
Abstract
The starch grains present in the dental calculus of four Neolithic individuals of the Dehesilla Cave (Cadiz, South of Spain) were extracted and observed with light microscopy. The variant of the extraction technique used proved to be very effective. A number of Poaceae-Triticeae taxa [wheat (Triticum sp., the main crop, including Triticum aestivum), barley (Hordeum sp.)], Fabaceae-Fabeae-Cicereae and Fagaceae (oaks = Quercus sp., which constitute new data for the area in question) were identified. Some pollen grains and fungal spores were also identified, which helped to provide information on the ecological framework of the crops associated with the archaeological site.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The Dehesilla Cave
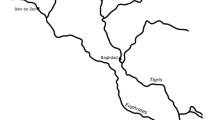
The Dehesilla Cave is located in the Cadiz province of Spain, in the South of the Iberian Peninsula (Fig. 1). It was first explored by Viñas Valverdú (1970, 1971), Martí et al. (1975) and Acosta and Pellicer (1990). In the 21th century, García-Rivero et al. (2018a, b, 2019, 2020, 2022a, b) carried out relevant interdisciplinary works of archaeological and paleoecological nature that allows a better understanding of the human occupation of this important site.

Geographical location map of Dehesilla Cave and location of the archaeological areas C003, C004, and C006 on the three-dimensional plan of the cave
Since 2016, new archaeological excavations took place in Dehesilla Cave, under the direction of Daniel García-Rivero, in six different areas, two outside the cave entrance (C001 and C002) and four inside the cave (C003, C004, C005, and C006) (García-Rivero et al. 2018a, b, 2019, 2020, 2021, 2022a, b; Taylor et al. 2018; Taylor and García-Rivero 2020). On the platform outside the mouth of the cave an Andalusian archaeological sequence, dating from the eleventh-twelfth centuries, was recognized (García-Rivero et al. 2018a; Taylor et al. 2018). Inside the cave, the excavation of two areas in the innermost room (C005 and C006) revealed a Neolithic occupation, with different stratigraphic levels dated from the Early to the Late Neolithic (García-Rivero et al. 2022a, 2023). In the test pit C003, besides a Neolithic sequence, from the Early to the Late Neolithic, a Chalcolithic stratigraphic level was identified as well as two upper historical phases (García-Rivero et al. 2018b, 2019, 2022b). Finally, in the archaeological area C004, the only one that is still under excavation, there is, until now, evidence of a broader occupation ranging from the Middle Neolithic, Late Neolithic, Chalcolithic, Protohistory, and the twentieth century, the latter in the uppermost layer.
Dental calculus
Dental calculus, or tartar, consists of a calcified bacterial dental plaque. It is essentially composed of calcium phosphate mineral salts (White 1997; Eden 2008; Fons-Badal et al. 2020) and originates from intricate interactions between bacteria and saliva (Radini et al. 2017). Dental calculus is very common in human teeth and its formation and quantity depend on a number of factors such as oral hygiene habits, ethnicity, and diet type (White 1997; Arora et al. 2017).
The nature and morphology of starch
Starch is a carbohydrate polymer, composed by two different molecules, both of them made of the monosacharide D-glucose: amylose (linear, constituting 5–35% of natural starches) and amylopectin (ramified, forming 95–65% of this polysaccharide) (Lodish et al. 2004; Seung 2020). It accumulates throughout the photosynthesis in the amyloplasts, and is commonly found, as energy storage granules, in various organs (e.g., tubers, rhizomes and seeds) of vascular plants (Chakraborty et al. 2020; Louderback et al. 2022), constituting the primary storage carbohydrate in the Plant Kingdom (Lodish et al. 2004). The morphology of these granular structures (shape, size, internal and surface characteristics) varies significantly within the different taxa, providing data for their identification (Perez et al. 2009; Ashogbon and Akintayo 2013; Yang and Perry 2013; Copeland and Hardy 2018; Gismondi et al. 2019; Louderback et al. 2022).
The relevance of starch in Archaeology
The importance of starch in Archaeology is mainly associate with two characteristics: a) its resistance, which allows it to be conserved in the dental calculus (Copeland and Hardy 2018). In fact, tartar frequently captures and preserves organic material (including starch) in its structure (Copeland and Hardy 2018; Allende and Samplonius 2021); b) its relevant variability among taxa, which frequently allows the identification of the taxa (family, tribe, genus, species) that produced it (Aceituno-Bocanegra and López-Sáez 2012; Ashogbon and Akintayo 2013; Yang and Perry 2013; Gismondi et al. 2019; Ahituv and Henry 2022; Louderback et al. 2022). In an anthropological context, starch can be extracted from teeth and various human tools, such as pottery and other artifacts (Louderback et al. 2022) and even from pre-historic stone tools (Copeland and Hardy 2018).
Material and methods
Material
The dental calculus founded in the teeth of four individuals of the Dehesilla Cave (Table 1) was used in this work. These teeth were retrieved during different archaeological campaigns in different areas inside the cave (Fig. 1): DH16-3-15C-4 was recovered in 2016, from the archaeological area C003, stratigraphic unit 15C; DH17-6-Locus 2–1 comes from the archaeological campaign of 2017, archaeological area C006, stratigraphic unit Locus 2, while DH19-4–74-Flot.69.2 and DH19-4–74-Flot.71 were recovered in 2019, from the archaeological area C004, stratigraphic unit 74.
With the exception of DH17-6-Locus 2–1, an upper right first premolar, found in situ, in a cranium that probably belonged to a female individual, aged between 24 to 45 years old, all other were loose teeth recovered directly from the sediment or indirectly from sediment flotation, and therefore it is not possible to provide more information pertaining sex or age at death of the individuals they belonged to besides the fact that they are adults.
Only DH17-6-Locus2-1 has a direct radiocarbon dating placing it in Middle Neolithic A (MNA) (García-Rivero et al. 2020). The radiocarbon dates of the remaining teeth are indirect ones, based on the dating of faunal teeth found in the same stratigraphic unit as the human teeth. The obtained date for DH16-3-15C-4 comes from the dating of a caprine/goat molar tooth (García-Rivero et al. 2019), while the dating of the two human teeth identified in the stratigraphic unit 74, from the archaeological area C004, was based on the dating of a caprine molar. This last one is an unpublished AMS date performed at the Centro Nacional de Aceleradores, University of Seville (Table 2).
Methods
After mechanical extraction, the dental calculus was triturated with a glass rod in a stainless-steel fine mesh filter (mesh size = 1/10 mm), to facilitate its maceration, washed with diluted (1N) hydrochloric acid and put in Eppendorf tubes (2 ml). Subsequently, the tubes were kept for 48 h at 4º C, and, afterwards, centrifugated (9000 rpm, 3 min) and decanted. Then, they were washed twice with distilled water, decanted and filled with glycerol. A drop of 50% glycerol with the sediment was transferred to a microscope slide and the cover was doubly sealed with nail-polish. The chemical identification of starch was made by the classical Lugol’s iodine solution. The observations and micrographs were made with a Motic BA 310 light microscope (LM) operating at × 600 and equipped with a digital camera. The material was kept in the reference pollen collection of the Life Sciences Department.
In the identification of starch granules, a number of articles (Aceituno Bocanegra and López Sáez 2012; Yang and Perry 2013; Gismondi et al. 2019; Chakraborty et al. 2020; Monge Calleja et al. 2020; Ahituv and Henry 2022; Louderback et al. 2022), dichotomous keys (Yang and Perry 2013; Monge Calleja et al. 2020; Ahituv and Henry 2022; Louderback et al. 2022) and specialized websites (Starch Grain Database 2022) were used. The modern starch collection of the Department of Life Sciences of the University of Coimbra was also employed (Figs. 2 and 3). The terminology follows essentially the International Code for Starch Nomenclature (ICSN 2011).

Modern starch grains of A Hordeum vulgare; B Triticum aestivum; C T. durum; D Cicer arietinum; E Lathyrus sativus; F Lens culinaris; G Vicia faba

Modern starch grains of Quercus (Fagaceae). A Q. coccifera; B Q. faginea; C Q. ilex; D Q. suber. Scale bar = 20 μm
Results
Starch grains were obtained from all the four individuals that were studied. Their main characteristics and identification are summarized in Table 3. As the starch of barley and wheat species share several features (e.g., a marked bimodal distribution of the sizes of the smaller and larger grains), Table 4 compares the dimensions of the largest modern starch grains of these species. There is a clear difference in size between the grains of the two wheat taxa (which are significantly larger) and those of barley.

Starch grains. A-F: Individual DH16-3-15C-4. A-B Triticum spp. (Poaceae, Triticeae) and Fabaceae-Fabeae (Vicia faba, Lathyrus spp. or, less probably, Lens culinaris); C Triticum spp. and Fagaceae (Quercus faginea or Q. suber, less probably Q. ilex); D Poaceae-Triticeae (probably Triticum spp.); E Triticum spp.; F Poaceae-Triticeae (Hordeum spp. or Triticum spp.). G-L: Individual DH17-6-Locus 2. G Triticum spp. (probably T. aestivum); H Triticum spp. and Hordeum spp.; I Triticum spp. and Fabaceae-Fabeae (V. faba, Lathyrus spp. or L. culinaris) or, less probably, Fabeae-Cicereae (Cicer arietinum); J Triticum spp. and Fagaceae (Q. faginea or Q. suber, less probably Q. ilex); K Triticum spp. and Fabaceae-Fabeae (V. faba, Lathyrus spp. or L. culinaris) or, less probably, Fabeae-Cicereae (C. arietinum); L Triticum spp. and Fabaceae-Fabeae (V. faba, Lathyrus spp. or L. culinaris). a) Triticum spp.; b) Fabaceae; c) Quercus spp.; d) Hordeum spp; 1) pore; 2) depressions; 3) hilum; 4) fissures. Scale bar = 20 μm

Starch grains. A-F: Individual DH19-4–74-Flot. 69.2. A Triticum spp. (probably T. aestivum) treated with Lugol’s iodine, showing a strong reaction; B Vicia faba or, less probably, Lens culinaris, treated with Lugol’s iodine, showing a strong reaction; C-E, F Lathyrus spp. (probably L. sativus) or Vicia faba. G-L: Individual DH19-4–74-Flot. 71. G Triticum spp.; H-I Hordeum spp.; J Fabaceae; K Triticum spp. (probably T. aestivum) treated with Lugol’s iodine, showing a mild reaction; L Triticum spp., less probably Hordeum spp., treated with Lugol’s iodine, showing a strong reaction; 3) hilum; 5) lamellae. Scale bar = 20 μm

A Individual DH17-6-Locus 2—Pollen grain of an Asteraceae-Gnaphalieae; B Individual DH19-4–74-Flot. 69.2—Phaeodidimospores of a fungus, possibly Cladosporium spp. or Asperisporium spp.; C-D Individual DH19-4–74-Flot. 71: C) Hyaloamerosporae of a fungus, probably Ustilago spp., less probably, Tilettia spp.; D—Hyalophragmospore of a fungus, possibly Cochliobolus spp. or Pirenophora spp. Scale bar = 20 μm
The percentages of wheat and barley starch grains identified in relation to the total number of Triticeae identified up to the genus, as well as the percentages of Triticeae, Fabaceae and Quercus in relation to the total number of identified starches for each individual analyzed are respectively presented in Figs. 7 and 8.

Cumulative bar chart with the percentages of wheat and barley in relation to the total number of Triticeae identified up to the genus

Cumulative bar chart with the percentages of Triticeae, Fabaceae and Quercus in relation to the total number of identified starches
Discussion
Starch extraction
The variant of the method for extracting the starch used in this work proved to be very effective. In fact, the use of a higher concentration of hydrochloric acid (1N instead of 0.1 N) for less time allowed the process to be faster (48 h instead of 96 h) without damaging the grains of this polysaccharide. Also, the mechanical actions described above facilitated the maceration of the calculus and the release of the starch without harming it. The success of this variant can be easily seen from Figs. 4 and 5. Indeed, in all cases the amount of starch grains found was high. It should be noted that they often constituted aggregates, including one or more genera or families.
Starch identification
To identify the taxa that produced the starch granules, all the qualitative and quantitative characteristics were considered. We agree with Yang and Perry (2013) that the larger starch grains are the most informative and the ones that are easiest to identify. However, in some cases, combining the characteristics of large grains and small grains can also be useful (see 4.2.1).
Poaceae-Triticeae
The dispersal of agriculture on the Iberian Peninsula is recorded from, at least, 5550 cal BC (the first evidence of farming—grains of Poaceae—belonging to the site of Mas d’Is, in the province of Alicante) (Peña-Chocarro et al. 2018), and cereals belonging to the Poaceae-Triticeae are among the first of these crops (Alonso Martínez 2000; Zapata et al. 2004; Peña-Chocarro et al. 2013, 2018). During the second half of the sixth-millennium cal BC, agriculture extended to almost the entire Peninsula, including Andalusia (Peña-Chocarro et al. 2018).
Triticum was, without doubt, the most important crop of all we found, and we could identify it in all the individuals (Figs. 4A-E, F-L, 5C-E, K-L). The presence of an observable hilum in the sub-polygonal smallest grains of Fig. 4(G, H, K, L), in conjunction with other characteristics (e.g., size) made it possible to identify them as belonging to this genus (Aceituno-Bocanegra and López-Sáez 2012), and, in some cases, with a good probability (larger dimensions), to the species T. aestivum. These identifications are in agreement with the results previously described by García-Rivero et al. (2019, 2022b) with the identification of wheat seeds (Triticum aestivum-durum) in all Neolithic sequence, since the ENB, in both analyzed archaeological areas (C003 and C006) of Dehesilla cave.
Nevertheless, Hordeum was also present (Fig. 5H-I), barley starch is quite similar to that of wheat, but in general, it is smaller, with more frequent abnormal shapes and better-defined lamellae (Aceituno-Bocanegra and López-Sáez 2012), which allowed us, in a number of cases, to separate, with a good probability, the two genera. García-Rivero et al. (2019: 225, 2022b) have found a similar balance between barley (Hordeum vulgare) and naked wheats (Triticum aestivum-durum) in the carpological analysis of the sediments of both archaeological areas (C003 and C006) attributed to Early Neolithic B, where the individual DH16-3-15C-4 comes from. However, from the calculus analysis, we see a clear predominance of wheat (96.3%) over barley (3.7%). This predominance was observed in the MNA period, when barley seeds appear very scarcely in the archaeological area C003 (García-Rivero et al. 2019), and were not identified in the archaeological area C006 (García-Rivero et al. 2022b). In the calculus analysis of the individual DH17-6-Locus2-1 this cereal was identified, although in less quantity (8.3%) than wheat (91.7%). In the MNB period, no barley seeds were identified in the archaeological area C003 (García-Rivero et al. 2019), but in the archaeological area C006 there is once again a clear predominance of wheat over barley (García-Rivero et al. 2022b). The individuals that we have from this period (MNB) were retrieved in the archaeological area C004 from which there is still no archaeobotanical data, but assuming a likely similar behavior between the coeval stratigraphic levels the data obtained for the archaeological areas C003 and C006 can be used as a proxy. From the calculus analysis for these two individuals (DH19-4–74-Flot.69.2 and DH19-4–74-Flot.71) we can see an increase of barley (45.5% and 42.9%) in relation to wheat, with a better balance between these two taxa.
In this context, it is relevant to mention that several authors (Alonso Martínez 2000; Zapata et al. 2004; Agrologia 2023) mentioned that wheat and barley were the first cereals cultivated in the Iberian Peninsula, already since the Early Neolithic, with the cultivation of rye (Secale cereale L.) and oat (Avena sativa L.) being quite later (Zapata et al. 2004; Coleto Martínez et al. 2016). Regarding these two species, it should be noted that oats have compound starch grains and the grains of rye present a fissured, commonly star-shaped hilum which is not the case with wheat or barley (Gismondi et al. 2019; Monge Calleja et al. 2020).
Fabaceae-Fabeae / Cicereae
After the Poaceae, Fabaceae (Leguminosae) is, certainly, the most important family in the feeding of humanity, their presence in the Iberian Peninsula being as ancient as that of cereals (Alonso Martínez 2000; Peña-Chocarro et al. 2018; Marinangeli 2020), and they were commonly cultivated together (Zapata et al. 2004; Peña-Chocarro et al. 2018; 2016—Anno Internazionale dei Legumi 2023). From the point of view of human nutrition, pulses complement cereals in a harmonious and healthy way, because they are very rich in proteins and amino acids [and other nutrients, such as fiber, potassium, and iron (Marinangeli 2020)], so they were—and still are—used as a meat substitute. On the other hand, by fixing nitrogen, they fertilize the fields where they are grown (Alonso Martínez 2000).
Fabeae is its more relevant tribe, including such important crops as broad-bean (Vicia faba L.), lentil (Lens culinaris L.), and pea (Pisum sativum L.). However, some other species, like grass-pea (Lathyrus sativus L.) and sweet-pea (Vicia sativa L.) are also edible. All of these taxa have a long history as foodstuffs, and have been consumed by humankind for thousands of years. It is worth noting that the starch of Fabaceae-Fabeae was founded in the dental calculus of all the individuals we studied (see Table 3 and Figs. 4A, B, K, L and 5B, F, J). The separation of the Fabeae species by the morphology of their starch grains is in general not easy, as there are more or less extensive overlaps between them. However, pea starch is an exception to this rule, because it presents grains of the compound type, that are absent in the other taxa. As for the other food species, some qualitative [e.g., distinct/indistinct lamellae and hilum (Ahituv and Henry 2022)] and quantitative characters [length and width of the grains (personal data)] and ratios [length/width (personal data)], may, in some cases, be useful.
The tribe Cicereae includes chickpea (Cicer arietinum L.), another important pulse. The presence of chickpea starch in the tartar of individual DH17-6-Locus 2–1 should not be ruled out. It must be pointed out that some grains (Fig. 4K) present a somewhat bifid hilum, a common feature with, for example, lentil (Lens culinaris L.), although, given the available data in the literature, we consider it more likely to correspond to this species or to fava beans.
Bearing in mind the relevant morphological diversity of Fabaceae starch found in this work, we consider it likely that, associated with the margins of the cereal fields (wheat and barley) in the Dehesilla area, several pulse species were cultivated together, a condition previously noted, for the Iberian Peninsula, by several authors (e.g., Alonso Martínez 2000; Zapata et al. 2004; Peña-Chocarro et al. 2013, 2018).
As already mentioned, Fabaceae starches were identified in the dental calculus of all the individuals under analysis and it looks like there was a slight increase over time (19.7% in DH16-3-15C-4, 23.8% in DH17-6-Locus 2–1, 20% in DH19-4–74-Flot.69.2 and 40% in DH19-4–74-Flot.71). From the archaeobotanical data available for Dehesilla cave, the presence of legumes in the ENB period from the archaeological area C003 is limited to one possible vetch (Lathyrus sp.) identified in the stratigraphic unit 15A, while in C006, two seeds of Fabaceae were identified. No legumes were identified in stratigraphic unit 15C from where the individual DH16-3-15C-4 was recovered. In the MNA period, a fragment attributed to Vicia/Lathyrus was recognized in the stratigraphic unit 14 of the archaeological area C003 (García-Rivero et al. 2019), but there is no record of Fabaceae in the MNA levels from archaeological area C006, namely in the sediment recovered from the stratigraphic unit Locus 2 (García-Rivero et al. 2020, 2022b), from where the individual DH17-6-Locus 2–1 comes from. This may result from the different use of both areas, while C006 was a unique funerary event, C003 seems to have served as a domestic space. For the MNB period, García-Rivero et al. (2019) only detected a faba bean seed in the archaeological area C003 and no legumes in C006, while from the calculus analysis, the calculated percentages of the starches of Fabaceae were 20% (DH19-4–74-Flot.69.2) and 40% (DH19-4–74-Flot.72).
Fagaceae
The human consumption of acorns is quite ancient. In fact, acorns have been associated with archaeological finds since the Mesolithic in Europe and the Mediterranean Basin, throughout much of the distribution area of the genus Quercus (Mason 1992).
Some starch grains of Quercus were identified in the dental calculus of individuals DH16-3-15C-4 (Fig. 4C) and DH17-6-Locus 2–1 (Fig. 4J). Its specific morphology (see Table 3) suggests that they probably belong to the Portuguese oak (Q. faginea Lam. – Fig. 3B) or the cork oak (Q. suber L. – Fig. 3D) or, less probably, to the holm (or holy) oak [Q. ilex L. subsp. ballota (Desf.) Samp. – Fig. 3C], all of them present in the Cadiz province (Galiano 1987; Franco 1990). The kermes oak (Q. coccifera L. – Fig. 3A), also present in the Cadiz province, seems to be excluded. In fact, the sub-triangular shape observable in the referred individuals (Fig. 3B, D) is less frequent in the holm oak and, mainly, in Q. coccifera (Fig. 3A). Also, the dimensions of the starch grains of this last species are clearly larger than those of our reference collection. We consider this as a relevant new data for the archaeological excavations of the Dehesilla cave. From the pollen analysis García-Rivero et al. (2019) have found in their palynological analysis high percentages of pollen of Quercus ilex type in the ENB (38.9%-42.7%), with similar values in the MNA, and a pronounced decrease in the MNB (15.7–18%). This decreasing of Quercus over time, apparently is what we see from the dental calculus analysis, since Quercus starches were identified in low quantities only in the dental calculus of the individuals from the ENB (DH16-3-15C-4: 3.28%) and MNA (DH17-6-Locus 2–1: 0.6%).
Other structures
Pollen grains
The pollen grains found in DH17-6-Locus 2–1 undoubtedly belong to an Asteraceae, and, more precisely, to the tribe Gnaphalieae (Fig. 6A). Indeed, the pollen of the Anthemideae does not present caveae (Dimon 1971; Diez 1987), that of Cynareae have larger dimensions and more robust spines (Dimon 1971; Diez 1987; Reille 1992) and, with the only exception of Scorzonera humilis the pollen of the Cichorieae present a very typical echinolophate pattern of sculpture (Blackmore 1984; Diez 1987; Reille 1992). As for the Astereae, Senecioneae, Eupatorieae, Inuleae and Heliantheae s.l., the polar axis, equatorial diameter, and spines are larger (Dimon 1971; Diez 1987; Reille 1992; Pereira Coutinho 2002). It is important to emphasize that a considerable number of species of Gnaphalieae [belonging, for instance, to the genera Bombycilaena (DC.) Smoljan., Filago Loefl. ex L., Gnaphalium L., and Logfia Cass.] live associated with or bordering crop fields (Pereira Coutinho 1939; Devesa 1987; Andrés-Sánchez et al. 2019), so, in this case, their presence in the dental calculus would probably be due to involuntary contamination of cereals with taxa of this tribe, excluding Helichrysum Mill. and Phagnalon Cass., which pollen spines are larger and less numerous (Dimon 1971; Pereira Coutinho 2002). A less likely hypothesis is the intentional consumption of plants from this tribe for health reasons. In fact, some species of Gnaphalium are used in folk medicine, because they have applications such as expectorant, suppressor, or reliever of coughing or anti-inflammatory (Zheng et al. 2013). This would also explain their consumption and capture of the grains by the dental tartar.
García-Rivero et al. (2019) referred to the presence of a small number of pollen grains of Aster type (4–4.6%) in the MNA. This type corresponds to the Bellis annua type described by Díez (1987) that includes the genera of Gnaphalieae above mentioned.
Fungal spores
It is worth noting that although the spore diversity in the Kingdom Fungi is huge, in all cases the spores found in the dental calculus of the studied individuals could be associated, with a relevant probability, with some graminicolous genera. In fact, the phaeodidimospores extracted from the individual DH19-4–74-Flot.69.2 could be associated with the genera Cladosporium Link (Perelló et al. 2003; Smith 2000; Fungi of Great Britains and Ireland 2023) or Asperisporium Maubl (Fig. 6B). The hyaline, 4-septed phragmospore that was found in the DH19-4–74-Flot.71 individual, could easily belong to Cochliobolus sativus (S. Ito & Kurib.) Drechsler ex Dastur, Pirenophora tritici-repentis (Died.) Drechsler (Benslimane 2015; Smith 2000; Rózewicz et al. 2021) or Ustilago tritici (Pers.) C. N. Jensen, Kellerm. & Swingle [all infecting wheat (Benslimane 2015)] or Pirenophora teres Drechsler that infects barley (Herbario Virtual Fitopatología 2023) (Fig. 6C). Likewise, the presence of the hyaline, microreticulate, or microspinulate amerospore that was present in the same individual may, with good probability, be associated with the crop-pathogenic genus Ustilago (Pers.) Roussel (Fig. 6D) (Kruse et al. 2018; Fungal Spores 2023). Tilletia Tul. & C. Tul. could also be a good candidate, but its teliospores are larger [ca. 16–24 μm (Trione 1974; Pimentel et al. 2000; Župunski et al. 2012)] so this would be difficult unless the specimen photographed was quite immature, which is not probable. It is rather unlikely that this last type of spores belongs to Puccinia Pers., a genus that commonly infects wheat, causing rust. In fact, among their different types of spores, the aeciospores and urediniospores are similar in morphology, but not in size, while the opposite is true for basidiospores (Smith 2000; Anikster et al. 2005). All the aforementioned fungi constitute common crop pests that could easily be present in the cultivated fields of the Dehesilla region. It must be noted that at least part of the identified wheat grains, i.e., the larger, belong to the free-threshing species bread wheat (Triticum aestivum) or macaroni wheat (T. durum), which are more prone to fungal infections than the hulled-wheats einkorn (T. monococcum) and emmer (T. dicoccum), because the latter species, unlike the first two, retain their tough glumes associated with the caryopses when they are released, which gives them some protection against fungal attacks (Nesbitt and Samuel 1996; Curzon et al. 2021).
Dental calculus analysis represents direct evidence of feeding behavior. Besides assisting in the paleoenvironment reconstruction, it also allows for a glimpse of past individual dietary habits, however, its interpretation is not straightforward. Several aspects must be pondered. First, with this technique only starchy foods can be identified so we are far from scrutinizing the entire diet; secondly, we should be aware that not all starchy foods are trapped in dental calculus, therefore the use of proportions must be interpreted with caution, the reason why there are researchers, such as D’Agostino et al. (2022) that consider dental calculus analysis as a qualitative method; and lastly, another important issue to be considered is that it is not possible to determine when dental calculus deposits were formed, there is great variability in respect to its formation and composition among individuals (Dobney and Brothwell 1988; Henry and Piperno 2008; D’Agostino et al. 2022).
Our study is a preliminary analysis, given the small sample size, however, some important aspects are emerging. Our data shows that wheat and barley were available in the Dehesilla cave in the Neolithic period, which is in accordance with the archaeobotanical date for this site (García-Rivero et al. 2019, 2022b). Assuming that these proportions observed are reliable, apparently, the diet of the individuals from the ENB and MNA periods had a strong emphasis on wheat, the predominant cereal consumed, but the relative importance of this cereal seems to decrease in the MNB period, in which there is an increase in the consumption of barley towards more balanced levels. This could be explained by a greater availability of wheat over barley in the surroundings of Dehesilla cave, in the ENB and MNA, however the results obtained from the archaeobotanical analysis points to balance between wheat and barley in the ENB, which means that the much higher proportion of wheat observed in the individual dated from the ENB may reflect an individual consumption preference or a differential access to these cereals, or in fact the individual consumed both cereals in similar proportions but barley starches were not trapped in dental calculus deposits. The archaeobotanical analysis (García-Rivero et al. 2019, 2022b) shows a notable predominance of wheat over barley in MNA the archaeological area C003, which is in accordance with our results, but no barley in the archaeological area C006, what, as previously mentioned, most probably results from the fact that Locus 2, in archaeological C006, was an intentional deposition of two crania associated to a juvenile sheep/goat skeleton with ritual purposes (García-Rivero et al. 2020) and not a residence area. Nevertheless, our results for MNB period slightly disagree with the archaeobotanical data, since for the archaeological area C003 no barley seeds were identified in MNB, and a predominance of wheat over barley is observed in the archaeological area C006 for this period (García-Rivero et al. 2022b), while dental calculus analysis denote a significant increase in barley consumption when compared to the individuals from the ENB and MNA, with a balance between wheat and barley. Personal choices seem to be the most plausible explanation, since it looks like there was no differential access to both cereals, as they appear in similar proportions in dental calculus, and the possibility that the individuals DH19-4–74-Flot.69.2 and DH19-4–74-Flot.71were actually consuming more wheat, whose starch grains for some reason were not retained in dental calculus, loses strength since we have two individuals with the same behavior. However, it should be noted that in C003, immediately after MNB period, there is a part of the stratigraphic sequence dating from the transition between MNB and LN (Late Neolithic) where barley is represented with the same ubiquity of wheat. Also, it must be noted that the analyzed individuals DH19-4–74-Flot.69.2 and DH19-4–74-Flot.71 come from a part of the stratigraphic sequence of C004 area dated from MNA-MNB, not exclusively MNB. The MNB period is currently the most difficult Neolithic period to define in terms not only of radiocarbon dating but also of material culture, as has been noted previously (García-Rivero et al. 2022a). We hope that the expected archaeobotanical data from the archaeological area C004, where these two individuals come from, may shed new light on this issue soon.
Cereals were indeed a very important component of the diet of these individuals, but the consumption of vegetables was also relevant and apparently much more so than anticipated by the archaeobotanical studies. While García-Rivero et al. (2022b) refers to the presence of few seeds or none in the different stratigraphic levels analyzed on C006 area, Fabaceae starches were recognized in the dental calculus of all the individuals, in orders of magnitude from approximately 20% to 40% in the MNB. This fact was better documented in MNB period of C003 area, where other botanical specimens (fruits, weeds, and legumes) were predominant over cereals, but, in any case, highlights the importance of dental calculus analysis as a complement to archaeobotanical research. In fact, we have observed less total starch grains in the MNB, which can be justified by a higher proportion of fruits in the diet, in line with the results obtained by the archaeobotanical analysis that points to a significant increase in fruits (García-Rivero et al. 2022b) like wild apples [Malus sylvestris (L.) Mill. (Aedo et al. 1998)], plums [e.g., Prunus insititia L. (Blanca and Díaz de la Guardia 1998)] and grape vines [Vitis vinifera L. (Morales and Ocete 1998)], which have little or no starch.
Apparently, acorns were consumed only in the ENB and MNA, whereas in the MNB no Quercus starches were identified. This may be explained by the forest decline observed for this period related to animal grazing activities, as well as to the transition from an intensive to an extensive farming system (García-Rivero et al. 2019).
Conclusions
Our results largely agree with those obtained by other authors for the Dehesilla Cave. Namely, the presence of Poaceae-Triticeae (Triticum, Hordeum) starch grains. The presence of Fabaceae is however higher than anticipated by the archaeobotanical analysis for this site and the existence of Quercus starch grains in the dental calculus of the individuals is a new data, that demonstrates, for the first time, the consumption of acorns by the inhabitants of Dehesilla cave. By its turn, the identification of the pollen grains and fungal spores contained in the dental calculus are congruent with the existence of cultivated areas of cereals near the Dehesilla Cave.
References
2016 – Anno Internazionale dei Legumi (2023) https://www.fao.org/pulses-2016/news/news-detail/it/c/429320/. Accessed 25 Jan 2023
Aceituno-Bocanegra FJ, López-Sáez JA (2012) Caracterización morfológica de almidones de los géneros Triticum y Hordeum em la Península Ibérica. Trab De Prehist 69(2):332–348. https://doi.org/10.3989/tp.2012.12095
Acosta P, Pellicer M (1990) La cueva de la Dehesilla (Jerez de la Frontera). Las primeras civilizaciones productoras en Andalucía Occidental. CSIC, Jerez de la Frontera
Aedo C, Muñoz Garmendia F, Navarro C (1998) Malus L. In: Muñoz Garmendia F, Navarro C (eds) Flora Iberica – Plantas vasculares de la Península Ibérica e Islas Baleares, vol. VI – Rosaceae. Real Jardín Botánico, CSIC, pp 438–444
Agrologia (2023) Pre-historia de la agricultura en la Península Ibérica. https://agrologia.wordpress.com/2017/03/15/pre-historia-de-la-agricultura-en-la-peninsula-iberica/. Accessed 20 Jan 2023
Ahituv H, Henry AJ (2022) An initial key of starch grains from edible plants of the Eastern Mediterranean for use in identifying archaeological starches. J Archaeol Sci Rep 42:103396. https://doi.org/10.1016/j.jasrep.2022.103396
Allende MK, Samplonius A (2021) Dental anthropological report: exploring plant-based treatments through the analysis of dental calculus and sediment of dental caries in a woman from the Late Preceramic period. Peru Ann Anat 240:151849. https://doi.org/10.1016/j.aanat.2021.151849
Almaguer M, Aira MJ, Rodríguez-Rajo MJ, Fernandez-Gonzalez M, Rojas-Flores TI (2015) Thirty-four identifiable airborne fungal spores in Havana, Cuba. Ann Agric Environ Med 22:215–220. https://www.aaem.pl/pdf-72262-9490?filename=Thirty_four%20identifiable.pdf
Alonso Martínez N (2000) Cultivos y producción agrícola en época ibérica. III Reunió sobre Economia en el Món Ibèric. SAGVNTVM-PLAV, Extra-3:25–46
Andrés-Sánchez, Martinez Ortega MM, Rico E (2019) Filago Loefl. ex L. In: Benedi C, Buira A, Rico E, Crespo MB, Quintanar A, Aedo C (eds) Flora Iberica - Plantas vasculares de la Peninsula Ibérica e Islas Baleares. Real Jardín Botánico, CSIC, Madrid, pp 1672–1704
Anikster Y, Eilam T, Bushnell WR, Kosman E (2005) Spore dimensions of Puccinia species of cereal hosts as determined by image analysis. Mycol 97(2):474–484. https://www.jstor.org/stable/3762265. Accessed 12/05/2023
Arora G, Mackay DF, Conway DI, Pell JP (2017) Ethnic differences in oral health and use of dental services: cross-sectional study using the 2009 adult dental health survey. BMC Oral Health 17:1. https://doi.org/10.1186/s12903-016-0228-6
Ashogbon AO, Akintayo ET (2013) Morphological and functional properties of starches from cereal and legume: a comparative study. Int J Biotechnol Food Sci 1(4):72–83
Benslimane H (2015) Revue bibliographique: la maladie de la tache aureolée du blé. OEPP/EPPO Bull 45(1):52–65. https://doi.org/10.1111/epp.12191
Blackmore S (1984) Compositae – Lactuceae. Rev Palaeobot Palynol 42:45–85. https://doi.org/10.1016/0034-6667(84)90062-9
Blanca G, Díaz de la Guardia C (1998) Prunus L. In: Muñoz Garmendia F, Navarro C (eds) Flora Iberica – Plantas vasculares de la Península Ibérica e Islas Baleares, vol. VI – Rosaceae. Real Jardín Botánico, CSIC, pp 444–466
Boukhelkhal M, Moulai-Mostefa N (2017) Physicochemical characterization of starch isolated from soft acorns of holm oak (Quercus ilex subsp. ballota (Desf.) Samp.) grown in Algeria. J Food Meas Charact 11(4). https://doi.org/10.1007/s11694-017-9582-6
Chakraborty I, Pallen S, Shetty Y, Roy N, Mazumder N (2020) Advanced microscopy techniques for revealing molecular structure of starch granules. Biophys Rev 12:105–122. https://doi.org/10.1007/s12551-020-00614-7
Coleto Martínez JM, Bartolomé García T, Velásquez Otero R (2016) Historias de plantas (III): la historia del Centeno. In: La Agricultura y la Ganadería Extremeñas en 2016. Fundación CB, Badajoz, pp 263–278
Copeland L, Hardy K (2018) Archaeological Starch Agron 8(1):4. https://doi.org/10.3390/agronomy8010004
Curzon AY, Kattakota C, Nashef K, Abbo S, Bonfil DJ, Reifen R, Bar-El S, Rabinovitch O, Avneri A, Ben-David R (2021) Assessing adaptive requirements and breeding potential of spelt under Mediterranean environment. Sci Rep 11:7208. https://doi.org/10.1038/s41598-021-86276-1
D’Agostino A, Di Marco G, Marvelli S, Marchesini M, Rizzoli E, Rolfo MF, Canini A, Gismondi A (2022) Neolithic dental calculi provide evidence for environmental proxies and consumption of wild edible fruits and herbs in central Apennines. Commun Biol 5:1384. https://doi.org/10.1038/s42003-022-04354-0
Devesa JA (1987) Tribu Inuleae. In: Valdés B, Talavera S, Galiano EF (eds) Flora Vascular de Andalucía Occidental, vol 3. Ketres Editora S. A., pp 15–38
Diez MJ (1987) Asteraceae-Compositae. In: Valdes B, Diez MJ, Fernandez I (eds) Atlas polinico de Andalucia Occidental. Instituto de Desarollo Regional No.943. Universidad de Sevilla y Excma, Diputación de Cadiz, Sevilla, pp 332–357
Dimon MT (1971) Problèmes généraux soulevés par l’étude des Composées Mediterraneènes. Naturalia Monspeliensia, Série Botanique 22:129–144
Dobney K, Brothwell D (1988) A scanning electron microscope study of archaeological dental calculus. In: Olsen S (ed) Scanning electron microscopy in archaeology, BAR International Series, vol 452. UK BAR, Oxford, pp 372–385
Eden BD (2008) Prevention strategies for periodontal diseases. In: Capelli DP, Mobley CC (eds) Prevention in clinical oral health care. Elsevier, Amsterdam, pp 213–229
Fons-Badal C, Fons-Font A, Labaig-Rueda C, Solá-Ruiz MF, Selva-Otaolaurruchi E, Agustín-Panadero R (2020) Analysis of predisposing factors for rapid dental calculus formation. J Clin Med 9(3):858. https://doi.org/10.3390/jcm9030858
Franco JA (1990) Quercus. In: Castroviejo S, Laínz M, López González G, Montserrat P, Muñoz Garmendia F, Paiva J, Villar L (eds) Flora Iberica, vol. 2. Real Jardín Botánico, C.S.I.C., Madrid, pp 15–36
Fungal Spores (2023) Ustilago tritici. https://sites.google.com/site/fungalspores/welcome-to-the-fungal-spore-photo-site/basidiomycete-spores/ustilago-tritici. Accessed 10 Jan 2023
Fungi of Great Britains and Ireland (2023) https://fungi.myspecies.info/all-fungi/cladosporium-sp-p4871. Accessed 9 Jan 2023
Galiano EF (1987) Fagaceae. In: Valdés B, Talavera S, Fernández-Galiano E (eds) Flora vascular de Andalucía Occidental, vol. 1. Ketres Editora S. A., Barcelona, pp 159–163
García-Rivero D, Taylor R, Pérez-Aguilar LG, Pérez-Jordà G, García-Viñas E, López-Sáez JA, Zurro D, Peña-Chocarro L, Bernáldez-Sánchez E (2018a) Andalusi populations at La Dehesilla Cave (Sierra de Cadiz, Southern Iberia): and interdisciplinary approach to their rural economic systems. J Islam Archaeol 5(2):119–151
García-Rivero D, Vera-Rodríguez JC, Díaz-Rodríguez MJ, Barrera-Cruz M, Taylor R, Pérez-Aguilar LG, Umbelino C (2018b) La Cueva de la Dehesilla (Sierra de Cádiz): vuelta a un sitio clave para el Neolítico del sur de la península ibérica. Munibe Antropol-Arkeol 69:5–15. https://doi.org/10.21630/maa.2018.69.19
García-Rivero D, Pérez-Jordà G, García-Viñas E, López-Sáez JA, Taylor R, Peña-Chocarro L, Bernáldez-Sánchez E, Pérez-Díaz S (2019) Ecological patterns and use of natural resources during the Neolithic of the south of the Iberian Peninsula: An update from the 6th to 4th millennia cal BC sequence of Dehesilla Cave. Quat Sci Rev 219:218–235. https://doi.org/10.1016/j.quascirev.2019.07.010
García-Rivero D, Taylor R, Umbelino C, Price TD, García-Viñas E, Bernáldez-Sánchez E, Pérez-Jordà G, Peña-Chocarro L, Barrera-Cruz M, Gibaja-Bao JF, Díaz-Rodríguez MJ, Monteiro P, Vera-Rodríguez JC, Pérez-González J (2020) The exceptional finding of Locus 2 at Dehesilla Cave and the Middle Neolithic ritual funerary practices of the Iberian Peninsula. PLoS ONE 15(8):e0236961. https://doi.org/10.1371/journal.pone.0236961
García-Rivero D, Taylor R, Umbelino C, Cubas M, Barrera-Cruz M, Díaz-Rodríguez MJ (2021) Early Neolithic ritual funerary behaviours in the Westernmost regions of the Mediterranean: New insights from Dehesilla Cave (Southern Iberian Peninsula). Doc Praehist 48:298–327. https://doi.org/10.4312/dp.48.8
García-Rivero D, Barrera-Cruz M, Díaz-Rodríguez MJ, Vera-Rodríguez JC, Taylor R (2022a) Cueva de la Dehesilla: nuevos datos estratigráficos, materiales y radiocarbónicos (corte C006) y revisión de su secuencia neolítica. Spal 31(1):57–91
García-Rivero D, García-Viñas E, Pérez-Jordà G, Taylor R, Bernáldez-Sánchez E, Peña-Chocarro L (2022b) Human ecology and the Southern Iberian Neolithic: an approach from archaeobotany and archaeozoology. J Field Archaeol 47(8):536–555. https://doi.org/10.1080/00934690.2022.2135248
García-Rivero D, Taylor R, Barrera-Cruz M, López-Carmona D, Díaz-Rodríguez MJ, Moreno-Caballud A (2023) Estratigrafía, materiales arqueológicos y dataciones del corte C005 de Cueva de la Dehesilla: primicia pleistocena y nueva contribución a la secuencia neolítica. Lucentum 42:9–32. https://doi.org/10.14198/LVCENTVM.23443
Gismondi A, D’Agostino A, Canuti L, Di Marco G, Basoli F, Canini A (2019) Starch granules: a data collection of 40 food species. Plant Biosyst 153(2):273–279. https://doi.org/10.1080/11263504.2018.1473523
Henry AG, Piperno DR (2008) Using plant microfossils from dental calculus to recover human diet: a case study from Tell al-Raqā’i, Syria. J Archaeol Sci 35(7):1943–1950. https://doi.org/10.1016/j.jas.2007.12.005
Herbario Virtual Fitopatología (2023) https://herbariofitopatologia.agro.uba.ar/?page_id=97. Accessed 8 Jan 2023
ICSN (2011) The International Code for Starch Nomenclature. http://fossilfarm.org/ICSN/Code.html. Accessed 15 Nov 2022
Kruse J, Dietrich W, Zimmermann H, Klenke F, Richter U, Richter H, Thines M (2018) Ustilago species causing leaf-stripe smut revisited. IMA Fungus 9(1):49–73. https://doi.org/10.5598/imafungus.2018.09.01.05
Lodish H, Berk A, Matsudaira P, Kaiser C, Krieger M, Scott M, Zipursky SL, Darnell J (2004) Molecular Cell Biology, 5th edn. W. H. Freeman and Company, New York
Louderback LA, Wilks S, Herzog NM, Brown GH, Joyce K, Pavlik BM (2022) Morphometric identification of starch granules from archaeological contexts: diagnostic characteristics of seven major North American plant families. Front Earth Sci 10:897183. https://doi.org/10.3389/feart.2022.897183
Marinangeli CPF (2020) Complementing cereal grains with pulse grains to enhance the nutritional and environmental sustainability profiles of manufactured foods in Canada and the United States. Cereal Foods World 65(6). https://doi.org/10.1094/CFW-65-6-0063
Martí F, Sanmartí E, Viñas R (1975) La cueva de «La Dehesilla» en Jerez de la Frontera. In: Ripoll E, Llongueras M (eds.) XXV Aniversario de los Cursos Internacionales de Prehistoria y Arqueología en Ampurias (1947–1971). Miscelanea Arqueologica. Diputación Provincial de Barcelona, Barcelona, pp 13–47
Mason SLR (1992) Acorns in human subsistence. PhD thesis, University College London
Monge Calleja AM, Santos AL, Pereira Coutinho A (2020) A new method of starch extraction from dental calculus and creation of an identified collection for the reconstruction of paleodiets. Antropol Port 37:99–130. https://doi.org/10.14195/2182-7982_37_5
Morales R, Ocete R (1998) Vitis L. In: Muñoz Garmendia F, Navarro C (eds) Flora Iberica – Plantas vasculares de la Península Ibérica e Islas Baleares, vol. VI – Rosaceae. Real Jardín Botánico, CSIC, pp 58–62
Nesbitt N, Samuel D (1996) From staple crop to extinction? The archaeology and history of the hulled wheats. In: Padulosi S, Hemmer K, Heller J (eds) Hulled wheat, Proceedings of the First International Workshop on Hulled Wheats, Castelvecchio Pascoli, Tuscany, Italy, pp 40–99
Nitiu DS, Mallo AC, Gardella-Sambeth MC, Mrbelli MA (2010) Contribución a la identificación de esporas del Reino Fungi en la atmósfera de La Plata. Argentina Bol Soc Argent Bot 45(3–4):301–308
Oliveira TM, Pirozi MR, Borges JTS, Germani R, Fontes MPF (2009) Caracterização do amido de grão-de-bico (Cicer arietinum L.). Boletim do Centro de Estudos de Pesquisa de Processamento de Alimentos 27(1):27–42
Peña-Chocarro L, Pérez-Jordà G, Morales J (2018) Crops of the first farming communities in the Iberian Peninsula. Quat Int 470B:369–382. https://doi.org/10.1016/j.quaint.2017.06.002
Peña-Chocarro L, Pérez-Jordà G, Morales J, Vera J (2013) … Y llegaron los agricultores: agricultura y recolección en el Occidente del Mediterráneo. Menga: Revista de Prehistoria de Andalucía 4:15–33
Pereira Coutinho A (1939) Flora de Portugal, 2nd edn. Bertrand (Irmãos) Ltd., Lisboa
Pereira Coutinho A (2002) Palinologia das tribos Inuleae Cass. (sensu stricto), Gnaphalieae Benth., Helenieae Benth. e Heliantheae Cass. (Asteraceae) em Portugal Continental e Açores. PhD Thesis, Universidade de Coimbra
Perelló AE, Sisterna MV, Moreno MV (2003) Occurrence of Cladosporium herbarum on wheat leaves (Triticum aestivum) in Argentina. Australas Plant Pathol 32:327–328. https://doi.org/10.1071/AP03019
Perez S, Baldwin PM, Gallant DJ (2009) Structural features of starch granules I. In: BeMiller J, Whistler R (eds) Starch: chemistry and technology, 3rd edn. Academic Press, Amsterdam, pp 149–192
Pimentel G, Carris LM, Peever TL (2000) Characterization of interspecific hybrids between Tilletia controversa and T. bromi. Mycologia 92(3):411–420. https://doi.org/10.2307/3761499
Piperno DR, Weiss E, Holst I, Nadel D (2004) Processing of wild cereal grains in the Upper Palaeolithic revealed by starch grain analysis. Lett Nat 430:670–673. https://doi.org/10.1038/nature02734
Punia S, Dhull SB, Sandhu KS, Kaur M (2019) Faba bean (Vicia faba) starch: structure, properties, and in vitro digestibility – a review. Legum Sci e18. https://doi.org/10.1002/leg3.18
Radini A, Nikita E, Buckley S, Copeland L, Hardy K (2017) Beyond food: the multiple pathways of materials into ancient dental calculus. AJBA 162:71–83. https://doi.org/10.1002/ajpa.23147
Reille M (1992) Pollens et spores d’Europe et d’Afrique du Nord. Laboratoire de Botanique Historique et Palynologie, Marseille
Rózewicz M, Wizínska M, Grabínski J (2021) The most important fungal diseases of cereals - problems and possible solutions. Agronomy 11:714–726
Sáenz-Laín C, Gutiérrez-Bustillo M (2003) Esporas atmosféricas em la comunidad de Madrid. Industria Grafica MAE, Madrid
Seung D (2020) Amylose in starch: towards an understanding of biosynthesis, structure and function. New Phytol 228(5):1490–1504. https://doi.org/10.1111/nph.16858
Singh N, Sandhu KS, Kaur M (2004) Characterization of starches separated from Indian chickpea (Cicer arietinum L.) cultivars. J Food Eng 63(4):441–449. https://doi.org/10.1016/j.jfoodeng.2003.09.003
Smith EG (2000) Sampling and identifying allergenic pollens and molds. Blewstone Press, San Antonio
Sofi BA, Wani IA, Masoodi FA, Saba I, Muzaffar S (2013) Effect of gamma irradiation on physicochemical properties of broad bean (Vicia faba L.) starch. Food Sci Technol 54(1):63–72. https://doi.org/10.1016/j.lwt.2013.05.021
Taylor R, Pérez-Aguilar LG, García-Rivero D (2018) La secuencia arqueológica andalusí (siglos XI-XII) de la Cueva de La Dehesilla (Sierra de Cádiz, España). Arqueol Territ Mediev 25:107–143
Taylor R, García-Rivero D (2020) Evidencias sobre el posible contexto del horizonte impresso en la cueva de la Dehesilla: caracterización cerámica y definición respecto al Neolítico antiguo andaluz tradicional. In: Pardo-Gordo S, Gómez-Bach A, Molist M, Bernabeu J. (eds) Contextualizando la cerámica impressa. Horizontes culturales en la península ibérica. Universitat Autónoma de Barcelona, Barcelona, pp 159–172
Trione EJ (1974) Morphology and cytology of Tilletia caries and Tilletia controversa in axenic culture. Am J Bot 61(8):914–919. https://doi.org/10.2307/2441629
Viñas Vallverdú R (1970) La Cueva de la Dehesilla en Jerez de la Frontera (Cádiz). Actas del I Congreso Nacional de Espeleología, Barcelona, pp 151–153
Viñas Vallverdú R (1971) Prospecciones espeleoarqueologicas en la cueva de La Dehesilla, Jerez de la Frontera. Información Arqueológica 6:176–177
Watt B (2023) Loose smut of wheat or barley (Ustilago tritici). Bugwood.org, University of Maine. https://www.forestryimages.org/browse/detail.cfm?imgnum=5503666. Accessed 18/05/2023
White DJ (1997) Dental calculus: recent insights into occurrence, formation, prevention, removal and oral health effects of supragingival and subgingival deposits. Eur J Oral Sci 105:508–522
Yang C, Perry L (2013) Identification of ancient starch grains from the tribe Triticeae in the North China Plain. J Archaeol Sci 40(8):3170–3177. https://doi.org/10.1016/j.jas.2013.04.004
Zapata L, Peña-Chocarro L, Pérez-Jordá G, Stika HP (2004) Early Neolithic Agriculture in the Iberian Peninsula. J World Prehist 18(4):283–325. https://doi.org/10.1016/j.quaint.2017.06.002
Zarroug Y, Boulares M, Safayhi D, Slimi B, Stiti B, Zaieni K, Nefissi S, Kharrat M (2022) Structural and physicochemical properties of Tunisian Quercus suber L. starches for custard formulation: a comparative study. Polymers 14(3):556. https://doi.org/10.3390/polym14030556
Zheng X, Wang W, Piao H, Xu W, Shi H, Zhao C (2013) The genus Gnaphalium L. (Compositae): phytochemical and pharmacological characteristics. Molecules 18(7):8298–8318. https://doi.org/10.3390/molecules18078298
Zundel JL (1939) Studies on the Ustilaginales of the world. Mycologia 31:572–589
Župunski V, Jevtić R, Ignjatović-Micić D, Stanković S, Lević J, Ivanović D, Nikolić A (2012) Incidence of Tilletia species in non-processed seed of Triticum aestivum in Vojvodina, Serbia. Seed Sci Technol 40(3):320–332. https://doi.org/10.15258/sst.2012.40.3.04
Acknowledgements
The authors wish to thank all members of the archaeological team who participated in the different archaeological campaigns in Dehesilla Cave, namely Araceli Barrera Cruz, Daniela Pacheco, David López Carmona, Elena Trujillo Godoy, Esteban García-Viñas, Javier Pérez González, Jericó Mancera Jiménez, Leonor Vale, Luis Gethsemani Pérez-Aguilar, María Barrera Cruz, Manuel J. Díaz Rodríguez, Pablo Gutiérrez Franco, Patricia Virino Gabella and Ruth Taylor. Finally, we would like to thank the Editors of Archaeological and Anthropological Sciences for their interest in our work. The comments from the two anonymous reviewers helped to improve the initial version of the paper.
Funding
Open access funding provided by FCT|FCCN (b-on). This publication is part of the project ‘NeoProModels: Patterns and processes in the Neolithization of the South of the Iberian Peninsula’, Ref. PID2022-137946NB-I00, supported by MCIN/AEI/10.13039/501100011033/FEDER, UE. The data presented here is provided by the project “Cueva de la Dehesilla: Estudio arqueológico y paleoambiental para el conocimiento de la ocupación humana prehistórica de la Sierra de Cádiz.” These two projects are led by Daniel García-Rivero. Archaeological field seasons also received financial support from several funding programs of the University of Seville, as well as from CIAS, University of Coimbra. Cláudia Umbelino is funded by the Fundação para a Ciência e Tecnologia (FCT, reference UIDB/00283/2020). António Pereira Coutinho was funded by the Fundação para a Ciência e a Tecnologia (FCT, reference UID/BIA/04004/ 2020).
Author information
Authors and Affiliations
Contributions
The dental calculus was extracted by Cláudia Umbelino and prepared by António Pereira Coutinho, and Eduarda Silva. The microstructures were identified by António Pereira Coutinho and Eduarda Silva. Margarida Moreira prepared the figures of starch grains, pollen and fungal spores. António Pereira Coutinho, Cláudia Umbelino and Daniel García-Rivero wrote the original text of the manuscript and revised it.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Coutinho, A.P., Moreira, M., Silva, E. et al. Identification of Neolithic diet by the morphology of the starch grains of dental calculus found in the Dehesilla Cave (Cadiz – South of the Iberian Peninsula). Archaeol Anthropol Sci 16, 62 (2024). https://doi.org/10.1007/s12520-024-01972-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-024-01972-z