
Galton, P.M. 1983. The cranial anatomy of Dryosaurus, a hypsilophodontid dinosaur from the Upper Jurassic of North America and East Africa, with a review of hypsilophodontids from the Upper Jurassic of North America. Geologica Et Palaeontologica 17: 207–243.
Galton, P.M. 1989. Crania and endocranial casts from ornithopod dinosaurs of the families Dryosauridae and Hypsilophodontidae (Reptilia: Ornithischia). Geologica Et Palaeontologica 23: 207–243.
Galton, P.M. 2001. The skull of Hypsilophodon, the small ornithopod dinosaur from the Lower Cretaceous of England. Dinopress 4: 118–127.
Galton, P.M. 2009. Notes on Neocomian (Lower Cretaceous) ornithopod dinosaurs from England - Hypsilophodon, Valdosaurus, “Camptosaurus”, “Iguanodon” - and referred specimens from Romania and elsewhere. Revue De Paléobiologie 28: 211–273.
Galton, P.M., and H.P. Powell. 1980. The ornithischian dinosaur Camptosaurus prestwichii from the Upper Jurassic of England. Palaeontology 23: 411–443.
Gates, T.A., and S.D. Sampson. 2007. A new species of Gryposaurus (Dinosauria: Hadrosauridae) from the late Campanian Kaiparowits Formation, southern Utah, USA. Zoological Journal of the Linnean Society 151: 351–376. https://doi.org/10.1111/j.1096-3642.2007.00349.x.
Gates, T.A., J.R. Horner, R.R. Hanna, and C.R. Nelson. 2011. New unadorned hadrosaurine hadrosaurid (Dinosauria, Ornithopoda) from the Campanian of North America. Journal of Vertebrate Paleontology 31: 798–811. https://doi.org/10.1080/02724634.2011.577854.
Gates, T.A., D.C. Evans, and J.J.W. Sertich. 2021. Description and rediagnosis of the crested hadrosaurid (Ornithopoda) dinosaur Parasaurolophus cyrtocristatus on the basis of new cranial remains. PeerJ 9: e10669. https://doi.org/10.7717/peerj.10669.
Gilmore, C.W. 1909. Osteology of the Jurassic reptile Camptosaurus, with a revision of the species of the genus, and descriptions of two new species. Proceedings of the United States National Museum 36: 197–332.
Gilmore, C.W. 1933. On the dinosaurian fauna of the Iren Dabasu Formation. Bulletin of the American Museum of Natural History 67: 23–78.
Godefroit, P., Y.L. Bolotsky, and J. Van Itterbeeck. 2004. The lambeosaurine dinosaur Amurosaurus riabinini, from the Maastrichtian of Far Eastern Russia. Acta Palaeontologica Polonica 49: 585–618.
Godefroit, P., H. Shulin, Y. Tingxiang, and P. Lauters. 2008. New hadrosaurid dinosaurs from the uppermost Cretaceous of northeastern China. Acta Palaeontologica Polonica 53: 47–74. https://doi.org/10.4202/app.2008.0103.
Godefroit, P., V.A. Codrea, and D.B. Weishampel. 2009. Osteology of Zalmoxes shqiperorum (Dinosauria, Ornithopoda), based on new specimens from the Upper Cretaceous of Nălaţ-Vad (Romania). Geodiversitas 31: 525–553. https://doi.org/10.5252/g2009n3a3.
Godefroit, P., F. Escuillié, Y.L. Bolotsky, and P. Lauters. 2012a. A new basal hadrosauroid dinosaur from the Upper Cretaceous of Kazakhstan. In Bernissart dinosaurs and early Cretaceous terrestrial ecosystems, ed. P. Godefroit, 335–358. Life of the Past. Bloomington: Indiana University Press.
Godefroit, P., Y.L. Bolotsky, and I.Y. Bolotsky. 2012b. Osteology and relationships of Olorotitan arharensis, a hollow-crested hadrosaurid dinosaur from the latest Cretaceous of far eastern Russia. Acta Palaeontologica Polonica 57: 527–560. https://doi.org/10.4202/app.2011.0051.
Godefroit, P., G. Garcia, B. Gomez, K. Stein, A. Cincotta, U. Lefèvre, and X. Valentin. 2017. Extreme tooth enlargement in a new Late Cretaceous rhabdodontid dinosaur from Southern France. Scientific Reports 7: 13098. https://doi.org/10.1038/s41598-017-13160-2.
Grigorescu, D. 1983. A stratigraphic, taphonomic and paleoecologic approach to a “forgotten land”: The dinosaur-bearing deposits from the Haţeg Basin (Transylvania-Romania). Acta Palaeontologica Polonica 28: 103–121.
Grigorescu, D. 2010. The latest Cretaceous fauna with dinosaurs and mammals from the Haţeg Basin—a historical overview. Palaeogeography, Palaeoclimatology, Palaeoecology 293: 271–282. https://doi.org/10.1016/j.palaeo.2010.01.030.
Hübner, T.R., and O.W.M. Rauhut. 2010. A juvenile skull of Dysalotosaurus lettowvorbecki (Ornithischia: Iguanodontia), and implications for cranial ontogeny, phylogeny, and taxonomy in ornithopod dinosaurs. Zoological Journal of the Linnean Society 160: 366–396. https://doi.org/10.1111/j.1096-3642.2010.00620.x.
Hulke, J.W. 1880. Iguanodon prestwichii, a new species from the Kimmeridge Clay founded on numerous fossil remains lately discovered at Cumnor, near Oxford. Quarterly Journal of the Geological Society of London 36: 433–456.
Liyong, J., C. Jun, Z. Shuqin, R.J. Butler, and P. Godefroit. 2010. Cranial anatomy of the small ornithischian dinosaur Changchunsaurus parvus from the Quantou Formation (Cretaceous: Aptian–Cenomanian) of Jilin Province, northeastern China. Journal of Vertebrate Paleontology 30: 196–214. https://doi.org/10.1080/02724630903412372.
Lull, R.S., and N. Wright. 1942. Hadrosaurian dinosaurs of North America. Geological Society of America Special Papers 40: 1–242.
Mallison, H., and O. Wings. 2014. Photogrammetry in paleontology—a practical guide. Journal of Paleontological Techniques 12: 1–31.
Marsh, O.C. 1881. Principal characters of American Jurassic dinosaurs. Part IV. Spinal cord, pelvis and limbs of Stegosaurus. American Journal of Science 21: 167–170.
Martin, J.E., M. Rabi, Z. Csiki-Sava, and Ş Vasile. 2014. Cranial morphology of Theriosuchus sympiestodon (Mesoeucrocodylia, Atoposauridae) and the widespread occurrence of Theriosuchus in the Late Cretaceous of Europe. Journal of Paleontology 88: 444–456. https://doi.org/10.1666/13-106.
Mayr, G., V.A. Codrea, A. Solomon, M. Bordeianu, and T. Smith. 2020. A well-preserved pelvis from the Maastrichtian of Romania suggests that the enigmatic Gargantuavis is neither an ornithurine bird nor an insular endemic. Cretaceous Research 106: 104271. https://doi.org/10.1016/j.cretres.2019.104271.
McDonald, A.T., J. Bird, J.I. Kirkland, and P. Dodson. 2012. Osteology of the basal hadrosauroid Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation of Utah. PLoS ONE 7: e45712. https://doi.org/10.1371/journal.pone.0045712.
McDonald, A.T., D.G. Wolfe, E.A. Freedman Fowler, and T.A. Gates. 2021. A new brachylophosaurin (Dinosauria: Hadrosauridae) from the Upper Cretaceous Menefee Formation of New Mexico. PeerJ 9: e11084. https://doi.org/10.7717/peerj.11084.
McFeeters, B., D. Evans, and H. Maddin. 2021. Ontogeny and variation in the skull roof and braincase of Maiasaura peeblesorum from the Two Medicine Formation of Montana, U.S.A. Acta Palaeontologica Polonica 66: 485–507. https://doi.org/10.4202/app.00698.2019.
Nopcsa, F. 1900. Dinosaurierreste aus Siebenbürgen I: Schädel von Limnosaurus transsylvanicus nov. gen et sp. Denkschriften Der Königlichen Akademie Der Wissenschaften 68: 555–591.
Nopcsa, F. 1902a. Dinosaurierreste aus Siebenbürgen II. Schädelreste von Mochlodon. Mit einem Anhange: zur Phylogenie der Ornithopodiden. Denkschriften der königlichen Akademie der Wissenschaften. Mathematisch-Naturwissenschaftliche Klasse 72: 149–175.
Nopcsa, F. 1902b. Über das Vorkommen der Dinosaurier bei Szentpéterfalva. Zeitschrift Der Deutschen Geologischen Gesellschaft 54: 34–39.
Nopcsa, F. 1904. Dinosaurierreste aus Siebenbürgen III. Weitere Schädelreste von Mochlodon. Denkschriften der königlichen Akademie der Wissenschaften. Mathematisch-Naturwissenschaftliche Klasse. Wien 74: 229–263.
Nopcsa, F. 1914. Über das Vorkommen der Dinosaurier in Siebenbürgen. Verhandlungen Der Zoologisch-Botanischen Gesellschaft Wien 54: 12–14.
Nopcsa, F. 1915. Die Dinosaurier der siebenbürgischen Landesteile Ungarns. Mitteilungen Aus Dem Jahrbuche Der Königlich Ungarischen Geologischen Reichsanstalt, Budapest 23: 3–24.
Nopcsa, F. 1923. Kallokibotion, a primitive amphichelydean tortoise from the Upper Cretaceous of Hungary. Palaeontologica Hungarica 1: 1–34.
Nopcsa, F. 1928. Dinosaurierreste aus Siebenbürgen IV. Die Wirbelsäule von Rhabdodon und Orthomerus. Palaeontologica Hungarica 1: 273–304.
Nopcsa, F. 1929. Dinosaurierreste aus Siebenbürgen V. Struthiosaurus transsylvanicus. Geologica Hungarica 4: 1–76.
Norman, D.B. 1980. On the ornithischian dinosaur Iguanodon bernissartensis of Bernissart (Belgium). Mémoires Institut Royal Des Sciences Naturelles De Belgique 178: 1–103.
Norman, D.B. 1986. On the anatomy of Iguanodon atherfieldensis (Ornithischia : Ornithopoda). Bulletin De L’institut Royal Des Sciences Naturelles De Belgique 56: 281–372.
Norman, D.B., A.W. Crompton, R.J. Butler, L.B. Porro, and A.J. Charig. 2011. The Lower Jurassic ornithischian dinosaur Heterodontosaurus tucki Crompton & Charig, 1962: Cranial anatomy, functional morphology, taxonomy, and relationships. Zoological Journal of the Linnean Society 163: 182–276. https://doi.org/10.1111/j.1096-3642.2011.00697.x.
Ősi, A., E. Prondvai, R.J. Butler, and D.B. Weishampel. 2012. Phylogeny, histology and inferred body size evolution in a new rhabdodontid dinosaur from the Late Cretaceous of Hungary. PLoS ONE 7: e44318. https://doi.org/10.1371/journal.pone.0044318.
Ostrom, J.H. 1961. Cranial morphology of the hadrosaurian dinosaurs of North America. Bulletin of the American Museum of Natural History 122: 33–186.
Owen, R. 1842. Report on British fossil reptiles. Part II. Reports of the British Association for the Advancement of Science 11: 60–204.
Panaiotu, C.G., and C.E. Panaiotu. 2010. Palaeomagnetism of the Upper Cretaceous Sânpetru Formation (Haţeg Basin, South Carpathians). Palaeogeography, Palaeoclimatology, Palaeoecology 293: 343–352. https://doi.org/10.1016/j.palaeo.2009.11.017.
Panaiotu, A.G., D. Ciobanete, C.G. Panaiotu, and C.E. Panaiotu. 2011. New palaeomagnetic data from the Haţeg Basin, Romania. In Abstract volume, Eighth Romanian Symposium on Palaeontology, 84–85. Bucharest, Romania: Ars Docendi.
Párraga, J., and A. Prieto-Márquez. 2019. Pareisactus evrostos, a new basal iguanodontian (Dinosauria: Ornithopoda) from the Upper Cretaceous of southwestern Europe. Zootaxa 4555: 247. https://doi.org/10.11646/zootaxa.4555.2.5.
Pereda-Suberbiola, X. 1999. Las faunas finicretácicas de dinosaurios ibéricos. Zubia 17: 259–279.
Pereda-Suberbiola, X., J.I. Canudo, P. Cruzado-Caballero, J.L. Barco, N. López-Martínez, O. Oms, and J.I. Ruiz-Omeñaca. 2009. The last hadrosaurid dinosaurs of Europe: A new lambeosaurine from the uppermost Cretaceous of Aren (Huesca, Spain). Comptes Rendus Palevol 8: 559–572. https://doi.org/10.1016/j.crpv.2009.05.002.
Porro, L.B., L.M. Witmer, and P.M. Barrett. 2015. Digital preparation and osteology of the skull of Lesothosaurus diagnosticus (Ornithischia: Dinosauria). PeerJ 3: e1494. https://doi.org/10.7717/peerj.1494.
Prieto-Márquez, A. 2005. New information on the cranium of Brachylophosaurus canadensis (Dinosauria, Hadrosauridae), with a revision of its phylogenetic position. Journal of Vertebrate Paleontology 25: 144–156. https://doi.org/10.1671/0272-4634(2005)025[0144:NIOTCO]2.0.CO;2.
Prieto-Márquez, A. 2010. The braincase and skull roof of Gryposaurus notabilis (Dinosauria, Hadrosauridae), with a taxonomic revision of the genus. Journal of Vertebrate Paleontology 30: 838–854.
Prieto-Márquez, A., and M. Carrera Farias. 2021. The late-surviving early diverging Ibero-Armorican ‘duck-billed’ dinosaur Fylax and the role of the Late Cretaceous European Archipelago in hadrosauroid biogeography. Acta Palaeontologica Polonica 66: 425–435. https://doi.org/10.4202/app.00821.2020.
Prieto-Márquez, A., and M.A. Norell. 2010. Anatomy and relationships of Gilmoreosaurus mongoliensis (Dinosauria: Hadrosauroidea) from the Late Cretaceous of Central Asia. American Museum Novitates 3694: 1–49. https://doi.org/10.1206/3694.2.
Prieto-Márquez, A., and G.C. Salinas. 2010. A re-evaluation of Secernosaurus koerneri and Kritosaurus australis (Dinosauria, Hadrosauridae) from the Late Cretaceous of Argentina. Journal of Vertebrate Paleontology 30: 813–837. https://doi.org/10.1080/02724631003763508.
Prieto-Márquez, A., F.M. Dalla Vecchia, R. Gaete, and À. Galobart. 2013. Diversity, relationships, and biogeography of the lambeosaurine dinosaurs from the European Archipelago, with description of the new aralosaurin Canardia garonnensis. PLoS ONE 8: e69835. https://doi.org/10.1371/journal.pone.0069835.
Prieto-Márquez, A., G.M. Erickson, and J.A. Ebersole. 2016. Anatomy and osteohistology of the basal hadrosaurid dinosaur Eotrachodon from the uppermost Santonian (Cretaceous) of southern Appalachia. PeerJ 4: e1872. https://doi.org/10.7717/peerj.1872.
Prieto-Márquez, A., V. Fondevilla, A.G. Sellés, J.R. Wagner, and À. Galobart. 2019. Adynomosaurus arcanus, a new lambeosaurine dinosaur from the Late Cretaceous Ibero-Armorican Island of the European archipelago. Cretaceous Research 96: 19–37. https://doi.org/10.1016/j.cretres.2018.12.002.
Săsăran, E., V.A. Codrea, and Z. Csiki. 2011. Fluvial systems e meandering rivers: a case study from Nălaţ-Vad area (Haţeg Basin, Romania). In Abstract book, 8th Romanian Symposium of Paleontology, Bucharest (Romania), 106–107. Bucharest, Romania: Ars Docendi.
Seeley, H.G. 1888. The classification of the Dinosauria. Report British Association for the Advancement of Science 1887: 698–699.
Shibata, M., P. Jintasakul, Y. Azuma, and H.-L. You. 2015. A new basal hadrosauroid dinosaur from the Lower Cretaceous Khok Kruat Formation in Nakhon Ratchasima Province, northeastern Thailand. PLoS ONE 10: e0145904. https://doi.org/10.1371/journal.pone.0145904.
Smith, T., V.A. Codrea, E. Săsăran, J. Van Itterbeeck, P. Bultynck, Z. Csiki, P. Dica, C. Fărcaș, A. Folie, G. Garcia, and P. Godefroit. 2002. A new exceptional vertebrate site from the Late Cretaceous of the Haţeg Basin (Romania). Studia Universitatis Babeș-Bolyai, Geologia, Special Issue 1: 321–330.
Sues, H.-D., and A. Averianov. 2009. A new basal hadrosauroid dinosaur from the Late Cretaceous of Uzbekistan and the early radiation of duck-billed dinosaurs. Proceedings of the Royal Society B: Biological Sciences 276: 2549–2555. https://doi.org/10.1098/rspb.2009.0229.
Sullivan, R.M., and T.E. Williamson. 1999. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus. New Mexico Museum of Natural History and Science Bulletin 15: 1–52.
Taquet, P. 1976. Géologie et paleontologie du gisement de Gadoufaoua (Aptien du Niger). Cahiers de Paléontologie. C.N.R.S.: 1–191.
Therrien, F. 2004. Paleoenvironments and magnetostratigraphy of the Maastrichtian Sînpetru Formation (Romania): Is the disappearance of dinosaurs indicative of the K/T Boundary? Journal of Vertebrate Paleontology 24: 7.
Therrien, F. 2006. Depositional environments and fluvial system changes in the dinosaur-bearing Sânpetru Formation (Late Cretaceous, Romania): Post-orogenic sedimentation in an active extensional basin. Sedimentary Geology 192: 183–205. https://doi.org/10.1016/j.sedgeo.2006.04.002.
Therrien, F., D.K. Zelenitsky, and D.B. Weishampel. 2009. Palaeoenvironmental reconstruction of the Late Cretaceous Sânpetru Formation (Haţeg Basin, Romania) using paleosols and implications for the “disappearance” of dinosaurs. Palaeogeography, Palaeoclimatology, Palaeoecology 272: 37–52. https://doi.org/10.1016/j.palaeo.2008.10.023.
Tsogtbaatar, K., D.B. Weishampel, D.C. Evans, and M. Watabe. 2014. A new hadrosauroid (Plesiohadros djadokhtaensis) from the Late Cretaceous Djadokhtan Fauna of southern Mongolia. In Hadrosaurs, ed. David A. Eberth, David C. Evans, and Patricia E. Ralrick, 108–135. Life of the Past. Bloomington: Indiana University Press.
Tsogtbaatar, K., D.B. Weishampel, D.C. Evans, and M. Watabe. 2019. A new hadrosauroid (Dinosauria: Ornithopoda) from the Late Cretaceous Baynshire Formation of the Gobi Desert (Mongolia). PLoS ONE 14: e0208480. https://doi.org/10.1371/journal.pone.0208480.
Van Itterbeeck, J., V.S. Markevich, and V.A. Codrea. 2005. Palynostratigraphy of the Maastrichtian dinosaur- and mammal sites of the Râul Mare and Bărbat Valleys (Haţeg Basin, Romania). Geologica Carpathica 56: 137–147.
Wang, X., Z. Csiki, A. Ősi, and G.J. Dyke. 2011. The first definitive record of a fossil bird from the Upper Cretaceous (Maastrichtian) of the Haţeg Basin, Romania. Journal of Vertebrate Paleontology 31: 227–230. https://doi.org/10.1080/02724634.2011.541013.
Weishampel, D.B., and P.R. Bjork. 1989. The first indisputable remains of Iguanodon (Ornithischia: Ornithopoda) from North America: Iguanodon lakotaensis, sp. nov. Journal of Vertebrate Paleontology 9: 56–66. https://doi.org/10.1080/02724634.1989.10011738.
Weishampel, D.B., D. Grigorescu, and D.B. Norman. 1991. The dinosaurs of Transylvania. National Geographic Research & Exploration 7: 196–215.
Weishampel, D.B., D.B. Norman, and D. Grigorescu. 1993. Telmatosaurus transsylvanicus from the late Cretaceous of Romania: The most basal hadrosaurid dinosaur. Palaeontology 36: 361–385.
Weishampel, D.B., C.-M. Jianu, Z. Csiki, and D.B. Norman. 2003. Osteology and phylogeny of Zalmoxes (n. g.), an unusual euornithopod dinosaur from the latest Cretaceous of Romania. Journal of Systematic Palaeontology 1: 65–123. https://doi.org/10.1017/S1477201903001032.
Wiman, C. 1929. Die Kreide-Dinosaurier aus Shantung. Palaeontologia Sinica. Serie C 6: 1–67.
Winkler, D.A., P.A. Murry, and L.L. Jacobs. 1997. A new species of Tenontosaurus (Dinosauria: Ornithopoda) from the Early Cretaceous of Texas. Journal of Vertebrate Paleontology 17: 330–348.
Xing, H., J.C. Mallon, and M.L. Currie. 2017. Supplementary cranial description of the types of Edmontosaurus regalis (Ornithischia: Hadrosauridae), with comments on the phylogenetics and biogeography of Hadrosaurinae. PLoS ONE 12: e0175253. https://doi.org/10.1371/journal.pone.0175253.
Acknowledgements
We would like to thank Panagiotis Kampouridis and Josephina Hartung (both University of Tübingen) for extensive discussions that surely improved the manuscript. Additionally, we are grateful to Andreas Matzke (University of Tübingen) for insightful discussions about braincase anatomy. We acknowledge the support of the Centre of Visualisation, Digitisation and Replication at the Eberhard Karls Universität in Tübingen (VDR) for instrument use, as well as scientific and technical assistance. More specifically, we want to thank Adrian Tröscher, Christina Kyriakouli, Verena Pietzsch and Gabriel Ferreira (all VDR) for producing high-resolution 3D prints of the basicrania that are deposited in the Palaeontological Collection of the University of Tübingen. DB was partially funded through the Swiss National Science Foundation (Grant no. 31003 A_179401 to T. Scheyer). Furthermore, we would like to thank the two reviewers Filippo M. Rotatori and Andrew T. McDonald for their constructive feedback that helped to improve the quality of the manuscript, as well as associate-editor Hans-Dieter Sues, editor-in-chief Mike Reich and editor-in-chief Michael Rasser for comments on the manuscript and their assistance during the publication process.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Prof. Hans-Dieter Sues.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Augustin, F.J., Dumbravă, M.D., Bastiaans, D. et al. Reappraisal of the braincase anatomy of the ornithopod dinosaurs Telmatosaurus and Zalmoxes from the Upper Cretaceous of the Haţeg Basin (Romania) and the taxonomic reassessment of some previously referred specimens. PalZ 97, 129–145 (2023). https://doi.org/10.1007/s12542-022-00621-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-022-00621-x