
Abstract
The Devonian–Carboniferous boundary beds in the Berchogur (Birshogyr) sections in the Mugodzhary (Mugalzhary) Mountains in western Kazakhstan, known to contain various fossil groups, including ammonoids, conodonts, foraminifers, corals, crinoids, and trilobites, are re-examined. The siliciclastic-carbonate succession of the Zhangana Formation reveals the presence of several ammonoid, conodont, and foraminiferal zones. The succession contains the Acutimitoceras ammonoid Genozone, equivalent to the level of the Stockum ammonoid fauna of Germany, with the conodont Siphonodella sulcata appearing within the Genozone. The same beds show mass occurrences of the foraminifer Tournayellina pseudobeata. The study of the Berchogur sections began in the 1980s; these sections are among very few successions globally with ammonoids of the Acutimitoceras Genozone in association with conodonts and foraminifers. At that time, several outcrops in a small area in the upper reaches of Burtybai (Zhangansai) Creek and two boreholes drilled near the sections were studied, and new taxa of ammonoids, foraminifers, ostracods, conodonts, algae, and spores were described. New excavations in 2018–2020, in conjunction with a search for a new definition of the Devonian–Carboniferous boundary, provided abundant new information on these taxa, on the lithology, and on crinoids, trilobites, and corals. The exact position of marker fossils and lithological changes are documented in several sections along Burtybai Creek allowing an amended correlation with sections of the D–C boundary beds in Western Europe.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Berchogur (Birshogyr) sections in the Mugodzhary (Mugalzhary) Mountains (western Kazakhstan) (Fig. 1) are among a few sections of the Devonian–Carboniferous (D–C) boundary beds with a variety of fossil remains including ammonoids, conodonts, foraminifers, corals, brachiopods, ostracods, trilobites, and other groups (Maslov 1987, etc.). Hence, these sections have been the focus of D–C boundary research for a number of years. Although many authors (Barskov et al. 1984, 1986, 1988; Kusina 1985; Maslov 1987; Vorozhbitov et al. 1992; Barskov and Kusina 1996) recognized their high importance for D–C boundary studies, no field research was undertaken on these sections between 1984 and 2018. This was due to the seemingly stable situation with this boundary following its ratification as a GSSP in the La Serre section in France at the supposed level of the first appearance datum (FAD) of the conodont Siphonodella sulcata (Huddle, 1934) (Paproth et al. 1991). Since S. sulcata was subsequently discovered in the La Serre GSSP section below the ratified level, the definition of the boundary and the GSSP section have been under re-consideration (see Kaiser 2009; Becker et al. 2016; Kaiser et al. 2019 for references). Several international teams are currently working on both these tasks. It is likely that the composite level corresponding to the base of the Protognathodus kockeli Zone, the beginning of radiation, and the end of major regression (top of Hangenberg Sandstone), will be the primary new boundary markers (Corradini et al. 2011, 2017; Becker et al. 2016; Kaiser et al. 2017; Spalletta et al. 2017). For the South Urals in Russia and western Kazakhstan, it means that the composite nature of the proposed boundary definition is the best option because it can be applied even where Pr. kockeli (Bischoff, 1957) is absent. This is important because protognathids are extremely rare in the Uralian and Northern Eurasian D–C sections. Our current research in western Kazakhstan aims to contribute to the study of D–C successions by re-examination of the sections in the Berchogur (Birshogyr) Depression (Mugodzhary Mountains), from which the Acutimitoceras ammonoid fauna and the conodont Siphonodella sulcata were previously reported (Barskov et al. 1984, 1988; Maslov 1987). These sections were studied by several teams of researchers in the 1950s and 1980s (see references below), but the precise position of the localities and the fossiliferous levels needed updating because of the disruption of previous topography by the extensive limestone quarrying in the eastern area of the Berchogur Depression (Alabas quarries), construction of new roads, changing of the course of local waterways, and destruction of several published outcrops. We have re-examined and resampled several remaining sections in the upper reaches of Burtybai Creek in the Berchogur Depression, recording the position of fossil occurrences in the sections as precisely as possible.

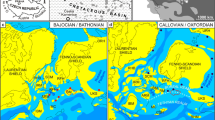
Maps of Kazakhstan and Shalkar District of the Aktobe Region and a satellite image (Google) of the vicinity of the village of Alabas. The map of the Shalkar District shows the main rivers and creeks mentioned on the text; it also shows the Burtybai section and “Outcrop 8” of Maslov (1987); the satellite image shows the position of trenches and boreholes: BK-1 (Trench BK-1, top): 48º34′02.6′′ N, 58º40′26.1′′ E, BK-3 (Trench BK-3, top): 48º34′08′′ N, 58º40′23′′ E, BK-4 (Trench BK-4, top): 48º34′03.8′′ N; 58º40′26.6′′ E; Loc. 3 (Locality 3 of 2018); Loc. 5 (Locality 5; Member 3c, top, 10 m north of Loc. 3), Loc. 6 (Locality 6; Member 3, top, 48º34′15.77′′ N, 58º40′26.01′′ E), Loc. 7 (Locality 7; Member 4, loose, 48º34′09.7′′ N, 58º40′22.3′′ E). Note that BK-1 is nearly the same as BK-1 of Barskov et al. (1984, 1988), Trench BK-3 was extended downwards in 2019 after Barskov et al. (1984, 1988), whereas “BK-2” and “BS-2” of Barskov et al. (1988) were not re-examined and are not mapped here. Locality 5 of 2019 is very close to Locality 3 of 2018. Locality 7 of 2019 is presumably very close to the ammonoid locality in Member 4 of Kusina (1985); Locality 4 is not shown, as it is near the top of BK-3 and would not have been visible at this scale. Boreholes BS-1 and BS-2 are after Barskov et al. (1988)
Geological background
The Berchogur sections of the D–C boundary beds are studied in western Kazakhstan (Aktobe Region, Shalkar District), in the Berchogur Depression (Syncline) between the Main Mugodzhary Mountains and Zhan-Gana Range (Ivanov et al. 1986). The D–C boundary beds crop out throughout the Berchogur Depression (Petrenko 1946; Balashova 1953), but the most complete sections are known in its western and eastern wings. The best studied sections are located at Burtybai Creek (= Zhangansai Creek), right tributary of the Shyuldak (= Chuuldak or Shuldak) River (Fig. 1). These sections are on the left bank of Burtybai Creek in its meridional course, north of the road connecting the old and new Alabas quarries, 5 km east of the village of Alabas and 15 km northeast of Berchogur (Birshogyr) railway station. Here, fossil occurrences are found in the Zhangana Formation, studied in seven localities (including four trenches excavated in 1981, 1983, and 2019) and two boreholes (drilled in 1983) (Barskov et al. 1984, 1988; Nikolaeva and Mustapaeva 2018) (Fig. 1).
The Berchogur Depression is one of the prominent structures of the South Urals, a meridional syncline (ca. 30 × 60 km) with Middle and Upper Devonian and lower Carboniferous sediments in the middle surrounded by thick Lower–Middle Devonian volcanic series of the Mugodzhary, Kurkuduk, and Milgashin formations (Petrenko 1946; Ivanov et al. 1986; Maslov 1987). The lower Carboniferous beds include the Zhangana, Berchogur, and Karabulak formations, represented by 900 m-thick siliciclastics and carbonate-siliciclastics with coal and coalified shale beds in the upper horizons. Tectonically, the Berchogur Depression is part of the Western Mugodzhary (Puchkov 2000, 2010), an area frequently interpreted as a continuation of the Magnitogorsk Megasynclinorium, whereas some reconstructions (Zonenshain et al. 1990; Ivanov et al. 2014) interpret this area as occurring near the western border of the Mugodzhary Microcontinent separating the latter from the Palaeoasian Ocean. However, the discussion of the tectonics of this area is beyond the scope of this paper.
There are two main regions with fossiliferous D–C boundary interval sections exposed in the Aktobe Region (Mugodzhary Mountains), in the east and west of the Berchogur Depression. Balashova (1945a, b, 1953, 1956) described the stratigraphy and fossils of these sections, which she began to study in 1939 (see Petrenko 1946, p. 2) and described ammonoids from the western regions of the depression: (1) Aktan and Kust-Kara dry creeks; (2) middle reaches of Sarysai Creek; (3) west of the Zhan-Gana Range, Tungulyk-Bulak Gully; and (4) Kabaksai Creek (Fig. 1). Balashova (1945a, p. 189) and Balashova in Petrenko (1946) considered the ammonoid assemblage containing mainly species of a genus that she identified as “Imitoceras” (most of these species are currently assigned to Acutimitoceras, while some need further revision and are referred to as “Imitoceras”, see the discussion below) to indicate a lower Gattendorfia ammonoid fauna. She drew the D–C boundary 10 m below the limestones with ammonoids, at the base of limestones containing the brachiopod Paulonia ranovensis (Peetz, 1893), at the level coinciding with the top of the limestones with the brachiopod Retzia ulentica Nalivkin, 1937. Rozman (1960, 1962, p. 52) studied in detail the D–C boundary beds in the Berchogur Depression, assembled a collection of ammonoids, brachiopods, and foraminifers, and placed the D–C boundary considerably lower, at the base of the limestones with the brachiopod Ptychomaletoechia panderi (Semenov and Moeller, 1864), at the level corresponding to the base of the Zhangana Formation. In 1981, a research group led by B.I. Bogoslovsky re-examined one of Rozman’s sections in the upper reaches of Burtybai Creek (Locality 3, Fig. 1, Trench BK-3 in Barskov et al. 1984, 1988) and collected ca. 200 specimens of ammonoids later described by Kusina (Barskov et al. 1984; Kusina 1985; Bogoslovsky 1988). In the 1980s, trench BK-3 was essentially a pit that only exposed Member 3. In 1979 and 1981, a team led by N.M. Kochetkova, studied the sections along the middle reaches of Burtybai Creek (Kulagina 1982; Kochetkova and Kulagina in Maslov, 1987). These were the Burtybai section and “Outcrop 8”. The Burtybai section extended from 2.5 km west of the mouth of Burtybai Creek to the valley in its upper reaches (currently crossed by a new road and not easily identifiable). Deposits of the Burtybai section were subdivided into 11 lithological units. “Outcrop 8” was located ca. 3 km south-west of the mouth of Burtybai Creek (confluence with the Shyuldak River. The thickness of “Outcrop 8” is 5.3 m and it includes six beds (Kochetkova and Kulagina in Maslov, 1987). Bed 1 (2.5 m-thick) of “Outcrop 8” correlates with the uppermost part of “Unit VIII” of the Burtybai section (Beds with the ostracods Armenites asiaticus–Selebratina subtarchanica –Bairdia turgida) and Beds 2–6 of “Outcrop 8” were described as “Unit IX” in the Burtybai section (upper part of the Zhangana Formation with ostracods of the Pseudoleperditia tuberculifera–Coryellina alba– Cribroconcha primaris assemblage, the foraminifer Tournayellina, and the calcareous alga Garwoodia) (Maslov 1987). In 1983, a team led by I.S. Barskov and colleagues from the Aktobe Geological Survey focussed on the section in the upper reaches of Burtybai Creek, excavated two trenches (BK-1 and BK-2), and drilled two boreholes (BS-1 and BS-2) (Barskov et al. 1984, 1988) (Fig. 1). They referred to these sections as the “Berchogur sections” and recognized a different set of members in the Zhangana Formation (From 1 to 4), which we follow here. It was established that the limestones with ammonoids (Member 3) are underlain (Member 2) and overlain (Member 4) by shales with lenses and interbeds of argillaceous limestones. A second ammonoid-bearing horizon (here “Locality 7”) was discovered on the slope ca. 18.5 m above Trench BK-3, and in Borehole BS-2. Foraminifers (E.A. Reitlinger and E.I. Kulagina) and ostracods (N.M. Kochetkova) were re-examined based on the joint collections of all the teams that had worked in the region and were published as collective research (Simakov et al. 1985; Maslov 1987), where several foraminiferal and ostracod-based units were recognized. Barskov et al. (1984, p. 213) proposed that the D–C boundary in Berchogur can be drawn “within the lineage of the earliest Acutimitoceras. This boundary may be translated at least in term of conodonts (base of the Siphonodella sulcata Zone)…”. This level is within Member 3, at the same horizon as ammonoid Sample 3/5 of Barskov et al. (1984), and also at the level of Member 3c (this paper, Figs. 2, 3). Barskov et al. (1984, 1988) referred to this earlier finding, but also considered that samples BS-2/51 from the depth of 29.0 m contained “specimens of Pseudopolygnathus similar to P. fusiformis”. Barskov et al. (1984, 1988) went on to suggest that this level can therefore correspond to the interval of the Siphonodella sulcata Zone, and already be Carboniferous, as it would agree with the decision of the International Working Group referred to by Paproth and Streel (1985). Therefore, Barskov et al. (1988, p. 178) suggested that the conodont-based boundary as the “boundary between the B. costatus and S. sulcata zones in the Berchogur section can be expected within Member 2b in the argillaceous series 70-cm thick above the base of the “sandstone-like” limestone of Member 2a”. However, subsequent searches have not confirmed these expectations, and the base of the S. sulcata Zone is currently drawn at the base of Member 3 (which is slightly below the FOD of S. sulcata in Member 3c (Sample BK-3/5a) in Trench BK-3 and in Member 3b (Sample 19/12-3) in Trench BK-1, as it is likely that the earliest entry of S. sulcata in Member 3b is not its evolutionary first appearance) (Fig. 3).
In 2018–2019, we re-excavated Trench BK-1 of Barskov et al. (1984, 1988) and excavated two more new trenches in the upper reaches of Burtybai Creek. One of these new trenches is an expansion of Trench BK-3 of Barskov et al. (1984, 1988; Figs. 2–3). The presented research re-examined a series of parallel sections of the upper part of the Zhangana Formation (members 2 and 3 in Barskov et al. 1984, 1988) in the meridional portion of Burtybai Creek that were referred to as the “Berchogur section” by Barskov et al. (1984, 1988) and Simakov et al. (1985). This interval of the succession overlies the Zhangana limestones (algal-foraminiferal wackestones of Member 1 of Barskov et al. (1984, 1988) and Unit VIII in the Burtybai section of Maslov (1987)) and contains two major parts. The lower part is mainly shale with interbeds of limestone and is referred to as Members 2a and 2b exposed in trenches BK-1, BK-3 (Member 2a is not exposed in BK-3) (see Figs. 2, 3), BK-4 (Fig. 3), and boreholes BS-1, BS-2 (Fig. 4). The upper part is Member 3 represented by peloid wackestones forming a good marker horizon, 0.45 cm thick that yielded ammonoids of the Acutimitoceras Genozone, the conodont Siphonodella sulcata, and the foraminifer Tournayellina pseudobeata. Limestones of Member 3 are overlain by shales of Member 4, which are studied in isolated outcrops. The total thickness of the succession is ca. 35–40 m.

Berchogur (Birshogyr) sections studied in Trenches BK-1, BK-3, and BK-4 (not discussed in this paper), and position of Borehole BS-1 on the left bank of Burtybai Creek in the northeast of the Berchogur Depression (Syncline)

Stratigraphic logs and distribution of conodonts, ammonoids and selected taxa of foraminifers in Trenches BK-1 and BK-3. The upper part of BK-3 (Member 4, shaded) is studied in isolated outcrops. (1) limestone; (2) marly limestone; (3) shales; (4) alternation of shale and thin beds of limestone with bioclast aggregates; (5) carbonate concretions; (6) beds of gravelstone; (7) plant debris; (8) algae and cyanobacteria; (9) foraminifers; (10) ammonoids; (11) ostracods; (12) bryozoans; (13) bivalves; (14) conodonts; (15) crinoids; (16) brachiopods

Stratigraphic logs and distribution of conodonts and selected taxa of foraminifers in Boreholes BS-1 and BS-2. Explanation as in Fig. 3 and additionally (1) covered interval; (2) shale with bioclasts
The interval of members 2b and 3 corresponds to Unit IX of the Burtybai section and “Outcrop 8” (Maslov 1987, pp. 10–36).
Lithology and depositional settings
The depositional settings were interpreted based on the study of thin sections and microfossils (other than conodonts and foraminifers) and by manual isolation from the residue of several samples from Members 2a and 2b boiled in fresh water with no acid added, and then washed and settled and separated using a light microscope. Importantly, all samples throughout the section lacked any visible siliciclastics or silt- or sand-size particles. Previously published reports of “arenaceous shale” (e.g., Maslov 1987) were based on field observations of some “grains” in shales and limestones, which represent aggregations of calcareous bioclasts, mainly algae. Below, the results of new observations are included in the description of the recognized lithological units, in stratigraphic order:
Member 1. (Figs. 3, 4) (> 10 m thick). Light-grey, pinkish-grey, and greenish-grey, medium- to thick-bedded limestone with foraminifers, stromatoporoids, rugose corals, ostracods, shallow-water conodonts of the Polygnathus-Icriodus biofacies (Barskov et al. 1988). Microfacies: algal-foraminiferal wackestone.
Member 2a. (1.3–1.5 m). Dark-green, laminated shale, followed by gravelstone, brown and brownish-grey, algal grainstones with interbeds of arenaceous shale (Unit 2a1). The fissile shales (Bed 3) contain a very diverse benthic assemblage: brachiopods, bryozoans, ostracods, gastropods, crinoids, ophiuroids, echinoids, holothurians, machaeridians, as well as the microproblematic Menselina Antropov, 1967, characteristic of the Q. kobeitusana Zone in Europe and Asia (Mamet 1991; Ivanova and Stepanova 2011). Shales composing this unit are studied in samples BK-1-19/3-1 and BK-1-19/3-2 (Trench BK-1, year 2019) (Fig. 3). They contain various bioclasts, including crinoidal columnals, ostracod tests, tubular calcareous algae, and the problematic (alga?) of the genus Menselina. The latter were previously cited from the Burtybai section, Member VII, Samples 43 and 48 (Maslov 1987). Branching bryozoans are uncommon; the residue contains small fragments of brachiopod shells, holothurian sclerites, remains of ophiuroids, echinoids and asteroids, moulds of gastropod shells, machaeridian sclerites, and conodonts. The diversity of the benthos and presence of calcareous algae suggest a normal marine shallow environment of the inner shelf. The nearly total absence of fish remains is very striking (only one or two small conical teeth have been found). The scarcity of fishes is characteristic of all lithological members of the Berchogur sections.
A bed of gravelstone (Bed 5 in Fig. 3), 1–5 mm thick, is present in the upper part of the unit and is immediately followed by algal grainstone (limestone) of Unit 2a2 (0.5–1 m thick, Bed 6) terminating the member. The upper bedding surface of this limestone is slightly irregular, brownish, and represents a hardground suggesting a possible gap in sedimentation. Bedding surfaces of limestones and shale of Bed 6 contain accumulations of small coalified plant remains. Limestones contain algae, foraminifers, ostracods, and spores (Barskov et al. 1988).
Unit 2a2 corresponds to the so-called “sandstone-like limestone” (Maslov 1987). The bioclast composition was studied in samples BK-1/11 (Trench 1) (Fig. 3) and BS-1/25 (Borehole BS-1) (Fig. 4). This grainstone is mainly composed of remains of calcareous dasycladaceans and Menselina. The residue also contains occasional crinoidal columnals, fragmented and juvenile brachiopod shells, remains of echinoids, ophiuroids, and plates of machaeridians. The assemblage is less diverse than in the underlying Unit 2a1, but still marine, although indicating a shallower depth. It is likely that the basin somewhere had algal meadows producing a very large amount of carbonate bioclasts. At the same time the residue contains some altered mica plates suggesting an influx of some clastic components from the land. Interestingly, the assemblage contained one spirally coiled microconchid tube, which might indicate a slight decrease in salinity (Gierlowski-Kordesch and Cassle 2015).
Member 2b. (2–2.5 m) Dark-brown, greenish-grey to black shale, yellowish brown where weathered. In the middle of the member there is a very thin (1–3 cm) interbed of marly limestone with abundant bivalve imprints (Bed 8, Sample BK-14) (Fig. 3). Within Bed 10 there is a layer with numerous carbonate concretions and abundant fossils, including tabulate and rugose corals, brachiopods, bryozoans, and crinoidal columnals.
Two samples were studied from Trench BK-2 from the year 1983 (BK-2/5 and BK-2/6). Shales of this unit contain smaller and larger (up to 3.5 × 3 × 1.5 cm) carbonate concretions. Some concretions contain moulds and, in some cases, fragments (up to 1 cm) of turriform gastropod shells. The shale contains many fragments of brachiopod shells and ferruginous moulds of juvenile brachiopods. There are hundreds or thousands of small plates of the cirripedian Pabulum Whyte (1976) described from the upper Viséan of Yorkshire and Scotland, crinoidal columnals, ostracod tests, branching bryozoans, and asteroid plates, the latter with somewhat different morphology to the plates from Unit 2a1. Ostracod tests and bivalve moulds, and echinoid, ophiuroid, holothurian, and machaeridian remains are rare. The residues contained fragments of orthoconic non-ammonoid cephalopod shells, and one small conical fish tooth. No identifiable remains of calcareous algae or Menselina were found. This composition of bioclasts clearly indicates a fast deepening of the basin during accumulation of the sediments of Member 2b and increased salinity, while the general taxonomic composition of the biota remained largely unchanged compared to Member 2a. Again, almost no fish remains were found. This suggests that the eustatic sea-level fall event between Members 2a and 2b was not long and did not result in a fundamental change in the environment. It is likely to be related to a significant depth increase, with the depth of the seafloor below the photic zone.
In summary, Member 2a reflects a single short-term stage in the evolution of the basin, showing a significant shallowing trend. Deposits of Member 2b were accumulated after a short gap in deposition in a considerably deeper basin, although retaining the marine biota of previous times.
Beds 9–11 are represented by greenish-blue shale and are one unit. Before our excavations “Bed 9” and “Bed 11” were covered intervals separated by an exposed interval of “Bed 10” (Barskov et al. 1988, text-fig. 2). Therefore, we use bed nos. 9 and 11 for continuity. It would be more logical to have a boundary between beds 10 and 11 lower, at the level within Bed 10 with numerous carbonate concretions and abundant fossils. However, at this stage we focus on maintaining correspondence with previous studies, especially because in 2019 we used the same samples nos. as in Barskov et al. (1988). A detailed conodont study including processing samples of 5 kg throughout the section is in progress and will be published in a separate paper.
Member 3. (0.45–0.5 m) Dark grey to yellowish-grey marly limestone with rare accumulations of ammonoids in the upper part. Apart from ammonoids, the limestone contains brachiopods, trilobites, ostracods, foraminifers, conodonts, and spores (Barskov et al. 1988). In Trench BK-3, the member is subdivided into three distinct units: 3a, 3b, and 3c, with ammonoids only in 3c (see Figs. 3, 5). S. sulcata also comes from the upper part of Member (3b and 3c). This level corresponds to Sample 3/5 of Barskov et al. (1984).

Photograph of the upper part of Trench BK-3 in 2019 showing units 3a, 3b, 3c, localities 3 and 5. Seen from the south
3a (0.2 m). Micropeloid wackestone, in places marly, with rare foraminifers, crinoids, and the algae Garwoodia gregaria (Nicholson, 1888). Occasionally, the rock contains accumulations of small skeletal remains (gastropods, thin-shelled ostracods) (Fig. 6D, H, I).

Microstructures, foraminifers, and algae from Trench BK3, Member 3 (a, b, d, f–i), and Member 4 (c, e), collected in 2019. Collection no. 123 is housed in the Institute of Geology, Russian Academy of Sciences, Ufa (IG UFRC RAS); a Earlandia minima Birina, 1948 in peloidal wackestone, longitudinal section, Sample 19BK3b (1); b Tournayellina pseudobeata Reitlinger and Kulagina in Maslov, 1987, median sections, Sample 19BK3a (1); c, e Bioclastic peloidal wackestone, Locality 7, Sample BK3-4a (1); c with Tournayellina pseudobeata (T), thin-walled ostracods (O), and a single echinoid spine (E); e with Garwoodia gregaria (Nicholson, 1888), d Mudstone with crinoids and juvenile shell of ammonoid, Sample 19BK3c (1); f Peloidal wackestone with bioclasts of bryozoans (B), crinoids (C), and ostracods (O), Sample 19BK3b (2); g Peloidal wackestone with rare foraminifers, the same thin section as in A with lower magnification; h Garwoodia gregaria Nicholson, 1888, Sample 19BK3a (2); i Fine-grained bioclastic peloidal wackestone, same thin section as in B with lower magnification; scale bar for a–b 0.2 mm; scale bars for c–i are shown in mm in each image
3b (0.12 m). Wackestone/mudstone with rare unilocular foraminifers, Earlandia (Fig. 3A), bioclasts of bryozoans, crinoids, and ostracods, with peloidal microsparite matrix (Fig. 6F, G).
There is a 10-cm thick bed of shale between 3b and 3c.
3c (0.1 m). Mudstone with rare juvenile ammonoid shells and microbioclasts (Fig. 6D).
In summary, sediments of Member 3 were accumulated below the fair-weather wave base in an oxic environment, suggested by the presence of benthic organisms and traces of bioturbation.
Member 4. (18.5 m) Dark grey to black calcareous shale, sometimes with thin (1 cm) beds of argillaceous limestone or marl and microbioclastic wackestone. Thin sections show numerous thin-walled ostracods, foraminifers, algae, and echinoids fragments (Fig. 6C, E). Fossils are found at the base and in the top part and include ammonoids, nautiloids, brachiopods, corals, conularians, and crinoids. Conodonts are mainly found at the base of the member (Barskov et al. 1988). In borehole BS-2, Member 4 is overlain by sandstones with shale interbeds of the Berchogur Formation, probably of continental origin. This member is studied in several poorly exposed intervals above 3c.
Fossils
Conodonts
In 2019 we resampled the Berchogur section (carbonates and shale members) for conodonts (trenches BK-1, BK-3, BK-4). Trench BK-2 (of Barskov et al. 1984, 1988) has not been resampled. The conodont processing is ongoing, but we can present here our preliminary results. The studied samples contained 36 conodont specimens of which 18 are platform elements and the rest are ramiform ones. Seven conodont genera are identified: Polygnathus, Neopolygnathus, Bispathodus, Siphonodella, Mehlina, Icriodus, and Apatognathus. The most important find was a discovery in Unit 3b in Trench BK-1 (Sample BK1-19-12/3) of Siphonodella (Eosiphonodella) sulcata, which allowed this member to be assigned to the sulcata Zone, as it was established previously in Trench BK-3 (Barskov et al. 1988) (Fig. 7).

Famennian-Tournaisian conodonts from the Berchogur (Birshogyr) sections; a, b Siphonodella (Eosiphonodella) sulcata (Huddle, 1934); a BK-3/5a; also in Barskov et al. 1984, pl. 6, fig. 7a; also in Maslov, 1987, pl. 27, fig. 12a Member 3, Bed 4, Sample 5a; lower Tournaisian, sulcata Zone; b BK-3/5a; Member 3, Bed 4, Sample 5a; lower Tournaisian, sulcata Zone; c, e Icriodus costatus (Thomas, 1949): c BS-2/54; also in Barskov et al. 1984, pl. 7, fig. 2a; Maslov, 1987, pl. 25, fig. 9; Member 2a2, Sample 54; uppermost Famennian, Lower–Middle praesulcata zones (top praesulcata Zone sensu Kaiser et al. 2009); e BS-2/54; Sample 54; uppermost Famennian, Lower–Middle praesulcata zones (top praesulcata Zone sensu Kaiser et al. 2009); d Neopolygnathus communis (Branson and Mehl, 1934b); BK-2/10; also in Maslov 1987, pl. 26, fig. 1a; Member 3, Sample 10; lower Tournaisian, sulcata Zone; f, g Polygnathus inornatus inornatus Branson, 1934; f BS-2/54; also in Maslov 1987, pl. 27, fig. 1a; Member 2a2, Sample 54; uppermost Famennian, Lower–Middle praesulcata zones (top praesulcata Zone sensu Kaiser et al. 2009); g BS-1/24; Member 2a2, Sample 24; uppermost Famennian, Lower–Middle praesulcata zones (top praesulcata Zone sensu Kaiser et al. 2009); h–k Siphonodella cf. praesulcata Sandberg in Sandberg et al., 1972; h BS-1/7; also in Maslov, 1987, pl. 27, fig. 6a; Member 2b, Sample 7; uppermost Famennian, Middle praesulcata Zone (costatus-kockeli Interregnum); i BS-1/12; also in Maslov, 1987, pl. 27, fig. 7a; Member 2b, Sample 12; uppermost Famennian, Upper praesulcata Zone (costatus-kockeli Interregnum); j BS-1/10; also in Barskov et al. 1984, pl. 7, fig. 2b; Member 2b, Sample 10; uppermost Famennian, Upper praesulcata Zone (costatus-kockeli Interregnum); k BK-1/15; Member 2b, Bed 10, Sample 15; uppermost Famennian, Middle praesulcata Zone (costatus-kockeli Interregnum); l–n Neopolygnathus mugodzaricus (Gagiev, Kononova and Pazukhin in Maslov, 1987): l BS-2/54; Member 2a2, Sample 54; uppermost Famennian, Lower–Middle praesulcata zones (top praesulcata Zone sensu Kaiser et al. 2009); m BS-2/54; Member 2a2, Sample 54; uppermost Famennian, Lower–Middle praesulcata zones (top praesulcata Zone sensu Kaiser et al. 2009); n BS-2/54; also in Barskov et al. 1984, pl. 7, fig. 6b; Maslov 1987, pl. 26, fig. 5b; Member 2a2, Sample 54; uppermost Famennian, Lower–Middle praesulcata zones (top praesulcata Zone sensu Kaiser et al. 2009); o, p Bispathodus spinulicostatus (Branson, 1934): o BK-1/22; Member 3, Bed 12, Sample 22; uppermost Famennian, possible equivalent of the Upper praesulcata Zone (= kockeli Zone); p BK-3/2; Member 3, Bed 4, Sample 2; uppermost Famennian, possible equivalent of the Upper praesulcata Zone (= kockeli Zone); q–s Bispathodus aculeatus aculeatus (Branson and Mehl, 1934a): q BK-1/22; also in Maslov 1987, pl. 28, fig. 4a; Member 3, Bed 12, Sample 22; uppermost Famennian, possible equivalent of the Upper praesulcata Zone (= kockeli Zone); r BK-1/22; Member 3, Bed 12, Sample 22; uppermost Famennian, possible equivalent of the Upper praesulcata Zone (= kockeli Zone); s BK-3/1; Member 3, Bed 4, Sample 1; uppermost Famennian, possible equivalent of the Upper praesulcata Zone (= kockeli Zone); t Bispathodus stabilis vulgaris (Dzik, 2006), BK-3/3; also in Maslov 1987, pl. 28, fig. 3; Member 3, Bed 4, Sample 3; uppermost Famennian, possible equivalent of the Upper praesulcata Zone (= kockeli Zone); u Mehlina strigosa (Branson and Mehl, 1934a, b), BS-1/37; Member 1, Sample 37; uppermost Famennian, Lower–Middle praesulcata zones (top praesulcata Zone sensu Kaiser et al. 2009); scale bar for all figs. 100 μm
In this paper we also add the distribution of conodonts based on the sampling of 1981–1983 from three trenches and two boreholes based on ca. 1 kg samples. The conodonts are extremely rare, discovered only in a few samples. Some conodont elements from these samples were already illustrated in Maslov (1987). Conodont collection no. 243 is housed at the Department of Paleontology, Lomonosov Moscow State University (MSU). The analysis of the conodont distribution in the Berchogur sections allows the recognition of the following conodont zones (Fig. 8):
-
1.
the interval of the Upper expansa to basal part of the Middle praesulcata zones (= Bispathodus ultimus ultimus and Siphonodella (Eosiphonodella) praesulcata s.l. zones sensu Kaiser et al. 2009; Becker et al. 2016; Söte et al. 2017) in Members 1, 2a1 and 2a2.
-
2.
the main part of the former Middle praesulcata Zone (= costatus-kockeli Interregnum sensu Kaiser et al. 2016; Becker et al. 2016) in Member 2b
-
3.
possible equivalents of the Upper praesulcata Zone (= Protognathodus kockeli Zone) in Member 2b.
-
4.
sulcata Zone in Member 3 (FO in 3b-c) and in Member 4 (Figs. 3, 4).

Conodont zonal correlation across the Devonian–Carboniferous boundary, showing the span of the trenches and boreholes. Bi., Bispathodus; Eo., Eosiphonodella, Pr.; Protognathodus; S., Siphonodella
These conodont-based units in the Berchogur sections are considered below:
-
1.
The undivided interval of the Upper expansa to basal Middle praesulcata zones (= Bispathodus ultimus ultimus and Siphonodella (Eosiphonodella) praesulcata s.l. zones) (Figs. 3, 4) is recognized based on the presence of Icriodus costatus (Thomas, 1949) (Fig. 7c, e), Poly-gnathus inornatus (Branson, 1934) (Fig. 7f, g), Mehlina strigosa (Branson and Mehl, 1934a) (Fig. 7u), and Apatognathus varians cipitis Kononova in Bushmina and Kononova, 1981. Of these, Apatognathus varians cipitis was originally described from the upper/uppermost Famennian of the Kuznetsk Basin (Western Siberia, Russia), from beds with Icriodus costatus and Polygnathus parapetus (Druce, 1969), originally correlated with the upper part of the B. costatus Zone (Bushmina and Kononova 1981, pp. 25, 34, Table 6). This subspecies is also found in the South Urals and occurs in the local Pseudopolygnathus trigonicus Zone, which corresponds to the Upper expansa to basal Middle praesulcata zones (Artyushkova et al. 2011, text-fig. 20). Also, Pseudopolygnathus trigonicus Ziegler, 1962 became extinct at the base of the Hangenberg Black Shale interval (e.g., Kaiser et al. 2009, text-fig. 3). Specimens identified as Icriodus costatus are most similar to I. costatus darbyensis Klapper, 1958 Morphotype 2, which occurs in the expansa Zone and continues to the Lower and basal Middle praesulcata zones (Sandberg and Dreesen 1984; top of the praesulcata Zone sensu Kaiser et al. 2009). The species Polygnathus inornatus is known from many regions of the world; it first appears in the upper Famennian Middle expansa Zone (base of the Bispathodus costatus Zone, see the range chart of Ziegler and Sandberg 1984) and continues to the upper Tournaisian (e.g., Spalletta et al. 2017).
We have no local record of the praesulcata Zone in Members 1 and 2a that fall in the interval of the B. ultimus and S. (Eo.) praesulcata s.l. zones. Considering the conodont data, we provisionally correlate Members 1 and 2a with the Wocklum Limestone of the Rhenish Massif (cf. recent review by Becker et al. 2016 and Kaiser et al. 2016) (Fig. 8). The endemic species Neognathodus mugodzaricus (Gagiev, Kononova and Pazukhin in Maslov, 1987) (samples BK1-19-6/3; BK1-19-1/1, etc.) (Fig. 7L–N) was described from the upper Famennian of the Berchogur Depression (Maslov 1987, p. 95, pl. 26, figs. 4–8) and has not been recorded from any other region. This species belongs to the N. communis group, which is classified in the genus Neopolygnathus by Vorontzova in Barskov et al. (1991). The presence in Sample BK-19-1/1 (top of Member 1) of Mehlina strigosa and Apatognathus varians cipitis Kononova in Bushmina and Kononova, 1981 supports the assignment of this part of the section to the uppermost Famennian (Bushmina and Kononova 1981; Ziegler and Sandberg 1984).
-
2.
The shallowing upwards trend at the top of Member 2a2 (Beds 4/5) may indicate the regressive Hangenberg Crisis Prelude, such as one at the top of the Wocklum Limestone. The range of icriodids to the top of Member 2a1 suggests that this is the main Hangenberg extinction level and, therefore, the top of the praesulcata Zone s.l. or the level equivalent to the base of the costatus-kockeli Interregnum. The onlap of dark shales (lower Member 2b) above a small disconformity at the top of Member 2a2 can correspond to a transgressive phase, and the lower part of Member 2b correlates with the Lower Crisis Interval (lower costatus-kockeli Interregnum). In the Berchogur sections, the earliest occurrence of Siphonodella (Eosiphonodella) praesulcata is recorded from Member 2b (Borehole BS-1, Fig. 4), whereas in other regions its appearance defines the Lower praesulcata Zone (s.l., see discussion in Söte et al. 2017). The assemblage of Member 2b also contains Pseudopolygnathus cf. fusiformis (Branson and Mehl, 1934b) (see Barskov et al. 1984), Polygnathus inornatus, and Neopolygnathus mugodzaricus (BK-1, Sample 17). Our specimens of S. (Eo). praesulcata are not very similar to specimens of S. (Eo.) praesulcata illustrated by Sandberg et al. (1972). Specimens from Berchogur appear to be more derived in the praesulcata/sulcata lineage, as can be judged from Flajs and Feist’s (1988, p. 74) summary of differences between these two species based on the angle of the carina curvature. In specimens from Member 2b, this angle is less than 12°, hence we identified them as S. (Eo.) praesulcata. The specimen illustrated in Fig. 6H, with a curvature of ca. 10°, is an intermediate form. This specimen appears to be similar to the specimen illustrated by Becker et al. (2013, pl. 1, figs. 6a–b) from Lalla Mimouna as S. (Eo.) sp. A. In Fig. 6i–k the angles appear to be less than 12° as in 6H, however, this is an artefact of the photography, i.e., the specimens are slightly inclined, so the carinae appear to be more strongly curved. All these specimens possess a pseudokeel and can therefore be assigned to S. (Eo.) praesulcata. The Last Appearance Datum (LAD) of Icriodus costatus, Mehlina strigosa, and Apatognathus varians cipitis at the top of Member 2a probably correlates with the main conodont extinction defining the base of the costatus-kockeli Interregnum of Kaiser et al. (2009). The lower part of Member 2b is interpreted as the probable record of a transgression, which can probably be correlated with the top part of the Hangenberg Sandstone (see Becker et al. 2016).
-
3.
The Upper praesulcata Zone correlates with the interval of the Lower Protognathodus fauna of Ziegler (1962), the Upper praesulcata Zone of Ziegler and Sandberg (1990) and Corradini (2003), the kockeli Zone sensu Kaiser et al. (2016), and the lower part of the Pr. kockeli Zone sensu Corradini et al. (2017) and Spalletta et al. (2017). The entry of S. (Eo.) cf. praesulcata in Member 2b (Fig. 7hk) is not the FAD of the zonal index species characterizing the Lower praesulcata Zone because the Berchogur specimens have a more derived morphology. The assemblage also contains Polygnathus inornatus (Fig. 7f, g), which can also indicate the upper part of the praesulcata Zone. In the Berchogur sections there are no records of Protognathodus kockeli. However, this level (= kockeli Zone) is known to contain associations of Bispathodus, whereas one of these, B. spinulicostatus (Branson, 1934) (Fig. 7o, p) frequently occurs in this zone (Corradini et al. 2017). By correlation, largely based on sea-level changes, the level of the FAD of Pr. kockeli could be expected in the uppermost beds of Member 2b (dark shale unit above the limestone with bivalves) (Fig. 3).
-
4.
The sulcata Zone is recognized based on the presence of the index species Siphonodella (Eosiphonodella) sulcata (Fig. 7a, b) and several representatives of the genus Bispathodus (Bispathodus aculeatus aculeatus (Branson and Mehl, 1934a) (Fig. 7q–s), B. aculeatus anteposicornis (Scott, 1961), B. spinulicostatus and B. stabilis vulgaris (Dzik, 2006) (Fig. 7t)). This zone also contains Neopolygnathus communis (Branson and Mehl, 1934b) (Fig. 7d). It is noteworthy that the first appearance of S. (Eo.) sulcata in this section is fixed ca. 20 cm above the base of the limestone beds of Member 3 (Unit 3b) in Trench BK-1 (Sample BK-19-12/3) and in Unit 3c in Trench BK-3 (the latter based on Barskov et al. 1984). The assemblage contains Pseudopolygnathus cf. fusiformis, which occurs in the uppermost Famennian and basal Tournaisian. This occurrence is at approximately the same level as the first ammonoids that we found in 2018–2019 and agrees with Barskov et al.’s (1988) conclusion that the sulcata Zone can only be reliably identified in Member 3. The level of the base of the sulcata Zone within Member 2b (as suggested in Maslov 1987) is not upheld in this paper, as we so far have no evidence of that. Considering the lithology (change from clay to wackestones with algae and rare foraminifers and then to wackestones with ammonoids) and the presence of Acutimitoceras and Siphonodella, Member 3 can be interpreted as a transgressive phase of the evolution of the basin, and at least its upper part (3c) can be considered an equivalent of the Stockum Limestone of the Rhenish Massif (Clausen et al. 1994) and basal Hastière Formation of the Ardennes (Becker et al. 2016, etc.). The genus Bispathodus is represented by Bispathodus aculeatus aculeatus found in the lower part of Member 3 (Sample BK1-19-12/1). This subspecies appeared at the base of the Middle expansa Zone and continued to the lower Tournaisian (Ziegler and Sandberg 1984; Corradini et al. 2017). Polygnathids are represented by Neopolygnathus communis (Branson and Mehl, 1934b) (Sample BK1-19-12/1) (Fig. 3). Neopolygnathus communis ranges from the lower Famennian to the lower Tournaisian (Corradini et al. 2017). It is possible that there is a short hiatus between Members 2b and 3, as Yu. Gatovsky observed in 2019 a ferruginous crust 1–2 cm thick between clays of Member 2b and limestones of Member 3, whereas the basal Member 3a is slightly brecciated.
Foraminifers
Foraminifers of the Zhangana Formation were first studied by Reitlinger (1961) based on Rozman’s (1960, 1962) collections from the Berchogur Depression. More than 20 years later, Reitlinger and Kulagina (in Simakov et al. 1985; Barskov et al. 1984; Maslov 1987) published the results of their study of foraminifers from the Burtybai and Berchogur sections based on material collected in 1978–1983. Foraminifers of boreholes BS-1, BS-2 and trenches BK-1, BK-2, and BK-3 were studied by Reitlinger, and those of “Outcrop 8” were studied by Kulagina, Pazukhin and Kochetkova (Maslov 1987, Table 5). Based on all these data, the following foraminiferal beds were recognized from bottom to top: (1) Beds with unilocular foraminifers and rare tournayellids (89.5 m, units I-IV, Kushelgian Regional Substage), (2) Beds with Septatournayella potensa—rare Quasiendothyra communis simplex (22 m, Unit V, Lytvian Regional Substage), (3) Beds with Quasiendothyra regularis (Units VI and VII, 85.5 m in total thickness, including covered intervals); (Lytvian Regional Substage), and (4) Beds with Quasiendothyra kobeitusana substricta and Endothyra imminuta (later Endoglomospiranella imminuta in Kulagina 2013) (30 m, Unit VIII of the Burtybai section, Members 1 and 2a in Borehole BS-1, and Member 2a in Borehole BS-2, Lytvian Regional Substage), (5) Beds with Tournayellina pseudobeata (about 4.5 m, Unit IX of the Burtybai section and in “Outcrop 8”, and Member 3 in the Berchogur section, Gumerovian Regional Substage). However, Unit IX in the Burtybai section was studied in isolated exposures, and in “Outcrop 8” it included a covered basal interval 1.3 m thick, possibly composed of shale.
In the correlation scheme in Maslov (1987, text-fig. 7, insert), Beds 3–5 in “Outcrop 8” (total thickness ca. 1.5 m) were correlated with the upper part of Member 2b and Member 3 of the Berchogur sections and the “Beds with T. pseudobeata” were recognized in Beds 4–6, 0.8 m thick. Later the base of the Beds with T. pseudobeata in “Outcrop 8” was drawn at Bed 1 = upper part of Unit VIII, approximately m below its top based on the first appearance of the zonal index (Kulagina 2013). This boundary corresponds to a level in the lower part of Member 2a in the Berchogur sections (Maslov 1987).
Specimens identified as T. primitiva Lipina, 1965 by Barskov et al. (1984, pl. 3, figs. 1–4) and Simakov et al. (1985, p. 18) were later reidentified as T. pseudobeata Reitlinger and Kulagina in Maslov, 1987 (p. 73).
In 2018, Mustapayeva collected foraminiferal samples from natural outcrops along the upper reaches of Burtybai Creek and Locality 3, approximately at places where in 2019 we re-excavated Trenches BK-1 and BK-3. In 2019 we received new material from re-excavated trenches BK-1 and BK-3, re-examined thin sections from the collection of Reitlinger (Geological Institute, Russian Academy of Sciences, Moscow) from samples collected by Kononova from Borehole BS-1 in 1983.
In this paper we recognize: (1) Quasiendothyra kobei-tusana substricta Zone and (2) Tournayellina pseudobeata Acme Zone in the Berchogur sections. Trench BK-1 (Fig. 3) and Borehole BS-1 (Fig. 4) revealed the upper 2–3 m of the Quasiendothyra kobeitusana substricta Zone. In Borehole BS-2 (Fig. 4), samples 54 and 55 (see Maslov 1987, Table 5) contained an impoverished assemblage with long-ranging species, including Glomospiranella rara Lipina, 1955, Septatournayella potensa Durkina, 1959, Septaglomospiranella bouckaerti Conil and Lys, 1964 and Quasiendothyra (Eoendothyra) communis (Rauser-Chernousova, 1948). The Tournayellina pseudobeata Acme Zone is studied in Trenches BK-1 and BK-3.
Quasiendothyra kobeitusana substricta Zone. This unit is defined by the presence of the zonal subspecies Quasiendothyra (Quasiendothyra) kobeitusana substricta Conil and Lys, 1964 and the upper boundary is drawn by its disappearance. We recognize the Quasiendothyra kobeitusana substricta Zone, rather than the Beds with Quasiendothyra kobeitusana substricta—Endoglomospiranella imminuta, because Endoglomospiranella imminuta is found in the upper part of Unit VIII in “Outcrop 8” of Maslov (1987), where Q. (Q.) kobeitusana substricta is not found and is correlated with Unit 2a2 in the Berchogur sections. E. imminuta apparently continues higher up the section, compared to Q. (Q.) kobeitusana substricta.
Only the uppermost part of the Quasiendothyra kobeitusana substricta Zone is recognized in Borehole BS-1 where it corresponds to Members 1 and 2a (about 3 m).
In Member 1, the algal-foraminiferal wackestones contain dasycladaceans, abundant foraminifers represented mainly by tournayellids and frequent specimens of the genus Paracaligelloides, rare Quasiendothyra (Eoendothyra) communis (Rauser-Chernousova, 1948) is also present. Representatives of Q. (Eoendothyra) regularis (Lipina, 1955) are rare and small-sized (Maslov 1987). In the uppermost part of Member 1 the first occasional specimens of T. pseudobeata appear (Sample 36, in Maslov 1987, Table 5; Fig. 9q in this paper) (Fig. 4). In the upper horizons of Member 2a1, thin interbeds of argillaceous and nodular limestones (samples 27, 26) are dominated by Q. (Eoendothyra) communis and Q. (Eoendothyra) regularis, less commonly Q. (Quasiendothyra) kobeitusana substricta and Q. (Eoendothyra) delicata Durkina, 1959; Septaglomospiranella spp. (Maslov 1987, Table 5). In Bed 2a (Member 2a2, samples 23, 24) algal limestones contain dasycladaceans and the calcified cyanobacterium Girvanella. The abundance and diversity of specimens of Quasiendothyra considerably decrease. The bed contains only small-sized Q. (E.) communis and juvenile specimens of Quasiendothyra unidentifiable to species.

Foraminifers of the Berchogur Section: a–m Trench BK-1; p Trench BK-3; n, o locality 3; a–p collected by S. N. Mustapayeva, 2018–2019; collection no. 7/2019 is housed at the Institute of Geological Sciences, Almaty; q from collection of E.A. Reitlinger, Geological Institute, Russian Academy of Sciences (Moscow) (GIN RAS) (Kulagina et al. 2021, text-Fig. 7b); a Parathurammina aff. dagmarae Suleimanov, 1945, Member 3, Sample 19-1/2a; b Bisphaera malevkensis Birina, 1948, Member 2a2, sample19-6/4; c Baituganella ex gr. tchernyshinensis Lipina, 1955, Member 1, Sample 19-1/1; d Tournayella (Eotournayella) aff. jubra Lipina and Pronina in Lipina, 1965, median section, Member 1, Sample 19-1/1; e Septaglomospiranella sp., nearly axial section, Member 1, Sample 19-1/1; f Septabrunsiina cf. bertchogurica Reitlinger and Kulagina, 1987, nearly axial section, Member 2a2, Sample 19-6/4; g Quasiendothyra (Eoendothyra) ex gr. regularis (Lipina, 1955), axial section, Member 1, Sample 19-1/1; h Quasiendothyra (Quasiendothyra) kobei-tusana substricta Conil and Lys, 1964, tangential section, Member 1, Sample 19-1/1; i, j, m Tournayellina pseudobeata Reitlinger and Kulagina, 1987, median sections, Member 3: i Sample 19-12/5, j, m Sample 19-12/1; k cyanobacterial limestone, consisting of Girvanella sp., Member 1, Sample 18-1/1 (level of 19-1/1); l Garwoodia gregaria mugodzharica Reitlinger and Kulagina 1987, Member 3, Sample 19-12/1; n Tournayellina vulgaris Lipina, 1955, Member 3, Sample 18-Bg-7; o Eochernyshinella sp., Berchogur, Locality 3, Member 3, Sample 18-Bg-7; p Tournayellina pseudobeata Reitlinger and Kulagina in Maslov, 1987, median section, BK-3, Member 3, Sample BK-3C; q Tournayellina pseudobeata Reitlinger and Kulagina, 1987, median section, specimen GIN RAS, BC-1/36, Borehole BS-1, Sample 36. Scale bar 0.2 mm (except for k)
In Trench BK-1, Member 1 lies at the base of the trench and is also exposed in the dried channel of the creek. It contains foraminifers, including the zonal subspecies (Fig. 10c–h), and the abundant cyanobacterium Girvanella (Fig. 9k). Limestones of Unit 2a2 contain rare Bisphaera and Tournayelloidea (Fig. 9b, f).

Foraminiferal zonal correlation across the Devonian–Carboniferous boundary, showing the span of the trenches and boreholes. Reg. sub., regional substages; C., Clubovella; P., Prochernyshinella; Q., Quasiendothyra; T., Tournayellina
No foraminifers were found in the overlying Member 2b.
Tournayellina pseudobeata Acme Zone. The lower boundary of this unit is recognized based on the disappearance of the index subspecies of the previous unit and most species of the underlying beds and on the mass occurrence of T. pseudobeata.
These beds are identified in Member 3a of Borehole BS-1 and in trenches BK-1, BK-2, and BK-3 (Simakov et al. 1985; Maslov 1987; Barskov et al. 1988; Kulagina et al. 2016) (Figs. 3, 4), and Locality 7 (Sample 19BK3-4a). This unit is characterized by the disappearance of the genera Septaglomospiranella, Septabrunsiina, Quasiendothyra and Endoglomospiranella occurring in the underlying beds, whereas Tournayellina pseudobeata becomes abundant (Fig. 9i, j, m, p), and Archaesphaera and Paracaligelloides are found less commonly. A sample collected from Member 3 in 2018, before Trench BK-3 was re-excavated in 2019, contained Tournayellina vulgaris Lipina, 1955 (Fig. 9n, and Eochernyshinella sp. (Fig. 9o). The samples also contain abundant algae Garwoodia gregaria mugodzharica Reitlinger and Kulagina in Maslov, 1987 (Fig. 9l).
In Trench BK-3, Member 3b contains rare specimens of Earlandia minima Birina, 1948 (see Fig. 8a).
The Q. kobeitusana Zone is represented in most sections of the D–C boundary beds in Russia and Kazakhstan. This zone correlates with the Q. kobeitusana substricta Zone of the Berchogur sections, whereas Tournayellina pseudobeata is found in a very limited number of sections (Kulagina et al. 2016, 2018).
In the Timan-Pechora Province (Kamenka section), the first appearance of T. pseudobeata is recorded beginning from the base of the Siphonodella sulcata Zone (Vevel 2009; Vevel et al. 2012; Kulagina et al. 2021). In the South Urals, in the Sikaza section, two small specimens of this species are found in the upper part of the Gumerovian Regional Substage in association with S. sulcata (Kulagina et al. 2016). In Borehole Melekess no. 1 (Volga-Urals Province) the first occurrence of T. pseudobeata was recorded at approximately the base of the Siphonodella sulcata conodont Zone (Kulagina et al. 2021).
The Beds with Q. kobeitusana substricta correlate with the uppermost Famennian DFZ7 of Poty et al. (2006; but included in the basal Tournaisian in Poty 2016) with Quasiendothyra kobeitusana (Rauser-Chernousova, 1948), Q. konensis (Lebedeva, 1956), Klubovella konensis Lebedeva, 1956, Laxoendothyra parakosvensis (Lipina, 1955), Septabrunsiina kingirica (Reitlinger, 1961), S. krainica (Lipina, 1948), abundant Paracaligelloides sp. (Poty et al. 2006, text-fig. 4), and with Quasiendothyra kobeitusana-Q. konensis Zone of the Moravian Karst (Czech Republic) (Kalvoda et al. 2015) (Fig. 10).
The Tournayellina pseudobeata Acme Zone most likely correlates with DFZ8 of the Avesnelles Formation of Belgium and northern France with key elements: T. pseudobeata, Chernyshinella spp., Ch. gutta (Conil and Lys, 1964); Laxoendothyra. parakosvensis nigra (Conil and Lys, 1964) (= Endoglomospiranella nigra), Septabrunsiina spp., S. kingirica (Reitlinger, 1961), Septaglomospiranella spp., and Earlandia moderata (Malakhova, 1954) (Poty et al. 2006, p. 832, text-fig. 4). Species of the genus Quasiendothyra (Poty et al. 2006, text-fig. 5) do not continue into this zone. Therefore, it is possible to propose a correlation based on the disappearance of Quasiendothyra and abundant occurrence of T. pseudobeata. Note that the foraminiferal assemblages in the Avesnelles Formation is considerably richer and contains species, which appear higher in the Uralian and Kazakhstanian successions, in the Chernyshinella disputabilis Zone.
In the Moravian Karst, T. pseudobeata is known in several sections (Kalvoda et al. 2015). In the Mokrá Quarry composite section, the first appearance of this species is fixed at the base of the S. sulcata Zone in the oolitic bioclastic calciturbidites, where various quasiendothyrids are associated with that species, including Q. kobeitusana, Q. konensis, Q. dentata, Eoseptatournayella spp., Laxoendothyra alta (Conil and Lys, 1964), and other species (Kalvoda et al. 2015, text-fig. 10). However, the underlying beds corresponding to the Ckl conodont interval (costatus-kockeli Interregnum and kockeli Zone) is represented by mudstone/marlstone without foraminifers. In the Lesní lom and Anaklety composite section T. pseudobeata first appears at the boundary of the costatus-kockeli Interregnum and kockeli Zone also in association with the assemblage of Quasien-dothyridae (Kalvoda et al. 2015, fig, 14). In South China, T. pseudobeata is found in the MFZ1—Unilocular Zone of the Malanbian Province, Menggongao Formation (Hance et al. 2011).
Ammonoids
Balashova (1953) listed ammonoids from the Berchogur Depression, but her “Imitoceras subbilobatum” has been reidentified as Acutimitoceras dzhanganense Nikolaeva, 2020, and her “Imitoceras substriatum” as A. alabasense Nikolaeva, 2020. Her localities and taxa reported there are as follows: (1) Aktan and Kust-Kara dry creeks: Imitoceras sp., “I.” cf. rotiforme Librovitch, 1940, “I. subbilobatum (Münster, 1839)”, (2) middle reaches of Sarysai Creek: “I.” cf. rotiforme, “I. subbilobatum”, (3) west of the Zhan-Gana Range, Tungulyk-Bulak Gully: “I. substriatum”, “I.” bertchogurense (= “I. angustiferus” Balashova, 1953 nom. nud. in the caption of pl. XII), “I. subbilobatum”; (4) Kabaksai Creek: “I. subbilobatum”. The same ammonoid species were cited by Librovitch (1940, 1957) and Rozman (1962). The precise position of these localities is unknown, but the rivers and creeks near the localities indicated by Balashova and Rozman in their publications are shown in Fig. 1.
Ammonoids collected in 1981 and 1983 by several teams of researchers were identified by Kusina (in Barskov et al. 1984; Kusina, 1985) and discussed by Bogoslovsky (1986) and Bogoslovsky in Maslov (1987). The taxonomy of the species-rich genus Acutimitoceras is still debatable. It comprises a variety of species with and without constrictions, with a long or short ventral lobe, and with or without a carinate venter. Becker (1996) proposed several subgenera within Acutimitoceras, apart from the nominal subgenus (type species A. acutum Schindewolf, 1923). He named Stockumites, with the type species Imitoceras intermedium Schindewolf, 1923 and Streeliceras (type species Imitoceras heterolobatum Vöhringer, 1960), and considered Sulcimitoceras Kusina, 1985 as a subgenus of Acutimitoceras Librovitch, 1957. Korn and Klug (2002) considered Stockumites Becker as a junior synonym of Acutimitoceras and the genus Streeliceras Becker as a junior synonym of Nicimitoceras Korn, 1993 (type species Imitoceras subacre Vöhringer, 1960). Korn and Klug (2002) considered Sulcimitoceras Kusina, 1985 as a separate genus. The range of morphological variation in species currently included in Stockumites is quite wide and is likely to warrant further subdivision. The two Berchogur species A. dzhanganense and A. alabasense, show some affinity with the type species A. (Stockumites) intermedium, but their cross sections and sutures are quite different (see Korn 1994, text-figs. 59B, C; Nikolaeva, 2020, text-figs. 3 and 5), and it is possible that these two species could be assigned to a different subgenus; we do not make a final decision on their subgeneric assignment in this paper. We tentatively assign a single loosely collected specimen identified as Imitoceras (Acutimitoceras) carinatum Schmidt, 1924 by Kusina (1985, text-fig. 3b, 1i), to the genus ?Nicimitoceras, taking into consideration that the early stages are noticeably less evolute than in Streeliceras heterolobatum (see Korn 1994, text-fig. 59E). In addition, the ventral lobe in the Berchogur specimen (Kusina, 1985, text-fig. 3b) is not shortened, as in Streeliceras. At the same time, the early whorls of this specimen (Kusina 1985, text-fig. 1i) are more evolute than in N. subacre (Vöhringer 1960, p. text-fig. 2; Korn 1994, text-fig. 59C). Therefore, the generic affinity of this specimen remains problematic. In the description, Kusina (1985) noted that this specimen develops a carinate venter on the sixth whorl (later than in the type material of carinatum Schmidt) and has a wider shell; the Berchogur specimen is likely to represent a new species, but more material is needed for a positive identification.
In 2018–2019 ammonoids were again collected from Members 3 and 4. The assemblage is similar to those described by Kusina (1985) from these members and contains “Imitoceras” bertchogurense (Balashova, 1953) (Fig. 11o, p); Acutimitoceras (Stockumites?) dzhanganense Nikolaeva, 2020 (Fig. 11i–n) [identified as “I. (A). subbilobatum” by Kusina, 1985], A. (Stockumites?) alabasense Nikolaeva, 2020 (Fig. 11a–h) [identified as “I. (A.) substriatum” (Münster, 1840)” by Kusina 1985], A. (Stockumites?) mugodzharense Kusina in Barskov et al., 1984, A.? or N.? cf. carinatum (Schmidt, 1924), and Sulcimitoceras yatskovi Kusina, 1985 (re-illustrated by House, 1993, text-fig. 2). Member 4 yielded A. (Stockumites?) pulchrum Kusina, 1985 (formerly identified as “A. aff. rotiforme (Librovitch, 1940)” in Barskov et al. 1984)) and possibly A. (Stockumites?) alabasense Nikolaeva, 2020 (previously identified by Kusina (1985) as “I. (A.) substriatum”) (see Kusina 1985; Barskov et al. 1988). In Member 3 we were able to identify only one level with ammonoids (in Member 3c), as opposed to two levels indicated by Barskov et al. (1984), despite considerable excavations. As indicated by Barskov et al. (1984) both ammonoid horizons were confined to 15 cm interval in Member 3 (their samples 3/4 and 3/5). After profound excavation in 2019 using a mechanical digger it has become apparent that Member 3 is composed of three limestone beds separated by clay intervals (Fig. 5). In 2019 ammonoids were only found in Member 3c.

Ammonoids from the Berchogur (Birshogyr) sections (including loosely collected specimens), Zhangana Formation; a–h Acutimitoceras alabasense Nikolaeva, 2020: a, b holotype PIN, no. 4005/147; Member 3; c, d specimen PIN, no. 4005/122; Member 3; e, f specimen PIN, no. 4005/129; Member 3; g, h specimen PIN, no. 4005/115; Member 3; i–n Acutimitoceras dzhanganense Nikolaeva, 2020; i, j specimen PIN, no. 4005/195; Member 3; k, l holotype PIN, no. 4005/126; Member 3; m, n specimen PIN, no. 4005/196; Member 3; o, p “Imitoceras” bertchogurense (Balashova, 1953), PIN, no. 5643/39; Member 3; scale bars a–f, i–p 1 cm; g, h 0.5 cm
The ammonoid assemblage of Member 3c belongs to the Acutimitoceras Genozone based on the occurrence of the index genus. In addition to Acutimitoceras (prionoceratids with evolute inner whorls), the assemblage contains species with involute or semiinvolute inner whorls, “I.” bertchogurense, and N.? (or A.?) cf. carinatum; the latter is similar to specimens from the prorsum Zone in its type region in Germany (Korn 1994; Becker et al. 2016). It should be also noted that A. dzhanganense is similar to specimens commonly assigned to A. (Stockumites) subbilobatum (even though A. dzhanganense has no constrictions), which are found in the prorsum Zone in Franconia, in the Rhenish Massif, and in Morocco (Becker et al. 2016). It is not clear whether the German and Moroccan specimens attributed to A. (S.) subbilobatum represent the same species. Therefore, the assemblage from Member 3 probably correlates with that of the prorsum Zone. Becker (1996) proposed to consider the size of the shells developing a carinate venter, suggesting that the development of the oxyconic morphotypes could be shifted to earlier ontogenetic stages in the evolution of the genus Acutimitoceras. For instance, a previously figured specimen of Acutimitoceras sp. from Berchogur (Acutimitoceras sp., Barskov et al. 1984, pl. 1, figs. 8a–b, “Imitoceras (A.)” sp., Kusina 1985, pl. III, fig. 8), developed an oxyconic shell at 15 mm shell diameter, whereas in A. acutum from the eponymous zone in its type region in Germany, such shell morphology is present at the shell diameter of 6–8 mm, in the fourth to fifth (see Korn 1994, text-fig. 50B) whorl. More similar to the Berchogur specimen is A. (A.) oxynotum Bartzsch and Weyer, 1996 from Thuringia, which becomes oxyconic at ca. 15 mm in diameter or even somewhat later (paratype MB.C.1976). Both forms can be distinguished by different types of shell constrictions (very short and only ventral in the Berchogur specimen). Unfortunately, the precise level of occurrence of A. (Acutimitoceras) sp. at Berchogur is not clear (Kusina 1985; Bartzsch and Weyer 1996, p. 53); the specimen was not collected in situ. The ammonoid fauna of Berchogur can be tentatively correlated with that of the lower part of the Wangyou Formation in China in the Muhua sections (Ruan 1981; Qie et al. 2015), containing A. wangyuense Sun and Shen, 1965. Becker and Weyer (2004, pp. 29–30) discussed A. wangyuense and excluded it from Acutimitoceras. It is most likely an early karagandoceratid species and was cited as ?Bartzschiceras wangyuense. It was also re-emphasized (following Bartzsch and Weyer 1996, p. 93) that Sun and Chen’s original specimen and the supposed later wangyuense specimen of Ruan (1981) are not conspecific. In China, Acutimitoceras sp. were found in the same formation in the S. sulcata Zone, or at the base of the S. duplicata Zone (Member C in the Gedongguan section (Youjiang Basin, Changshun County, Guizhou) (Qie et al. 2015, text-fig. 4). Similar shell morphology is observed in ammonoids from Faunule 1 (Bed RTB 10) of the Gara Bou Tlidat section, in the lower part of the Fezzou Formation in Morocco (Becker et al. 2018) but this level also includes gattendorfiids and is younger than the Stockum Limestone (Fig. 12). The position of the ammonoid fauna in Berchogur in relation to the conodont zones is currently under investigation (see “Discussion”).
In summary, this ammonoid fauna indicates that the Carboniferous ammonoids appeared in this section during an apparent phase of explosive radiation in the “upper Crisis Interval” (sensu Becker et al. 2016).

Corals
Corals were reported from the Berchogur sections by Barskov et al. (1984). From Members 1 and 2, they listed Cyathaxonia aff. cornu Michelin, 1846 in Michelin 1840–1847, Cyathaxonia sp., ?Clisiophyllum sp., Axophyllum aff. praecursor (Frech, 1885), ?Cyathoclisia sp. and Campophyllum flexuosum (Goldfuss, 1826). The latter is typically Strunian (uppermost Famennian) taxon, as well as Axophyllum aff. praecursor (Bounophyllum aff. praecursor in updated taxonomy), and probably “Clisiophyllum” sp. Cyathaxonia is a long-ranging taxon with limited use in stratigraphy. As the corals were not illustrated, attributions remain uncertain. Nevertheless, this association still indicates a Devonian age for the lower two members of the section.
The present ca. 180 specimens were mostly collected at the top of Member 3, either in trenches BK-1 and BK-3 and Locality 5 (Fig. 1) or as loose specimens on the surface. Besides fragments of indeterminable syringoporids s.l., the corals are mostly represented by small solitary and some, larger, dissepimented rugose corals. A short description of this fauna is provided here.
Cyathaxonia cornu (Fig. 13g–h) is abundant, ranging from very small (5 mm-high, < 1 mm diameter) to relatively larger (1.8 mm-high, 4 mm in diameter) specimens. Larger specimens (6–8 mm in diameter, 30 major septa) with an elliptic columella can be related to Cyathaxonia aff. rushiana (Vaughan, 1908) but that species needs to be revised.

Rugose corals from the Berchogur sections (abbreviations: CV, external calicular view; LS, longitudinal thin section; TS, transversal thin section); a Molophyllum sp. (specimen BK.1.L.1., loose from Trench BK-1, TS); b–e Conilophyllum priscum (Münster, 1840); (b–c specimen BK1.3a.1 from Trench BK-1, Member 3a, TS, LS; d BK1.3L.2, loose from Trench BK-1, Member 3, TS; e BK1.L.4, loose from Trench BK-1, TS); f Proheterelasma omaliusi De Koninck, 1872 (specimen BK1.3L.44, loose from Trench BK-1, Member 3, CV); g–h Cyathaxonia cornu Michelin, 1846 in Michelin 1840–1847 (specimen BK5.3.37, from Locality 5, Member 3, external views); i–l Rylstonia brevisepta Hudson, 1942 (i–j specimen BK1.L.7, loose from Trench BK-1, TS, LS; k BK1.3L.34, loose from Trench BK-1, Member 3, CV; l BK1.3L.42, loose from Trench BK-1, Member 3, CV); m unidentified uraliniid coral (specimen BK1.L.3, loose from Trench BK-1, Member 3, TS); n–q Caninia cf. cornucopiae Michelin, 1840 in Michelin 1840–1847 (n specimen BK1.3L.50, loose from Trench BK-1, Member 3, CV showing early growth stage with elongated counter septum; o BK1.3.6, from Trench BK-1, Member 3, TS; p BK1.3L.14, loose from Trench BK-1, Member 3, CV, late growth stages; q BK1.3L.61, loose from Trench BK-1, Member 3, CV, columellate late growth stage); r-s “Permia” cavernula Hudson, 1943 (r specimen BK5.12, from Locality 5, TS; s BK1.3L.26, loose from Trench BK-1, Member 3, CV)
Proheterelasma omaliusi De Koninck, 1872 (Fig. 13f) is represented by a single solitary ceratoid coral, 4–5 mm in diameter and with 22 major septa grouped in four bundles joining the axial end of the cardinal columella.
Caninia cf. cornucopiae Michelin, 1840 in Michelin 1840–1847 (Fig. 13n–q) is trochoid or trocho-ceratoid, 8–20 mm-high, 4–15 mm in diameter and with 22–26 major septa in mature stages. It displays thickened septa in the cardinal quadrants (convex side of the corals) and conspicuous septal furrows on the outer surface. The major septa are usually withdrawn in adult stages, but the counter septum is commonly elongated and reaches the axis where it can form a rudimentary axial structure with the upturned tabulae. The cardinal septum in the well-developed cardinal fossula is short. The minor septa appear as crests on the external wall. Dissepiments occur only in corals > 10 mm in diameter. These corals display a wide morphological variability but the presence of thickened cardinal quadrants, cardinal fossula, and elongated counter septa are relatively stable characters. We refer this population to C. cornucopiae with caution, as the material is poorly preserved, hampering the proper observation of internal characters, and because their size is relatively small. Similar corals occurring in the Sultsiferovyi (sulcifer) Horizon and Karakingir Beds (Upper Famennian) of Mount Aktas were described as Caninia cornucopiae by Ulitina (1975), whereas Volkova (1941) reported similar specimens under the name Laccophyllum fossulatum from the beds in 1941 assigned to the “lower Tournaisian” of the Shiderty River, Karaganda region of Kazakhstan. If Volkova’s material could be re-examined, these specimens could be attributed to the species Canina fossulatum (Volkova, 1941); the illustration does not allow suitable comparison.
Rylstonia brevisepta Hudson, 1942 (Fig. 13i–l) is a small (< 10 mm in diameter, 10–15 mm-high, 18–22 major septa) ceratoid solitary coral with frequent constrictions and episodes of rejuvenescence. Its axial structure made of thickened axial plate and radial lamellae is prominent in the calices. Its dimensions and narrow dissepimentarium make it comparable to specimens of R. brevisepta described by Hudson (1942) (as a variety of the late Tournaisian species R. benecompacta Hudson and Platt, 1927) from the upper Courceyan of the British Isles. The columellate corals from the upper Kassinian Regional Substage (lower Tournaisian) of the Kyzylzhal Mountains of Kazakhstan, described as Rylstonia cystosa Keller, 1959, are probably a junior synonym of Hudson’s (1942) species as well as Litvophyllum karakingiria Keller, 1959 and possibly Dibunophyllum urunsaica Keller, 1959, also from the upper Kassinian, which differs from R. cystosa only in the development of few transeptal dissepiments.
Conilophyllum priscum (Münster, 1840) (Fig. 13b–e) is a cylindrical coral 10–14 mm in diameter, having weakly thickened 24–30 major septa. The minor septa are short and commonly contraclinant. The dissepimentarium comprises rare interseptal or transeptal dissepiments. The cardinal fossula is inconspicuous and the tabulae are flat, mesa-shaped. The outer wall is festooned. Despite the rarity of dissepiments, the material can be attributed with confidence to the Hastarian (lower Tournaisian) species Conilophyllum priscum of Western Europe, which commonly displays a wide range of variation (Poty and Boland 1994). The species was described as Kassinella longiseptata Keller, 1959 from the upper Kassinian Regional Substage (lower Tournaisian), reported from Mount Kyzylzhal in the Zhezkazgan (formerly Dzhezkazgan) area and identified as Guerichiphyllum kazakhstanicum Ulitina, 1975 from the Sultsiferovyi (sulcifer) Regional Substage (Upper Famennian) of Mount Aktas (Ulitina 1975). Both should be regarded as junior synonyms of Conilophyllum priscum.
“Permia” cavernula Hudson, 1943 (Fig. 13r–s) is ceratoid to patellate, up to 10 mm in diameter, having 24 major septa. In the early growth stages, the septa are long and rhopaloid and form an axial column with a narrow lumen. In later stages, the septa withdraw and form an aulos. Fedorowski (1971) synonymized Permia with Aulophyllum Milne-Edwards and Haime, 1850; hence a new generic name is required for these species formerly assigned to ‘Permia’. Ulitina (1975) figured Amplexocarinia muralis from the Sultsiferovyi (sulcifer) Regional Substage (Upper Famennian) of Mount Aktas, that could be conspecific with the present material.
One fragment of a cylindrical coral 20 mm in diameter can be attributed to Molophyllum sp. (Fig. 13a). It has 33 major septa, strongly thickened in all quadrants of the tabularium, joined in bundles that almost reach the axis of the corals. The minor septa are short and thin. The cardinal and counter septa are of similar length and there is no conspicuous fossula. The dissepimentarium comprises 1–2 rows of transeptal dissepiments and 2–3 rows of concentric interseptal dissepiments. The wall is thick and smooth. These characters point towards the genus Molophyllum Onoprienko, 1979, but no published species of that genus is so small. Therefore, it seems appropriate to retain it in open nomenclature until more material is available.
A last very eroded fragment of cylindrical coral (Fig. 13m) displays only the thickened tabular part of the major septa divided in four quadrants by four open fossulae. This single specimen is possibly a uralinid (or keyserlingophyllid) coral, but its preservation precludes a more precise attribution.
Of these eight coral taxa, only Molophyllum sp. has an Upper Devonian affinity as it was described from the uppermost Famennian deposits of the Omolon (Onoprienko 1979; Shilo et al. 1984). Conilophyllum priscum also occurs in the uppermost Famennian of Omolon (Poty 1999) but only in the lowermost Tournaisian (Hastarian) of Western Europe, where it is the guide-taxa of the RC1α subzone of Poty et al. (2006). Cyathaxonia cornu, Caninia cornucopiae, and uralinids typically occur in lower Tournaisian strata in Eurasia (Poty et al. 2006) whereas “Permia” cavernicula was collected from the lower Courceyan shales of Ireland, attributed to the RC2 zone. Columellate Caninia cornucopiae, Rylstonia brevisepta, and Proheterelasma omaliusi appear in the lower upper Tournaisian (Ivorian Substage of NW Europe) RC3α zone, but early occurrences in Central Asia cannot be excluded.
In summary, most of the coral fauna recovered from the Berchogur trenches point to a (lower or basal upper) Tournaisian age, with the notable exception of the possibly Devonian Molophyllum sp.
Crinoids
Numerous well-preserved crinoid columnals and pluricolumnals are found in the D–C boundary beds of the Berchogur section (mostly at the top of Bed 3c and loose). The collection is housed at the TsNIGR Museum, St. Petersburg, no. 12735.
The assemblage is characterized by the family Floricyclidae (Fig. 14a–e, h). This family was established by Moore and Jeffords (1968) based on crinoid stems from various Mississippian and Pennsylvanian localities of North America. It represents a morphologically well-defined taxonomic group with a clearly outlined pentalobate lumen surrounded by a perilumen, a wide areola, and a coarse crenularium. Later, members of this family were identified from the (mainly lower) Carboniferous of many regions of Kazakhstan, South Tien-Shan, South China, Mongolia, Transbaikal, Altai, South Urals, and Poland (Dubatolova and Shao 1959; Stukalina 1973; Yeltyschewa and Poljarnaja 1975; Dubatolova 1976; Głuchowsky 1981, etc.). In Kazakhstan, the earliest occurrences of this family are from the Sultsiferovyi (sulcifer) Regional Substage of the Upper Famennian and, in addition, they are common in the Simorinian Regional Substage, and also in Tournaisian and Viséan beds (Sisova 1979, 1983; Chernova and Stukalina 1989). In Transbaikal, members of Floricyclidae are also recorded in the uppermost Famennian (Kotikhinian Regional Substage) with a brachiopod assemblage including Sphenospira julii (Dehée, 1929), the index species of the uppermost Famennian Sphenospira julii—Spinocarinifera nigra Zone (Kurilenko et al. 2002; Kurilenko and Kulkov 2008), although the crinoid assemblage of this interval is mixed, as in addition to the earliest occurrences of the typical Carboniferous species it contains the latest occurrences of Late Devonian taxa. Species of Floricyclidae are also identified in the lower Tournaisian beds of Eastern Transbaikal containing a lower Tournaisian crinoid assemblage. In the Berchogur sections, we identified representatives of the genera Floricyclus Moore and Jeffords, 1968 (F. kazangapicus Sisova, 1983 (Fig. 14a–e); Floricyclus sp.) and Lamprosterigma Moore and Jeffords, 1968 (L. cf. welleri (Moore and Jeffords, 1968)) (Fig. 14h). Floricyclus has an axial canal with long club-like lobes. Lamprosterigma is distinguished by a wide axial canal with short, weakly differentiated lobes. Chernova and Stukalina (1989) showed a trend of successive changes in morphological elements depending on the position of columnals in a stem. Distally, the lumen lobes become narrower, the areola increases, and the crenularium decreases. This trend is also observed in the material from Berchogur (Fig. 14a–c). The species Floricyclus kazangapicus is typical of the Tournaisian of South Kazakhstan (upper part of the Kassinian Regional Substage) and the lower part of the Rusakovian Regional Substage) (Sisova 1983; Chernova and Stukalina 1989) and Floricyclus cf. kazangapicus is known from Eastern Transbaikal (Pavlovian Regional Substage) (Kurilenko et al. 2002). In Transbaikal, deposits of the Pavlovian Regional Substage contain an abundant lower Tournaisian fauna of bryozoans, brachiopods, and crinoids (Kurilenko et al. 2002; Kurilenko and Kulkov 2008). The genus Lamprosterigma is found in the lower Carboniferous of Kazakhstan and North America. Lamprosterigma welleri (Moore and Jeffords, 1968) was described from the lower Mississippian of Kentucky (Brodhead Formation, Borden Group, Osagean) and from the Viséan of Holy Cross Mountains, Poland (Moore and Jeffords 1968; Głuchowsky 1981; Chernova and Stukalina 1989).

Crinoids from the Berchogur (Birshogyr) sections; a–e Floricyclus kazangapicus Sisova, 1983: a, b articular facets of the proximal columnals (Loc. 5); c articular facet of a distal columnal (Member 3); d lateral view of a distal pluricolumnal (Member 3); e lateral view of a proximal pluricolumnal (Loc. 5); f, i Baryschyr sp. (Loc. 5): f articular facet of a columnal, i lateral view of a pluricolumnal: g, j, k Cyclocaudex cf. aptus Moore and Jeffords, 1968: g, j articular facets of the columnals (Loc. 5, loose), k lateral view of a pluricolumnal (Loc. 6); h Lamprosterigma cf. welleri (Moore and Jeffords, 1968): articular facet of a columnal (loosely collected in Trench BK-3, Member 3); l–n Platycrinites? cf. subtuberosus Stukalina, 1973 (loosely collected in Trench BK-3, Member 3). l, m articular facets of the columnals; n side view of a columnal; o–s Gilbertsocrinus? circumvallatus (Yeltyschewa in Dubatolova and Shao, 1959): o articular facet of a proximal columnal (Member 3); s articular facets of a distal columnal (Member 4); p–r lateral views of the proximal pluricolumnals (Member 3); t, u Taranshicrinus sp.: t articular facet of a columnal; u side view of a columnal (Loc. 5); v Goniostathmus sp.: lateral view of a pluricolumnal (Member 3)
The Berchogur assemblage also includes other lower Carboniferous species: specimens similar to Cyclocaudex aptus Moore and Jeffords, 1968 (Fig. 14g, j, k), described from the Lower Mississippian of Kentucky and lower Carboniferous of Poland, Baryschyr sp. (Fig. 14f, i), originally described from the Pennsylvanian of Oklahoma (Moore and Jeffords, 1968), and also Gilbertsocrinus? circumvallatus (Yeltyschewa in Dubatolova and Shao, 1959) (Fig. 14o–s), specimens similar to Platycrinites? subtuberosus Stukalina, 1973 (Fig. 14l–n), and Taranshicrinus sp. (Fig. 14t, u), typical of the Upper Famennian−lower Tournaisian beds (Dubatolova and Shao 1959; Moore and Jeffords 1968; Głuchowsky 1981, 2002; Sisova 1988, etc.).
Columnals of the family Platycrinitidae Austin and Austin, 1842 (Fig. 14l–n) are easily identified. They are bilaterally symmetrical, subdivided by a fulcral ridge into two halves with an areola in each (Stukalina 1973; Dubatolova 1976; Kurilenko 1989; Kurilenko et al. 2002, etc.). The genus Platycrinites Miller, 1821 was established based on calices; hence, when it is identified from columnals, a question mark is traditionally placed next to the generic name. Members of the family are widespread in the Carboniferous of many regions of Kazakhstan, Central Asia, Russia, and North America. The species from Berchogur, similar to Platycrinites? subtuberosus, is distinguished from typical specimens of this species in the presence of columnals of two orders and their smaller size. In Transbaikal, Platycrinites? subtuberosus is typically found in the D–C boundary beds in association with Gilbertsocrinus? circumvallatus, which is known from the Tournaisian of Western Siberia, Kuznetsk Basin, southern Transcaucasia, Altai, Kazakhstan, Far East, Mongolia, and South China (Stukalina 1973; Dubatolova 1976, etc.). The assemblage also includes Goniostathmus sp. (Fig. 14v), a genus also known from the Upper Famennian of Central Kazakhstan and Lower Mississippian of Iowa (Burlington Limestone, Osagean) (Moore and Jeffords 1968).
Columnals of Cyclocaudex, Floricyclus, Gilbertsocrinus and Platycrinites (mostly identified to genus) have been previously recorded from the Tournaisian (sulcata Conodont Zone of Pilton Beds, Fremington Quay, North Devon (England)) (Głuchowsky and Racki 2005) in an assemblage similar to those of Poland, Czech Republic, North America, and Kazakhstan. In summary, the crinoid assemblage from the Berchogur sections supports an early Tournaisian age for the host deposits, as indicated by the conodonts, ammonoids, and corals.
Trilobites
The first Carboniferous trilobites were found in the Berchogur Depression by Jaworski (1916) “in the lowermost Carboniferous”. Weber (1932) based on a considerable number of cranidia collected by Jaworski (1916), librigenae and pygidia, recognized a new species, Phillipsia bitumulata Weber, 1932 (p. 45, pl. IV, figs. 1‒15, text-fig. 8). Later, in a review Weber (1937, p. 58, pl. VI, figs. 33‒35) briefly characterized this species and figured only three specimens (cranidium, librigena, and pygidium). In 1939‒1940, E.A. Balashova and Z.G. Balashov studied Carboniferous outcrops in Mugodzhary (Mugalzhary) and collected ca. 30 trilobites at several stratigraphic levels in a number of localities. This collection was described by Balashova (1956), who identified in total seven species, including three new ones. She identified Typhloproetus (?) granulineatus Balashova, 1956, Cyrtosymbole (Waribole) baiburensis (Weber, 1937), Phillipsia coronata Balashova, 1956, and Ph. carringtonensis Etheridge in Woodward, 1884 from the lower part of the Zhangana Formation, approximately near the Devonian–Carboniferous boundary. It is noteworthy that one rock fragment collected by Balashova contained two cranidia and several pygidia of Ph. carringtonensis, and a cranidium identified as belonging to Cyrtosymbole (Waribole) baiburensis. The upper (lower Tournaisian) part of the Zhangana Formation yielded Phillipsia mugodjarica Balashova, 1956. Two more taxa were identified from considerably younger Tournaisian beds: Phillipsia bitumulata Weber, 1932 in the lower part of the Karabulak Formation (upper Tournaisian) (Petrenko 1977, p. 65) and Ph. cf. kassini Weber, 1937 in the lower part of the “middle” Tournaisian Berchogur Formation (Petrenko and Sultanaev 1977, p. 173).
Trilobites collected in 2019 come from Member 3 (Trench BK-3) of the Berchogur sections. Previously, a few trilobites were found in the member and were identified as Archegonus (Waribole) sp. (Maslov 1987, p. 48; Barskov et al. 1988, p. 175) (this collection is presumed to be lost). Additional trilobites that were not identified were recovered from the yellowish-grey limestone with Tournayellina pseudobeata (Bed 4, Unit IX, “Outcrop 8”), which is located 3 km south of the Burtybai section of Maslov (1987, p. 27).
Our present collection contains two rock samples with trilobites. The first contains an isolated pygidium, the second is a small rock fragment with exuviae of a pygidium, cranidium, and possibly of a free cheek (Fig. 15). In our opinion they are identical to those that were assigned by Balashova (1956) to Phillipsia carringtonensis and Cyrtosymbole (Waribole) baiburensis, which also were found as aggregated group of remains in the Kust-Kara locality (south of Burtybai Creek). Cranidia of all these taxa have a moderately narrow glabella with a very weakly pronounced constriction and a smoothly rounded front margin. The glabella surface, mainly centrally, is covered with small tubercles, which in the studied cranidium were only preserved in the posterior right corner of the glabella (Fig. 15d). An important character is a very wide pre-glabella field with relatively straight front margin. In addition, the fixigenae are also relatively wide. The pygidia are also similar in our specimen from the aggregation of remains; the lateral border is very wide, and the pleural ribs end far from the edge of the pygidium. The number of rings of the axis is 10‒11 (Fig. 15a), whereas in Balashova’s specimens it is 11‒12, and there are 8‒9 pleural ribs. Hahn and Hahn’s (1972, p. 344) revision states that the cranidium identified by Balashova as Cyrtosymbole (Waribole) baiburensis, most likely belongs to Cummingella carringtonensis, but later Hahn and Hahn (2008, p. 80) excluded Balashova’s specimens from C. carringtonensis. It is possible that the cranidium recognized by Balashova (1956) as the new species Phillipsia coronata also belongs to the above species. She described two small incomplete cranidia (ca. 3 mm long) that come from a different locality in the upper reaches of the Shyuldak River. Based on morphology, especially the position of furrows on the glabella, its shape, and raised cornea layer, it is possible to interpret Ph. coronata as young specimens of C. carringtonensis. The generic assignment of Ph. coronata remains debatable: Osmólska (1970, p. 94) tentatively assigned this species to the genus Piltonia, whereas Hahn and Hahn (1972, p. 391) assigned it to the subgenus Phillipsia (Phillipsia). The analysis of Hahn and Hahn’s (2008) catalogue shows that the Berchogur trilobite is most similar to early Tournaisian species of the subgenus Liobolina (Liobolina) Richter and Richter, 1951. In having a considerable wide preglabellar field, they are most similar to the common species Liobolina (Liobolina) nebulosa Richter and Richter, 1951 (Hahn and Hahn 2008, text-figs. 220‒222), from the lower Tournaisian (Hangenberg Limestone), of the duplicata/sandbergi conodont zone of Germany, Austria, and England. However, in that species, the glabella is smooth, in contrast to the Berchogur taxon, which displays a prominently ornamented cranidium. More specimens are needed to identify the Berchogur trilobites more precisely, but until that, this taxon can be provisionally identified as Liobolina (Liobolina) coronata (Balashova, 1956).

Trilobites from Berchogur (Birshogyr). Trench BK-1. Liobolina (Liobolina) coronata (Balashova, 1956); Member 3, lower Tournaisian: a pygidium, specimen PIN, unnumbered; b limestone fragment with group of exuviae of trilobites; c pygidium, specimen PIN, unnumbered; d cranidium, specimen PIN, unnumbered; scale bar 5 mm
Discussion
Renewed interest in the Berchogur sections was a result of searching for and testing of new criteria to correlate the proposed levels for the D–C boundary in regions where it is not possible to use the FAD of the conodont Protognathodus kockeli. This conodont species is common in Western Europe, where it enters slightly above the main regressive phase represented by the “Hangenberg Sandstone Event” (Middle Crisis Interval), i.e., at the base of the Lower Stock-um Limestone = base of the “Upper Crisis Interval” (Becker et al. 2016). This level is above the Hangenberg Black Shale (= main part of the Lower Crisis Interval) separated from it by the Hangenberg Shale/Sandstone (= Regression Interval). The search for the new proposed boundary level, identified by the end of the “Hangenberg Sandstone regressive event” and near the FAD of Pr. kockeli is a current task of the working group of the Devonian–Carboniferous boundary (see references in Becker et al. 2016 and Kaiser et al. 2016). Protognathodus kockeli, like all species of the genus Proto-gnathodus, is extremely rare in the Middle and South Urals and Kazakhstan. In this situation, correlations might be possible by identifying coeval faunas among other groups of organisms and correlating them and finding traces of transgressive–regressive episodes. Previous studies focussed on the first appearance datum of the conodont S. (Eo). sulcata (see Barskov et al. 1984), which was then considered as a potential boundary marker, but was later devalued due to its facies dependence (absence in shallow-water intervals and extreme rarity in pelagic Stockum Limestone facies) and an earlier occurrence in La Serre (Kaiser 2009). The mostly shallow-water facies of the Berchogur sections contains a diversity of fossils allowing the interpretation of the biota and environments of the South Urals-western Kazakhstan basins at the D–C boundary. The base of the section is represented by shallow-water algal-foraminiferal limestones with stromatoporoids (lower Zhangana Formation, Member 1) overlain by an alternation of shales and marly limestones (Member 2), then by three beds of limestone (Member 3, 45 cm thick in total in Trench BK-3), in the upper part with ammonoids, mostly of the genus Acutimitoceras, and then by shale with rare interbeds of limestone (Member 4). It is not possible to document the occurrence of Pr. kockeli in this section, but both the S. (Eo.) praesulcata and S. (Eo.) sulcata conodont zones are present in the section and it is possible to identify the level of the FO (First Occurrence) of the Acutimitoceras fauna. The undivided Upper expansa to praesulcata zones (= B. ultimus ultimus to S. (Eo.) praesulcata s.l. zones) encompass the lower part of this succession represented by algal-foraminiferal limestone and shale member (Members 1 and 2, with a possible gap between 2a and 2b), while the FO of S. (Eo.) sulcata is recorded in the upper part of Member 3 (near the upper Acutimitoceras level of Barskov et al. 1984; Simakov et al. 1985; although later excavations did not confirm the occurrence of two separate ammonoid levels). Main ammonoid, coral and crinoid occurrences are located at the top of Bed 3c. This level possibly correlates with the sulcata FOs at Seiler, Müssenberg and Hasselbachtal in the Rhenish Massif (Paproth and Streel 1985; Clausen et al. 1989; Bless et al. 1993; Kaiser et al. 2006; Becker et al. 2016; etc.). The correlation of the FO of S. (Eo.) praesulcata with the FO of Pr. kuehni (Upper Protognathodus Fauna of Alberti et al. 1974) was proven in some sections, for example at Seiler (Clausen et al. 1989; compare Kaiser et al. 2019). In the Berchogur Depression, no ammonoids are found within the praesulcata zones (Members 1 and 2, B. ultimus ultimus Zone to ckl), which is characterized by shallowing upwards within Member 2a, a sharp deepening at the base of Member 2b (most likely the transgressive Lower Crisis Interval and Hangenberg Black Shale equivalent). We draw the base of the S. (Eo.) sulcata Zone in Trench BK-1 and Trench BK-3 at the base of Member 3. The presence of carbonates (mudstones) with ammonoids in Bed 3c re-presents the continuation of a transgressive trend after the end of a regressive episode in the upper part of Member 2b (Simakov et al. 1985). An important element in understanding this succession is the occurrence of reworked material forming the gravelstone horizon (between samples 5 and 6) in Member 2a below the limestones forming Unit 2a2. Limestones of Members 3a-b, in addition to conodonts, contain foraminifers (Tournayellina and Paracaligelloides) together with the alga Garwoodia (trenches BK-1 and BK-3). The latter suggests a shallow-water depositional environment. Considering that the Acutimitoceras fauna is found only within Member 3c, it is possible to suggest a quick succession of changes in bathymetry. This agrees with the data from the European successions, where a small-scale regression (Hangenberg Sandstone regression) is presumed near the FAD of S. (Eo.) sulcata. It is presently known that the first appearance of the conodont S. (Eo.) cf. praesulcata is in Member 2b approximately one metre below Member 3, and the interval of ammonoids in Unit 3c shows the presence of S. (Eo.) sulcata in its upper part (Barskov et al. 1984, 1988; Maslov 1987; our own observation). By correlation based on sea-level changes, the FAD of Pr. kockeli could be expected in the uppermost beds of Member 2b (dark shale unit above the limestone with neritic faunas), and we draw the base of the sulcata zone at the base of Member 3 slightly below the FO of S. (Eo.) sulcata. In any case, the upper part of Member 3c can be correlated with the upper part of the prorsum Zone, as S. (Eo.) sulcata enters within the prorsum Zone in Western Europe (Korn and Weyer 2003; Clausen and Korn 2008). It is also possible that beds of Member 4 correlate with higher horizons of the lower Tournaisian, but so far ammonoid data for Member 4 (a series of shale and marly limestones with another level with Acutimitoceras) are inconclusive. This higher level could correspond to the Gattendorfia Zone, but so far, no gattendorfiids were found in this section, possibly due to the very poor exposure of this level. In this respect, it would be important to know the precise level of A. (Acutimitoceras), which is so far only known from Gattendorfia levels of Germany. The correlation of Unit 3c with the upper Stockum Limestone (Tournaisian) is supported by the presence of basal Tournaisian corals and crinoids. It is evident that the succession shows small-scale depositional cycles that can be roughly correlated with those in Western Europe, as episodes dominated by siliciclastic deposition alternated with short events of carbonate accumulation.
Conclusions
The re-excavation and re-examination of the Berchogur sections revealed an interesting history of evolution of a shallow-water mixed terrigenous-carbonate basin showing cycles of limestone and shale deposition around the Devonian–Carboniferous boundary in Western Kazakhstan. The integrated approach allows its correlation with the up-to-date stratigraphic schemes of the Rhenish Massif and other regions and the rough locality of the future D–C boundary in the absence of conodont markers, including Pr. kockeli. This agrees with the currently prevalent approach to search for a composite set of criteria denoting the D–C boundary, rather than a single species that may or may not be present in remote successions. Based on the identification of a regressive event that most probably records the “Hangenberg Sandstone event”, and with the clear distinction of Devonian and Carboniferous faunal associations (foraminifers, conodonts, ammonoids, etc.), the future D–C boundary is likely to be within the upper part of Member 2b, somewhat below the base of Member 3, as this is the end of the regressive episode and beginning of a new transgression. The current D–C boundary is located within Member 3c and is drawn by the FO of S. (Eo.) sulcata. Member 3c contains ammonoids similar to those of the A. prorsum Zone of the Rhenish Stockum Limestone, which ranges from the Pr. kockeli to the basal S. (Eo.) sulcata zones. Crinoids and corals, which are published for the first time from Member 3, show clear lower Tournaisian affinities suggesting that the new Mississippian benthic faunas appeared at Berchogur as early as in the S. (Eo.) sulcata Zone. This pattern agrees with observations in other regions (Denayer and Webb 2017).
Change history
16 June 2022
The missed Rp correctionds were updated
References
Alberti, H., H. Groos-Uffenorde, M. Streel, H. Uffenorde, and O.H. Walliser. 1974. The stratigraphical significance of the Proto-gnathodus fauna from Stockum (Devonian/Carboniferous boundary, Rhenish Schiefergebirge). Newsletters on Stratigraphy 3: 263–276.
Antropov, I.A. 1967. Algae of the Devonian and Lower Carboniferous (Tournaisian) of the central part of the Russian Platform. In Iskopayemye Vodorosli SSSR [Fossil algae of the USSR], ed. A.P. Zhuze, 118–125. Moscow: Nauka (in Russian).
Artyushkova, O.V., V.A. Maslov, V.N. Pazukhin, E.I. Kulagina, R.C. Tagarieva, L.I. Mizens, and A.G. Mizens. 2011. Devonian and Lower Carboniferous type sections of the western South Urals. In Pre-Conference Field Excursion Guidebook. International Conference “Biostratigraphy, Paleogeography and Events in Devonian and Lower Carboniferous”, Ufa, Sterlitamak, Russia, July 20–25, 2011, ed. V.N. Puchkov, 1–92. Ufa: Institute of Geology of the Russian Academy of Sciences.
Austin, T., and T. Austin. 1842. Proposed arrangement of the Echinodermata, particularly as regards of the Crinoidea, and subdivision of the class Adelostella (Echinidae). Annals and Magazine of Natural History, Series 1 10(69): 106–113.
Balashova, E.A. 1945a. On the Fauna of the Carboniferous of Ber-Chogur. Uchenye Zapiski Leningradskogo Gosudarstvennogo Universiteta 65, Seriya Geologo-Pochvennykh Nauk 13: 184–192 (in Russian).
Balashova, E.A. 1945b. On the fauna and stratigraphy of the Carboni-ferous of Ber-Chogur. Nauchnyi Byulleten Leningradskogo Gosudarstvennogo Universiteta 3: 22–28 (in Russian).
Balashova, E.A. 1953. Goniatites of the Carboniferous of Berchogur. Ezhegodnik Vsesoyuznogo Paleontologicheskogo Obshchestva 14: 189–202 (in Russian).
Balashova, E.A. 1956. Some Tournaisian brachiopods of Ber-Chogur (Mugodzhary). Uchenye Zapiski Leningradskogo Gosudarstvennogo Universiteta 209, Seriya Geologo-Pochvennykh Nauk 7: 101–120 (in Russian).
Barskov, I.S., K.V. Simakov, A.S. Alekseev, B.I. Bogoslovsky, T.V. Byvsheva, M.H. Gagiev, L.I. Kononova, N.M. Kochetkova, L.F. Kuzina, and E.A. Reitlinger. 1984. Devonian-Carboniferous transitional deposits of the Berchogur section, Mugodzhary, USSR. Courier Forschungsinstitut Senckenberg 67: 207–230.
Barskov, I.S., and L.F. Kuzina. 1996. The Dinantian ammonoid succession in Northern Eurasia. In European Dinantian Environments II. Developments in Lower Carboniferous Geology, University College Dublin, Ireland, 6th–8th September 1994, Abstracts Volume, ed. G.L. Jones: 44; Dublin: University College Dublin.
Barskov, I.S., L.I. Kononova, T.V. Byvsheva, and A.S. Alekseev. 1988. Granitsa devona i karbona v razreze Berchogur (Mugodzhary) [The Devonian–Carboniferous boundary in the Berchogur section (Mugodzhary)]. In Granitsa devona i karbona na territorii SSSR [The Devonian–Carboniferous boundary in the USSR], ed. V.K. Golubtsov, 171–180. Minsk: Nauka i Tekhnika (in Russian).
Barskov, I.S., T.N. Vorontzova, L.I. Kononova, and A.V. Kuzmin. 1991. Opredelitel konodontov Devona i nizhnego karbona [Key to Devonian and Carboniferous conodonts]. Moscow: Izdatelstvo Moskovskogo Universiteta. (in Russian).
Bartzsch, K., and D. Weyer. 1996. Acutimitoceras acutum (Schindewolf 1923). Leitart der ersten unterkarbonischen Ammonoidea-Zone im Bohlen-Profil von Saalfeld (Thüringisches Schiefergebirge). Beiträge zur Geologie von Thüringen, Neue Folge 3: 91–103.
Becker, R.T. 1996. New faunal records and holostratigraphic correlation of the Hasselbachtal D/C-boundary auxiliary stratotype (Germany). Annales de la Société Géologique de Belgique 117: 19–45.
Becker, R.T., and D. Weyer. 2004. Bartzschiceras n. gen. (Ammonoidea) from the Lower Tournaisian of Southern France. Mitteilungen des Geologisch-Paläontologischen Instituts der Universität Hamburg 88: 11–36.
Becker, R.T., S.I. Kaiser, and M. Aretz. 2016. Review of chrono-, litho- and biostratigraphy across the global Hangenberg Crisis and Devonian-Carboniferous boundary. In Devonian climate, sea level and evolutionary events, eds. R.T. Becker, P. Königshof, and C.E. Brett. Geological Society of London Special Publications 423: 355–386.
Becker, R.T., S. Hartenfels, Ch. Klug, Z.S. Aboussalam, and L. Afhüppe. 2018. The cephalopod-rich Famennian and Tournaisian of the Aguelmous Syncline (southern Maïder). Münstersche Forschungen zur Geologie und Paläontologie 110: 273–306.
Becker, R.T., S. Hartenfels, Z.S. Aboussalam, H. Tragelehn, D. Brice, D., and A. El Hassani. 2013. The Devonian—Carboniferous boundary at Lalla Mimouna (Northern Maider) – a progress report. In International Field Symposium The Devonian and Carboniferous of Northern Gondwana - Morocco 2013, eds. R.T. Becker, A. El Hassani, and A. Tahiri. Documents de l'Institut Scientifique Rabat 27:109−120.
Becker, R.T. 1993. Anoxia, eustatic changes, and Upper Devonian to lowermost Carboniferous global ammonoid diversity. In The Ammonoidea, environment, ecology, and evolutionary change, ed. M.R. House. Systematics Association Special Volume 47: 115–163.
Birina, L.M. 1948. New types of calcareous algae and foraminifera of the boundary layers of the Devonian and Carboniferous. Sovetskaya Geologiya 11(28): 154–159 (in Russian).
Bischoff, G. 1957. Die Conodonten-Stratigraphie des rhenoherzynischen Unterkarbons mit Berücksichtigung der Wocklumeria-Stufe und der Devon/Karbon-Grenze. Abhandlungen des Hessischen Landesamtes für Bodenforschung 19: 1–64.
Bless, M.J.M., R.T. Becker, K. Higgs, E. Paproth, and M. Streel. 1993. Eustatic cycles around the Devonian-Carboniferous boundary and the sedimentary and fossil record in Sauerland (Federal Republic of Germany). Annales de la Société Géologique de Belgique 115: 689–702.
Bogoslovsky, B.I. 1986. Stages in the evolution of ammonoids and the Devonian Carboniferous boundary. In Granitsa devona i karbona na territorii SSSR. Materialy k vsesoyuznomu soveshchaniyu, Minsk, April, 1986 [The Devonian–Carboniferous boundary in the territory of the USSR. Materials for the All-Union Conference], ed. V.I. Avkhimovich, V.K. Golubtsov, Z.A. Gorelik, A.M. Sinichka, and V.A. Chizhova. 24–26. Minsk: Belorusskiy Nauchno-Issledovatelskiy Geologorazvedochnyi Institut. (in Russian).
Bogoslovsky, B.I. 1988. Evolution and biochronology of ammonoids at the Devonian–Carboniferous boundary. In Granitsa devona i karbona na territorii SSSR [The Devonian–Carboniferous boundary in the USSR], ed. V.K. Golubtsov, 248–252. Minsk: Nauka i Tekhnika. (in Russian).
Branson, E.R. 1934. Conodonts from the Hannibal Formation of Missouri. The University of Missouri Studies 8(4): 301–334.
Branson, E.B., and M.G. Mehl. 1934a. Conodonts from the Grassy Creek shale of Missouri. University of Missouri Studies 8(3): 171–259.
Branson, E.B., and M.G. Mehl. 1934b. Conodonts from the Bushberg Sandstone and equivalent formations of Missouri. University of Missouri Studies 8(4): 265–289.
Bushmina, L.S., and L.I. Kononova. 1981. Mikrofauna i biostratigrafiya pogranichnykh sloyev devona i karbona (yug Zapadnoy Sibiri) [Microfauna and biostratigraphy of the boundary beds of the Devonian and Carboniferous (south of Western Siberia)]. In Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniya Akademii Nauk SSSR 459: 1–123. Moscow: Nauka. (in Russian).
Chernova, G.N., and G.A. Stukalina. 1989. On the taxonomy of the Late Paleozoic sea lilies of the family Floricyclidae. In Problemy izuchenia iskopaemykh i sovremennykh iglokozhikh [Problems of studying fossil and modern echinoderms], ed. D.L. Kaljo, 111–127. Tallinn: Valgus. (in Russian).
Clausen, C.-D., K. Leuteritz, W. Ziegler, and D. Korn. 1989. Ausgewählte Profile an der Devon/Karbon-Grenze im Sauerland (Rheinisches Schiefergebirge). Fortschritte in der Geologie von Rheinland und Westfalen 35: 161–226.
Clausen, C.-D., D. Korn, R. Feist, K. Leuschner, H. Groos-Uffenorde, F.W. Luppold, D. Stoppel, K. Higgs, and M. Streel. 1994. Die Devon/Karbon-Grenze bei Stockum (Rheinisches Schiefergebirge). Geologie Und Paläontologie in Westfalen 29: 71–95.
Clausen, C.-D., and D. Korn. 2008. Höheres Mitteldevon und Oberdevon des nördlichen Rheinischen Schiefergebirges (mit Velberter Sattel und Kellerwald). Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften, 52 (Stratigraphie von Deutschland VIII): 439–481.
Conil, R., and M. Lys. 1964. Matériaux pour l’étude micropaléontologique du Dinantien de la Belgique et de la France (Avesnois): Partie 1, algues et foraminifères, et partie 2, foraminifères. Mémoires de l'Institut Géologique de l'Universite de Louvain 23: 1–296.
Corradini, C. 2003. Late Devonian (Famennian) conodonts from the Corona Mizziu sections near Villasalto (Sardinia, Italy). Palaeontographia Italica 98: 65–116.
Corradini, C., S.I. Kaiser, M.C. Perri, and C. Spalletta. 2011. Proto-gnathodus (Conodonta) and its potential as a tool for defining the Devonian/Carboniferous boundary. Rivista Italiana di Paleontologia e Stratigrafia 117(1): 15–28.
Corradini, C., C. Spalletta, A. Mossoni, H. Matyja, and D.J. Over. 2017. Conodont across the Devonian/Carboniferous boundary: A review and implication for the redefinition of the boundary and a proposal for an updated conodont zonation. Geological Magazine 154(4): 888–902.
De Koninck, L.-G. 1872. Nouvelles recherches sur les animaux fossiles du terrain carbonifère de la Belgique. Mémoires de l'Académie Royale de Belgique 39: 1–78.
Dehée, R. 1929. Description de la faune d’Etrœungt—Faune de passage du Dévonien au Carbonifère. Mémoires de la Société Géologique de France, Nouvelle Série 5: 1–62.
Denayer, J., and G. Webb. 2017. Earliest Mississippian rugose corals of eastern Australia: Post-disaster fauna across the Devonian-Carboniferous Boundary? Palaeobiodiversity and Palaeoenvironments 97(3): 367–374.
Druce, E.C. 1969. Devonian and Carboniferous conodonts from the Bonaparte Gulf Basin, northern Australia. Bureau of Mineral Resources Bulletin 98: 1–242.
Dubatolova, Yu.A. 1976. Opisaniye fauny: typ Echinodermata. [Descriptions of fauna: type Echinodermata] in Pribalkhashje—perekhodnaya zona biogeographicheskikh poyasov pozdnego karbona [Balkhash area—a transitional zone of Late Carboniferous biogeographical belts], ed. V.N. Dubatolov. Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniya Akademii Nauk SSSR 285: 81–98 (in Russian).
Dubatolova, Yu.A., and Shao Jie. 1959. Stems of sea lilies of the southern China Carboniferous, Permian and Triassic deposits. Acta Palaeontologica Sinica 7(1): 41–76 ((in Russian and Chinese)).
Durkina, A.V. 1959. Foraminifera of the Lower Carboniferous Deposits of the Timan-Pechora Province. Trudy VINGRI, Novaya Seriya 136 (Mikrofauna SSSR): 132–335. (in Russian).
Dzik, J. 2006. The Famennian “Golden Age” of conodonts and ammonoids in the Polish part of the Variscan Sea. Palaeontologica Polonica 63: 1–360.
Fedorowski, J. 1971. Aulophyllidae (Tetracoralla) from the Upper Viséan of Sudetes and Holy Cross Mountains. Palaeontologica Polonica 24: 1–137.
Flajs, G., and R. Feist. 1988. Index conodonts, trilobites and environment of the Devonian-Carboniferous Boundary beds at La Serre (Montagne Noire, France). Courier Forschungsinstitut Senckenberg 100: 53–107.
Frech, F. 1885. Die Korallenfauna der Oberdevons in Deutschland. Zeitschrift der Deutschen Geologischen Gesellschaft 37: 21–130.
Gierlowski-Kordesch, E.H., and C.F. Cassle. 2015. The “Spirorbis” problem revisited: Sedimentology and biology of microconchids in marine-nonmarine transitions. Earth-Sciences Reviews 148: 209–227.
Głuchowski, E. 1981. Paleozoic crinoid columnals and pluricolumnals from Poland. Geologia 7(3): 29–57.
Głuchowski, E. 2002. Crinoids from the Famennian of the Holy Cross Mountains, Poland. Acta Palaeontologica Polonica 47: 319–328.
Głuchowski, E., and G. Racki. 2005. Disarticulated crinoid stems from the Devonian and Carboniferous of North Devon, England. Proceedings of the Yorkshire Geological Society 55(3): 161–172.
Goldfuss, G.A. 1826. Petrefacta Germaniae. 1‒122. Düsseldorf: Anz and Co.
Hahn, G., and R. Hahn. 1972. Trilobitae carbonici et permici III. Fossilium Catalogus 120: 336–531.
Hahn, G., and R. Hahn. 2008. Catalogus trilobitorum cum figures (Trilobitae carbonici et permici, VI. Cummingellinae). Fossilium Catalogus 145: 1‒432.
Hance, L., H. Hou, D. Vachard, F.X. Devuyst, J. Kalvoda, E. Poty, and X. Wu. 2011. Upper Famennian to Viséan foraminifers and some carbonate Microproblematica from South China – Hunan, Guangxi and Guizhou, eds. L. Hance, H. Hou, and D. Vachard. 1‒359. Beijing: Geological Publishing House.
House, M.R. 1993. Earliest Carboniferous goniatite recovery after the Hangenberg Event. Annales de la Société Géologique de Belgique 115(2): 559–579.
Huddle, J.W. 1934. Conodonts from the New Albany Shale of Indiana. Bulletins of American Paleontology 21(72): 1–136.
Hudson, R.G.S. 1943. On the lower Carboniferous coral Permia cavernula sp. n. Annals and Magazine of Natural History Series 11 10(66): 361–368.
Hudson, R.G.S., and M.I. Platt. 1927. On the Lower Carboniferous corals: The development of Rylstonia benecompacta, gen. et. sp. n. Annals and Magazine of Natural History Series 9 19(109): 39–48.
Hudson, R.G.S. 1942. On the Lower Carboniferous corals: Rylstonia benecompacta var. breviseptata, var. nov. Proceedings of the Yorkshire Geological Society 24(5): 373–382.
Ivanov, K.S., V.A. Kontorovich, V.N. Puchkov, Yu.N. Fedorov, and Yu.V. Erokhin. 2014. Tectonics of the Urals and the basement of Western Siberia: The main features of the geological structure and development. Geologiya i Mineralno-Syryevyye Resursy Sibiri 2C: 22–35 (in Russian).
Ivanov, S.N., V.N. Puchkov, K.S. Ivanov, G.I. Samarkin, I.V. Semenov, A.I. Pumpyansky, A.M. Dymkin, Yu.A. Poltavets, A.I. Rusin, and A.A. Krasnobaev. 1986. Formirovanie zemnoy kory Urala [Ural crust formation]. 1–248. Moscow: Nauka. (in Russian).
Ivanova, R.M., and T.I. Stepanova. 2011. Upper Devonian calcareous algae of the southern West Siberia (Quasiendothyra kobeitusana Zone). Litosfera 3: 22–38 (in Russian).
Jaworski, V.I. 1916. Several words on the Ber-Chogur coal deposit in Mugodzhary. Poverkhnost’ i Nedra 6: 241–245 (in Russian).
Kaiser, S.I. 2009. The Devonian/Carboniferous stratotype section La Serre (Montagne Noire) revisited. Newsletters on Stratigraphy 43: 195–205.
Kaiser, S.I., T. Steuber, R.T. Becker, and M. Joachimski. 2006. Geochemical evidence for major environmental change at the Devonian-Carboniferous boundary in the Carnic Alps and the Rhenish Massif. Palaeogeography, Palaeoclimatology, Palaeoecology 240(1–2): 146–160.
Kaiser, S.I., R.T. Becker, C. Spalletta, and T. Steuber. 2009. High-resolution conodont stratigraphy, biofacies and extinctions around the Hangenberg Event in pelagic successions from Austria, Italy and France. Palaeontographica Americana 63: 97–139.
Kaiser, S.I., M. Aretz, and R.T. Becker. 2016. The global Hangenberg Crisis (Devonian–Carboniferous transition): Review of a first-order mass extinction. In Devonian climate, sea level and evolutionary events, eds. R.T. Becker, P. Königshof, and C.E. Brett. Geological Society of London Special Publications 423: 387–437.
Kaiser, S., T. Kumpan, and V. Cígler. 2017. New unornamented siphonodellids (Conodonta) of the lower Tournaisian from the Rhenish Massif and Moravian Karst (Germany and Czech Republic). Neues Jahrbuch für Geologie und Paläontologie 286(1): 1–33.
Kaiser, S., T. Kumpan, and M.W. Rasser. 2019. High resolution conodont biostratigraphy in two key sections from the Carnic Alps (Grüne Schneid) and Graz Paleozoic (Trolp)—implication for biozonation concept at the Devonian-Carboniferous boundary. Newsletters on Stratigraphy 53(3): 249–274.
Kalvoda, J., T. Kumpan, and O. Bábek. 2015. Upper Famennian and Lower Tournaisian sections of the Moravian Karst (Moravo-Silesian Zone, Czech Republic): a proposed key area for correlation of the conodont and foraminiferal zonations. Geological Journal 50: 17–38.
Keller, N.B. 1959. New lower Carboniferous tetracorals of Dzhezkazgan Area (Kazakhstan). Paleontologicheskii Zhurnal 1959 (4): 90–99 (in Russian).
Klapper, G. 1958. An upper Devonian conodont fauna from the Darby formation of the Wind River mountains, Wyoming. Journal of Paleontology 32(6): 1082–1093.
Korn, D. 1986. Ammonoid evolution in Late Famennian and Early Tournaisian. Annales de la Société Géologique de Belgique 109: 49–54.
Korn, D. 1994. Devonische und karbonische Prionoceraten (Cephalopoda, Ammonoidea) aus dem Rheinischen Schiefergebirge. Geologie und Paläontologie in Westfalen 30: 1–85.
Korn, D. 2002. Historical subdivisions of the Middle and Late Devonian sedimentary rocks in the Rhenish Mountains by ammonoid faunas. Senckenbergiana Lethaea 82(2): 545–555.
Korn, D., and Ch. Klug. 2015. Paleozoic ammonoid biostratigraphy. In Ammonoid paleobiology, from macroevolution to paleogeography, ed. Ch. Klug, D. Korn, K. De Baets, I. Kruta, and R.H. Mapes, 299–328. Dordrecht: Springer.
Korn, D., and D. Weyer. 2003. High resolution stratigraphy of the Devonian-Carboniferous transitional beds in the Rhenish Mountains. Mitteilungen aus dem museum für Naturkunde in Berlin, Geowissenschaftliche Reihe 6: 79–124.
Kulagina, E.I. 2013. Taxonomic diversity of foraminifers of the Devonian-Carboniferous boundary interval in the South Urals. Bulletin of Geosciences 88(2): 265–282.
Kulagina, E.I., Y.A. Vevel, T.I. Stepanova, and E.L. Zaytseva. 2016. Foraminifers of the genus Tournayellina Lipina, 1955. Paleontological Journal 50: 557–572.
Kulagina, E.I., E.I. Zaytseva, Y.A. Vevel, T.I. Stepanova, N.B. Gibshman, S.V. Nikolaeva, L.I. Kononova, and A.N. Plotitsyn. 2021. The foraminiferal zonal scale of the Devonian-Carboniferous boundary beds in Russia and western Kazakhstan and its correlation with ammonoid and conodont scales. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-020-00439-y.
Kulagina, E.I., T.I. Stepanova, E.I. Zaytseva, E.N. Gorozhanina, N.B. Gibshman, R.M. Ivanova, Ya.A. Vevel, G.Yu. Ponomareva, and T.V. Filimonova. 2018. Atlas of Upper Devonian and Lower Carboniferous foraminifers and microfacies of northern Eurasia. Famennian and Tournaisian. 1–221. Moscow: Paleontological Institute, Russian Academy Sciences, pp. http://ig.ufaras.ru/File/mono/Atlas_2018.pdf (in Russian).
Kulagina, E.I. 1982. Depositional settings of the Zhangana Formation of the Berchogur Depression (Mugodzhary). In Verkhniy dokembriy i paleozoy Yuzhnogo Urala: (Stratigrafiya i litologiya) [Upper Precambrian and Paleozoic of the South Urals: (Stratigraphy and lithology)], ed. M.A. Garris, N.M. Kochetkova, et al. 116–122. Ufa: BFAN SSSR. (in Russian).
Kurilenko, A.V., G.V. Kotlyar, N.P. Kulkov, N.I. Raitina, N.G. Yadrischenskaya, L.P. Starukhina, Ye.M. Markovich, T.M. Okuneva, T.A. Dol'nik, L.I. Popeko, G.V. Belyayeva, A.S. Byakov, N.F. Bashurova, A.V. Timokhin, I.V. Korovnikov, N.K. Mogucheva, N.G. Izokh, S.A. Anisimova, T.V. Klets, R.M. Ivanova, and G.A. Stukalina. 2002. Atlas fauny i flory paleozoya-mezozoya Zabaykal'ya [Atlas of fauna and flora of Paleozoic-Mesozoic in Transbaikal]. Novosibirsk: Nauka (in Russian).
Kurilenko, A.V., and N.P. Kulkov. 2008. A proposed crinoid zonation of the Devonian deposits of Eastern Transbaikal. Bulletin of Geosciences 83(4): 461–472.
Kurilenko, A.V. 1989. Finds of Platycrinites in the Lower Carboniferous of Eastern Transbaikal. In Problemy izuchenia iskopaemykh i sovremennykh iglokozhikh [Problems of studying fossil and modern echinoderms], ed. D.L. Kaljo. 128–133. Tallinn: Valgus. (In Russian).
Kusina, L.F. 1985. On the revision of the genus Imitoceras. Paleontologicheskiy Zhurnal 1985(3): 35–48 (in Russian).
Lebedeva, N.S. 1956. Foraminifera of the Etroeungt deposits of the Tengiz Basin. Trudy Vsesoyuznogo Neftyanogo Nauchno-Issledovatelskogo Geologo-Razvedochnogo Instituta (VNIGRI). Novaya Seriya 98: 39–60 (in Russian).
Librovitch, L.S. 1957. On some new groups of goniatites from the Carboniferous deposits of the USSR. Ezhegodnik Vsesoyuznogo Paleontologicheskogo Obshchestva 16: 246–272 (in Russian).
Librovitch, L.S. 1940. Ammonoidea from the Carboniferous beds of Northern Kazakhstan. In Paleontologiya SSSR, 4 (Part 9, no. 1), 1–394. Moscow-Leningrad: Izdatelstvo Akademii Nauk SSSR. (in Russian).
Lipina, O.A. 1948. Foraminifers of the Chernyshino Formation of the Tournaisian Stage of the Lower Carboniferous in the Moscow Basin. Trudy Instituta Geologicheskikh Nauk Akademii Nauk SSSR, Geologicheskaya Seriya 19(62): 251–259 (in Russian).
Lipina, O.A. 1955. Foraminifers from the Tournaisian Stage and the upper part of the Devonian of the Volga-Ural region and the western slope of the Middle Urals. Trudy Instituta Geologicheskikh Nauk Akademii Nauk SSSR 163: 1–96 (in Russian).
Lipina, O.A. 1965. Systematics of tournayellids. Trudy Instituta Geologicheskikh, Nauk Akademii Nauk SSSR 139: 1–116 (in Russian).
Malakhova, N.P. 1954. Foraminifers from the Kizelovian Limestone of the western slopes of the Urals. Byulleten Moskovskovo Obshchesta Ispytatelei Prirody, N.s., Otdel Geologicheskii 29(1): 49–60 (in Russian).
Mamet, B. 1991. Carboniferous calcareous algae. In Calcareous algae and stromatolites, ed. R. Riding, 370–451. Berlin: Springer.
Maslov, V.A. Ed. 1987. Fauna i Biostratigrafiya pogranichnykh otlozhenii devona i karbona Berchogura (Mugodzhary) [Fauna and biostratigraphy of the Devonian–Carboniferous boundary beds in Berchogur (Mugodzhary)]. Moscow: Nauka (In Russian).
Michelin, J.L.H. 1840–1847. Iconographie Zoophytologique. xii + 348. pp. 79 pl. Paris: P. Bertrand.
Miller, J.S. 1821. A Natural History of the Crinoidea, Or Lily-shaped Animals: With Observations on the Genera, Asteria, Euryale, Comatula & Marsupites. Bristol: Bryan & Co.
Milne-Edwards, H. and Haime, J. 1850–1854. A monograph of Britisch fossil corals. Palaeontographical Society London: 3(1850): i-lxxxv, 1–71; 5(1851): 73–145; 6(1852): 147–210; 7(1853): 211–244; 8(1854): 245–322.
Moore, R.C., and R.M. Jeffords. 1968. Classification and nomenclature of fossil crinoids based on studies of dissociated parts of their columns. University of Kansas Paleontological Contributions, Supplement of 46 (Echinodermata: 9):1–86 .
Münster, G.G. 1839. Nachtrag zu den Goniatiten des Fichtelgebirges. Beiträge Zur Petrefactenkunde 1: 16–31.
Münster, G.G. 1840. Die Versteinerungen des Uebergangskalkes mit Clymenien und Orthoceratiten von Oberfranken. Beiträge zur Petrefacten-Kunde 3: 33–121.
Nalivkin, D.N. 1937. Brachiopods of the Upper and Middle Devonian and Lower Carboniferous of Northeastern Kazakhstan. Trudy Tsentralnogo Nauchno-Issledovatelskogo Geologorazvedochnogo Instituta 99: 1–200 (in Russian).
Nicholson, H.A. 1888. On certain anomalous organisms which are concerned in the formation of some of the Palaeozoic limestones. Geological Magazine 5:15.
Nikolaeva, S.V. 2020. New ammonoids from the Devonian-Carboniferous boundary section in Berchogur (western Kazakhstan). Paleontological Journal 54(5): 464–476.
Nikolaeva, S.V., and S.N. Mustapaeva. 2018. The Devonian-Carboniferous boundary locality in Berchogur (western Kazakhstan) revisited. Subcommission on Devonian Stratigraphy, Newsletter 33: 31–34.
Onoprienko, Y. 1979. New Rugosa findings in the Devonian–Carboniferous transitional deposits of the Omolon Massif. Field excursion guidebook tour IX, XIV, Pacific Science Congress, Khabarovsk, 1979, supplement 2: 5–13. Magadan: Magadan Publishing House (in Russian).
Osmólska, H. 1970. Revision of non-cyrtosymbolinid trilobites from the Tournaisian-Namurian of Eurasia. Palaeontologica Polonica 23: 1–165.
Paproth, E., and M. Streel. 1985. In search of a Devonian-Carboniferous boundary. Episodes 8(2): 110–111.
Paproth, E., R. Feist, and G. Flajs. 1991. Decision on the Devonian-Carboniferous boundary stratotype. Episodes 14(4): 110–336.
Petrenko, A.A. 1946. Lower Carboniferous coal and anthracite deposits of the South Urals. Trudy Instituta Geologicheskikh Nauk 79: 1–176.
Petrenko, A.A., and A.A. Sultanaev. 1977. Karabulak Formation. In Stratigraficheskii slovar SSSR. Karbon, Perm [Stratigraphic dictionary of the USSR. Carboniferous, Permian], ed. G.V. Kotlyar, 173. Leningrad: Nedra. (in Russian).
Petrenko, A.A. 1977. Berchogur Formation. In Stratigraficheskii slovar SSSR. Karbon, Perm [Stratigraphic dictionary of the USSR. Carboniferous, Permian], ed. G.V. Kotlyar. 65. Leningrad: Nedra. (in Russian).
Poty, E. 1999. Famennian and Tournaisian recoveries of shallow water Rugosa following late Frasnian and late Strunian major crises, southern Belgium and surrounding areas, Hunan (South China) and the Omolon region (NE Siberia). Palaeogeography, Palaeoclimatology, Palaeoecology 154(1–2): 11–26.
Poty, E. 2016. The Dinantian (Mississippian) succession of southern Belgium and surrounding areas: Stratigraphy improvement and inferred climate reconstruction. Geologica Belgica 19(1–2): 177–200.
Poty, E., and K. Boland. 1994. Révision des tétracoralliaires caninomorphes de l’Hastarien (Tournaisien) belge. Annales de la Société Géologique de Belgique 117(1): 201–225.
Poty, E., F.X. Devuyst, and L. Hance. 2006. Upper Devonian and Mississippian foraminiferal and rugose coral zonations of Belgium and northern France: A tool for Eurasian correlations. Geological Magazine 143(6): 829–857.
Puchkov, V.N. 2000. Paleogeodinamika Tsentralnogo i Yuzhnogo Urala [Paleogeodynamics of the Central and South Urals]. Ufa: Dauria. (in Russian).
Puchkov, V.N. 2010. Geologiya Urala i Priural'ya (aktual'nyye voprosy stratigrafii, tektoniki, geodinamiki i metallogenii) [Geology of the Urals and Cisuralia, topical issues of stratigraphy, tectonics, geodynamics and metallogeny]. Ufa: DizainPoligrafServis. (in Russian).
Qie, W., J. Liu, J. Chen, X. Wang, H. Mii, X.X. Zhang, X. Huang, L. Yao, and T.J.G. Algeo, and G. Luo. 2015. Local overprints on the global carbonate δ13C signal in Devonian-Carboniferous boundary successions of South China. Palaeogeography, Palaeoclimatology, Palaeoecology 418: 290–303.
Rauser-Chernousova, D.M. 1948. Contributions to the foraminiferal fauna of the Carboniferous deposits of Central Kazakhstan. Trudy Instituta Geologicheskikh Nauk, Akademii Nauk SSSR, Geologicheskaya Seriya 21(66): 1–28 (in Russian).
Reitlinger, E.A. 1961. Some questions of the systematics of Quasiendothyra. Voprosy Mikropaleontologii 5: 31–68 (in Russian).
Reitlinger, E.A., and E.I. Kulagina. 1987. Foraminifers. In Fauna i Biostratigrafiya pogranichnykh otlozhenii devona i karbona Berchogura (Mugodzhary) [Fauna and biostratigraphy of the Devonian–Carboniferous boundary beds in Berchogur (Mugodzhary)], ed. V.A. Maslov, 48–52. Moscow: Nauka. (in Russian).
Richter, R., and E.E. Richter. 1951. Der Beginn des Karbons im Wechsel der Trilobiten. Senckenbergiana 32(1/4): 219–266.
Rozman, Kh.S. 1960. Stratigraphy of the Famennian and Lower Tournaisian deposits of Mugodzhary and adjacent regions of the Urals. Izvestiya Akademii Nauk SSSR, Seriya Geologicheskaya 12: 42–51 (in Russian).
Rozman, Kh.S. 1962. Stratigraphy and brachiopods of the Famennian Stage of Mugodzhary and adjacent regions. Trudy Geologicheskogo Instituta Akademii Nauk SSSR 50: 1–196 (in Russian).
Ruan, Y.-P. 1981. Devonian and earliest Carboniferous ammonoids from Guangxi and Guizhou. Memoirs of the Nanjing Institute of Geology and Palaeontology, Academia Sinica 15: 1–152.
Ruzhencev, V.E., and M.F. Bogoslovskaya. 1971. Namurian stage in evolution of ammonoids. Early Namurian ammonoids. Trudy Paleontologicheskogo Instituta, Akademiya Nauk SSSR 133: 1–382 (in Russian).
Sandberg, C.A., and R. Dreesen. 1984. Late Devonian icriodontid biofacies models and alternate shallow-water conodont zonation. Geological Society of America, Special Paper 196: 143–178.
Sandberg, C.A., M. Streel, and R.A. Scott. 1972. Comparison between conodont zonation and spore assemblages in the Devonian-Carboniferous boundary in the western and central United States and in Europe. Compte Rendu Septième Congrès International de Stratigraphie et de géologie du Carbonifère, Krefeld 1971(1): 179–203.
Schindewolf, O.H. 1923. Beiträge zur Kenntnis des Paläozoikums in Oberfranken, Ostthüringen und dem Sächsischen Vogtlande. I. Stratigraphie und Ammoneenfauna des Oberdevons von Hof a.S. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Beilage-Band 49: 250–357.
Schmidt, H. 1924. Zwei Cephalopodenfaunen an der Devon-Carbon-Grenze im Sauerland. Jahrbuch der Preußischen Geologischen Landesanstalt 44(1923): 98–171.
Scott, A.J. 1961. Three new conodonts from the Louisiana Limestone (Upper Devonian) of western Illinois. Journal of Paleontology 35: 1223–1227.
Semenov, P., and V.I. Moeller. 1864. On the Upper Devonian strata of Middle Russia. Gornyi Zhurnal 2: 187–233 (in Russian).
Shilo, N.A., J. Bouckaert, G.A. Afanasjeva, M.J. Bless, R. Conil, O.A. Erlanger, M.H. Gagiev, S.S. Lazarev, Y.I. Onoprienko, E. Poty, T.P. Razina, K.V. Simakov, L.V. Smirnova, M.M. Streel, and T.P. Razina. 1984. Sedimentological and paleontological atlas of the late Famennian and Tournaisian deposits in the Omolon region (NE-USSR). Annales de la Société Géologique de Belgique 107: 137–247.
Simakov, K.V., A.S. Alekseev, I.S. Barskov, B.I. Bogoslovsky, T.V. Byvsheva, M.H. Gagiev, L.I. Kononova, L.I. Kochetkova, L.F. Kuzina, E.I. Kulagina, Yu.I. Onoprienko, V.N. Pazukhin, E.R. Radionova, T.P. Razina, E.A. Reutlinger, L.V. Smirnova, and M.G. Yanbulatova. 1985. Biostratigrafiya pogranichnykh otlozhenii devona i karbona. Vypusk 9. Pogranichnye otlozheniya devona i karbona Mugodzhar [Biostratigraphy of the Devonian–Carboniferous boundary beds. Issue 9. Devonian–Carboniferous boundary beds in Mugodzhary], preprint. 1–55. Magadan: SVKNII, Dalnevostochnyi Nauchnyi Tsentr. (in Russian).
Sisova, E.N. 1979. Late Devonian floricyclides of Eastern Kazakhstan. Ezhegodnik Vsesoyuznogo Paleontologicheskogo Obshchestva 22: 166–171 (in Russian).
Sisova, E.N. 1983. About some Devonian and Carboniferous floricyclides in Central and Southern Kazakhstan (Crinoids). Ezhegodnik Vsesoyuznogo Paleontologicheskogo Obshchestva 25: 263–275 (in Russian).
Sisova, E.N. 1988. Late Famennian crinoids of Central Kazakhstan. Ezhegodnik Vsesoyuznogo Paleontologicheskogo Obshchestva 31: 146–158 (in Russian).
Söte, T., S. Hartenfels, and R.T. Becker. 2017. Uppermost Famennian stratigraphy and facies development of the Reigern Quarry near Hachen (northern Rhenish Massif, Germany). Palaeobiodiversity and Palaeoenvironments 97: 633–654.
Spalletta, C., M.C. Perri, D.J. Over, and C. Corradini. 2017. Famennian (Upper Devonian) conodont zonation: Revised global standard. Bulletin of Geosciences 92(1): 31–57.
Stukalina, G.A. 1973. Late Paleozoic sea lilies of Transbaikal and Mongolia. In Stratigrafiya i paleontologiya osadochnykh geologicheskikh formatsiy Zabaikal'ya [Stratigraphy and paleontology of sedimentary geological formations of Transbaikal], ed. S.M. Sinitsa, 16–55; Chita: Vostochno-Sibirskoe knizhnoe Izdatelstvo. (in Russian)
Suleimanov, I.S. 1945. New species of small foraminifera from the Tournaisian Stage of the Ishimbayevo District. Doklady Akademii Nauk SSSR 48(2): 130–134 (in Russian).
Sun, Y.-Z., and Y.-T. Shen. 1965. On the late Upper Devonian ammonite fauna of the Wocklumeria beds of south Kweichow and its stratigraphical significance. Professional Papers of the Academy of Geological Science, Ministry of Geology, Section B 1: 33–100 (in Chinese).
Thomas, L.A. 1949. Devonian-Mississippian formations of southeast Iowa. Geological Society of America Bulletin 60 (3): 403–438.
Ulitina, L.M. 1975. Rugose corals. In Fauna pogranichnykh otlozheniy devona i karbona Tsentralnogo Kazakhstana [Fauna of the Devonian–Carboniferous boundary beds of Central Kazakhstan], ed. T.A. Gorokhova. Materialy po geologii Centralnogo Kazakhstana, vol. 18, pp. 36–41 (in Russian).
Vaughan, A. 1908. An account of the faunal succession and correlation. Pp. 436–474 in: C.A. Matley and A. Vaughan. The Carboniferous rocks at Loughshinny (County Dublin). Quarterly Journal of Geological Society of London 64: 413–474.
Vevel, Ya.A., A.V. Zhuravlev, and V.V. Popov. 2012. Devonian–Carboniferous boundary beds in the section on the Kamenka River (Pechoro-Kozhvinsky Mega-Swell, Timan-Pechora Province). Neftegazovaya geologiya. Teoriya i praktika 7:1 [Online publication] http://www.ngtp.ru/rub/2/6_2012.pdf (in Russian).
Vevel, Ya.A. 2009. A discovery of the foraminifera Tournayellina beata pseudobeata Reitlinger and Kulagina in the Lower Carboniferous of the Pechora Ridge. Neftegazovaya Geologiya. Teoriya i praktika 4: 2 [Online publication] http://www.ngtp.ru/rub/2/17_2009.pdf (in Russian).
Vöhringer, E. 1960. Die Goniatiten der unterkarbonischen Gattendorfia-Stufe im Hönnetal (Sauerland). Fortschritte in Der Geologie Von Rheinland Und Westfalen 3 (1): 107–196.
Volkova, M.S. 1941. Nizhnekamennougol’nye korally tsentral’nogo Kazakhstana (Lower Carboniferous corals of Central Kazakhstan). Materialy Po Geologii i Poleznym Iskopayemym Kazakhstana 11: 1–120 (in Russian and English).
von Peetz, H. 1893. Materials for the study of the fauna of the Malevko-Muraevinsk Stage. Travaux De La Société Des Naturalistes De St.-Pétersbourg, Section De Géologie Et De Mineralogie 22(2): 29–118 (in Russian).
Vorozhbitov, A.M., and A.S. Alekseev. 1992. New Conularia from the Tournaisian of Berchogur (Mugodzhary). Paleontologicheskiy Zhurnal 1993 (3): 94–96 (in Russian).
Weber, V.N. 1932. Trilobity Turkestana (Trilobites of Turkestan). Trudy Vsesoyuznogo Geologo-Razvedochnogo Objedineniya. 1–158 pp. Moskva-Leningrad. (in Russian).
Weber, V.N. 1937. Trilobity kamennougolnych i permskich otlozenij SSSR. Kamennougolnye trilobity (Trilobites of the Carboniferous and Permian Deposits of the USSR. Carboniferous trilobites). Monografii po paleontologii SSSR 71(1): 1–160; Leningrad-Moskva: Glavnaya redaktsiya geologo-razvedochnoy i geodezicheskoy literatury (in Russian).
Whyte, M.A. 1976. A Carboniferous pedunculate barnacle. Proceedings of the Yorkshire Geological Society 41: 1–12.
Woodward, H. 1884. A Monograph of British Carboniferous trilobites. Monographs of the Palaeontographical Society. Part II: 39–86.
Yeltyschewa, R.S. and Poljarnaja, Z.A. 1975. Crinoidea (sea lilies). Trudy Vsesoyuznogo Neftyanogo Geologo-Razvedochnogo Instituta (VNIGRI) 383 (Paleontological Atlas of Carboniferous deposits of Urals): 205–207 (in Russian).
Ziegler, W. 1962. Taxionomie und Phylogenie oberdevonischer Conodonten und ihre stratigraphische Bedeutung. Abhandlungen des Hessischen Landesamtes für Bodenforschung 38: 1–166.
Ziegler, W., and C.A. Sandberg. 1984. Palmatolepis-based revision of upper part of standard Late Devonian conodont zonation. Geological Society of America, Special Paper 196: 179–194.
Ziegler, W., and C.A. Sandberg. 1990. The late Devonian standard conodont zonation. Courier Forschungsinstitut Senckenberg 121: 1–115.
Zonenshain, L.P., M.I. Kuzmin, and L.M. Natapov. 1990. Tektonika litosfernykh plit territorii SSSR [Tectonics of lithospheric plates of the USSR], vols. 1, 2. 1‒328, 1‒334. Moscow: Nedra. (in Russian).
Acknowledgements
We are grateful to the colleagues from the Alabas Quarry who helped us to excavate the trenches and to our colleagues from Satbayev University (Kazakh National Research Technical University) who participated in the field trip. We thank S. Bagirov (Paleontological Institute, Moscow) for trilobite photography, the Photography Unit (NHM, London) for ammonoid photography and N.G. Yadrishchenskaya (AO “Urangeo”, Chita, Russia) for crinoid photography. We also thank Max Barclay (NHM, London) for checking the English of the manuscript. We are deeply grateful to H.-G. Herbig (Cologne) and R.T. Becker (Münster) for their thorough reviews. The fieldwork was supported by a grant from the Satbayev University (Kazakh National Research Technical University) to S.N. Mustapayeva and S.V. Nikolaeva (no. AP05131610 “Major chronostratigraphic levels in the lower Carboniferous in the sections of Western Kazakhstan (Aktobe Region): potential for international standards”. The work by E. Kulagina is supported by State Programs 0246-2019-0118, IG UFRC R. This study was partly funded by a subsidy of the Russian Government to support the Program of Competitive Growth of Kazan Federal University among the World’s Leading Academic Centres.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Hans-Georg Herbig.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nikolaeva, S.V., Kulagina, E.I., Mustapayeva, S.N. et al. Progress in the study of the Devonian–Carboniferous boundary sections in the Berchogur Depression (Mugodzhary Mountains, western Kazakhstan). PalZ 96, 415–447 (2022). https://doi.org/10.1007/s12542-022-00611-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-022-00611-z