
Abstract
Balma Guilanyà shelter (north-eastern Iberian Peninsula, Spain) contains a stratigraphic sequence spanning the Late Upper Palaeolithic to early Holocene. During excavations from 1992 to 2008, seventeen human teeth, and several cranial and postcranial bones were unearthed from the layer E (Late Glacial). In this contribution, we report new unpublished dental remains, which add to the previously dental assemblage from level E at Balma Guilanyà. The purpose of this article is to provide an updated description of the morphology of this material, as well as an analysis of the MNI, age at death, and the presence of several pathologies, approaching it from a holistic vision of the teeth assemblage from the Late Glacial. This review has made it possible to diagnose a dens invaginatus, the persistence of a deciduous upper central incisor (possibly caused by a mesiodens), teeth with cavities, deep occlusal pits conducive to cariogenesis, an atypical lower second molar and several molars with fused roots. This pattern of dental anomalies attributed to a single individual (Individual III) is not usual among hunter-gatherers who lived 13,000 years ago. Likewise, the presence of enamel hypoplasia in several teeth allows us to discuss this aetiology. These new identifiers derived from the review of the dental assemblage from Balma Guilanyà can help extend our knowledge of dental pathologies and stress markers in Late Glacial Homo sapiens populations in Western Europe. Additionally, an accumulation of teeth attributed to two individuals found in a confined area of the excavation was examined. Potential scenarios were assessed regarding the arrangement of this grouping, which presents challenges in attributing its formation to post-depositional or natural processes. This pattern would have the potential to elucidate behavioural funerary practices among late glacial hunter-gatherer populations.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The remains of Late Pleistocene Homo sapiens are crucial to anthropologically, demographically and genetically reconstructing Western European hunter-gatherers (Posth et al. 2023; Villalba-Mouco et al. 2019; Villalba-Mouco et al. 2023). These proxies originate from skeletal remains found in intentional burials, often containing single individuals, although instances of double or multiple burials exist. These burial contexts, which offer crucial palaeoanthropological and cultural insights, are relatively uncommon (Pettitt 2011; Orschiedt 2018). Conversely, the discovery of isolated human remains, typically challenging to correlate with funerary practices, is a more common phenomenon (Arenas del Amo et al. 2024). There is growing interest in this type of remains since it cannot be ruled out that they correspond to crypto-funerary behaviours that are difficult to explain (Kerner 2018). However, taphonomic, contextual and cultural processes mask the diversity of factors at work on the archaeological record, making it difficult to identify the peri/post-mortem treatment of these individuals (Fahlander and Oestigaard 2008; Orschiedt 2013).
The interpretation of human remains retrieved from ancient excavations or poorly documented contexts poses challenges due to potential ambiguities. However, while employing direct 14C dating could effectively resolve uncertainties regarding their age, these analyses are infrequently conducted. This circumstance may impact the comprehension of human remains discovered at Balma Guilanyà. While their antiquity remains undisputed, elucidating their cultural significance has proven elusive (Meiklejohn 2009; Peyroteo 2016). The assemblage was recovered from an archaeological level that several 14C (AMS) dates, including two direct dates on human remains, confirming them as being Late Glacial. However, the causes related to the formation of this accumulation may be the result of natural processes, or alternatively, they could be associated with mortuary practices (García-Guixé et al. 2009; Martínez-Moreno and Mora 2009).
This article reviews the contextual and chronometric data justifying the attribution of the human remains to the Late Glacial. Our review focuses on the dental assemblage to evaluate the count of individuals, revising previous estimates (Lalueza 1996; Ruíz et al. 2005). In addition, preliminar identified dental pathologies (Lalueza 1996) have made it possible to diagnose new dental anomalies that are little known in hunter-gatherers. These results extend the record of stress indicators and oral pathologies in Late Pleistocene human populations in Western Europe.
General setting of the human tooth assemblage from Level E at Balma Guilanyà
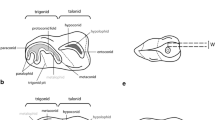
Balma Guilanyà is located in the Serra de Busa (X=385087 Y=4660546, UTM H31N ED50) at 1,157 m above sea level, on the first escarpment of the southern slopes of the Prepyrenees, in Lleida in the north-eastern Iberian Peninsula (Fig. 1A). This Eocene conglomerate shelter was partially destroyed by the construction of a forest road (Fig. 1B), exposing a 15x 5 m sedimentary deposit. The lithic technocomplexes allow us to differentiate an Upper Unit, assigned to the Early Holocene, and a Lower Unit, assigned to the Late Glacial; these attributions have been validated radiometrically (Martínez-Moreno et al. 2006; Martínez-Moreno and Mora 2009). In this 2.5-3 m thick sequence, several block falls can be detected, and at the archaeostratigraphic level, there was a collapse of the conglomerate wall shelter overhang on the roof of the Lower Unit, on top of which the Upper Unit is located. This event prevents the vertical migration and mixing between Late Glacial and Holocene artefacts and bones (Martínez-Moreno and Mora 2009, Fig. 2).

General setting. A) Location of Balma Guilanyà on the southern slope of the Pyrenees, north-eastern Iberian Peninsula. B) General view of the rock shelter

Calibrated 14C ASM intervals. The grey area corresponds to the date UBAR-367 which is not relevant to the discussion. The Ua-34297 and Ua-34298 distributions correspond to the direct dates from the human remains. The blue line marks the Gi 1a/ Gs 1 boundary (Rasmussen et al. 2014). Image produced using OxCal v4.4.2 (Bronk Ramsey 2020)
The excavation of a 25 m2 area made it possible to define five stratified archaeological units. In the Upper Unit, Mesolithic levels C and C1 were identified, while in the Lower Unit, occupations E, EJ and K, attributed to the Azilian, followed one another. These assemblages contain lithic remains (Martínez-Moreno and Mora 2009), fauna (Martínez-Moreno and Mora 2011), charcoal (Allué et al. 2012), and in level E were recovered some marine shell ornaments (Dentalium sp., Columbella rustica, Cyclope neritea, and Nassarius incrassatus) that come from more than 100 km away (Martínez-Moreno et al. 2010).
In these levels there are no indicators related to the installation in the shelter of features related to dwelling as hearths; however, fire use is evidenced by abundant charcoal, as well as burned lithics and bones. No pit construction was identified, and the archaeological material were found integrated into the sediment. Level E is below the before mentioned fall of the shelter ledge and has an average thickness of 25-30 cm. This vertical dispersion suggests repeated seasonal visits to the shelter to hunting wild goats (Martínez-Moreno and Mora 2011). These materials, including human remains, form low-resolution palimpsests accumulated from several events that took place over an imprecise time range. On the top of level E, within a confined area of approximately 20 cm2 and exhibiting minimal vertical dispersion, an intriguing collection of nine isolated human teeth was recovered proximate to a fragmented and poorly preserved cranial vault. The archaeological record indicates potential disturbance from the construction of a rural road in the vicinity of the site. However, the impact of this disturbance on the archaeological deposit will be considered in the analysis of the teeth accumulation processes. Apart from this consideration, no evidence of natural depressions or deliberately excavated pits indicative of intentional deposition of these human remains was discerned during surface excavation. Notably, the human teeth were found interspersed with lithic and faunal remains originating from the same stratigraphic level (Terradas et al. 1993).
There are six 14C AMS dates for the assemblage, obtained from two pieces of charcoal, one Corylus shell, the collagen from a molar of Capra pyrenaica, and two samples of collagen extracted from human remains (right M1 (E-3998) and a skull fragment (E-NC)) (Table 1). This series of BP dates has been calibrated with the IntCal 20 atmospheric curve (Reimer et al. 2020) and transformed into 2δ chronometric intervals using OxCal v4.4.2 software routines (Bronk Ramsey 2020).
We have integrated these dates into our discussion of the chronometry of Level E, except UBAR-367, with a radiometric range very imprecise. Although the human tooth collagen date E-3998 (Ua 34297) also has a high δ, its chronometric range is compatible with the ranges derived from the charcoal samples (Beta-247706), Corylus (Beta-210729), and the collagen from the molar of a Capra treated with ultrafiltration (UF) (Beta-252286), which situate the Level E occupations between 13,100-12,750 cal BP. The high δ date obtained from the human skull fragment (Ua 34298) presents a notable deviation from the rest of the sample series, affirming its antiquity while also suggesting potential issues with collagen preservation owing to bone damage by significant root activity. This date, although indicative of antiquity, warrants caution and could benefit from further investigation during subsequent site visits. Direct radiometric dating of human crania yields broad and imprecise temporal ranges, underscoring the need for refined dating methods. Employing collagen purification protocols, such as those outlined by Bronk Ramsey et al. (2004), not utilized in previous analyses at the Uppsala laboratory, could provide a clearer understanding of the temporal scope of Level E. The application of ultrafixation protocol on tooth collagen from a Capra pyrenaica specimen (Beta-252286) highlight the potential for improved dating precision and warrant consideration in future analyses.
The chronometric position of Level E is not a secondary issue. In previous studies, we have proposed Level E -and consequently the human bones- should be integrated in the end of the Gi 1a chronozone (Allerod) (Martínez-Moreno and Mora 2009). Following the chronostratigraphic review proposed by the INTIMATE group, the onset of Gs 1 in the NGRIP core was defined as 12,896 + 4 b2k (Rasmussen et al. 2014). In line with this new boundary, the formation of Level E would be interpolated between the end of Gi 1a interstadial and the beginning of Gs 1 stadial (Fig. 2). Assuming this event of abrupt climate change affected deeply ecosystems and humans, it is a notion not easy to evaluate from the archaeological palimpsest of Level E. However, some dental pathologies detected in this paleoanthropological set, allow discussing the possible impact of environmental degradation over the human populations as it has been proposed for central Europe hunter-gatherers in this period (Weber et al. 2011).
Material and methods
The palaeoanthropological assemblage from Level E comprises the mentioned poorly preserved cranial vault, where fragments of the temporal, parietal, and parietal-temporal suture have been identified along with three manual phalanges, and a radius diaphysis. The 17 teeth, central focus of this revision, have been the subject of several studies, with a focus on their odontological identification and the estimation of the number of individuals (Lalueza 1996; Ruíz et al. 2005).
The initial findings in 1992 of nine permanent teeth were presented in a first study by Lalueza (1996). Based on the dental wear and eruption, as well as the morphology of the postcranial bones, Lalueza proposed that all the assemblage could be attributed to a unique individual, specifically a woman of between 20 and 25 years old. He mentioned several teeth affected by various pathologies, including mild hypoplasia affecting almost all teeth, carious perforations on an upper right third molar (NE-3381), and an upper deciduous canine (E-1184) interpreted as an impacted tooth on the permanent canine (Table 2).
A later study reported four recently discovered teeth (E-3998, E-4111, E-4182, E-5020). The identification, dental repetition and dental wear degree on the fourteen teeth, were attributed to one individual aged between 25 and 35; a second between 17 and 25, and a five years old child. However, the wear on some of the teeth could be compatible with a possible fourth individual. That study mentions no dental pathologies, and the deciduous canine is defined as an upper central incisor (Ruíz et al. 2005).
Subsequently, three skeletal remains that do not necessarily correspond to the same individual were analysed for 13C/15N stable isotope content (García-Guixé et al. 2009). Isotopes were extracted from the radius diaphysis (E-1204) at the Dept. of Archaeological Sciences at the University of Bradford (UK), while an upper molar (E-3998) and a cranial fragment (E-NC) were investigated at the Archaeological Research Laboratory at Stockholm University (Sweden). The collagen extracted of a tooth and a cranial fragment were dated by 14C AMS in-dated by the Radiocarbon Laboratory in Uppsala (Sweden), and these results have been discussed above.
Thus, in addition to the previously published teeth, we report four additional dental remains discovered in 2008 (E-13146, E-11259, E-11710, E-12040). These unpublished teeth were found at the same level E as the rest of the assemblage, as suggested by their contextual and spatial distribution and location within the stratigraphy of the site. Therefore, the complete assemblage consists of 16 permanent teeth, and one deciduous molar (Figs. 3, 4, 5, and 6).

Tooth from Individual I from Balma Guilanyà: left M1 (E-5020). Views from left to right are occlusal, buccal, mesial, lingual, and distal. B: buccal, M: mesial

Teeth from Individual II from Balma Guilanyà. From top to bottom: left I1 (E-13146), left I1 (E-5019), right I2 (E-1232), right C1 (E-5018), left P4 (E-4111), left M1 (E-4182). Views from left to right are occlusal, buccal, mesial, lingual, and distal. B: buccal, M: mesial

Teeth from Individual III from Balma Guilanyà. From top to bottom: left i1 (E-1184), left P3 (E-1224), left P4 (E-11259), right P4 (E-1216), right M1 (E-3998), right M1 (E-5016), right M2 (E-1173), right M2 (E-5017), right M3 (E-3381), left M3 (E-11710 and E-12040). Views from left to right are occlusal, buccal, mesial, lingual, and distal. B: buccal, M: mesial. The holes visible in the root of M1, M2 and M3 show a perforation made to extract samples for stable isotope analysis (García-Guixé et al. 2009)

Three-dimensional digital models of i1 (E-1184). B: buccal, O: occlusal, M: mesial, S: section
To describe them, the non-metric characters of the deciduous and permanent dentition were assessed following the morphometric guidelines defined by Nelson (2020). The criteria of Zoubov (1968) were applied for tooth wear Zoubov (1968), who expanded Gerasimov (1955) tooth wear assessment system. While maintaining the scale, Zoubov altered certain aspects in the development of crown components, and proposed separate scales for premolars/molars and incisors/canines. We used the classification proposed by the FDI (1992) for enamel hypoplasia defects. In the permanent teeth of modern humans, dental chronology is estimated from tooth eruption, temporal root resorption sequences and developmental patterns assessed according to Moorrees et al. (1963), which was modified by AlQahtani et al. (2010) in four stages of classification detailed with descriptions and illustrations that highlight changes in enamel, dentin, and pulp.
Crown dimensions (height, and mesiodistal and buccolingual diameter), neck dimensions (mesiodistal and buccolingual diameter), and vertical curvature measurements (root length, and mesial and distal curvature of the cervical line) were recorded using a standard digital calliper with an accuracy of 0.01 mm. The dental nomenclature in the text, figures and tables employs superscripts to denote maxillary teeth and subscripts to denote mandibular teeth.
Bivariate crown plots obtained from the mesiodistal (MD) and buccolingual (BL) diameters were compared with measurements for Homo sapiens from various chronologies (late Pleistocene and early Holocene, Neolithic, and medieval times) compiled at http://anthropologicaldata.free.fr (Voisin et al. 2012) (SM Table 1). This comparative analysis excluded remains for which the full dimensions of the crown could not be obtained or with extreme wear. The data were processed with Past 4.03 software (Hammer et al. 2001).
Eleven remains were scanned using high-resolution RX-XWT computed microtomography (μCT) at the Laboratory for the Non-Destructive Analysis of Geological Materials-CORELAB (Univ. Barcelona, Spain). The X-ray tube was set at 70kV and 30 mA using an aluminium filter (0.5 mm thick) and teeth were scanned at a magnification of 8.46, for between 16 and 19 minutes. The volumetric data was reconstructed using isometric voxels ca. 15 mm, being treated with Avizo software (FEI-Visualization Sciences Group Inc.) to generate 2D and 3D reconstructions as well as 3D digital models to examine the internal structure of the teeth. These elements bring the catalogue of dental pieces up to date and make it possible to identify new odontological diagnostic elements and define morphological elements, conservation status and taphonomic alterations more precisely. These descriptions were compared metrically with other prehistoric populations.
Results
MNI
The minimum number of individuals (MNI) and age-at-death were estimated, as well as several dental pathologies, emphasising their diagnosis. Based on the degree of development, dental wear and associated anomalies, the 17 teeth from Balma Guilanyà allow us to estimate a minimum of three individuals, with two groups of teeth attributed to two adult individuals, and one isolated tooth that belonged to a child (Table 3). Nevertheless, while we propose a tentative MNI, it is crucial to discuss the possibility of a fourth individual represented, given the differences in taphonomic alteration and the slight morphological difference of two lower molars. Even when our hypothesis is supported by the exhaustive description of teeth, considering the absence of maxilla or mandible remains, we acknowledge the need for further contextual data to our conclusions.
Here we will summarize the key morphological characters of the Balma Guilanyà dental assemblage. A detailed description and visual material of each tooth can be found in the Supplementary Material (SM Text 1 and SM Figures 1-29) and the basic metric parameters are presented in Supplementary Material (SM Table 2).
Individual I
The only tooth that corresponds to Individual I is the left M1 (E-5020) (Fig. 3). The root of M1 is in the formative stage with only the coronal third and the bifurcation area visible, close to grade R1/4 of Moorrees et al. (1963), a stage that corresponds to an age at death of approximately 5 years ± 9 months based on dental development standards for modern humans (AlQahtani et al. 2010). Due to its recent eruption, crown wear is non-existent (grade 0). The mesial surface of the mesio-buccal cusp has a small contact surface with the left m2. The differences in the dental chronology with respect to the rest of the assemblage point to a child represented by a single remain.
Lower left first molar with well-preserved crown and still-forming root with only the coronal third. Considering the root finishes its formation at 9-10 years of age and the crown is completely calcified, this could correspond to a child of 5 years ± 9 months. Due to its recent eruption, crown wear is non-existent (grade 0). The mesial surface of the mesio-buccal cusp has a small contact surface with the left m2 (SM Figure 1).
Individual II
This individual is represented by left I1 (E-13146), left I1 (E-5019), right I2 (E-1232), right C1 (E-5018), left P4 (E-4111) and left M1 (E-4182) (Fig. 4). The contact facets of I2 and C are compatible, while the preservation, appearance, hypoplastic lines and wear of I1 (E-13146) and I1 (E-5019) are similar to the former. The wear of M1 (E-4182) is compatible with the anterior teeth and points to an adult individual of about 40 years of age. Although only a minor portion of the P4 (E-4111) and M1 (E-4182) has been preserved, they exhibit subtle characteristics for their identification. The lingual fragment from the P4 exhibits a longer mesial side of the lingual cusp, distinguishing it from a buccal fragment. Meanwhile, the fragment from the M1 reveals the mesio-lingual and part of the disto-lingual cusps. Notably, it presents extensive wear (grade 4) and retains traces of the contact point with the lower left fourth premolar.
Individual III
The following dental remains are assigned to Individual III: left i1 (E-1184), left P3 (E-1224), left P4 (E-11259), right P4 (E-1216), right M1 (E-3998), right M1 (E-5016), right M2 (E-1173), right M2 (E-5017), right M3 (E-3381), and two fragments from the same left M3 (E-11710 and E-12040) (Fig. 5). Several dental anomalies are assigned to this individual, including the prolonged persistence of i1 in the maxilla (maybe as a consequence of an odontoma or mesiodens). Equally the dens invaginatus of P3 (Beynon 1982; Hegde et al. 2014; Morfis 1993; Ruprecht et al. 1986; Shifman and Tamir 1979) can correspond with the abnormal development of the deep pits observed in P4, P3, P4, M3 and the two fragments of M3. This pattern of anomalies will be discussed below (see Discussion). The contact points between M1, M2 and M3 articulate perfectly and show a degree of dental wear suggesting an age of 20-25 years. Although deciduous tooth i1 could be linked to Individual I, the excessive wear and lack of root resorption contrast with the apical resorption of the permanent incisor that occurs at around 5 years of age.
The morphological characteristics displayed by the right M1 (E-5016) and the right M2 (E-5017) could suggest the presence of a fourth individual. These teeth notably differ from those attributed to Individual II in terms of occlusal wear, tooth dimensions, and preservation status. Conversely, they align more closely with those of Individual III. However, these two teeth seem to have undergone taphonomic processes that have altered their surface morphology. Their preservation status notably differs from the other pieces attributed to Individual III, despite being discovered alongside the teeth assigned to Individuals II and III within a confined area of 20 cm2. If these two teeth indeed belong to Individual III, they should exhibit occlusal wear facets consistent with the right M1 (E-3998) and the right M2 (E-1173), both attributed to this individual. However, such facets are not discernible on the occlusal surface of the lower molars, possibly due to their poor preservation status. Nevertheless, considering their dimensions, degree of wear, and radiographic characteristics, it is plausible to assign these teeth to Individual III. Given the use of a minimum number of individuals and the absence of substantial evidence to introduce an Individual IV based solely on these two teeth, we decided to maintain a conservative stance, proposing a minimum number of three individuals.
Metric comparison
This issue has been previously addressed (Ruíz et al. 2005) and we thought it appropriate to return to it. The comparative metric data of MD and BL crown diameters for the upper (SM Table 2) and lower dentition (SM Table 3) indicate that the Balma Guilanyà teeth are slightly small, although they fit within the variability of Holocene humans (HH). The plots indicate that the Balma Guilanyà teeth are smaller than other Pleistocene specimens but close to European populations from the Late Pleistocene and Holocene (SM Fig. 30-41). The crown dimensions of M2 (E-5017, BL 11.3 mm) are at the upper end of the HH range and fall within those of Upper Palaeolithic populations of Homo sapiens (UPHS). However, the BL diameters of I1 (E-5019, 5.35mm) and I1 (E-13146, 6.48mm) appear at the lower end of the HH range, these being small teeth within their respective tooth classes.
Dental anomalies
In this catalogue, the anomalies detected in i1 and P3 stand out and should be discussed in more detail.
i1 (E-1184)
This significant tooth in the study was diagnosed as an upper decidual canine (Lalueza 1996). It presents very advanced wear (grade 6) with a small cervical portion of the crown preserved where a thin layer of enamel can be distinguished delimiting the CEJ whose measurements, as mentioned, are consistent with the average for deciduous upper central incisors in modern humans (Black 1897). The root is wider mesiodistally than buccolingually, and the above measurements rule out a decidual canine (Fig. 6). Although it could be mistaken for a microdontic definitive upper incisor, the prominent cervical enamel ridges, the very thin enamel and the unaccentuated curvature of the cervical line correspond to a deciduous left upper incisor. The X-ray examination revealed a very wide root canal and narrow root walls. The absence of root resorption and the advanced wear indicate that it was a long-lasting functional tooth in an adult individual.
Hyperdontia defines an excessive number of teeth in relation to the normal dental configuration. One or more teeth may appear in the dental arch, either unilaterally or bilaterally on the mandible or maxilla, or both, and may be erupted or unerupted. One of the reasons of persistence of primary teeth are lack of space caused by the presence of a mesiodens (supernumerary tooth) and an odontoma (Brin et al. 1982; da Costa et al. 2008; Mercedes Gallas and García 2000). These are often associated with other anomalies such as dens invaginatus (SM Text 2). In European populations the incidence of this is estimated to be between 0.1-3.8% (Rajab and Hamdan 2002), with mesiodens being the most frequent supernumerary tooth (prevalence between 0.15%-1.9%), usually between the maxillary central incisors (Sedano and Gorlin 1969). Other possible causes could be bone or mucosal barriers that interfere with normal eruption, dilacerations caused by traumatic injury to deciduous teeth, ectopic development of the permanent germ or the strengthening and enlargement of the follicle around the tooth (Brin et al. 1982; Manilla and Katagiri 2017; Tanaka et al. 2008; Becktor et al. 2002; Savage and Kokich 2002).
P3 (E-1224)
The analysis of the occlusal surface of the P3 crown revealed two abnormally deep pits, indicating the possible presence of a dens invaginatus (or dens in dente). X-ray analysis confirmed modified tooth development resulting in the infolding of the enamel and dentine towards the pulp, a malformation that occurs during early morphogenetic stages prior to the completion of tooth mineralisation (Fig. 7). Invagination begins at the crown, manifesting as an opening on the coronal surface that develops apically at different depths through a pit, a tiny duct that ends in a fold, forming a pocket of organic material beneath the enamel surface (SM Text 2).

Three-dimensional digital models of P3. The invagination fossa has a coronal entrance and exceeds the cementoenamel junction (CEJ) producing a cavity inside the root. M: mesial; O: occlusal; S: section
High resolution μCT 2D and 3D images reveal a Type II invagination that extends apically beyond the CEJ, generating a narrow canal that connects to a confined cavity at the root (Oehlers 1957). The apical end of the root has been partially destroyed by infection, and in life the tooth was probably affected by pulpitis arising from the communication between the invaginated cavity and the pulp cavity, infecting the dental pulp through the root canal to the apex. This process would have led to apical periodontitis which was responsible for the root defect (Kendrick 1971; Chen et al. 1990; Whyman and MacFadye 1994). Additionally, the internal cavity in P3 shows reduced enamel density and irregularity, indicating hypomineralisation and the altered chemical structure of the enamel (Beynon 1982; Morfis 1993).
In present-day populations, the prevalence of dens invaginatus is between 0.3%-10% (Backman and Wahlin 2001; Hamasha and Al-Omari 2004; Ezoddini et al. 2007). The most typically affected tooth is the upper permanent lateral incisor, accounting for up to 90% of cases, and to a lesser extent in canines; its presence in maxillary premolars and molars is extremely scarce (Conklin 1978; Hamasha and Al-Omari 2004). Although cases have been reported in the lower dentition, its incidence on mandibular teeth is rare.
The various known archaeological cases (SM Text 2) confirm that the diagnosis of this aetiology on the left P3 is not only unusual, but it also dates this dental pathology in human remains back to the end of the Pleistocene; at 13,000 years old, it is the first known case. This statement does not mean that invagination did not occur prior to this, as this pathology does not result in easily visible external elements, but it does highlight the need to observe internal tooth structures using radiological imaging, preferably applying 3D computed tomography.
Discussion
The study of the teeth assemblage recovered at Level E shows that they correspond to at least three individuals: one tooth is attributed to a 5-year-old child (Individual I); a second group consisting of six teeth are from an adult/older individual of about 40 years of age (Individual II); and ten dental remains are associated with a 20–25-year-old adult (Individual III). Additionally, there is a possibility of a fourth individual constituted by two teeth, a right M1 (E-5016) and right M2 (E-5017), based on their morphological characteristics and the observed taphonomic damage. This characterisation allows us to discuss the following aspects:
-
a)
the integration of the dental assemblage in relation to the process of dental reduction in Pleistocene and Holocene populations;
-
b)
the pathologies related to the pattern of anomalies seen in Individual III;
-
c)
causes of cariogenic processes;
-
d)
other stress indicators;
-
e)
contextual implications related to the tooth accumulation from Level E.
Evolutionary trends in the Level E dentition assemblage
The dentition assemblage is metrically consistent with the mesiodistal and buccolingual dimensions of the Holocene populations and fall slightly below the Pleistocene populations, and that is related to the reduction pattern affecting the upper and lower dentition and their crown dimensions (Pinhasi et al. 2006; Holt and Formicola 2008), a trend that seems to increase from the Holocene onwards and which is not very different from present-day populations. Except for teeth M1 (E-4182) and M2 (E-5017), the buccolingual dimensions observed in the Level E assemblage are in the lower range for Pleistocene populations, especially I1 (E-5019) and I1 (E-13146). This set confirms a moderate trend in Late Pleistocene European populations towards a reduction in the dimensions of dental crowns, which is more accentuated in the buccolingual dimensions.
Dental Pathologies and Anomalies in Individual III
A series of teeth exhibiting various pathologies have tentatively been attributed to the same individual, although the possibility of the existence of an additional individual is suggested by the right M1 (E-5016) and the right M2 (E-5017) within the assemblage that do not entirely conform to the characteristics of the rest of Individual III. One of these teeth displays several pathologies (E-5017). Considering the identification remains subject to potential uncertainties, should the existence of a fourth individual be confirmed, Individual III would not encompass all the pathologies present in the teeth assemblage from Balma Guilanyà. Nonetheless, this combination of pathologies within a relatively small assemblage is noteworthy, and even in some cases, to our knowledge, is unprecedented within the paleoanthropological record (see Trinkaus 2018; Lacy 2014). We propose to highlight the elements that contribute to this pattern of anomalies while acknowledging the need for cautious interpretation.
In view of their particular characteristics, the dental pathologies attributed to Individual III should be described in detail. At the odontological level, the identification of a dens invaginatus in P3 stands out. This is a dental anomaly that is difficult to diagnose and is detected by the presence of abnormally deep pits on the occlusal surface of the crown. X-ray and μCT imaging allow us to recognise a protrusion inside the root forming an enamel-lined space that connects the internal cavity with the outside of the tooth through a narrow canal. This anomaly is not common in present-day populations (less than 10%) and is rarely mentioned in the archaeological record (SM Text 2), this being the first case in which dental invagination has been described in populations of Homo sapiens from the Upper Palaeolithic. Rather than a null prevalence in Upper Pleistocene populations, we believe that detecting this pathology calls for the analysis of the internal dental structure using radiological images and 3D computed tomography, a technique which is becoming more widely used in odonto-palaeoanthropological studies.
We propose that this pathology warns of a pattern of dental alteration that affects other teeth. At the odonto-pathological level, dental invagination is linked to the persistence of dentition, and the appearance of supernumerary teeth (Shifman and Tamir 1979; Beynon 1982; Ruprecht et al. 1986; Morfis, 1993; Hegde et al. 2014). The i1 located on the midline of the maxillary arch must have caused the uneruption and impaction of the upper permanent central incisor, preventing the normal exfoliation of the primary incisors. Likewise, the abnormally deep pits observed in the crowns of P3 (E-1224), P4 (E-1216), M3 (E-11710), and M3 (NE-3381) lead us to suggest that rather than cariogenic alterations, as discussed in the following section, they could be considered incipient invaginations congruent with dens invaginatus (Fig. 8).

3D digital models of M3, P4, P3 and M3 showing the development of abnormally deep pits on the occlusal surface
Another anomaly observed is the fusion of the roots affecting molars M1 (E-3998), M2 (E-1173), M3 (NE-3381), along with the M2 (E-5017) provisionally ascribed to Individual III (Fig. 9). Although this is not exceptional in the case of a M3, which can develop one or two roots (Sidow et al., 2000; Čosić et al., 2008), the prevalence of fused roots in M1 and M2 (respectively 7% and 21%) and in lower first and second molars (0.7% and 12.6%) is an interesting feature (Martins et al., 2016). This phenomenon affecting the morphology of the roots made it difficult to identify the E-5017 tooth, which could correspond to either M2 or M3, while the morphology of the occlusal face could correspond to that of an M1. The M1 and M3 contact points confirm that this is an "atypical" M2.

3D digital models of M2, M2, M1 and M3 showing fused roots
This combination of pathologies suggests a potential aetiology that may indicate populations with a history of inbreeding, a characteristic often associated with hunter-gatherer groups inhabiting regions of low population density (Holliday et al. 2014). This hypothesis could be evaluated from paleogenetic studies on individuals from Level E.
Cariogenic processes
Carious lesions in living humans are most frequently initiated in the enamel of tooth crowns, particularly in the occlusal surfaces of molars and the areas just below the contact points between neighbouring teeth, known as the approximal areas (Hillson 2001). The occurrence of caries in M2 (E-5017) and M3 (NE-3381) is another anomaly observed in Individual III. The progressive destruction of the tooth by bacteria (e.g., Streptococcus mutans) has a complex aetiology involving various factors such as saliva components, tooth morphology, hygiene, hormone levels, and dental plaque ecology (Nikiforouk 1985; Seif 1997; Love and Jenkinson 2002; Lukacs and Largaespada 2006). Its development is closely linked to diets rich in sugar and carbohydrates (Hillson 2001; Hillson 2005; Pezo and Eggers 2010), while in hunter-gatherer populations, its frequency is low due to the more varied diet (Oelze et al. 2011). However, it has been reported that hunter-gatherers in Western Europe and the Near East processed carbohydrate-rich wild plants prior to the widespread cultivation of cereals (Piperno et al. 2004; Revedin et al. 2010; Liu et al. 2013). The prevalence of caries in Taforalt (Morocco), 15-12.6 ka cal BP, has therefore been related to the intake of carbohydrate-rich wild acorns and pine nuts (Humphrey et al. 2014). Likewise, in the Late Glacial sites of Villabruna and Fredian (Prealps, Italy), a similar context to level E at Balma Guilanyà, there is evidence of intentional manipulation aimed at sanitising decayed teeth (Oxilia et al. 2015; Oxilia et al. 2017).
Even so, the cariogenic capacity of sugars in honey and sweet fruits in the diet of hunter-gatherers should not be underestimated (Bernal et al. 2007; Nelson et al. 1999; Neves and Cornero 1997). However, the 15N/13C stable isotope results from three skeletal remains from Balma Guilanyà, including M1 (E-3998) attributed to Individual III, suggest little contribution from plant-based resources as opposed to the intake of proteins obtained from terrestrial herbivores (García-Guixé et al. 2009), a diet with a typically low cariogenic capacity (van Palenstein et al. 1996).
Therefore, in the current case, we suggest that the carious lesions in both teeth should not be related to a sugar-rich diet but rather to a dental morphological predisposition (Fig. 10). The progression of carious lesions on occlusal surfaces follows a distinctive pattern, first with the appearance of tiny opaque spots that gradually grow in size, leading to a slightly roughened enamel surface. The carious process continues as the enamel surface breaks down, forming a cavity. Furthermore, this cavity tends to widen as it reaches the enamel-dentine junction (EDJ), with the enamel cavity's walls being undermined (Hillson 2005).

Molars assigned to Individual III. Upper row: M2; lower row: M3. Photographs (a) and 3D digital model (b) in occlusal view, and micro-computed tomography sections (c). Sections show a carious lesion in the central fossa of the occlusal surface of the M2 crown, and the abnormally deep pits on the occlusal surface of M3. The incipient caries has progressed within the small cavities; the infection has extended into the underlying dentine. B: buccal, M: mesial, L: lingual, D: distal
M2 and M3 would have had small cavities in the enamel that would be ideal for the accumulation of food debris enabling the development of infection. The deep occlusal pits of M3 possibly indicate the origin of the infection that could have caused caries to develop on the occlusal surface of the premolars and molars (Hillson 2001; Hillson 2005). In fact, an incipient caries is visible in a deep occlusal fossa of M3, while the infection in M2 overshadows the preceding occlusal fossa. Also, one of the M3 fragments (E-11710) has a small cavity underneath a small occlusal fossa (SM Fig. 11). These indicators allow us to propose that the carious lesions identified in M3 and M2 derived from a pre-existing dental pathology that favoured the development of the disease in these teeth. These carious lesions result from a complex interplay of factors, including bacterial activity, dietary habits, tooth morphology, and ecological influences. While the diet, particularly the consumption of sugars and carbohydrates, has historically been implicated in the development of dental caries, the dental morphology of Individual III, including deep occlusal pits in the M3, likely played a significant role in facilitating the onset and progression of caries. This case underscores the importance of considering both intrinsic and extrinsic factors in understanding cariogenic processes. The historical dietary context of this individual, as revealed by stable isotope analysis, further emphasizes the role of diet in dental health. These findings highlight the need for a comprehensive approach when examining dental pathologies and their aetiology, considering both genetic predispositions and environmental factors.
The unique accumulation of pathologies detected in Individual III, which contributed to the development of morphogenic caries, raises intriguing questions about potential genetic anomalies that favoured the development of infectious processes, pulpitis and other oral diseases that would have affected oral health and quality of life of this individual. However, given the uncertainties surrounding the dental assignation to a fourth individual, these observations warrant further investigation and should be approached with caution. This issue could alter the interpretation of the development of cariogenic processes in late glacial populations, and should be the subject of future research, aiming to elucidate the underlying factors influencing the dental health of Late Glacial populations.
Other stress indicators
Enamel hypoplasia is another prominent pathology (Lalueza 1996). This is a term used to describe a defect, or the absence, of enamel on a region of the tooth. It has a wide range of causes related to nutritional stress, infection, and vitamin deficiencies during the formation of tooth enamel. Lines or grooves corresponding to mild enamel hypoplasia were identified on the buccal side of P3, P4, P4, M1, M3, and M3 of Individual III, and I1, I1, I2, and C of Individual II.
These rare or not very severe alterations in the dentition of European Upper Palaeolithic hunter-gatherers (Trinkaus et al. 2006) increased in frequency during Late Glacial times and have been associated with vitamin deficiencies at the end of the Pleistocene (Holt and Formicola 2008), linked to environmental crisis events such as the Younger Dryas (Weber et al. 2011). In this possible scenario (chronometrically, Level E is positioned at the boundary between Gi 1a/Gs 1), we believe that hypoplasies in these individuals do not show the consequences of degrading climate conditions affecting their quality of life. Alternatively, the appearance of enamel hypoplasia may be linked to situations of nutritional stress during childhood as a result of introducing solid food into the diet (Hillson 2014). This transitional period when breast milk is gradually withdrawn, and solid foods are consumed in increasing quantities is known as ‘weaning’.
A significant divergence has been reported in the developmental stress patterns related to enamel hypoplasia between Upper Palaeolithic and Middle Palaeolithic populations (Skinner 1996). Notably, the Upper Palaeolithic group exhibits the highest stress levels during early infancy. This distinction in stress patterns could suggest that Upper Palaeolithic communities endured different stressors. Weaning is known to trigger the highest metabolic stress during early childhood in traditional societies (Moggi-Cecchi et al., 1994). Insights from dental data indicate that the Late Upper Palaeolithic population in western Europe displays the highest known prevalence of linear enamel hypoplasia (LEH), affecting 7 out of 10 individuals, experiencing significantly more stress in the first two years of life (Skinner 1986). Also, an early enamel hypoplasia research revealed that individuals in contemporary populations primarily exhibited enamel defects in the cuspal region, whereas those in the prehistoric communities exhibited hypoplasia in the cervical part of the enamel (Rose 1977), a pattern observe in some Balma Guilanyà teeth.
The report of osteopenia (bone mass deficiencies) in newborn facial structures, suggests that weaning alone might not be the sole factor at play. Research into this matter strongly implicates vitamin A deficiency (Skinner et al. 1994). The availability of vitamin A is influenced by maternal diet and the exposure of both mothers and infants to diseases, known to deplete vitamin A reserves (Stephensen et al. 1994). However, recent high resolution pollen studies in paleolakes on the southern Pyrenean slope point to mild forests in the area during Gi 1 (Rull et al. 2023). So, considering the relatively stable growing seasons in Late Pleistocene Europe, the natural availability of vitamin A would likely remain constant.
Changes in climate conditions does not appear to provide a satisfactory explanation for the observed stress patterns. There are indications of a population expansion that might have resulted in nutritional and demographic stress at some point. Demographic simulations, based on ethnographic and palaeoclimate data, suggest that climate was a major driver of population dynamics, leading the population size to reach about 330,000 people in 30,000 BP (Tallavaara et al. 2015). Moreover, a noticeable uptick in the number and size of archaeological sites during the Upper Palaeolithic has been observed (Arenas del Amo et al. 2024). Nevertheless, later, with the progressive cooling of temperatures, there was a significant drop to 130,000 people. The contraction of European populations is consistent with genetic studies involving mtDNA with Y-chromosome patterns (Pala et al. 2012; Soares et al. 2010; Torroni et al. 2001). Even most of the evidence supports the notion of a population increase during the Pleniglacial Upper Palaeolithic, differential access to limited dietary resources does not seem to be a possible explanation of the observed pattern of stress in the Upper Palaeolithic sample. In the chronological framework of Balma Guilanyà, Late Upper Palaeolithic populations were in a process of recolonization and growth after the population decline during the LGM, a growth that was likely gradual rather than sudden (Posth et al. 2023).
It is also necessary to address the correction regarding tooth wear in the European Upper Palaeolithic individuals, as variations in dental wear within prehistoric populations introduce a significant challenge in accurately interpreting age at death based solely on tooth wear patterns. Dental wear analysis, particularly in the context of Palaeolithic populations, is influenced by various factors such as diet, environment, and innate dental characteristics. The prevalence of dentine exposure, as observed in modern industrialized societies, typically occurs around the age of 6 or 7 years, with minimal exposure even among older children (Dugmore and Rock 2004; Milosevic et al. 1994). However, in Palaeolithic populations, a substantial proportion of children exhibit exposed dentine on deciduous molars at a much younger age, indicating accelerated dental wear (Bas et al. 2023). This early onset and intensity of dental wear are attributed to dietary factors, with Palaeolithic diets being significantly coarser and more abrasive compared to modern diets. Additionally, variations in dental wear within the population suggest differences in childhood diets either at a given time or over generations, reflecting the dynamic nature of dietary practices. Therefore, while diet influences overall wear intensity, the distinct wear patterns observed in UP and Mesolithic samples suggest a more nuanced understanding of dental use beyond dietary factors. Despite variations in dental wear, it's crucial to acknowledge the limitations of macroscopic analysis, as it may not fully capture underlying factors such as innate differences in enamel thickness and dental tissue proportions. Further research incorporating imaging of internal dental structures is needed to better understand the interaction between genetic and environmental factors shaping dental wear in Palaeolithic populations.
Origin of teeth accumulation
The discovery of isolated human remains in contexts from the end of the Upper Palaeolithic is an issue recently analysed in detail. Although in some cases various types of peri/ post-mortem treatment or manipulations are identified (see Kerner 2018; Marsh and Bello 2023; and references), in many cases the causes that explain their presence in the archaeological record are not evident. Within this perspective, the anthropological complex of Level E exemplifies the difficulties related to the identification of processes and agents involved in the configuration of this accumulation.
At the beginning of this article, we stated that we did not question the Late Glacial dating for these remains, but it was not easy to explain the contextual significance of the dental assemblage. A portion of these remains, namely the fragments of a poorly preserved cranial vault and nine isolated teeth, form an intriguing grouping recovered in an area of 20 cm2, in a preliminary test pit made on the roof of Level E. During the excavation, no natural or intentional depression was detected to explain this accumulation, but contextual observations provide elements to the discussion.
Archaeological remains appeared at the limit of the preserved surface of the shelter. In the archaeological sequence, the vertical dispersion of the artifacts in a N-S direction evidencing two stratigraphic levels separated by the massive fall of the cornice, separating an archaeological sequence in the upper part assigned to the Holocene, from another lower archaeological assemblage that corresponds to the levels assigned to Late Glacial. This conglomerate wall seals the development of Level E preventing the vertical migration of artifacts and bones between these sedimentary components.
At the same time, the vertical distribution suggests that the archaeological record does not have continuity towards the South and were destroyed during the construction of a rural road that affects the upper sequence (Level C), and partially the lower sequence (Level E). The position of the paleoanthropological complex on the roof of Level E and below the fall of the block suggests that this accumulation would correspond to an initial occupation of the shelter between the end of Gi 1a and Gs 1 stadial during the Late Glacial. At the same time, he warns that the human remains were affected prior to the beginning of the fieldwork by the construction of the road that led to the discovery of the shelter (Fig. 11).

A) Schematic N-S vertical plot projecting the materials of levels C and E on the axis Y=100.800 – 101.200 mm along the squares Y= 511.000-507.000 (see orange box Fig. 12B). The conglomerate wall is positioned as well as the abrupt disruption to the South of Levels C and E, affected by the construction of the rural road. The position of human bone accumulation on the roof of Level E is indicated. B) Plan of Level E showing the positions of the teeth from Individuals I, II, and III. The positions of the M3 refit fragments (E-11710 and E-12040) are noted with a discontinuous red line. Enlargement of the tooth accumulation located in the lower left corner (X=100, Y=509) showing the position of the isolated teeth attributed to Individuals II and III
The accumulation of human remains overrepresents the abundance of fragmented skull remains systematically identified as isolated points recovered during the excavation. Adjacent to the cranial fragment were identified the isolated upper and lower teeth, specifically i1, P3 (E-1224), P4 (E-1216), M1 (E-5016), M2 (E-1173) and M2 (E-5017) attributed to Individual III and I2 (E-1232), I1 (E-5019) and C (E-5018) from Individual II. Although the P4, M1 and M2 series from Individual III could correspond to a partial mandibular remnant, the teeth appear disconnected without retaining their respective anatomical order. The i1 from Individual III is slightly displaced with respect to the other teeth with anomalies such as P3 (dens invaginatus) and P4, which has carious occlusal pits. However, the two fragments that refit to form M3 with this pathology are separated by 1 m and displaced by more than 2 m from the rest of the accumulation (Fig. 11).
These observations indicate that this assemblage has a complex taphonomic history. Apparently, the teeth are scattered randomly over the excavated surface, with no specific dental arrangement, making it difficult to infer the causes that explain the presence of the dental remains attributed to Individuals II and III in a discrete area in relation to the excavated surface. It is not easy to pinpoint possible causal agents of this accumulation possibly affected by the construction of the rural road. There are no identifiable alterations or marks on the dental remains, skull, or other skeletal remains that would indicate the involvement of carnivores or scavengers (taphonomical damage from vegetation roots has only been observed) acting on possibly inhumed bodies or individuals that had died accidentally.
The presence of isolated teeth from Individuals II and III that are not related to portions of the mandible or maxilla makes it difficult to propose peri/post-depositional causes that could have led to this grouping in a discrete area. Furthermore, the potential existence of two individuals raises several questions: did they both die in the same time interval, and does this association correspond to some kind of mortuary practice, as it has been revealed in similar chronocultural context (Kerner 2018; Marsh and Bello 2023). Although there are no indicators to answer these questions, the contextual information provided by this anomalous accumulation should be the subject of future reflections.
Conclusions
The human remains of H. sapiens from the end of the Pleistocene are the key to anthropologically, demographically and genetically reconstructing Western Europe hunter-gatherers. Level E at Balma Guilanyà has yielded bioanthropological indicators that provide information on the health conditions of these individuals. This assemblage, formed at the end of Gi 1a/start of Gs 1 (13,100-12,750 cal BP) comprises one deciduous and 16 permanent teeth that show several pathologies not commonly seen in Late Glacial hunter-gatherers. Morphological analysis, developmental stage observations, duplications, dental wear and the various pathologies point to the presence of at least three individuals: a child aged 5 years ± 9 months, a young adult aged 20-25 years, and a 40-year-old mature adult. This demographic composition is interesting in a shelter located at an altitude of over 1150 m, which would be visited by groups of adults and children. This diversity in age, rather than indicating the displacement of task groups specialising in the capture of mountain ungulates, suggests that domestic activities were also carried out in the shelter.
The dental anomalies affecting Individual III revealing a dens invaginatus, an overretention, and several teeth with varying degrees of caries. We propose that dental invagination explains the incipient caries in some teeth indicated by deep pits and advanced caries in two molars. In this case, we suggest that this pathology is not related to the ingestion of carbohydrates; instead, we believe that its aetiology is linked to these dental anomalies. It is possible that a genomic study of these remains could provide some explanation for the origin of these dental alterations, which are little known in Late Glacial human populations.
The record of enamel hypoplasia in a significant number of teeth attributed to Individuals II and III is equally significant. These indicate stressful situations related to seasonal crises or famines that these individuals suffered during their early childhood.
Finally, it is worth noting the anomaly represented by the group of isolated teeth recovered in a limited zone of the excavation. The assemblage of dental remains tentatively assigned to three distinct individuals, were found intermingled in a space in which no natural or constructed container has been identified makes it difficult to explain the origin and meaning of this accumulation. Rather than an unclear context (Meiklejohn 2009; Peyroteo 2016), it seems that the deposit was profoundly affected prior to the beginning of the excavation of the site, making it difficult to identify possible cultural processes involved in the formation of this accumulation. However, we believe that these indicators merit discussion to detect similar situations that could define possible crypto-funerary behaviour.
These issues indicate the importance of restudying the odontological assemblage from Balma Guilanyà Level E, which has substantially expanded our knowledge of the living conditions and pathologies of the Late Glacial hunter-gatherers in Western Europe.
Data Availability
No datasets were generated or analysed during the current study.
References
Allué E, Martínez-Moreno J, Alonso N, Mora R (2012) Changes in the vegetation and human management of forest resources in mountain ecosystems at the beginning of MIS 1 (14.7-8 ka cal BP) in Balma Guilanyà (Southeastern Pre-Pyrenees, Spain). C R Palevol 11:507–518. https://doi.org/10.1016/j.crpv.2012.04.004
AlQahtani SJ, Hector MP, Liversidge HM (2010) The London atlas of human tooth development and eruption. Am J Phys Anthropol 142:481–490. https://doi.org/10.1002/ajpa.21258
Arenas del Amo S, Armentano ON, Daura J, Sanz M (2024) Overview of the European Upper Palaeolithic: The Homo sapiens bone record. J Archaeol Sci Rep 53:104391. https://doi.org/10.1016/j.jasrep.2024.104391
Backman B, Wahlin YB (2001) Variations in number and morphology of permanent teeth in 7-year-old Swedish children. Int J Paediatr Dent 11:11–17. https://doi.org/10.1046/j.1365-263x.2001.00205.x
Bas M, Kurzmann C, Willman J, Pany-Kucera D, Rebay-Salisbury K, Kanz F (2023) Dental wear and oral pathology among sex determined Early Bronze-Age children from Franzhausen I, Lower Austria. PLoS One 18(2):e0280769. https://doi.org/10.1371/journal.pone.0280769
Becktor KB, Bangstrup MI, Rølling S, Kjær I (2002) Unilateral primary or secondary retention of permanent teeth, and dental malformations. Eur J Orthod 24:205–214. https://doi.org/10.1093/ejo/24.2.205
Bernal V, Novellino P, Gonzalez P, Pérez I (2007) Role of wild plant foods among Late Holocene hunter-gatherers from central and north Patagonia (South America). Am J Phys Anthropol 133:1047–1059. https://doi.org/10.1002/ajpa.20638
Beynon AD (1982) Developing dens invaginatus (dens in dente). A quantitative microradiographic study and a reconsideration of the histogenesis of this condition. Br Dent J 153:255–260. https://doi.org/10.1038/sj.bdj.4804912
Black GV (1897) Descriptive Anatomy of the Human Teeth. White Dental Company, Philadelphia
Brin I, Zilberman Y, Azaz B (1982) The unerupted maxillary central incisor: review of its etiology and treatment. ASDC J Dent Children 49:352–356
Bronk Ramsey C (2020) https://c14.arch.ox.ac.uk/. Accessed 10 Dec 2023
Bronk Ramsey C, Higham T, Bowles A, Hedges R (2004) Improvements to the pretreatment of bone at Oxford. Radiocarbon 46:155–163. https://doi.org/10.1017/S0033822200039473
Chen RJ, Yang JF, Chao TC (1990) Invaginated tooth associated with periodontal abscess. Oral Surg Oral Med Oral Pathol 69:659. https://doi.org/10.1016/0030-4220(90)90256-R
Conklin WW (1978) Bilateral dens invaginatus in the mandibular incisor region. Oral Surg Oral Med Oral Pathol 45(6):905–908. https://doi.org/10.1016/S0030-4220(78)80014-0
Čosić J, Galic N, Njemirovskij V, Vodanovic, M (2008) Root canal anatomy of third molars. In London: PEF/IADR International Dental Research Meeting
da Costa CT, Torriani DD, Torriani MA, da Silva RB (2008) Central incisor impacted by an odontoma. J Contemp Dent Pract 9:122–128. https://doi.org/10.5005/jcdp-9-6-122
Dugmore CR, Rock WP (2004) The prevalence of tooth erosion in 12-year-old children. Br Dent J 196(5):279–282. https://doi.org/10.1038/sj.bdj.4811040
Ezoddini AF, Sheikhha MH, Ahmadi H (2007) Prevalence of dental developmental anomalies: a radiographic study. Community Dent Health 24:140–144
Fahlander F, Oestigaard T (2008) The materiality of death: Bodies, burials, beliefs. In: Fahlander F, Oestigaard T (eds) The materiality of death: Bodies, burials, beliefs, BAR International Series 1768, Oxford, pp 1–16. https://doi.org/10.30861/9781407302577
FDI (1992) A review of the developmental defects of enamel index (DDE Index). Commission on Oral Health, Research & Epidemiology. Report of an FDI Working Group. Int Dent J 42:411–426
García-Guixé E, Martínez-Moreno J, Mora R, Núñez M, Richards MP (2009) Stable isotope analysis of human and animal remains from the Late Upper Palaeolithic site of Balma Guilanyà, southeastern Pre-Pyrenees, Spain. J Archaeol Sci 36:1018–1026. https://doi.org/10.1016/j.jas.2008.12.001
Gerasimov M (1955) Face reconstruction by a skull. Nauka, Moscow
Hamasha AA, Al-Omari QD (2004) Prevalence of dens invaginatus in Jordanian adults. Int Endod J 37:307–310. https://doi.org/10.1111/j.0143-2885.2004.00797.x
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol Electron 4:9
Hegde S, Jain M, Shubha AB (2014) A Rare Bilateral Presentation of Multiple Dens Invaginatus, Shovel-Shaped Incisor and Talon Cusp with Mesiodens. Kathmandu Univ Med J 12:292–295. https://doi.org/10.3126/kumj.v12i4.13737
Hillson S (2001) Recording dental caries in archaeological human remains. Int J Osteoarchaeol 11:249–289. https://doi.org/10.1002/oa.538
Hillson S (2005) Teeth. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511614477
Hillson S (2014) Dental markers of disease and malnutrition. In: Tooth Development in Human Evolution and Bioarchaeology. Cambridge University Press, pp 162–197. https://doi.org/10.1017/CBO9780511894916.007
Holliday TW, Gautney JR, Friedl L (2014) Right for the Wrong Reasons: Reflections on Modern Human Origins in the Post-Neanderthal Genome Era. Curr Anthropol 55:696–724. https://doi.org/10.1086/679068
Holt BM, Formicola V (2008) Hunters of the ice age: the biology of Upper Palaeolithic people. Yearb Phys Anthropol 51:70–99. https://doi.org/10.1002/ajpa.20950
Humphrey LT, de Groote I, Morale J, Barton N, Collcutt S, Bronk Ramsey C, Bouzouggar A (2014) Earliest evidence for caries and exploitation of starchy plant foods in Pleistocene hunter-gatherers from Morocco. Proc Natl Acad Sci 111:954–959. https://doi.org/10.1073/pnas.1318176111
Kendrick JK (1971) Periapical abscess from dens in dente. Oral Surg Oral Med Oral Pathol 31:838. https://doi.org/10.1016/0030-4220(71)90142-3
Kerner J (2018) Manipulations post-mortem du corps humain: Implications archéologiques et anthropologiques. Sidestone Press, Leiden
Lacy SA (2014) Oral Health and its Implications in Late Pleistocene Western Eurasian Humans. Dissertation, Washington University in St. Louis. https://doi.org/10.7936/K7RJ4GFQ
Lalueza C (1996) Les restes humanes del jaciment epipaleolític de la Balma de Guilanyà (Navès, Solsonès). Pyrenae 27:279–285
Liu L, Bestel S, Shi J, Song Y, Chen X (2013) Paleolithic human exploitation of plant foods during the last glacial maximum in North China. Proc Natl Acad Sci 110(14):5380–5385. https://doi.org/10.1073/pnas.1217864110
Love RM, Jenkinson HF (2002) Invasion of dental tubules by oral bacteria. Crit Rev Oral Biol Med 13:171–179. https://doi.org/10.1177/154411130201300207
Lukacs JR, Largaespada L (2006) Explaining sex differences in dental caries prevalence: saliva, hormones, and “life-history” aetiologies. Am J Hum Biol 18:540–555. https://doi.org/10.1002/ajhb.20530
Manilla AE, Katagiri MK (2017) Orthodontic-surgical treatment of an impacted central incisor. Rev Mexicana Ortodoncia 5:e184–e189. https://doi.org/10.1016/j.rmo.2017.12.018
Marsh W, Bello S (2023) Cannibalism and burial in the late Upper Palaeolithic: Combining archaeological and genetic evidence. Quat Sci Rev 319. https://doi.org/10.1016/j.quascirev.2023.108309
Martins JN, Mata A, Marques D, Anderson C, Caramês J (2016) Prevalence and Characteristics of the Maxillary C-shaped Molar. J Endodontics 42(3):383–389. https://doi.org/10.1016/j.joen.2015.12.013
Martínez-Moreno J, Mora R (2009) Balma Guilanyà (Prepirineo de Lleida) y el Aziliense en el noreste de la Península Ibérica. Trab Prehist 66:45–60. https://doi.org/10.3989/tp.2009.09021
Martínez-Moreno J, Mora R (2011) In the kingdom of ibex: continuities and discontinuities in Late Glacial hunter-gatherer lifeways at Guilanyà (south-eastern Pyrenees). Palethnology 3:211–227. https://doi.org/10.4000/palethnologie.7641
Martínez-Moreno J, Mora R, Casanova J (2006) Balma Guilanyà y la ocupación de la vertiente sur del Prepirineo del Noreste de la Península Ibérica durante el Tardiglaciar. In: Sanchidrián JL (ed) La cuenca Mediterránea durante el Paleolítico Superior. IV Simposio de Prehistoria Cueva de Nerja. Fundación Cueva de Nerja, Nerja, pp 444–457
Martínez-Moreno J, Mora R, Casanova J (2010) Lost in the mountains?: marine ornaments in the Mesolithic of the northeast of the Iberian Peninsula. In: Álvarez-Fernández E, Carvajal D (eds) Not only food, vol 31. Munibe Suplemento, pp 100–109
Meiklejohn C (2009) Radiocarbon dating of Mesolithic human remains in Spain. Mesolithic Miscellany 20(2):2–20
Mercedes Gallas M, García A (2000) Retention of permanent incisors by mesiodens: a family affair. Br Dent J 188:63–64. https://doi.org/10.1038/sj.bdj.4800390a
Milosevic A, Young PJ, Lennon MA (1994) The prevalence of tooth wear in 14-year-old school children in Liverpool. Community Dent Health 11(2):83–86
Moggi-Cecchi J, Pacciani E, Pinto-Cisternas J (1994) Enamel hypoplasia and age at weaning in 19th-century Florence, Italy. Am J Phys Anthropol 93(3):299–306. https://doi.org/10.1002/ajpa.1330930303
Moorrees CF, Fanning EA, Hunt EE (1963) Formation and resorption of three deciduous teeth in children. Am J Phys Anthropol 21:205–213. https://doi.org/10.1002/ajpa.1330210212
Morfis AS (1993) Chemical analysis of a dens invaginatus by SEM microanalyses. J Clin Pediatr Dent 17:79–82
Nelson GC, Lukacs JR, Yule P (1999) Dates, caries, and early tooth loss during the Iron Age of Oman. Am J Phys Anthropol 108:333–343
Nelson SJ (2020) Wheeler's Dental Anatomy, Physiology and Occlusion. Elsevier, Barcelona
Neves WA, Cornero S (1997) What did South American paleoinidans eat? Curr Res Pleistocene 14:93–96
Nikiforouk G (1985) Understanding dental caries. Karger, New York
Oehlers FA (1957) Dens invaginatus (dilated composite odontome). I. Variations of the invagination process and associated anterior crown forms. Oral Surg Oral Med Oral Pathol 10(11):1204–1218. https://doi.org/10.1016/0030-4220(57)90077-4
Oelze VM, Siebert A, Nicklisch N, Meller H, Dresely V, Alt KW (2011) Early Neolithic diet and animal husbandry: stable isotope evidence from three Linearbandkeramik (LBK) sites in Central Germany. J Archaeol Sci 38(2):270–279. https://doi.org/10.1016/j.jas.2010.08.027
Orschiedt J (2013) Bodies, bits and pieces: Burials from the Magdalenian and the Late Palaeolithic. In: Pastoors A, Aufferman B (eds) Pleistocene Foragers: their culture and environment. Wissenschftliche Schriften des Neanderthal Museums 6, Mettmann, pp 117–132
Orschiedt J (2018) The Late Upper Palaeolithic and earliest Mesolithic evidence of burials in Europe. Philos Trans R Soc 373. https://doi.org/10.1098/rstb.2017.0264
Oxilia G, Fiorillo F, Boschin F, Boaretto E, Apicella SA, Matteucci C, Panetta D, Pistocchi R, Guerrini F, Margherita C, Andretta M, Sorrentino R, Boschian G, Arrighi S, Dori I, Mancuso G, Crezzini J, Riga A, Serrangeli MC et al (2017) The dawn of dentistry in the late upper Paleolithic: An early case of pathological intervention at Riparo Fredian. Am J Phys Anthropol 163:446–461. https://doi.org/10.1002/ajpa.23216
Oxilia G, Peresani M, Romandini MC, Spiteri CD, Henry AG, Schulz D, Archer W, Crezzini J, Boschin F, Boscato P, Jaouen K, Dogandzic T, Broglio A, Moggi-Cecchi J, Fiorenza L, Hublin J-J, Kullmer O, Benazzi S (2015) Earliest evidence of dental caries manipulation in the Late Upper Palaeolithic. Sci Rep 5:12150. https://doi.org/10.1038/srep12150
Pala M, Olivieri A, Achilli A, Accetturo M, Metspalu E, Reidla M, Tamm E, Karmin M, Reisberg T, Hooshiar Kashani B, Perego UA, Carossa V, Gandini F, Pereira JB, Soares P, Angerhofer N, Rychkov S, Al-Zahery N, Carelli V et al (2012) Mitochondrial DNA signals of late glacial recolonization of Europe from near eastern refugia. Am J Hum Genet 90:915–924. https://doi.org/10.1016/j.ajhg.2012.04.003
Pettitt P (2011) The Palaeolithic Origins of Human Burial. Routledge, London. https://doi.org/10.4324/9780203813300
Peyroteo R (2016) Roots of death: origins of human burial and the research on Early Holocene mortuary practices in the Iberian Peninsula. In: Grünberg J, Gramsch B, Larsson L, Orschiedt J, Meller H (eds) Mesolithic burials -Rites, symbols and social organisation of early postglacial communities, pp 629–654
Pezo L, Eggers S (2010) The Usefulness of Caries Frequency, Depth, and Location in Determining Cariogenicity and Past Subsistence: A Test on Early and Later Agriculturalists from the Peruvian Coast. Am J Phys Anthropol 143:75–91. https://doi.org/10.1002/ajpa.21296
Pinhasi R, Teschler-Nicola M, Prossinger H, Shaw P (2006) Diachronic trends in dental dimensions of Late Pleistocene and Early Holocene European and Near Eastern Populations, vol 4. Biennial Books of European Anthropological Association, pp 1–18
Piperno D, Weiss E, Holst I, Nadel D (2004) Processing of wild cereal grains in the Upper Palaeolithic revealed by starch grain analysis. Nature 430:670–673. https://doi.org/10.1038/nature02734
Posth C, Yu H, Ghalichi A, Rougier H, Crevecoeur I, Huang Y, Ringbauer H, Rohrlach AB, Nägele K, Villalba-Mouco V, Radzeviciute R, Ferraz T, Stoessel A, Tukhbatova R, Drucker DG, Lari M, Modi A, Vai S, Saupe T et al (2023) Palaeogenomics of Upper Palaeolithic to Neolithic European hunter-gatherers. Nature 615(7950):117–126. https://doi.org/10.1038/s41586-023-05726-0
Rajab LD, Hamdan MA (2002) Supernumerary teeth: review of the literature and a survey of 152 cases. Int J Paediatr Dent 12:244–254. https://doi.org/10.1046/j.1365-263X.2002.00366.x
Rasmussen S, Bigler M, Blockley SP, Blunier T, Buchardt SL, Clausen HB, Cvijanovic I, Dahl-Jensen D, Johnsen SJ, Fischer H, Gkinis V, Guillevic M, Hoek WZ, Lowe JJ, Pedro JB, Popp T, Seierstad IK, Steffensen JP, Svensson AM et al (2014) A stratigraphic framework for abrupt climatic changes during the Last Glacial period based on three synchronized Greenland ice-core records: refining and extending the INTIMATE event stratigraphy. Quat Sci Rev 106:14–28. https://doi.org/10.1016/j.quascirev.2014.09.007
Reimer PJ, Austin WEN, Bard E, Bayliss A, Blackwell PG, Bronk Ramsey C, Butzin M, Cheng H, Edwards RL, Friedrich M, Grootes PM, Guilderson TP, Hajdas I, Heaton TJ, Hogg AG, Hughen KA, Kromer B, Manning SW, Muscheler R et al (2020) The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0-55 cal kBP). Radiocarbon 62:725–757. https://doi.org/10.1017/RDC.2020.41
Revedin A, Aranguren B, Becattini R, Longo L, Marconi E, Lippi MM, Skakun N, Sinitsyn A, Spiridonova E, Svoboda J (2010) Thirty-thousand-year-old evidence of plant food processing. Proc Natl Acad Sci 107(44):18815–18819. https://doi.org/10.1073/pnas.1006993107
Rose JC (1977) Defective enamel histology of prehistoric teeth from Illinois. Am J Phys Anthropol 46:439–446
Ruíz J, García C, Martínez-Moreno J, Subirà E (2005) Los restos humanos del Tardiglaciar de Balma Guilanyà. In: Sanchidrián JL, Márquez AM, Fullola JM (eds) La cuenca Mediterránea durante el Paleolítico Superior. IV Simposio de Prehistoria Cueva de Nerja. Fundación Cueva de Nerja, Nerja, pp 458–467
Rull V, Blasco A, Calero MÁ, Blaauw M, Vegas-Vilarrúbia TA (2023) Continuous Centennial Late Glacial-Early Holocene (15–10 cal kyr BP) Palynological Record from the Iberian Pyrenees and Regional Comparisons. Plants 12(20):3644. https://doi.org/10.3390/plants12203644
Ruprecht A, Batniji S, Sastry KA, el-Neweihi E, (1986) The incidence of dental invagination. J Pedod 10:265–272
Savage RR, Kokich VG (2002) Restoration and retention of maxillary anteriors with severe root resorption. J Am Dent Assoc 133:67–71. https://doi.org/10.14219/jada.archive.2002.0023
Sedano HO, Gorlin RJ (1969) Familial occurrence of mesiodens. Oral Surg Oral Med Oral Pathol 27:360–361. https://doi.org/10.1016/0030-4220(69)90366-1
Seif T (1997) Cardiología: prevención, diagnóstico y tratamiento contemporáneo de la caries dental. Actualidades Médico Odontológicas Latinoamérica, Bogotá
Sidow SJ, West LA, Liewehr FR, Loushine RJ (2000) Root canal morphology of human maxillary and mandibular third molars. J Endod 26(11):675–8. https://doi.org/10.1097/00004770-200011000-00011
Shifman A, Tamir A (1979) Dens invaginatus with concrescent supernumerary tooth. Oral Surg Oral Med Oral Pathol 47:391. https://doi.org/10.1016/0030-4220(79)90266-4
Skinner M, Hadaway W, Dickie J (1994) Effects of ethnicity and birth month on localized enamel hypoplasia of the primary canine. J Dent Child 61:109–113
Skinner MF (1986) An enigmatic hypoplastic defect of the deciduous canine. Am J Phys Anthropol 69:59–69. https://doi.org/10.1002/ajpa.1330690108
Skinner MF (1996) Developmental stress in immature hominines from Late Pleistocene Eurasia: evidence from enamel hypoplasia. J Archaeol Sci 23:833–852. https://doi.org/10.1006/jasc.1996.0078
Soares P, Achilli A, Semino O, Davies W, Macaulay V, Bandelt H-J, Torroni A, Richards MB (2010) The archaeogenetics of Europe. Curr Biol 20(4):R174–R183. https://doi.org/10.1016/j.cub.2009.11.054
Stephensen CB, Alvarez JO, Kohatsu J, Hardmeier R, Kennedy JI, Gammon RB (1994) Vitamin A is excreted in the urine during acute infection. Am J Clin Nutr 60:388–392. https://doi.org/10.1093/ajcn/60.3.388
Tallavaara M, Luoto M, Korhonen N, Järvinen H, Seppä H (2015) Human population dynamics in Europe over the Last Glacial Maximum. Proc Natl Acad Sci 112:8232–8237. https://doi.org/10.1073/pnas.1503784112
Tanaka E, Kawazoe A, Nakamura S, Ito G, Hirose N, Tanne Y, Kawai N, Tanimoto K, Tanne K (2008) An adolescent patient with multiple impacted teeth. Angle Orthod 78:1110–1118. https://doi.org/10.2319/121007-581.1
Terradas X, Pallarés M, Mora R, Martínez J (1993) Estudi preliminar de les ocupacions humanes de la balma Guilanyà. Rev d'Arqueol Ponent 3:231–248
Torroni A, Bandelt HJ, Macaulay V, Richards M, Cruciani F, Rengo C, Martinez-Cabrera V, Villems R, Kivisild T, Metspalu E, Parik J, Tolk HV, Tambets K, Forster P, Karger B, Francalacci P, Rudan P, Janicijevic B, Rickards O et al (2001) A signal, from human mtDNA, of postglacial recolonization in Europe. Am J Hum Genet 69(4):844–852. https://doi.org/10.1086/323485
Trinkaus E (2018) An abundance of developmental anomalies and abnormalities in Pleistocene people. Proc Natl Acad Sci 115(47):11941–11946. https://doi.org/10.1073/pnas.1814989115
Trinkaus E, Hillson SW, Franciscus RG, Holliday TW (2006) Skeletal and dental paleopathology. In: Trinkaus E, Svoboda J (eds) Early modern human evolution in Central Europe: the people of Dolní Věstonice and Pavlov. Oxford University Press, Oxford, pp 419–458
van Palenstein W, Matee M, van der Hoeven J, Mikx F (1996) Cariogenicity Depends More on Diet than the Prevailing Mutans Streptococcal Specie. J Dent Res 75(1):535–545. https://doi.org/10.1177/00220345960750010501
Villalba-Mouco V, Loosdrecht MS, Posth C, Mora R, Martínez-Moreno J, Rojo-Guerra M, Salazar-García DC, Royo-Guillén JI, Kunst M, Rougier H, Crevecoeur I, Arcusa-Magallón H, Tejedor-Rodríguez C, García-Martínez de Lagrán I, Garrido-Pena R, Alt KW, Jeong C, Schiffels S, Utrilla P et al (2019) Survival of Late Pleistocene Hunter-Gatherer Ancestry in the Iberian Peninsula. Curr Biol 29(7):1169–1177.e7. https://doi.org/10.1016/j.cub.2019.02.006
Villalba-Mouco V, van de Loosdrecht MS, Rohrlach B, Fewlass H, Talamo S, Yu H, Aron F, Lalueza-Fox C, Cabello L, Cantalejo Duarte P, Ramos-Muñoz J, Posth C, Krause J, Weniger GC, Haak W (2023) A 23,000-year-old southern Iberian individual links human groups that lived in Western Europe before and after the Last Glacial Maximum. Nat Ecol Evol 7(4):597–609. https://doi.org/10.1038/s41559-023-01987-0
Voisin J-L, Condemi S, Wolpoff MH, Frayer DW (2012) A new online database (http://anthropologicaldata.free.fr) and a short reflection about the productive use of compiling internet data. PaleoAnthropology 241–244. Accessed 4 Mar 2023
Weber M-J, Grimm S, Baales M (2011) Between warm and cold: Impact of the Younger Dryas on human behaviour in Central Europe. Quat Int 242:277–301. https://doi.org/10.1016/j.quaint.2010.12.002
Whyman RA, MacFadye EE (1994) Dens in dente associated with infective endocarditis. Oral Surg Oral Med Oral Pathol 78:47–50. https://doi.org/10.1016/0030-4220(94)90116-3
Zoubov AA (1968) Odontology: methods in anthropological research. Nauka Press, Moscow (in Russian)
Acknowledgements
We wish to acknowledge the fieldwork was supported by the Government of Catalonia’s Archaeology Service. We appreciate the facilities provided by the Laboratory for the Non-Destructive Analysis of Geological Materials-CORELAB (Barcelona University). We also thank the Guilanyà family for their kind permission to give access to the site. We would like to extend our gratitude to the anonymous reviewers whose insightful comments and constructive feedback greatly contributed to enhance the quality and clarity of our work.
Funding
Open Access Funding provided by Universitat Autonoma de Barcelona. Balma Guilanyà is part of the project Human settlement during the Upper Pleistocene and Holocene in the south-eastern Pyrenees recognised by the Spanish Ministry of Science and Innovation, Spanish Government (PID 2019-104843GB-I00, PID2022-136363NB-I00) and the Government of Catalonia’s Agency for Management of University and Research Grants (2021 SGR 00190).
Author information
Authors and Affiliations
Contributions
S.A. and J.M. wrote the main manuscript text and prepared the figures and table, along with the supplementary material. J.L. analysed the material (teeth). All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information

ESM 1
(PNG 204 kb)

ESM 2
(PNG 265 kb)

ESM 3
(PNG 135 kb)

ESM 4
(PNG 155 kb)

ESM 5
(PNG 722 kb)

ESM 6
(PNG 194 kb)

ESM 7
(PNG 1218 kb)

ESM 8
(PNG 52 kb)

ESM 9
(PNG 58 kb)

ESM 10
(PNG 145 kb)

ESM 11
(PNG 1028 kb)

ESM 12
(PNG 214 kb)

ESM 13
(PNG 961 kb)

ESM 14
(PNG 240 kb)

ESM 15
(PNG 246 kb)

ESM 16
(PNG 879 kb)

ESM 17
(PNG 427 kb)

ESM 18
(PNG 1352 kb)

ESM 19
(PNG 331 kb)

ESM 20
(PNG 348 kb)

ESM 21
(PNG 1307 kb)

ESM 22
(PNG 329 kb)

ESM 23
(PNG 1212 kb)

ESM 24
(PNG 307 kb)

ESM 25
(PNG 1103 kb)

ESM 26
(PNG 51 kb)

ESM 27
(PNG 1208 kb)

ESM 28
(PNG 63 kb)

ESM 29
(PNG 1430 kb)

ESM 30
(PNG 44 kb)

ESM 31
(PNG 55 kb)

ESM 32
(PNG 59 kb)

ESM 33
(PNG 66 kb)

ESM 34
(PNG 59 kb)

ESM 35
(PNG 75 kb)

ESM 36
(PNG 70 kb)

ESM 37
(PNG 69 kb)

ESM 38
(PNG 68 kb)

ESM 39
(PNG 61 kb)

ESM 40
(PNG 54 kb)

ESM 41
(PNG 48 kb)
ESM 42
(XLSX 85 kb)
ESM 43
(XLSX 12 kb)
ESM 44
(XLSX 10 kb)
ESM 45
(XLSX 11 kb)
ESM 46
(DOCX 78 kb)
(MP4 11562 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Arenas del Amo, S., López, J.L., Martínez-Moreno, J. et al. Odontological, pathological and contextual patterns of the Late Glacial human tooth assemblage from Level E at Balma Guilanyà (south-eastern Pyrenees, Iberian Peninsula). Archaeol Anthropol Sci 16, 147 (2024). https://doi.org/10.1007/s12520-024-02021-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-024-02021-5