
Abstract
Recent research on human exploitation of molluscs, echinoderms and crustaceans during the Mesolithic in the Cantabrian region (northern Iberia) has shown significant variability in the intensity of the use of coastal resources by the last hunter-fisher-gatherers throughout the ~ 4000-year expanse of the Mesolithic (10,700 – 6,700 cal BP). Previous studies have proposed hypotheses related to demographic changes to explain intensification events. In this paper we aim to unravel whether climatic and environmental changes, or other social factors, such as demography, were involved in the increase in the use of marine resources at certain times. We employed species representation, quantification and biometric analysis of the shell assemblages from the El Toral III archaeological site (Asturias, northern Iberia) to identify patterns in shell exploitation throughout the stratigraphic sequence. To establish the chronology and distinguish occupation phases of the site, we employed Bayesian modelling of radiocarbon measurements. Faunal results show that marine gastropods such as limpets of the Patella genus and topshells Phorcus lineatus (da Costa, 1778) were the most exploited species, while bivalves, echinoids and crustaceans were present in smaller quantities. The comparison of the results for El Toral III with other Mesolithic sites in the region reveals significant temporal differences in the intensity of collection of marine shell species. Results showed that intensification processes are not clearly related to climate change but to other social factors, such as demography, suggesting that intensification and relaxation in shell collection events were strategies adopted depending on the needs of the human groups at different times.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Archaeological and faunal evidence suggest that from the Upper Palaeolithic to the Mesolithic, there was an intensification in the exploitation of marine resources (Álvarez-Fernández 2009; 2013a; Cuenca-Solana et al. 2013; Gutiérrez-Zugasti 2009; Gutiérrez-Zugasti et al. 2013; García-Escárzaga 2020). During the Mesolithic, the number and size of shell middens, archaeological sites characterised by the predominance of shells in the stratigraphic matrix, increased across the coasts of Atlantic Europe (Milner et al. 2007; Gutiérrez-Zugasti et al. 2011), suggesting an intensive use of marine resources. In the Cantabrian region (northern Iberia), these deposits are commonly found in caves and rock shelters located less than 5 km from the modern coastline (Bailey and Craighead 2003; Gutiérrez-Zugasti et al. 2011; Fano 2019). The quantity and quality of the archaeological record makes this region a key area for the study of shell middens and long-term changes in subsistence strategies.
Studies of Mesolithic sites in the Cantabrian region started at the beginning of the 20th century with the excavations of shell-midden sites, primarily in eastern Asturias, by the Count of Vega del Sella, from which the Asturian culture was defined (Vega del Sella 1923). The number of studies rose during the 1970s and 1980s with the excavation of sites such as La Riera, Balmori, Mazaculos II, and La Llana (González-Morales 1982; Clark 1983; Straus and Clark 1986). Towards the end of the 20th century and the beginning of the 21st century, new excavations of Mesolithic sites were carried out in Cantabria, at sites such as La Garma A (Arias et al. 2000), La Fragua (González-Morales 2000), and El Perro (González-Morales and Díaz Casado 2000), and in Asturias at sites such as Poza l'Egua (Arias et al. 2007), El Alloru (Arias et al. 2015), El Toral III (Noval-Fonseca 2013) and El Mazo (Gutiérrez-Zugasti et al. 2013). Excavations at the archaeological sites of El Mazo and El Toral III revealed the presence of exceptionally well-preserved Mesolithic shell middens which offered opportunities for the study of site formation processes, the exploitation of marine and terrestrial food resources (Marín-Arroyo et al. 2020; García-Escárzaga and Gutiérrez-Zugasti 2021), technological and symbolic developments (Rigaud and Gutiérrez-Zugasti 2016; Fuertes-Prieto et al. 2021) and palaeoenvironments (García-Escárzaga et al. 2022a).
The Mesolithic in northern Iberia ranged from 10,700 cal BP to 6,700 cal BP. Holocene enviromental conditions in coastal areas were characterised by increased temperatures but also by rapid abrupt cold climatic events, such as the 8.2 ka cal BP event, identified in the region at the site of El Mazo by decreased seawater temperatures and tree pollen reduction (Núñez de la Fuente 2018; García-Escárzaga et al. 2022a; b). Higher precipitation and humidity (Yanes et al. 2012), sea level rise throughout the period, with stabilisation of the coastline around 7.5-7 ka cal BP ago (Leorri et al. 2012) and the presence of forests predominantly composed by oaks, hazelnuts and birches (Núñez de la Fuente 2018) have also been recorded in the region. From a cultural point of view, the Mesolithic in the Western part of the region has been traditionally related to the Asturian culture. Lithic technology at this time was characterised by the predominant use of local raw materials for flake and laminar (blades and bladelets) production. Geometric microliths, although scarce, are also present in the archaeological record from nearly 8.3 ka cal BP ago (Fuertes Prieto et al. 2021). The most representative lithic tool is the Asturian pick, a quartzite cobble-made artefact knapped unifacially to form a pointed distal end (González-Morales 1982). Antler and bone tools are very rare, but fishing hooks and perforated antler batons have been found at some sites. Subsistence strategies were characterised by intense use of marine resources, especially mollucs (García-Escárzaga et al. 2022a; b), hunting of terrestrial mammals (Andreu-Alarcón 2013) and collection of plants (López-Dóriga 2016). The presence of local lithic raw materials and differences between coastal and inland sites have been interpreted as evidence of reduced mobility (Fano 2019; Arias et al. 2021).
Regarding intensification of coastal resource use, the first faunal studies in the Cantabrian region focused on changes in the distribution of shell sizes. Ortea (1986) showed a temporal decrease in shell size across the stratigraphic sequence at La Riera (Upper Palaeolithic and Mesolithic), which was attributed to human pressure. Bailey and Craighead (2003) revised Ortea's interpretation and proposed instead that changes in shell size resulted from climatic and environmental factors. Subsequently, Gutiérrez-Zugasti (2009; 2011a) corroborated the intensification hypothesis by performing statistical analysis of size distributions, size selection, species abundance and collection areas of Upper Palaeolithic and Mesolithic sites. More recent studies, such as those developed in the Asturian Mesolithic shell midden site of El Mazo (García-Escárzaga et al. 2022a), dated between c. 9,000 to 7, 600 cal BP, showed that the intensity of exploitation of marine resources varied significantly across the Mesolithic. However, the chronology of El Mazo does not cover the entire Mesolithic so data from other sites is still required to analyse exploitation patterns in the long term.
Here we present the results of a new archaeomalacological study for the shell-midden site of El Toral III. We employed Bayesian modelling of radiocarbon measurements to refine the chronology of the site. This allowed us to determine temporal variations in resource exploitation patterns and phases of intensification. We combined the data from El Toral III with other shell-midden sites, including El Mazo, to offer a broader temporal and spatial analysis of resource exploitation in the Cantabrian region during the Mesolithic.
El Toral III: description and stratigraphy
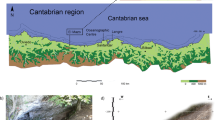
The El Toral III cave is located approximately 1 km from the modern coastline in the vicinity of the town of San Roque del Acebal within the municipality of Llanes (Asturias, Northern Spain) (Fig. 1). The cave is part of the Toral-Pellunarices karstic system, an active water circulation system consisting of more than 2.5 km of galleries.

Location of the study region and the shell midden site of El Toral III (Asturias, Northern Spain). MASL: meters above sea level
El Toral III consists of a wide rock shelter with a length of approximately 8 m (Fig. 2a) and a small cave with a chasm located at about 10 m from the entrance. The chasm connects the cave with an underground river that runs through the entire system. The archaeological deposit at El Toral III is located in the rock shelter and extensive salvage excavations were carried out in this area in 2009. Modern standard excavation procedures were used. The surface of the site was divided into 1 x 1 m squares, identified with a letter and a number (Fig. 2b), and those in four 0.5 x 0.5 subsquares (named from A to D). Stratigraphic units were individualised considering their composition and sedimentary features. Excavations revealed stratigraphic units informing on site formation and post-depositional processes (Noval-Fonseca 2013). Archaeological deposits are located in two separate areas (areas A and B). Area A, located in the south of the rock shelter, was impacted by water circulation and suffered heavy erosion and mixing of archaeological deposits (Supplementary Information 1A). Within this area, only stratigraphic unit 10 retained in situ intact human deposits (Fig. 2c). Unit 10 is stratigraphically the oldest archaeological unit of the stratigraphic sequence and is situated between two layers of sterile clay (units 9 and 11). Due to its relatively small size and the scarcity of archaeological materials found within, it is assumed that it was formed during a brief period of human occupation. Unit 13 lies just above unit 9 and consists of three subunits (13A, 13B and 13C) showing evidence for disturbances and the mixing of materials from different periods. Other stratigraphic units observed in area A were excluded from further study due to heavy erosion (e.g. units 4, 5 and 6) or because they were cemented (e.g. unit 8).

a View of the rockshelter. b Map of the excavation areas. c Area A of the excavation, vertical sequence with unit 13. d Area B of the excavation, vertical sequence with units 17-19. e Area B of the excavation, vertical sequence with units 20-22
Area B is situated in the northern part of the rock shelter and has a well-preserved stratigraphic sequence. For this area, units 17 and 19 show exceptional preservation (Fig. 2d), while units 20, 21 and 22 (Fig. 2e) have some features that indicate slight disturbances of the original deposits. For example, big stone blocks fell from the walls and roof of the rock shelter during the formation of units 21 and 22, altering in some way these deposits. The space between areas A and B was heavily eroded, and archaeological deposits have almost disappeared. Mesolithic shell-midden stratigraphic units contained a variety of materials, including mollusc shells, mammal bones, fish and bird bones, lithics, charcoal, plant seeds, and human remains. Radiocarbon dates and the typology of material culture indicate that the site was used during long periods of the Mesolithic. Mesolithic post-holes were identified in Area A (Noval-Fonseca 2013). The cave was used for human burial during the Chalcolithic and Bronze Ages (Estalrrich et al. 2019).
Materials and methods
Radiocarbon dating and Bayesian modelling
Bayesian chronological modelling of El Toral III stratigraphic sequences in areas A and B relied on a total of 36 radiocarbon dates (Supplementary Information 2A). This included eight previously published dates by Noval-Fonseca (2013), Rigaud and Gutiérrez-Zugasti (2016) and Allentoft et al. (2024), measured at the Center for Applied Isotope Studies of the University of Georgia (UGAMS), the International Chemical Analysis (ICA) and 14Chrono Centre of Queen’s University Belfast, and 28 new radiocarbon dates measured at the International Chemical Analysis (ICA). Dated radiocarbon samples included a mix of marine shells, plant charcoal, and herbivore and human bone samples. Collagen was extracted from bone samples for dating. Pre-treatment protocols, combustion, reduction, AMS measurement, and data analysis followed established methods (Brock et al. 2010).
We employed two separate Bayesian chronological models for areas A and B. These were run using the software OxCal v.4.4 (Ramsey 2001) and radiocarbon calibrations were done using the terrestrial IntCal curve for the northern hemisphere (Reimer et al. 2020) and the Marine20 curve (Heaton et al. 2020) for marine samples. Following the estimates of marine ∆R values for northern Iberia during the Early-Middle Holocene by García-Escárzaga et al. (2022b), a ∆R value of -134±49 yrs was applied to Patella vulgata Linnaeus, 1758 shells and -288±34 yrs to Phorcus lineatus (da Costa, 1778) shells. In the case of a single human bone collagen sample (UBA-40600), we employed a ∆R value of -186±163 yrs with a mixed terrestrial and marine curve. This ∆R value is a conservative estimate of mixed diet that may include P. vulgata and P. lineatus protein together with other marine species. To estimate the contribution from marine dietary carbon toward UBA-40600 we employed a model with two end-member references for carbon stable isotope ratios in human bone collagen under a fully terrestrial diet (δ13Cterrestrial= -20.5± 1‰) and a fully marine diet (δ13Cmarine= -7.8 ± 1‰) following Dewar and Pfeiffer (2010). Uncertainties for human bone collagen δ13C values were set conservatively at 0.5‰. Bayesian dietary estimates were done using the software ReSources, an upgraded version of the FRUITS software (Fernandes et al. 2014; Sołtysiak and Fernandes 2021). Outlier OxCal models were employed using a Student’s t distribution with five degrees of freedom (Ramsey 2009). Prior probabilities that radiocarbon dates were outliers was set at 5%. The OxCal model codes are given in Supplementary Information 1B.
Species representation
We studied the shell assemblage composed of molluscs, echinoderms and crustaceans from nine stratigraphic units in areas A and B. Materials from 331 litres of sediment were analysed. The archaeological remains were washed and screened using 4, 2 and 1-mm meshes. The materials that passed the 4 and 2 mm meshes were analysed, while those that passed the 1 mm mesh were not studied. The analysed remains came from stratigraphic units 17 and 19 located in square O8 (subsquares A and B), units 20, 21 and 22 located in square M9 (subsquares A and B), unit 10 from square O4 (subsquare D) and units 13A, 13B and 13C from square O6 (subsquares C and D). Although partial studies on shells from units 17, 21 and 13A have been previously carried out (Bello-Alonso et al. 2015; Martínez-Cuesta 2017), we have studied those remains again.
We used established references (Poppe and Goto 1991; Palacios Eguren and Vega De La Torre 1997) and the reference collection from the International Institute for Prehistoric Research of Cantabria (IIIPC) for species identification. Employed nomenclature was according to the World Register of Marine Species (WoRMS, http://www.marinespecies.org/). We classified anatomically 353,797 shell remains. Shellfish remains were included in the fragmentation categories proposed by Moreno (1994) and Gutiérrez-Zugasti (2011a) (Table 1).
To quantify the distribution of mollusc, we employed the abundance estimators based in fragmentation categories developed by Moreno (1994) to determine the Minimum Number of Individuals (MNI) for bivalves and gastropods, and by Gutiérrez-Zugasti (2011c) for echinoids and crustaceans. The formula used for calculating the MNI for bivalves was as follows:
-
MNI bivalves= COMV + FRAV + FCH + (AHF or PHF, whichever is higher). Lateralized valves with higher results were assumed as MNI.
-
MNI spiralled gastropods= COMI + FRAI + [APIF or (STOF + UMBF), whichever is higher]
-
MNI non-spiralled gastropods= COMI + FRAI + APIF
For echinoids and crustaceans, an individual MNI for each type of diagnostic remain was calculated. For sea urchins, we used different formulas for pyramids and teeth. In the case of epyphisis, rotulae and compasses, only complete remains were counted to calculate the MNI. The highest value of the five diagnostic remains was taken as MNI:
-
MNI sea urchins semi-pyramid= COMSP + (SPAF or SPBF, whichever is higher)/5
-
MNI sea urchins tooth= COMT + (TAF or TBF, whichever is higher) /5
For goose barnacles, we calculated the MNI from the Carina, Tergum and Scutum. The highest value was taken as MNI:
-
MNI goose barnacles= CC + (AFC or BFT, whichever is higher).
-
MNI goose barnacles= CT + (AFT or BFT, whichever is higher).
-
MNI goose barnacles= CS + (AFS or BFS, whichever is higher).
In the case of crabs, we calculated the MNI for each fragmentation category separately and the highest value was taken as MNI.
We also calculated the fragmentation index to classify shell preservation status and assess taphonomic processes. We followed the methods proposed by Gutiérrez-Zugasti (2011a).
Biometric analysis
Biometric analysis was performed to determine shell size variations and assess possible shell size selection across time. Size measurements were taken using a digital calliper to the nearest 0.1 mm on well-preserved shell specimens. Measurements of Height/Width/Length were taken for limpets (Patella depressa Pennant, 1777, P. vulgata, Patella ulyssiponensis Gmelin, 1791) and diameter for topshells (P. lineatus) following the methodology proposed by Gutiérrez-Zugasti (2009).
Statistical analyses were carried out using the software PAST (Hammer et al. 2001). We did not perform these when sample numbers were lower than 30. Kruskall-Wallis and Mann-Whitney pairwise tests were used for size comparisons across stratigraphic units. Normality was tested using the Shapiro-Wilk test and from this infer size selection. The skewness of the distributions was also used for interpretation of size selection. To expand the chronological range of our study, previously published data from other Mesolithic shell midden sites close to the study area was considered. Thus, only sites with absolute dates and a detailed study of the archaeomalacological assemblages, such as El Mazo (García-Escárzaga 2020) and Mazaculos II (Gutiérrez-Zugasti 2011b), were considered for discussion.
Collection areas, such as intertidal zonation (lower or higher shores) and the degree of the coastal exposure (exposed or sheltered), were established using biometric data from P. vulgata. Zonation and exposure equations were as proposed by Bailey and Craighead (2003). Exposure was calculated following the equation:
For exposures under -0.15 limpets were assumed to originate from exposed shores, while higher values were assigned to sheltered shores. Zonation of P. vulgata was determined by calculating Length/Height ratios (L/H) from each specimen. Data were compared with the 2.55 threshold value proposed by Gutiérrez-Zugasti (2009) for northern Iberia. If the result exceeds the threshold, the specimen is assumed to originate from lower shores and reversely to higher shores. The vertical zonation was calculated for the limpet P. depressa based on the studies conducted by García-Escárzaga (2020) on modern populations of this taxon. In this case, the Width/Height (W/H) ratio was used, following the criteria of García-Escárzaga (2020). The coefficient threshold was 2.53. Similarly, if the ratio exceeded the coefficient, the individual was assigned to lower shores and reversely to higher shores.
Results
Bayesian chronological modelling
A total of 32 14C AMS measurements from marine and terrestrial archaeological samples were used for the final model. Four measurements were excluded based on the poor agreement they showed during the process. The references of the excluded dates are: ICA-14C-6004, ICA-14C-6005, ICA-14C/0802 and UGAMS-5400 (Supplementary Information 2A). The Bayesian modelled dates place the formation of the Mesolithic shell midden site in two different times of occupation according to the excavation areas. The chronological sequence of Area A reflects occupations from the Early Mesolithic. The modelled earliest date was 9,891-9,494 cal BP (95.4% probability) corresponded to the boundary start of unit 10 with an interval of duration between 0 and 221 years cal BP. The modelled latest date was 9,549-9,417 cal BP (95.4% probability) and corresponded to the boundary end of unit 13C with an interval between 0 and 184 years cal BP (Fig. 3a) (Supplementary Information 2B). The Bayesian model from Area B shows a later occupation of the Mesolithic period. Sequence 22-21-20 showed the modelled earliest date between 7,724-7,580 cal BP (95.4% probability) corresponding to the boundary start of unit 22 with an interval of 0 and 114 years cal BP. Meanwhile, the boundary end of unit 20 was modelled between 7,511-7,150 cal BP with an interval of duration from 0 to 340 years cal BP. On the other hand, sequence 19-17 was modelled the earliest date between 7,780-7,571 cal BP (95.4% probability) corresponding to the start of unit 19 with an interval of duration from 0 to 253 years cal BP. The end of unit 17, and the end of sequence that we assumed, was modelled between 7,388-6,656 cal BP (95.4% probability) with an interval of duration from 0 to 812 years cal BP (Fig. 3b) (Supplementary Information 2C).

a Modelled chronology for Area A. b Modelled chronology for Area B. Radiocarbon dates for terrestrial samples are grey coloured, and marine dates are in green. Colours outliers of Oxcal v.4.4 version are used
Species representation
Results on species representation show the presence of marine bivalves and gastropods, as well as echinoderms and crustaceans at El Toral III. In most of the stratigraphic units, four species (P. vulgata, P. depressa and P. ulyssiponensis, and P. lineatus) account for more than 80% of each shell assemblage (Supplementary Information 3). When the abundance of the main species used for food is observed, it is possible to see three different temporal patterns in taxa representation (Fig. 4).

a Densities (MNI per dm3) of the main gastropod species present at the shell midden site of El Toral III. Red (P. depressa), grey (P. ulyssiponensis), yellow (P. vulgata) and blue (P. lineatus) bars show density values for the different stratigraphic units. b Images of the most represented species. Each coloured box represents a different species according to the colours in the graph. Scale 1: cm
First, unit 10 in Area A consists almost exclusively of limpets (only a single P. lineatus was recorded), with P. vulgata and P. depressa dominating the assemblage. The three subunits from unit 13 are dominated by topshells P. lineatus, followed by limpets P. depressa and P. vulgata, except unit 13C where the three species show similar abundance. There is a progressive increase in topshells, paralleled by a decrease in limpets P. vulgata, from unit 13C to unit 13A. In the younger units (17 to 22), located in Area B, the assemblages are dominated by the limpet P. depressa followed by P. vulgata and P. ulyssiponensis. In these units, a considerably lower density of topshells compared to units from Area A (i.e., 13A, 13B and 13C), is observed.
Crustaceans, echinoderms and other mollusc species are present in negligible numbers. Most of these species were not used for food, except the mussels Mytilus galloprovincialis Lamarck, 1819, the sea urchins Paracentrotus lividus (Lamarck, 1816) and crustaceans, such as crabs of the Brachyura order and goose barnacles Pollicipes pollicipes (Gmelin, 1790). The stratigraphic unit with the highest density of these species is unit 17, the youngest in the Mesolithic deposit. Despite their low abundance, these species are present in all stratigraphic units of the sequence, except for unit 10.
Shells were impacted by similar taphonomic processes throughout the sequence. Biodegradation, burning and fragmentation were the most common processes. Some units, such as 17 and 19 have a higher degree of cementation since they are closer to the walls of the rock shelter, where water circulation is more intense. Limpets, the most represented taxa in the archaeological record, display significant fragmentation, especially in units 13C, 13B and 13A. Topshells are comparatively less fragmented, but they show the higher degree of fragmentation in the younger units of the sequence (unit 20 and especially 17) (Table 2).
Biometrical analysis
The length of 2,424 P. depressa shells from the different stratigraphic units was measured (Fig. 5a) (Supplementary Information 4). Limpet shells of this species from unit 10 and the subunits from level 13 (Area A), have larger average sizes (between 26 – 24.2 mm) than limpets from units 17 to 22 (Area B) having similar average sizes (21.4 – 20.7 mm). Average size shows a decreasing trend in the earliest part of the sequence, but they remain almost the same in the youngest part. A similar pattern is visible for the length size of P. vulgata (7,735 specimens measured) (Fig. 5b) (Supplementary Information 4). In this case, average size varies between 26.2 - 24.4 mm in Area A and 23.2 - 22.5 in Area B. Thus, the same size pattern is observed for both species, although the absolute magnitude of the variation is larger in P. depressa. For topshells P. lineatus, the diameter of 4,735 specimens was measured (Supplementary Information 4). In this case, there is a slight increase in average sizes at the start of the sequence and a subsequent decrease, which is especially evident in the younger units (Fig. 5c). A Kruskal-Wallis test showed significant differences among stratigraphic units in all species (Supplementary Information 5).

Box plot of size distributions throughout the El Toral III sequence for P. depressa a, P. vulgata b and P. lineatus c shells. Boxes represent the interquartile range and include mean (crosses) and median (horizontal line). Vertical lines are whiskers and black filled circles identify outliers
Mann-Whitney pairwise tests, revealed significant differences among most of the stratigraphic units (Supplementary Information 5), although with some species differences. For P. depressa and P. lineatus there were significant differences among most units while for P. vulgata there were significant differences among units from Area A, but not among most units of Area B. Intentional size selection was checked using statistical analysis (Supplementary Information 5). In the case of P. depressa, test results show mostly non-normal and non-log-normal size distributions and positive skewness, except for unit 10 (normal), units 13A and 19 (log-normal), and the negative skewness observed for unit 13C. A similar pattern is visible in P. vulgata where most distributions are non-normal and non-log-normal, with positive skewness. Only unit 13C shows a normal distribution, while units 13B and 13A show log-normal distributions. P. ulyssiponensis show mostly normal distributions and negative skewness in units from Area A, and non-normal (non-log-normal) distributions and positive skewness in Area B. For topshells P. lineatus most of the stratigraphic sequence consists of non-normal and non-log-normal distributions, except units 22 and 20, which show normal size distributions. Skewness is positive in units from Area A and mostly negative in units from Area B.
The study of zonation shows that around 60% of P. depressa and 80% of P. vulgata were collected in the lower shore (Fig. 6a and b), while the rest of the specimens were collected from the higher shore. Exposure analysis shows that the collection of P. vulgata took place almost exclusively in exposed areas (Fig. 6c).

a Percentages of limpet shells P. depressa from El Toral III collected in the lower (orange) and upper (red) intertidal zones. b Percentages of limpet shells P. vulgata from El Toral III collected in the lower (orange) and upper (red) intertidal zones. c Percentages of limpet shells P. vulgata from El Toral III collected in exposed (green) and sheltered (purple) zones
Discussion
Post-depositional processes in shell midden formation
In general, stratigraphic units from Area A have undergone important post-depositional processes, probably due to multiple water circulation episodes, although anthropogenic activities during more recent prehistoric and historic periods, cannot be discarded (Noval-Fonseca 2013). These disturbance episodes contributed to the erosion of most of the original archaeological deposits from Area A and the mixture of materials of different ages. Units 4, 5 and 6 are clear examples of this problem and, for that reason, they have not been analysed in this work (Supplementary Information 1A). At first inspection, units 13A, 13B and 13C appeared better preserved. However, the sedimentary matrix of these three units, similar for all, consists of loose sediment and shells (unlike Area B units, whose matrices contain less sediment and are almost exclusively composed of shells). They contained abundant decimetric limestones, fallen from the walls and ceiling of the rock shelter. Such features suggest that these units have also undergone important post-depositional processes. This interpretation is also supported by the chronology of some materials recovered in units 13A, 13B and 13C, especially a human bone recovered in unit 13B that has been dated to the Early Bronze Age (Supplementary Information 2A). Moreover, the dates obtained from shells of P. vulgata and P. lineatus from unit 13C provided very different results. While the P. vulgata shells gave dates consistent with the stratigraphic position of the unit, the P. lineatus shells showed much more recent dates (they were rejected by the Bayesian model). This indicates that some (or all) shells of P. lineatus are not in their initial position and are intrusive.
The analysis of biometric data also supports the aforementioned interpretation, since the average size of P. lineatus from 13C is similar to that of unit 17 of Area B, while the sizes of the two species of limpets from 13C are larger than those of the units from Area B, and therefore it is probable that the remains of these species correspond to the actual chronology of the unit in which they were found. Moreover, an ungulate bone from unit 13A yielded a radiocarbon date consistent with the chronology of Area B (Supplementary Information 2A). However, despite these problems, the species representation of these units follows a very orderly pattern, with a predominance of P. lineatus followed by P. depressa. The same pattern has been observed in other sites, such as the rock shelter of El Mazo, in stratigraphic units with a chronology similar to that of the radiocarbon date obtained for unit 13A. Given such stratigraphic issues, we excluded these units from the discussion on temporal changes of species representation and biometric characteristics. These units exemplify the problems associated with the formation and erosion of shell middens, and the need to conduct investigations that combine the detailed study of stratigraphy, chronology and archaeological remains to identify alterations to original deposits.
Temporal changes in species representation
The study of mollusc shells from El Toral III shows a predominance of temperate-water species, such as the topshell P. lineatus and the limpets P. depressa. The species P. vulgata, traditionally considered a cold-water species, but capable of adapting to more temperate conditions (Poppe and Goto 1991), also appears in significant numbers. This pattern has been observed in other Mesolithic sites in the region, such as El Mazo, Mazaculos II, La Fragua and La Garma A, among others (Gutiérrez-Zugasti 2009; Álvarez-Fernández 2011; 2013b; Gutiérrez-Zugasti and Cuenca-Solana 2014; García-Escárzaga et al. 2022a). Temporal changes in species representation were investigated by combining data from El Toral III with that from the neighbouring site of El Mazo (Gutiérrez-Zugasti et al. 2013; García-Escárzaga et al. 2022a), located 1 km to the east from El Toral III. The oldest units of El Toral III range from 9,650 to 9,530 cal BP, while the sequence at El Mazo begins at 9,000 cal BP and ends at 7,635 cal BP. The most recent units of El Toral III range from 7,620 to 7,200 cal BP. Therefore, except for the 500 years between 9,500 and 9,000 cal BP, a period in which there is an information gap, the sequence of both sites covers more than 2,000 years of the Mesolithic period and makes it possible to trace the evolution of shellfish harvesting strategies in the region over an extended period (Fig. 7).

Densities of the minimum number of individuals per litre of excavated sediment for the most abundant gastropod species. Bars in red (P. depressa), grey (P. ulyssiponensis), yellow (P. vulgata) and blue (P. lineatus) show density values for the different stratigraphic units at the shell midden sites of El Toral III and El Mazo. Dashed lines through the graph indicate calibrated dates
The beginning of the sequence, dated at El Toral III around 9,650 cal BP (unit 10), shows a predominance of P. depressa and P. vulgata. Data originates from a small stratigraphic unit embedded between the base clays of the cave, having a low density of shells compared to other units. These characteristics suggest a short site occupation. After a gap of 500 years without data, the oldest unit of El Mazo shows a predominance of P. vulgata, with significant amounts of P. depressa and P. lineatus. Between 8,800 cal BP and 8,300 cal BP there is a predominance of P. lineatus, with significant amounts of P. vulgata and little abundance of P. depressa. During the 8.2 ka cal BP cold event P. vulgata dominates. Around 8 ka cal BP, following the 8.2 ka cold event, P. lineatus was once again the most abundant species. Around 7,620 cal BP, coinciding with the units located in Area B of El Toral III, there was a drastic change in species representation, with a clear predominance of P. depressa and a dramatic decrease of P. lineatus.
The study of molluscs from El Mazo reflects inversely proportional changes in the temporal abundance of P. vulgata and P. lineatus, due to their cold and temperate climatic preferences, respectively (García-Escárzaga et al. 2022a). The abundance of these species seems to follow the pattern of climatic variations visible in proxies such as δ18O from ice cores in Greenland (NGRIP members 2004), supporting the hypothesis that Mesolithic populations collected the most abundant species available on the intertidal areas. However, the case of P. depressa is more intriguing, since its abundance along the sequence does not follow climatic variations, as in the case of the other two species. At the beginning of the sequence its presence is scarce, and gradually increases until culminating in its predominance from 7,620 cal BP onwards, as observed in the most recent units of El Toral III. The species P. ulyssiponensis, with even more temperate preferences than P. depressa, follows a similar pattern although its densities are lower, probably because it lives in the lowest intertidal zone, and therefore, its collection was only possible during the most extreme spring tides, or perhaps because the presence of this species in the intertidal zone was reduced. The geographic distribution of these species shows that P. depressa and P. ulyssiponensis are species with clear temperate climatic preferences (Poppe and Goto 1991; Gutiérrez-Zugasti 2011b). Thus, it would be expected that these follow an abundance pattern like P. lineatus. However, this is not the case, which suggests that other factors, apart from seawater temperature, are influencing the presence of these species in the intertidal of the Cantabrian region. For example, increased coastal exposure, derived from sea level rise, could explain the abundance of P. depressa and P. ulyssiponensis, species better adapted to exposed shores than P. vulgata and P. lineatus. The position of the coastline in the region begins to stabilise around 7,500 - 7,000 cal BP (Leorri et al. 2012), coinciding with the time when these species predominate in the studied sequence. However, the results for collection areas shows that most of the P. vulgata specimens were obtained in exposed areas. This pattern is observed throughout the entire sequence, so exposed coasts seem to have been present in the region throughout the study period. Given this, it is difficult to determine the cause or causes of the different abundance patterns among the most abundant species. While in the case of P. lineatus and P. vulgata their abundance varies in relation to climatic changes, in the case of P. depressa and P. ulyssiponensis their abundance increases progressively across the sequence, independently of climatic changes.
Mesolithic shellfish exploitation intensification processes
The intensification of exploitation of marine resources during the Mesolithic has been previously suggested in various parts of the world, including in Western Europe during the Late Mesolithic (Maninno and Kenneth 2001). In the Mediterranean region, Colonese et al. (2011) also observed the intensification in the use of molluscs among Mesolithic hunter-gatherer communities. In Atlantic Europe, Gutiérrez-Zugasti (2011b) conducted an extensive review of Mesolithic shell midden sites in northern Iberia, identifying intensification processes related to human pressure and characterised by the intense exploitation of specific species, the collection in lower and exposed areas of the intertidal, and shell size and age decrease. Other areas of Atlantic Europe, such as Portugal, France, Denmark and the UK showed similar patterns of intensive shellfish collection (Gutiérrez-Zugasti et al. 2011). Similarly, beyond Europe, intensification has been observed on the West Coast of South Africa. Jerardino (2010) identified in Later Stone Age shell middens, processes of intensification in several species of limpets and black mussels around 2,600 BP, reflected in the reduction of mean sizes. Lastly, Hausmann et al. (2021) analysed size variation of a collection of over 15,000 specimens of Conomurex fasciatus (Born, 1778) from the Holocene shell mounds on the Farasan Islands (Saudi Arabia) in order to understand how intensification affected the resilience of molluscs to resource depletion between 7,360 cal BP and 4,780 cal BP. These examples show that intensification in the use of molluscs was a common strategy among hunter-gatherer communities.
Our results suggest that there was no gradual increase in shellfish exploitation during the Mesolithic in northern Iberia. The intensification on the exploitation of such resources was observed at certain time periods. Biometrics are here used to assess this process and the causes that lead human beings to adopt intensification strategies. For the temporal analysis of biometric features, we combined data from El Mazo, El Toral III, and the Mazaculos II shell midden (Gutiérrez-Zugasti and González-Morales 2010; Gutiérrez-Zugasti 2011b) allowing us to cover a wider chronological range (Fig. 8). During the interval 10,500 – 10,000 cal BP, the sizes of the three main species are the largest in the entire sequence, although a slight decrease is observed between the start and end of the interval. Around 9,000 cal BP a size decrease is observed, while between 9,000-8,300 cal BP the sizes are maintained or slightly smaller. Around 8,300 cal BP the sizes begin a downward trend, which has been related to an intensification event during the period corresponding to the 8.2 ka cal BP cold event. This intensification has been previously related to an increase in population in coastal areas due to the colder conditions of other neighbouring areas such as the Ebro Valley (García-Escárzaga et al. 2022a). From 8,050 cal BP, there is a shell size increase, suggesting a relaxation in reliance on marine resources. Around 7,800 cal BP, there was a new decrease in shell size, reflected in the most recent units of El Mazo, and in the units of Area B of El Toral III, which contain the smallest shell sizes of the entire sequence.

Chronological size variations from the Mesolithic shell midden sites of Mazaculos II (green), El Mazo (red) and El Toral III (blue). Measurements for P. depressa (a), P. vulgata (b) and P. lineatus (c). White circles represent the mean of the distributions and bars represent standard deviation. The grey colour line shows the trend and the grey shading shows the error using lm method
Information on demography during the Mesolithic in the region is limited. Some approaches to the population dynamics during the Holocene in the Cantabrian region has recently been proposed, based on the summed probability distributions of radiocarbon dates (Fernández-López de Pablo et al. 2019; Clark and Barton 2022; González-Rabanal 2022). These models suggest that during the 8.2 ka cal BP event, the population in the region remained stable or increased slightly, while shell sizes declined. Between 8,050 cal BP and 7,600 cal BP there was a population increase, in a period during which shell sizes increased first and then decreased, suggesting the existence of a relaxation on the reliance on molluscs followed by a new intensification event. After 7,600 cal BP there was a population decrease while shell sizes remained relatively stable.
Thus, according to these palaeodemographic models and data presented here, there is some correlation between a higher demographic pressure and a decrease in the size of molluscs. However, it is necessary to take these data with caution due to the limitations of the summed probability distributions of radiocarbon dates, which are influenced by aspects such as the intensity of the investigation or the archaeological visibility of sites and burials.
Except for the 8.2 ka cal BP event, changes in shell size do not seem to be directly correlated with significant climatic changes either. The three most abundant species, which have different climatic preferences, follow the same trend in terms of size across the stratigraphic sequence, which supports a cultural rather than climatic cause for size variations.
The analysis of collection areas (Fig. 6) shows a consistent pattern throughout the sequence, with a preference for the collection of limpets in lower and exposed intertidal areas. This pattern was previously observed in other Cantabrian Mesolithic sites, such as El Mazo, La Llana and Mazaculos II (Gutiérrez-Zugasti 2009; García-Escárzaga 2020; García-Escárzaga et al. 2022a) and related to the intensive use of shellfish, as lower and exposed areas remain less time emerged and are more dangerous for collection than higher and sheltered zones (Gutiérrez-Zugasti 2011a). This implies human knowledge of tide patterns, which made it possible to take advantage of the spring tides that occur every fortnight as the lower parts of the intertidal that are submerged during neap tides remain emerged during spring tides. Therefore, this pattern suggests organized collection strategies.
Most of the analysed archaeological units show non-normal or non-log normal shell size distributions, suggesting a size selection process as observed from field experiments (Gutiérrez-Zugasti 2009). The distribution of size data, which typically shows tails skewed towards larger sizes, as well as the positive skewness values, indicate that the larger sizes were targeted. The indiscriminate collection of all available sizes for a mollusc population could indicate collection intensification, as lower-yield specimens are also collected to satisfy the needs of the human groups. Under this assumption, size selection analysis does not show intensification features. However, the adoption of a strategy based on the collection of larger sizes could also be reflecting a certain intensification, by obtaining a higher meat yield with fewer shells collected. Collecting specimens with a higher meat yield involves harvesting females due to their larger dimensions, which, as previous studies in the region have indicated (Gutiérrez-Zugasti 2011b), may interfere with their reproductive capacity. This fact could subsequently impact size distributions if their numbers are lowered below a certain threshold. Therefore, selection of larger sizes could also be reflecting intensification. However, no temporal patterns through the Mesolithic are observed in this case, suggesting that collection of larger sizes was a strategy sustained through time.
Conclusions
Our study from the shell midden site of El Toral III showed that Mesolithic hunter-gatherers from northern Iberia had an organized and standardized strategy for shell collection. Larger sizes of limpets from lower and exposed shores were mostly targeted, as well as topshells collected in higher intertidal zones. Previous research on shell collection suggested that intensification in the use of molluscs was not gradual through the Mesolithic in northern Iberia, and that certain temporal variability occurred (García-Escárzaga et al. 2022a). Our data from El Toral III has provided new information on this issue, helping to increase the temporal range of the previous studies. Thus, here we relied on a data compilation from different shell midden sites to investigate the temporal intensity of shellfish harvesting in the area. Observed temporal variability could be related to climatic and environmental changes, demography, or choices related to human social organization. Despite de difficulties to unravel the causes of intensification, our results showed that intensification processes are not clearly related to climate change but to other social factors, such as demography, suggesting that intensification and relaxation in shell collection events were strategies adopted depending on the needs of the human groups at different times.
Data Availability
The data is available in the manuscript and in the supplementary information.
References
Allentoft ME, Sikora M, Refoyo-Martínez A, Irving-Pease EK, Fischer A, Barrie W, . . . Willerslev E (2024) Population genomics of post-glacial western Eurasia. Nature 625(7994):301-311. https://doi.org/10.1038/s41586-023-06865-0
Álvarez-Fernández E (2009) Análisis arqueomalacológico de la cueva de Altamira (Santillana del Mar, Cantabria): Excavaciones de J. González Echegaray y L.G. Freeman. Complutum 20:55–70
Álvarez-Fernández E (2011) Humans and marine resource interaction reappraised: Archaeofauna remains during the late Pleistocene and Holocene in Cantabrian Spain. Journal of Anthropological Archaeology 30:327–343. https://doi.org/10.1016/j.jaa.2011.05.005
Álvarez-Fernández E (2013a) La récolte des coquillages dans le región Cantabrique au Magdalénien: la grotte de <<Tito Bustillo>> (Asturies, Espagne). L’Anthropologie 117:62–93
Álvarez-Fernández E (2013b) Upper Pleistocene-Early Holocene Transition at La Garma A Cave (Omoño, Cantabria, Spain): Preliminary Report on the Marine Molluscs. In: Bailey GN, Hardy K, Camara A (eds) Shells Energy. Mollusc shells as coastal resources. Oxford: Oxbow Books: 167-181
Andreu Alarcón S (2013) La gestión de los recursos faunísticos en el Mesolítico del oriente de Asturias: Estudio arqueozoológico de los yacimientos de El Mazo y El Toral. Trabajo Fin de Máster Inédito, Universidad de Cantabria, Santander
Arias P, González-Sainz C, Moure JA, Ontañón R (2000) Estudio integral del Complejo Arqueológico de La Garma (Omoño, Ribamontán al Monte). In: Actuaciones Arqueológicas en Cantabria 1984-1999. Gobierno de Cantabria. Santander: 217-277
Arias P, Fernández-Tresguerres JA., Álvarez-Fernández E, Armendariz A, Cueto-Rapado M, Fano-Martínez MA, Fernández-García R, Garralda MD, Mensua C, Teira LC (2007) Excavación arqueológica de urgencia en la cueva de La Poza l’Egua (Lleídas, Llanes). In: Excavaciones Arqueológicas en Asturias 1999-2002. Consejería de Cultura, Comunicación Social y Turismo del Principado de Asturias, Oviedo: 227-239
Arias P, Cubas M, Fano M, Álvarez-Fernández E, Araújo A, Cueto M, Duarte C, Fernández-Sánchez P, Iriarte E, Jordá-Pardo J, López-Dóriga I, Núñez S, Salzmann C, Tapia J, Teichner F, Teira L, Uzquiano P, Vallejo J (2015) Une nouvelle aproche pour l’etude de l’habitat mésolithique dans le nord de la Péninsule Ibérique: Recherches dans le site de plein air d’El Alloru (Asturies, Espagne). Bulletin de la Société Préhistorique Francaise: 159-190
Arias P, Cubas M, Fano M, Álvarez-Fernández E, Araújo A, Cueto M, Fernández Sánchez P, Iriarte E, Teira L, Uzquiano P (2021) Looking for the ‘Asturian’ dwelling areas: New data from El Alloru and Sierra Plana de la Borbolla (Asturias, Spain). Foraging Assemblages 2: 169-177
Bailey GN, Craighead AS (2003) Late Pleistocene and Holocene coastal paleoeconomies. A reconsideration of the molluscan evidence from Northern Spain. Geoarchaeology :175-204
Bello-Alonso P, Ozkorta-Escribano L, Gutiérrez-Zugasti I (2015) Un acercamiento al aprovechamiento de los recursos litorales durante el Mesolítico: los invertebrados marinos del abrigo de El Toral III (Llanes, Asturias). In: La Investigación Arqueomalacológica en la Península Ibérica: Nuevas Aportaciones: 91-99. Nadir Ediciones. Eds. Igor Gutiérrez-Zugasti, David Cuenca Solana, Manuel González Morales
Brock F, Higham T, Ditchfield P, Bronk Ramsey C (2010) Current pretreatment methods for AMS radiocarbon dating at the Oxford radiocarbon accelator unit (Orau). Radiocarbon 52:103–112
Clark GA (1983) The Asturian of Cantabria. Early Holocene Hunter-Gatherers in Northern Spain. University of Arizona Press, 41
Clark GA, Barton CM (2022) The Mesolithic of Atlantic Coastal Spain – a comparison with the Middle Ebro Basin. Comptes Rendus Palevol 21(3):39–114
Colonese AC, Maninno MA, Bar-Yosef Mayer DE, Fa DA, Finlayson JC, Lubell D, Stiner MC (2011) Marine mollusc exploitation in Mediterranean prehistory: An overview. Quaternary International 239:86–103
Cuenca-Solana D, Gutiérrez-Zugasti I, González-Morales MR, Setién J, Ruiz-Martínez E, García-Moreno A, Clemente-Conte I (2013) Shell Technology, Rock Art, and the Role of Marine Resources during the Upper Palaeolithic. Current Anthropology 54:370–380
Dewar G, Pfeiffer S (2010) Approaches to estimating marine protein in human collagen for radiocarbon date calibration. Radiocarbon 52:1611–1625
Estalrrich A, González-Rabanal B, Marín-Arroyo AB, Gutiérrez-Zugasti I, González-Morales MR (2019) Los últimos cazadores recolectores y los primeros agricultores en el abrigo de El Toral III (Llanes, Asturias): una aproximación antropológica. XV Reunión Nacional de Cuaternario, Bizkaia Aretoa, Bilbao, 1-5 Julio 2019: libro de resúmenes
Fano M (2019) The Mesolithic “Asturian” culture (North Iberia), one century on. Quaternary International 515:159–175
Fernandes R, Millard AR, Brabec M, Nadeau MJ, Grootes P (2014) Food reconstruction using isotopic transferred signals (FRUITS): a Bayesian model for diet reconstruction. PloS one 9:87436
Fernández-López de Pablo J, Gutiérrez-Roig M, Gómez-Puche M, McLaughlin R, Silvia F, Lozano S (2019) Palaeodemographic modelling supports a population bottleneck during the Pleistocene-Holocene transition in Iberia. Nature 10
Fuertes-Prieto N, Risseto J, Gutiérrez-Zugasti I, Cuenca-Solana D, González-Morales M (2021) New perspectives on the Mesolithic technology in northern Iberia: data from El Mazo shell midden site (Asturias, Spain). In: Borić DA, Antonović D, Mihailović B (eds) For aging Assemblages, Serbian Archaeological Society, The Italian Academy for Advanced Studies in America, Columbia University, Belgrade - New York, pp 470–475
García-Escárzaga A (2020) Paleoclima y aprovechamiento de recursos costeros durante el Mesolítico en la región cantábrica (N de Iberia). British Archaeological Reports Limited International Series. Vol. 2976. BAR Publishing, Oxford
García-Escárzaga A, Gutiérrez-Zugasti I (2021) The role of shellfish in human subsistence during the Mesolithic of Atlantic Europe: An approach from meat yield estimations. Quaternary International 584:9–19
García-Escárzaga A, Gutiérrez-Zugasti I, Marín-Arroyo AB, Fernandes R, Núñez de la Fuente S, Cuenca-Solana D, Iriarte E, Simões C, Martín-Chivelet J, González-Morales MR, Roberts P (2022a) Human forager response to abrupt climate change at 8.2 ka on the Atlantic coast of Europe. Sci Rep 12: 6481
García-Escárzaga A, Gutiérrez-Zugasti I, Cuenca-Solana D, González-Morales MR, Hammann C, Roberts P, Fernandes R (2022b) Bayesian estimates of marine radiocarbon reservoir effect in northern Iberia during the Early and Middle Holocene. Quaternary Geochronology 67:101–232
González-Morales MR (1982) El Asturiense y otras culturas locales. La explotación de las áreas litorales de la región cantábrica en los tiempos epipaleolíticos. Monografía Nº7, Centro de Investigación y Museo de Altamira, Santander
González-Morales MR (2000) La Prehistoria de las Marismas: Excavaciones en la cueva de La Fragua (Santoña). Campañas de 1990, 1991, 1993, 1994 y 1996. Actuaciones Arqueológicas en Cantabria 1984-1999 (Ontañón Peredo, R. ed.), Consejería de Cultura y Deporte, Santander: 177-179
Gonzaléz-Rabanal B (2022) Dinámicas de población, dieta y prácticas funerarias de los últimos cazadores-recolectores y primeras sociedades campesinas de la Región Cantábrica. Escuela de Doctorado de la Universidad de Cantabria, Tesis Doctoral Inédita
González-Morales MR, Díaz Casado Y (2000) La Prehistoria de las Marismas: excavaciones arqueológicas en los abrigos de la Peña del Perro (Santoña). Actuaciones Arqueológicas en Cantabria 1984-1999 (Ontañón Peredo, R. ed.), Consejería de Cultura y Deporte, Santander, pp 93–96
Gutiérrez-Zugasti I (2009) La explotación de moluscos y otros recursos litorales en la región cantábrica durante el Pleistoceno final y el Holoceno inicial. Publican, Ediciones Universidad de Cantabria, Santander
Gutiérrez-Zugasti I (2011a) Shell fragmentation as a tool for quantification and identification of taphonomic processes in archaeomalacogical analysis: The case of the Cantabrian Region (Northern Spain). Archaeometry 53:614–630
Gutiérrez-Zugasti I (2011b) Coastal resource intensification across the Pleistocene-Holocene transition in Northern Spain: evidence from shell size and age distributions of marine gastropods. Quaternary International 244:54–66. https://doi.org/10.1016/j.quaint.2011.04.040
Gutiérez-Zugasti I (2011c) The use of echinoids and crustaceans as food during the Pleistocene-Holocene transition in Northern Spain: methodological contribution and dietary assessment. J Isl Coast Archaeol 6:115–133
Gutiérrez-Zugasti FI, González-Morales MR (2010) New data on Asturian shell midden sites: the cave of Mazaculos II (Asturias, Northern Spain). In: Álvarez-Fernández E et al (ed) Not only Food. Marine, Terrestial Molluscs in Archaeological Sites. In: Proceeding of the 2nd Meeting of the ICAZ Archaeomalacology Working Group (Santander, February 19th-22nd 2008) Donostia:Aranzadi Zientzia Elkartea, pp 110–118
Gutiérrez-Zugasti I, Cuenca-Solana D (2014) Biostratigraphy of shells and climate changes in the Cantabrian region (Northern Spain) during the Pleistocene-Holocene transition. In: Szabó K et al Archaeomalacology: Shells in the Archaeological Record. Oxford: Publishers of British Archaeological Reports, pp 225–234
Gutiérrez-Zugasti I, Andersen SH, Araújo AC, Monge AM (2011) Shell midden research in Atlantic Europe: State of the art, research problems and perspectives for the future. Quaternary International 239:70–85
Gutiérrez-Zugasti I, Cuenca-Solana D, González-Morales MR, García-Moreno A, Ortíz-Menéndez JE, Riseto J, De Torres T (2013) Back to the Asturian: First result from the Mesolithic shell midden site of El Mazo (Asturian, Northern Spain) In: Daire MY et al (ed) Ancient Maritime Communities and the Relationship between People and Environment along the European Atlantic Coasts. Oxford: Publishers of British Archaeological Reports, pp 483–490
Hammer Ø, Harper DAT, Ryan PD (2001) Past: Paleontological Statistics software package for Education and Date Analysis. Paleontologia Electronica 4:1–9
Hausmann N, Meredith-Williams M, Laurie E (2021) Shellfish resilience to prehistoric human consumption in the southern Red Sea: Variability in Conomurex fasciatus across time and space. Quaternary International 584:20–32
Heaton TJ, Köhler P, Butzin M, Bard., E., Reimer, R.W., Austin, W.E.N., Bronk Ramsey, C., Grootes P.M., Hughen, K.A., Kromer, B., Reimer, P.J., Adkins, J., Burke, A., Cook, M.S., Olsen, J., Skinner, L.C. (2020) Marine20- the marine radiocarbon age calibration curve (0–55,000 cal BP). Radiocarbon 62:779–820
Jerardino A (2010) Large shell middens in Lamberts Bay, South Africa: a case of hunter-gatherer resource intensification. J Archaeol Sci 37:2291–2302
Leorri E, Cearreta A, Milne G (2012) Field observations and modelling of the level changes in the southern Bay of Biscay: Implication for understanding current rates of relative sea-level change and vertical land motion along the Atlantic Europe. Quat Sci Rev 42:59–73
López-Dóriga I (2016) La utilización de los recursos vegetales durante el Mesolítico y Neolítico en la costa atlántica de la península ibérica. Tesis Doctoral Inédita, Universidad de Cantabria, Santander
Maninno MA, Kenneth DT (2001) Intensive mesolithic exploitation of coastal resources? Evidences from a shell deposit on the Isle of Portland (Southern England) for the impact of human foraging on populations of intertidal rocky shore molluscs. J Archaeol Sci 28:1101–1114
Marín-Arroyo AB, Gutiérrez-Zugasti I, Cuenca-Solana Adreu-Alarcón S (2020) Patrones de subsistencia de los grupos mesolíticos en la cueva de El Mazo (Asturias). Estudios de Arqueozoología y Tafonomía de la Península Ibérica. Homenaje al profesor Manuel Pérez Ripoll 21:227–229
Martínez-Cuesta R (2017) Estudio de los invertebrados marinos de la unidad estratigráfica 17 del yacimiento mesolítico de El Toral III (Asturias). Universidad de Cantabria, Santander, Trabajo de Fin de Máster Inédito
Milner N, Craig O, Bailey GN (2007) Shell middens in Atlantic Europe. Oxbow Books, Oxford
Moreno R (1994) Análisis Arqueomalacológicos en la Península Ibérica: Contribución Metodológica y Biocultural. Universidad Autonóma de Madrid, Madrid, Tesis Doctoral Inédita
Núñez de la Fuente S (2018) Dinámicas socio-ecológicas, resiliencia y vulnerabilidad en un paisaje Atlántico montañoso: La region cantábrica durante el Holoceno. Universidad de Cantabria, Santander, Tesis doctoral inedita
North Greenland Ice Core Project members (2004) High-resolution of Northern Hemisphere climate extending into the past interglacial period. Nature 431:147–151
Noval-Fonseca MA (2013) Excavación arqueológica en la Cueva de El Toral III (Andrín, Llanes). In: Excavaciones Arqueológicas en Asturias 2007-2012. En el centenario del descubrimiento de la caverna de Peña de Candamo. Consejería de Educación, Cultura y Deporte Gobierno del Principado de Asturias, Oviedo: 381-384
Ortea, J. (1986) The malacology of La Riera cave. In: Straus, L., G., Clark, G.A. (Eds.), La Riera Cave. Stone Age Hunter-Gatherer adaptations in Northern Spain. Arizona State University (Anthropological Research Papers 36), Tempe: 289-298.
Palacios Eguren N, Vega De La Torre JJ (1997) Guía de conchas de las playas y rías de Cantabria. Serie Naturaleza 1 Gobierno de Cantabria, Santander
Poppe GT, Goto Y (1991) European Seashells. (Polyplacophora, Caudofoveata, Solenogastra, Gastropoda), I. Verlag Christa Hemmen, Germany
Ramsey CB (2001) Development of the radiocarbon calibration program. Radiocarbon 43:355–363
Ramsey B (2009) Dealing with outliers and offsets in radiocarbon dating. Radiocarbon 51:1023–1045
Rigaud S, Gutiérrez-Zugasti I (2016) Symbolism among the last hunter-fisher-gatherers in northern Iberia: Personal ornaments form El Mazo and El Toral III. Quaternary International 407:131–144
Reimer PJ, Austin WEN, Bard E, Bayliss A, Blackwell PG, Bronk Ramsey C, Butzin M, Cheng H, Edwards RL, Friedich M, Grootes PM, Guilderson TP, Hajdas I, Heaton TJ, Hogg AG, Hughen KA, Kromer B, Manning SW, Muscheler R, Palmer JG, Pearson C, van der Plicht J, Reimer RW, Richards DA, Scott EM, Southon JR, Turney CSM, Wacker L, Adolphi F, Büntgen U, Capano Mm Fahrni SM, Fogtmann-Schulz A, Fiedrich R, Köhler P Kudsk S, Miyake F, Olsen J, Reinig F, Sakamoto M, Sookdeo A, Talamo S (2020) The IntCal20 northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62:725–757
Sołtysiak A, Fernandes R (2021) Much ado about nothing: assessing the impact of the 4.2 ka event on human subsistence patterns in northern Mesopotamia using stable isotope analysis. Antiquity 95(383):1145–1160
Straus LG, Clark GA (1986) La Riera Cave. Stone age hunter-gatherer adaptations in Northern Spain. Arizona State University, Tempe
Vega del Sella R (1923) El Asturiense. Nueva industria preneolítica. In: Serie Prehistórica 27, 32. Museo Nacional de Ciencias Naturales, Madrid
Yanes Y, Gutiérrez-Zugasti I, Delgado A (2012) Late-glacial to Holocene transition in northern Spain deduced from land-snail shelly accumulations. Quaternary Research 2:373–385
Acknowledgements
This research was supported by the projects HAR2016-75605-R and PID2021-124059NB-I00, both funded by the Spanish Ministry of Science and Innovation. RAM was supported by a predoctoral grant "Concepción Arenal" (grant number: 2019-8461). AGE during the development of this research was funded by the Catalonia Postdoctoral Programme through a Beatriu de Pinós fellowship (grant number 2020_BP_00240) and he is currently working in the framework of a Marie Sklodowska Curie – Postdoctoral Fellowship (grant reference: HEU-101064225-NEARCOAST), funded by the European Union. This work contributes to the ICTA-UAB “María de Maeztu” Programme for Units of Excellence of the Spanish Ministry of Science an Innovation (Ref.: CEX2019-00940-M). We thank Instituto Internacional de Investigaciones Prehistóricas de Cantabria (IIIPC) for providing technical support. We are also grateful to the scientific colleagues Jara Pascual-Revilla (IIIPC), Carlos García-Noriega (IIIPC), Mónica Fernández-García (Universitat de València) and Marco Vidal-Cordasco (Universidad de Cantabria) for their help during the graphic design process. Finally, the authors would like to thank María Noval-Fonseca and Orlando Morán for their help during fieldwork.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. Predoctoral grant "Concepción Arenal", CVE: 2019-8461, Beatriu de Pinós fellowship, 2020_BP_00240, Marie Sklodowska Curie – Postdoctoral Fellowship, HEU-101064225-NEARCOAST, ICTA-UAB “María de Maeztu” Programme for Units of Excellence of the Spanish Ministry of Science an Innovation, CEX2019-00940-M, project PID2021-124059NB-I00, project HAR2016-75605-R.
Author information
Authors and Affiliations
Contributions
We confirm that all authors have approved the final version of the manuscript and have made substantial contributions. RAM and IGZ designed the project. IGZ excavated at El Toral III cave. RF, RAM, AGE, IGZ designed the Bayesian modelling codes. RAM did the Bayesian models and the archaeomalacological and biometrical analysis. All authors interpreted the data. RAM and IGZ wrote the paper with contributions of all the co-authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Arniz-Mateos, R., García-Escárzaga, A., Fernandes, R. et al. Living in the shore: changes in coastal resource intensification during the Mesolithic in northern Iberia. Archaeol Anthropol Sci 16, 79 (2024). https://doi.org/10.1007/s12520-024-01982-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-024-01982-x