
Abstract
Evidence of dog consumption at the El Portalón de Cueva Mayor site (Sierra de Atapuerca, Spain) from the Holocene is revealed for the first time. The taxonomical and taphonomical studies of the animal bones from the El Portalón site have been carried out. The morphological and metrical analyses indicate that 130 dog bone remains have been identified from the El Portalón site, including from the Neolithic (NISP = 23), Chalcolithic (Pre-Bell Beaker Chalcolithic and Bell Beaker Chalcolithic) (26), Early Bronze Age, Middle Bronze Age and Late Bronze Age (81). The anthropic evidence encompasses cut marks, fresh bone fractures, human tooth marks and fire modifications, thus constituting clear evidence of cynophagy, at least in the Chalcolithic and Bronze Age levels in different contexts (habitat and funerary) from the El Portalón site (Atapuerca, Burgos). Furthermore, the fire alterations on two bone remains from the Neolithic suggest likely dog consumption due to the domestic character of the stratigraphical units where these bone remains were found. The taphonomic evidence suggests that domestic dogs were, at least occasionally, part of the diet of the humans who inhabited the El Portalón site, a fact that might be caused either by food shortages and hunger or as dog meat was considered as a delicacy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Nowadays, eating dog meat is considered taboo in many cultures, while it is perfectly acceptable, even considered a delicacy, in other cultures. Cynophagy is a common practice in different regions of the world: Southeast Asia (in the Philippines, there is a typical dish made of dog meat called Asocena), North and South America, the Pacific and in Northern Africa (the Amazigh are known for their custom of eating dogs: Simoons 1991, 1994). On the other hand, some societies (Hindus, Buddhists, Westerners and Muslims) are anti-dog eating for different reasons and Islamic law forbids dog consumption (The Encyclopaedia of Islam 1999) because it is considered an impure animal.
In Europe, there is evidence of cynophagy from the Mesolithic to the Iron Age (Böyönki 1975). Historical evidence for dog eating has been documented in both Rome and Greece (Simoons 1994), but only for medicinal purposes. During the nineteenth and twentieth centuries, in Munich (Germany), dog consumption was related to industrialization and population growth. Dogs were mainly consumed by poor and working-class people due to its lower cost compared with other kinds of meats (Geppert 1992). However, authors like Gautier (1990) believe that dogs are not an important source of protein due to their small size; however, according to Simoons (1994), dog meat contains as much protein and less fat than pork and is very tasty.
Regarding the zooarchaeological evidence, dog consumption in the Iberian Peninsula is documented during the Early Neolithic at the nearby El Mirador archaeological site in the Sierra de Atapuerca (Burgos, Spain) located 1 km from the El Portalón de Cueva Mayor site. It constitutes one of the oldest evidences found in the Iberian Peninsula (5230–4920 cal BC) and the first case where human tooth marks have been used to show dog consumption (Martín et al. 2014). In addition, evidence of cynophagy has been observed in another Iberian Neolithic site, the La Sarsa site (Mediterranean Iberia) (López and Molero 1984); however, this site does not provide a precise chronology. There are more dog remains with evidence of anthropic consumption (cut marks on the surface of the dog bones and fractures in fresh bones) from the Bronze Age period, such as the Lloma de Betxí; Pic del Corbs; the Cabezo Redondo sites in Comunidad Valenciana (Sanchis and Sarrión 2004); Castellón Alto; Terrera del Reloj (Milz 1986); the Cerro de la Encina sites (Friesch 1987) in Granada; the Palacios and Azuer sites in Ciudad Real (Driesch Von Den and Boessneck, 1980); the Gatas site in Almería; and La Bastida in Murcia (Andúgar Martínez 2016) (see Table 1).
Here, we present the taxonomic study of the canids and the taphonomic analysis in terms of the specific aspects of consumption found in the domestic canid remains from Neolithic to Bronze Age archaeostratigraphical sequence of the El Portalón de Cueva Mayor site (Sierra de Atapuerca, Burgos). The objective of the taphonomic study is to characterise the presence of human-induced modifications, carnivore activities and post-depositional signs on dog bones to trace the consumption of these canids from the Neolithic to Bronze Age periods at this site.
The El Portalón de Cueva Mayor archaeological site
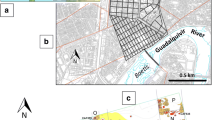
The El Portalón de Cueva Mayor site is located in a cave in the Sierra de Atapuerca (Burgos) on the Northern Plateau of Spain (Fig. 1A). It is the current entrance to the Cueva Mayor-Cueva del Silo karstic system. Prolonged human occupation has been documented, ranging from the Late Pleistocene to the Medieval Age (Clark et al. 1979; Carretero et al. 2008; Pérez-Romero et al. 2010, 2013, 2015). The archaeo-stratigraphic sequence exceeds 10 m of depth and is divided into 11 stratigraphic units, which are in turn grouped into two sedimentary units: the Late Pleistocene (level 11 and level 10, which is the Pleistocene/Holocene transition) and the Holocene. During the Holocene, there is evidence of human occupations in the Neolithic/Mesolithic (level 9); the Chalcolithic (levels 8, 7 and 6); the Early, Middle and Final Bronze Ages (level 5 and levels 3/4); Iron Age I (level 2); the Roman Age (level 1); and the Middle Ages (level 0) (Carretero et al. 2008; Pérez-Romero et al. 2015).

A Geographical location of the El Portalón de Cueva Mayor site. B Archaeo-stratigraphic sequence of the southern section of El Portalón de Cueva Mayor (modified from Pérez-Romero et al. 2016)
Ongoing archaeological excavations are being carried out in the earliest Neolithic layers located in level 9. The time between the Neolithic and Chalcolithic periods has been described as a short hiatus in human occupations of the cave, marked by an archaeologically sterile natural cave entrance and terrigenous silty sediments with abundant pellets. The Chalcolithic stratigraphic units are further divided into three archaeological contexts: the oldest corresponds to a funerary context during the early Pre-Bell Beaker Chalcolithic (level 8), an intermediate phase (level 7) during the Pre-Bell Beaker Chalcolithic includes sheepfold activity and the last unit during the Bell Beaker Chalcolithic (level 6) corresponds to the use of the site for a sheepfold (Pérez-Romero et al. 2017). Finally, the Bronze Age level has been further divided into Early, Middle and Final Bronze Age (levels 3/4 and 5) occupations (Pérez-Romero et al. 2016), all of them from a domestic habitat context (Fig. 1B).
Dating
Two carbonised Triticum sp. grains and two faunal bone remains from different Neolithic stratigraphic units were dated for this work (Table 2). Chalcolithic and Bronze Age stratigraphic units containing dog bone remains with evidence of consumption were previously dated (Carretero et al. 2008; Pérez-Romero et al. 2015) (Table 2). Four animal and human bone remains and one seed from different Chalcolithic stratigraphic units, where dog remains were found, were dated using accelerator mass spectrometry (AMS) at Beta Analytic Inc. (Miami, Florida) and six charcoal remains, one organic sediment sample and one animal bone from the Bronze Age levels were dated using AMS at different radiocarbon dating laboratories (Table 2). Dates were calculated and calibrated to years cal. BP using OxCal v4.4 software based on the IntCal20 radiocarbon age calibration curve.
Materials and methods
The identified faunal sample found in the Neolithic to Bronze Age levels from the El Portalón de Cueva Mayor site comprises 5431 determinable remains, of which 130 (2.4%) are attributed to dog remains.
The dog bones from the Bronze Age were recovered during excavations carried out from 1973 to 1983 by J.M. Apellániz, and were studied at the Museo de Burgos. The Chalcolithic and Neolithic level dog remains were recovered during recent excavations carried out by the current Atapuerca Research Project, specifically in field work conducted from 2007 to 2018. All the bone remains analysed in this work belong to the Colección Museística de Castilla y León of the Junta de Castilla y León.
Anatomical and taxonomic identification
The anatomical and taxonomic identification of the remains from El Portalón was carried out using atlases of animal anatomy (Schmid 1972; Pales and Garcia 1981; Barone 1999) and the collection of comparative anatomy at the Centro Mixto UCM-ISCIII de Evolución y Comportamiento Humanos in Madrid. The dog bones were quantified using the following criteria: NISP (number of identified specimens) (Binford 1978). The age of death was estimated in accordance with criteria for dental eruption and epiphyseal fusion criteria proposed by Schmid (1972) taking account the MNI (minimum number of individuals; Klein and Uribe 1984).
The diagnostic features of canid crania considered in the taxonomical identification were shorter snout, a more pronounced forehead area and a wide palate (Germonpré et al. 2009; Morey 2014; Sablin and Khlopachev 2002). The shortening of the snout in dogs suggests a strong mandible and well-developed carnassial (P4 and M1) (Germonpré et al. 2009), so the measurements of P4 and M1 were taken. Another criterion that is used as evidence of domestication is the crowding of teeth due to the absence of diastema (or shortening of the snout) (Boudadi-Maligne and Escarguel 2014).
The taxonomic identification was completed taking measurements of cranial and postcranial bone remains, to discriminate between Canis lupus lupus and Canis lupus familiaris. All measurements were documented using a Sylvac digital calliper (03.02/SYL-235-F, D, E/681.046–100) to the nearest 0.01 mm, following Driesch (1976).
Dental measurements from the length of M1 (carnassial) and mesio-distal diameter of P4 were taken and included within the metrical variation of teeth from recent and archaeological Eurasian dogs and wolves, which were collected from bibliographic sources: Benecke (1987); Böyönki (1975); Boudadi-Maligne et al. (2012); Chaix (2000); Clark (1995); Clutton-Brock (1962); Dayan (1994); Davis (1978); Davis (2006); Degerbol (1961); Detry and Cardoso (2010); Dimitrijević and Vuković (2015); Gaudry and Boule (1892); Germonpré et al. (2009); Germonpré et al. (2012); Germonpré et al. (2015); Germonpré et al. (2017); Harrison (1973); Huxley (1880); Janssens et al. (2019); Jeteiles (1877); Jolicoeur (1959); Kurtén (1968); Lawrence and Reed (1983); Morey (2014); Musil (2000); Napierala and Uerpmann (2012); Nehring (1888); NMBE database n 2217 dogs; Ovodov et al. (2011); Pidoplichko et al. (2001); Pionnier-Capitan (2010); Pionnier-Capitan et al. (2011); Rütimeyer (1861); Sablin and Khlopachev (2002); Sanchis and Sarrión (2004); Street (2002); Studer (1901); Tchernov and Valla (1997); Wolfgram (1894).
Postcranial measurements were taken and compared with a standard (a complete skeleton of a present-day female wolf from Italy, according to Boschin et al. 2020) using the log ratio methodology. The log ratio technique is used to increase sample size and allow for comparisons to be made between assemblages (Simpson et al. 1960; Meadow 1999). The log ratio is a size index scaling technique, comparing our measurements to the measurements of a standard individual or population.
Taphonomic analysis
In order to conduct the taphonomic analysis, we considered all the domestic dog remains (Canis l. familiaris) recovered from the different archaeological levels at the site: Neolithic (NISP = 23), Chalcolithic (NISP = 26) and Bronze Age (NISP = 81). For the microscopic study, a Nikon SMZ800 Stereoscopic zoom microscope and a DINO-LITE digital microscope were used. Photographs were taken with the digital video microscope DINO-LITE AM-TFVW-A (DinoCapture 2.0 software). The taphonomic study includes anthropic traces, fracture patterns, carnivore modifications and post-depositional alterations.
Stone tool modifications were classified as cut marks (including incisions or slicing cut marks, scrape marks and chop marks) and percussion marks (percussion pits, conchoidal scars and flakes, and adhered flakes) (Blumenschine et al. 1996; Blumenschine and Selvaggio 1988; Cáceres et al. 2007; Capaldo and Blumenschine 1994; Rodríguez-Hidalgo et al. 2015; Saladié et al. 2012; Shipman and Rose 1983). The location of cut marks was recorded—especially the muscle insertion areas or on tendons or ligaments—since these can be used as criteria for distinguishing different butchery activities (Binford 1981; Saladié et al. 2012; White 1992). In order to study the burned bones, we followed the stages defined by Stiner et al. (1995). Evidence of boiling was identified on the basis of the smoother, lighter and more transparent surfaces as opposed to the unboiled bones, as defined by Botella et al. (2000).
In order to analyse the breakage patterns, we focused on long bone fragments following the methodology proposed by Villa and Mahieu (1991) and Sala et al. (2015) in terms of fracture outline (longitudinal, transverse or oblique/curved), fracture angle (right or oblique), fracture edge (smooth or jagged), shaft circumference (1: less than half of the circumference; 2: more than half of the circumference; and 3: complete circumference) and shaft fragment (1: less than 1/4 of the total diaphysis; 2: between 1/4 and 1/2 of the total diaphysis; 3: between 1/2 and 3/4 of the diaphysis; and 4: increased from 3/4 of the diaphysis).
In addition to these features, the presence or absence of peeling was also considered. Peeling is defined as ‘a roughened surface with parallel grooves or fibrous texture’ (White 1992) produced when breaking the bone by bending the pieces with the hands or by cheek-tooth chewing the bones and bending with the hands (Fernández-Jalvo and Andrews 2011; Saladié et al. 2013). Pickering et al. (2013) distinguished three types of peeling: (i) classic peeling following the definition proposed by White (1992); (ii) general peeling defined as ‘an area of the whole dorsal or ventral cortex of a rib is peeled backed for some length, revealing the internal trabeculae of the rib’; and (iii) incipient peeling, a type of peeling ‘where a strip of lamella is only partially peeled back against the rib shaft, not fully removed from the specimen’. Classic and general peeling are considered particular features of anthropogenic breakage (Pickering et al. 2013).
Tooth marks on bone surfaces were classified as pits, punctures, furrowing, scores and dissolution due to gastric acids. Punctures, scores and pits were measured (length and width) in accordance with previous studies (Domínguez-Rodrigo and Piqueras 2003; Sala and Arsuaga 2018; Sala et al. 2014a; Selvaggio and Wilder 2001). The length and breadth of tooth marks were measured using DINO-LITE digital microscope software tools. Tooth mark measurements were compared with neo-taphonomic experimental assemblages of carnivores and humans (Delaney-Rivera et al. 2009; Fernández-Jalvo and Andrews 2011; Sala and Arsuaga 2018; Saladié et al. 2013). Recent studies (Andrés et al. 2012; Sala et al. 2014a, 2014b) show that tooth pit length on cortical surfaces is the best indicator of the taxa responsible for the tooth marks. For this reason, we will focus mainly on this variable when comparing the samples. For the univariate analysis, we performed a Welch test, comparing all possible pairs of samples to determine which differed significantly. Hominin tooth marks were identified following the criteria of Saladié et al. (2013), Fernández-Jalvo and Andrews (2011), Pickering et al. (2013) and Rodríguez Hidalgo et al. (2015), based on their morphological features, location and dimensions and their relationship with other anthropic modifications. In addition to the conspicuous marks, traces of the dissolution caused by gastric acids produced during the consumption of bone fragments are considered. Some carnivores (and also birds) are able to swallow bone fragments. These splinters can be recovered from faeces and they are also the product of regurgitation (Sala and Arsuaga, 2018). Bones affected by acid-etching display characteristic features described previously by Sutcliffe (1970), such as scalloping of the bone surface, presence of holes and/or fine and sharp edges.
Results
Anatomical and taxonomic identification
The M1 lengths of the canids from El Portalón (Table 3) fall within the metrical range of dogs from the different archaeological sites included in Janssens et al. (2019), within a variation ranging from 17 to 25.3 mm for the lower first molar.
The canid P4 lengths from El Portalón (Table 3) also fall within the metrical range of archaeological dogs, with a variation ranging from 14 to 21.8 mm for the upper four premolars. There is no overlap with the P4 mesio-distal diameter (22.5–29.2 mm) from fossil wolves and dogs, according to Janssens et al. (2019).
The data set of the postcranial elements is included in Supplementary Table S1 and Supplementary Figure S1. We can observe that the population of canids from the Neolithic, Chalcolithic and Bronze Age at El Portalón is smaller than the standard population (Supplementary Figure S1); therefore, the canid bone remains from the El Portalón site belong to Canis l. familiaris.
Thus, the morphology and metrical characteristics (Table 3; Supplementary Table S1 and Supplementary Figure S1) indicate that dogs were present in the Neolithic, the Pre-Beaker Chalcolithic (funerary and sheepfold phases), the Beaker Chalcolithic (sheepfold) and the Early, Middle and Final Bronze Age levels at the El Portalón site.
Twenty-three dog remains were recovered from the Neolithic level, 26 from the Chalcolithic (14 from a Pre-Beaker Chalcolithic funerary context, 10 from a Pre-Beaker Chalcolithic stabling context and two from the Beaker Chalcolithic) and 81 from the Bronze Age (28 from the Early Bronze Age, 47 from the Middle Bronze Age and six from the Final Bronze Age) (Table 4). There is no context from which dog remains were recovered where they exceeded 6% of NISP (the identified species) (Table 4).
From the Neolithic, at least two adults (fused anatomical elements), including the burnt phalanx (ATP17.UE536.425), the mandible (ATP15.UE507.392) and one juvenile (unfused elements), were found. From the Pre-Bell Beaker Chalcolithic (sheepfold context), at least two individuals were recovered (one unfused distal part of a tibia: ATPʹ08.UE21.83, suggesting an individual less than 15 months of age, and one fused ulna ATPʹ08.UE23.F10 + ATPʹ08.UE23.101, suggesting an individual older than 15 months of age). From the Pre-Bell Beaker Chalcolithic (funerary context), at least two adult individuals were recovered, which were calculated using left humeri. From the Bell Beaker Chalcolithic habitational context, the humerus found belongs to an individual over 6 months old. From the Early Bronze Age, two cranium fragments (zygomatic bones: CMI-A8-73–10; CMI-B2-79–2) were identified as one juvenile (unfused) and one adult (fused) individual. From the Middle Bronze Age, six maxilla fragments with permanent teeth (CMI-B2-64–2; CMI-C2-63–2; CMI-B4-41; CMI-B2-63–5; ATP18.2003 N.825) were identified and determined to be from at least five adult individuals. A metapodial (CMI-C2-33–2) was recovered from the Final Bronze Age and was fused, so this individual was over 6 months of age. It may belong to the same individual whose other anatomical elements were found in this level (see Table 5).
Taphonomic analysis
Neolithic (Fig. 2)

The Neolithic dog remains from the El Portalón de Cueva Mayor site. Photography and different views of fire alterations on the proximal phalanx ATP17.UE536.425 (A) (photo taken by Javier Trueba and the mandible ATP15.UE507.392 (B)). Scale bars 2 cm
From the Neolithic, a metacarpal III (AT16.UE524.302) has been documented with evidence of gastric acid dissolution (Table 6). One proximal phalanx (ATP17.UE536.425) and a mandible (ATP15.UE507.392) show evidence of burning on the proximal surface of the phalanx and the almost complete surface of the mandible. Following the burning damage categories of Stiner et al. (1995), the phalanx corresponds to stage 1 (less than half the surface is carbonised) and the mandible corresponds to stage 2 (more than half the surface is carbonised).
None of the bone remains from the Neolithic layers displays butchery evidence such as cut marks.
The reduced bone sample, together with the absence of conspicuous tooth marks, necessary for extrapolating useful metric data, hinders the accurate identification of the intervention of small carnivores and/or humans. Nevertheless, the intervention of later carnivores is demonstrated by the presence of bones with evidence of digestion.
Chalcolithic
The dog remain analysis from the Chalcolithic period was conducted in accordance with the archaeological context from which they were recovered: (i) Early (Pre-Bell Beaker) Chalcolithic corresponding to a funerary context; (ii) Early (Pre-Bell Beaker) Chalcolithic corresponding to a sheepfold and habitational context; and (iii) Late (Bell Beaker) Chalcolithic corresponding to a sheepfold. The results of the taphonomic analyses are detailed in Table 6 and Fig. 3.

Anthropic modifications on the Chalcolithic dog bone remains from the El Portalón de Cueva Mayor site. A Cut marks (white arrows) and human tooth marks (black arrow) on the tibia ATP08.UE21.83 from the Pre-Bell Beaker Chalcolithic sheepfold context. B Slicing marks on the humerus ATP10.466 from the Pre-Bell Beaker funerary context. C Slicing marks (white arrows) in combination with human tooth marks (black arrow) and burning (red arrow) on an ulna (ATP08.UE23.F10 + 101). D Peeling (blue arrows) and slicing marks (white arrows) on two ribs from the Pre-Beaker Chalcolithic funerary (ATP10.UE26.287) and sheepfold (ATP10.UE52.15) contexts. Scale bars 2 cm
Within the funerary context from the Pre-Bell Beaker Chalcolithic period, 10 dog remains were analysed and half of them display cut marks (slicing marks). In addition, tooth marks, burning and intentional breakage by humans (i.e. peeling) have been observed on these bone remains (Table 6). Only one bone displays conspicuous tooth marks that provided metric data (Table 7) and triangular-shaped morphology compatible with human tooth marks (Fernández-Jalvo and Andrews 2011; Saladié et al. 2013).
Similar frequencies of cut-marked bones are represented in combination with tooth marks and fresh bone breakage from the Pre-Bell Beaker Chalcolithic sheepfold and habitational contexts (Table 6). The tooth mark dimensions (Table 7), their triangular- or crescent-shaped morphology (Fig. 3), which was described previously as a diagnostic feature for human tooth marks by Fernández-Jalvo and Andrews (2011) and Saladié et al. (2013), together with other anthropic traces, all suggest that they are indeed human tooth marks, indicating that ‘sporadic’ consumption of dogs took place in funerary and sheepfold-habitational contexts during the Early Chalcolithic.
The locations of the cut marks from the Chalcolithic in both cases, the funerary and sheepfold contexts, are mainly concentrated in the diaphysis of long bone and ribs, though slicing marks have been identified additionally in the intertubercular groove of the humerus (Table 6). This indicates that defleshing, periosteum removal, evisceration and disarticulation took place amongst the butchering processes in these periods.
Evidence of fire modification is present on a burned surface of an ulna (ATPʹ08.UE23.101), which appears to be an isolated incident, as well as two dog remains that show characteristics typical of boiling (Table 6).
Within the herding and domestic habitat from the Bell Beaker Chalcolithic period, one bone shows fresh fractures compatible with human activity (Table 6).
Bronze Age
The material from the Bronze Age is the most abundant out of all the sequences. Separate analyses were performed for the three periods represented: the Early, Middle and Final Bronze. These provide frequencies of anthropic traces that are similar throughout the three periods. Cut marks mainly comprise slicing marks, but chop and percussion marks have been documented as well and are present in 25% and 27% of the Early and Middle Bronze Age periods, respectively. Fresh bone fractures, in some cases considered intentional breakage (i.e. peeling), have been documented in Early, Middle and Final Bronze Age periods in 35.7%, 25.5% and 25% of specimens, respectively. Tooth marks are only present in Early and Middle periods with frequencies of 21.4% and 17.6%, respectively, and in some cases are compatible with human tooth marks.
Lastly, by taking into consideration the burned and boiled bones, fire modification has been documented in the three periods as well, with a significantly higher frequency during the Middle Bronze Age period (25%, 81.25% and 25% for the Early, Middle and Final Bronze units, respectively). The detailed data regarding anthropic traces are portrayed in Table 6 and Fig. 4.

Anthropic modifications on the Early Bronze Age (A) and Middle Bronze Age (B) dog remains. A1 Slicing marks (white arrows) on the occipital bone CMI-A8-73–8 where rodent activity is also recorded (green arrow). A2 Innominate bone CMI-D2-104–10 with tooth pits (black arrow) and slicing marks (white arrows). A3 Peeling (blue arrow) in the CMI-D2-104–11 lumbar vertebra. A4 Astragalus CMI-B2-78–9 with partial calcination (red arrow). A5 General and detailed views of the slicing marks on the humerus CMI-C2-92–1 where the typical features of fresh bone (perimortem) breakage can be observed. B1 General and detailed views of the frontal and nasal remain CMI-B2-64–3 where slicing marks (black arrows) and tooth pits (black arrow) are marked. Note the superimposition of the tooth pit on the cut mark. B2 Numerous parallel slicing marks on the rib CMI-B2-64–4. All the scale bars of the microscope views correspond to 1 mm
The cut marks from the Bronze Age have been observed on all anatomical parts and are associated with the removal of soft tissues (flesh, periosteum, viscera and scalp), thus indicating butchering processes such as skinning, defleshing, evisceration and disarticulation (Table 7).
Tooth mark dimensions are represented in Table 7. The dimensions of tooth pits on the cortical surfaces corresponding to the Middle Bronze Age (the best represented period in terms of the number of measurements) do not differ statistically (p < 0.05) from the human tooth mark size data provided by Saladié et al. (2013), Delaney-Rivera et al. (2009), Romero et al. (2016) and Sala and Conard (2016). However, there are statistical differences regarding the large carnivore tooth pits, including for the wild canids (wolves), when compared with the data from Sala and Arsuaga (2018) and Sala et al. (2014a). Nevertheless, in some cases we cannot rule out the secondary intervention of small carnivores, such as foxes or dogs. Actually, in cranial remain CMI-B2-64–3, it is possible to observe a tooth pit (perhaps created by a small carnivore) superimposed on a slicing mark (Fig. 4). Moreover, rodent activity has been documented for one occipital fragment (CMI-A8-73–8).
The fracture patterns are dominated by oblique angles and smooth surfaces, typical of fresh bone breakage (perimortem). Furthermore, in some cases, there are traces of intentional breakage by humans such as peeling and percussion marks (Table 6).
Discussion
The role of dogs throughout Prehistory has changed over time. Dogs probably had considerable symbolic significance in the lives of hunter-gatherers, at least from the Mesolithic onwards and from the Mesolithic onwards (Larsson 1990). This special role is confirmed in the Iberian Peninsula, where, for instance, an almost complete dog skeleton was found close to human skeletons at Cabeço da Arruda, the final Mesolithic Muge shell middens (Portugal), suggesting it was buried intentionally (Detry and Cardoso 2010). From the second half of the 5th millennium cal BC, dogs played a very important role in the symbolic world. Dogs in the Neolithic (Detry and Cardoso 2010) are abundant, mainly in funerary contexts (Moreno-García 2003; Albizuri et al. 2019) (see Fig. 5 for location of the sites). It is in the subsequent Chalcolithic that their occurrence in the archaeological record begins to be abundant enough to draw conclusions as to morphology and dimensions. Dogs in funerary contexts are also very frequent throughout the Chalcolithic and Bronze Ages (see Fig. 5). There is also evidence that they served as draught animals, and on some occasions, they were consumed (Grandal-d’Anglade et al. 2019).

Location of the archaeological sites with dog remains associated with human consumption evidence (red stars) and funerary context (black pins). See Supplementary Table S2 for map legend
Evidences of dog consumption, including just cut and burn marks on dog bone remains, have been identified in European sites from the Upper Paleolithic (Pont d’Ambon; Pionnier-Capitan et al. 2011), the Neolithic and the Bronze Age in Hungary (Vretemark and Sten 2010) and the Iron Age in Slovakia (Chrószcz et al. 2013), the British Isles (Hambleton, 2008) and Gaul (Méniel 2006; Horard-Herbin 2014). In the Iberian Peninsula (Table 1; Fig. 5), these evidences (cut and burn marks) of dog consumption have been identified during the Neolithic (La Sarsa; López and Molero 1984), as well as during the Bronze Age at sites in Valencia (Sanchis and Sarrión 2004), Castellón Alto and Terrera del Reloj (Granada; Milz 1986), Los Palacios and Azuer (Ciudad Real; Driesch Von Den and Boessneck, 1980) and Cerro de La Encina (Granada; Friesch 1987) (Table 1; Fig. 5). Consequently, in the Iberian Peninsula it seems that a change occurred in the role of dogs from the Neolithic to the Bronze Age, since the consumption of dogs was rare during the Neolithic and became more frequent during the Bronze Age.
In the Iberian Peninsula, the Holocene archaeological sites of El Portalón de Cueva Mayor and El Mirador, in the Sierra de Atapuerca (Burgos), are the only ones in which human tooth marks have been documented on dog bone remains (Martín et al. 2014), unlike the rest of the peninsular sites with evidence of anthropic processing of dog bone remains, in which cuts and butchering marks are identified, as well as cremation features. While this last evidence is compatible with ritual offerings, the cases of the Sierra de Atapuerca sites clearly confirm the consumption of dogs at least from the Chalcolithic and probably from the Neolithic as well.
In the case of the Neolithic level from El Portalón, although the fire modifications on the phalanx and mandible do not constitute clear anthropic processing, they do suggest possible human consumption due to the character of the stratigraphical units where these bone remains (UE 507 and UE 536) were found; the burnt dog bones were recovered from activity floors, formed as the result of domestic activities inside the cave, cooking near several fireplaces and other consumed domestic animal bones.
The oldest Neolithic dog bone from the Neolithic level at the El Portalón site was found in UE 536 (Table 2) and dated 7240–7000 cal BP. This bone remain (ATP17.UE536.425) shows fire alteration and pits (Table 6) on its surface and is included within a domestic context. If these alterations are interpreted as indicative of human consumption, this bone, together with another one with similar evidence from the Neolithic at the nearby El Mirador site (MIR19: 7180–6970 cal BP) (Martín et al. 2014), would constitute the oldest Holocene evidence of cynophagy in the Iberian Peninsula.
The consumption of dogs at El Portalón is not decisive in terms of subsistence during the sequence of the El Portalón site, due to the low frequency of dog bones (less than 10% of NISP per level) in comparison with the high percentage of NISP of ovicaprines, bovines and pigs (Table 4) (Galindo-Pellicena et al. 2017, 2019, 2020; Francés-Negro et al. 2021). No faunal remains present pathologies that would indicate a health risk for the humans who lived there (just pathologies in cattle livestock from the Chalcolithic habitat context were detected due to the domestication process: Galindo-Pellicena et al. 2017). In terms of the age of the dogs when they were slaughtered, the vast majority of dogs from El Portalón are adults and not juveniles (11:3). Taking into account the variety and quantity of animals available at the site and the dogs’ age of death (not appropriate for eating, according to the Preclassic Mayan site in Belize, where dogs were deliberately bred and slaughtered for consumption around the first year of life: Clutton-Brock 1994), the dogs do not appear to be an effective meat source. Our observations could indicate sporadic dog consumption at the El Portalón site.
Therefore, the cynophagy in the domestic, funerary and sheepfold context from the Neolithic, Pre-Bell Beaker Chalcolithic, Bell Beaker Chalcolithic and Bronze Age levels at the El Portalón sites could be caused by the provision of extra food at sporadic periods of shortage or starvation (Beech 1995; Murphy 2001) or sporadic periods of dog consumption as a delicacy (Hayden 1990), as proposed in the nearby El Mirador site (Martín et al. 2014).
Nevertheless, in the case of the Pre-Bell Beaker Chalcolithic funerary context from El Portalón, a skeleton of a buried child was recovered with possible evidence of rickets and scurvy (Castilla et al. 2014). Three dog remains (IV and V metatarsals and one first phalanx) in anatomical connection (Pérez-Romero et al. 2017) were also found close to this burial. This possible association could suggest a symbolic offering in this funerary context (Pérez-Romero et al. 2017), with a portion of a dog either buried with the human or related to a ritual of commensalism (Larsson 1990).
The study of the Chalcolithic inhabitants’ dental microwear from El Portalón (García-González et al. 2019) indicates that their diet was based mainly on meat consumption (higher relative frequency of enamel vertical scratches than horizontal ones). In this context, the observation of dog meat consumption at this level is not out of the ordinary. This heavy meat consumption could be due to the higher intensification of animal husbandry at the expense of agriculture, as suggested by García-González et al. (2019), and may be related with the climatic conditions during the Chalcolithic, which were relatively dry, and with a decrease in the woodlands and nitrophilous taxa, together with a slight increase in xeric taxa, documented by Martínez-Pillado et al. (2014). Perhaps, these arid conditions could have increased the frequency of shortage, famine and finally starvation events, making small and not very profitable dogs a necessary resource of food.
Although it is still difficult to give a satisfactory interpretation, it is obvious that dogs were consumed at El Portalón during most of the Holocene, as suggested by the anthropogenic marks on the dog bone surfaces found at the site. And together with El Mirador, cynophagy is confirmed at least from Neolithic to Bronze Age in the Sierra de Atapuerca and by extension in the Iberian Peninsula.
Conclusions
The taxonomical analysis indicates that 130 Canis l. familiaris bone remains were present from the Neolithic (23), Chalcolithic (26) and Bronze Age (81) units at the El Portalón de Cueva Mayor site.
Signs of anthropic activity, such as cut marks, human tooth marks, intentional breakage and fire modifications, suggest sporadic consumption of domestic dogs during the Chalcolithic and Bronze Age and probably during the Neolithic. These anthropic signs have been identified in bone remains recovered from different archaeological contexts, such as domestic, sheepfold and funerary contexts.
These taphonomic evidences suggest that domestic dogs were, at least occasionally, also part of the diet of the humans who inhabited the El Portalón site; fact could be due to shortage, famine or the consideration of dog meat as a delicacy.
The El Portalón de Cueva Mayor and El Mirador Holocene archaeological sites from Sierra de Atapuerca constitute the oldest evidence of cynophagy in the Iberian Peninsula, and record the continued consumption of dogs over an extensive period of time (from the Neolithic to the Bronze Age) in a single region within the Iberian Peninsula.
References
Abril D, Nocete F, Riquelme JA, Bayona MR, Inácio N (2010) Zooarqueología del III Milenio A.N.E.: El barrio metalúrgico de Valencina de la Concepción (Sevilla). Complutum 21(1):87–100
Albizuri S, Nadal J, Martín P, Gibaja JF, Martín A, Esteve X, Oms FX, Martí M, Pou R, López D, Subirà ME (2019) Dogs in funerary contexts during the Middle Neolithic in the northeastern Iberian Peninsula (5th–early 4th millennium BCE). J Archaeol Sci Rep 24:198–207. https://doi.org/10.1016/j.jasrep.2019.01.004
Alférez Delgado F, Molero Gutiérrez G, Bustos Pretel V, Brea López P (1981) Apéndice II: La fauna de macromamíferos. Trabajos de Prehistoria 38:139e145
Andrés M, Gidna AO, Yravedra J, Domínguez-Rodrigo M (2012) A study of dimensional differences of tooth marks (pits and scores) on bones modified by small and large carnivores. Archaeol Anthropol Sci 4:209–219. https://doi.org/10.1007/s12520-012-0093-4
Andúgar Martínez L (2016) Análisis faunístico de los conjuntos calcolítico y argáricos de los yacimientos de Gatas (Turre, Almería) y La Bastida (Totana, Murcia). PhD. Universidad Autónoma de Barcelona, Departamento de Prehistoria
Baquedano MªI, Blanco JF, Alonso P, Álvarez D (2000) El Espinillo: un yacimiento calcolítico y de la Edad del Bronce en las terrazas del Manzanares. Arqueología, Paleontología y Etnografía, 8. Madrid
Barone R (1999). Anatomie Comparee des Mammiferes Domestiques, Tome I and II. Vigot Freges, Paris.
Beech M (1995) A Matter of Taste? Some evidence for the butchery of horses, dogs and bears at the La Tene settlement of Jenisuv Ujezd. In: Blazek J, Meduna P (eds) Archologické výzkumy v severozápdních Čechách v letech 1983–1992. Ustav archeologicke pamatkove pece severozapadnich Cech, Most, pp 165–170
Benecke N (1987) Studies on early dog remains from Northern Europe. J Archaeol Sci 14:31–49. https://doi.org/10.1016/S0305-4403(87)80004-3
Binford LR (1978) Nunamiut Ethnoarchaeology. Academic Press, New York
Binford LR (1981) Bones: ancient men and modern myths. Academic Press, London
Blasco MC, Capilla MLS, Calle J, Robles FJ, González VM, González A (1991) Enterramientos del horizonte protocogotas en el valle del Manzanares. Cuadernos de Prehistoria y Arqueología de la Universidad Autónoma Madrid 18:55–112. https://doi.org/10.15366/cupauam1991.18.004
Blumenschine RJ, Marean CW, Capaldo SD (1996) Blind tests of inter-analyst correspondence and accuracy in the identification of cut marks, percussion marks, and carnivore tooth marks on bone surfaces. J Archaeol Sci 23:493–507. https://doi.org/10.1006/jasc.1996.0047
Blumenschine RJ, Selvaggio MM (1988) Percussion marks on bone surfaces as a new diagnostic of hominid behaviour. Nature 333:763–765. https://doi.org/10.1038/333763a0
Boschin F, Bernardini F, Pilli E, Vai S, Zanolli C, Tagliacozzo A, Fico R, Fedi M, Corny J, Dreossi D, Lari M, Modi A, Vergata C, Tuniz C, Moroni A, Boscato P, Caramelli D, Ronchitelli A (2020) The first evidence for late Pleistocene for Late Pleistocene dogs in Italy. Sci Rep 10:13313. https://doi.org/10.1038/s41598-020-69940-w
Botella M, Alemán I, Jiménez SA (2000) Los huesos humanos. Manipulación y alteración. Bellaterra, Barcelona
Boudadi-Maligne M, Escarguel G (2014) A biometric re-evaluation of recent claims for Early Upper Palaeolithic wolf domestication in Eurasia. J Archaeol Sci 45:80–89. https://doi.org/10.1016/j.jas.2014.02.006
Boudadi-Maligne M, Mallye J-B, Langlais M, Barshay-Szmidt C (2012) Magdalenian dog remains from Le Morin rock-shelter (Gironde, France). Socio-economic implications of a zootechnical innovation. PALEO Rev. Archeol. Prehistorique 39–54. https://doi.org/10.4000/paleo.2465
Böyönki S (1975) Vlasac: an early site of dog domestication. North Holland Publishing Company.
Burgos A, Pérez C, Lizcano R (2001a) Actuación arqueológica realizada en la piscina comunitaria de los bloques A1, A2, A3, A6, A7, y A8 del sector UA-23 de Marroquíes Bajos de Jaén, Anuario arqueológico de Andalucía, Actividades de urgencia 1998 3(1):402–413
Burgos A, Pérez C, Lizcano R (2001b) Actuación arqueológica realizada en el espacio destinado a la instalación del ovoide del vial 4 de la UA-23, Marroquíes Bajos. Jaén, Anuario Arqueológico de Andalucía 1998 3(1):422–428
Cáceres I, Lozano M, Saladié P (2007) Evidence for Bronze Age cannibalism in El Mirador Cave (Sierra de Atapuerca, Burgos, Spain). Am J Phys Anthropol 133:899–917. https://doi.org/10.1002/ajpa.20610
Capaldo SD, Blumenschine RJ (1994) A quantitative diagnosis of notches made by hammerstone percussion and carnivore gnawing on bovid long bones. Am Antiq 59:724–748. https://doi.org/10.2307/282345
Carretero JM, Ortega AI, Juez L, Pérez-González A, Arsuaga JL, Pérez R, Ortega MC (2008) A Late Pleistocene-Early Holocene archaeological sequence of Portalón de Cueva Mayor (Sierra de Atapuerca, Burgos, Spain). Munibe Antropologia-Arkeologia 5:67–80
Castilla M, Carretero JM, Gracia A, Arsuaga JL (2014) Evidence of rickets and/or scurry in a complete. Chalcolithic Child Skeleton from the El Portalón Site (Sierra de Atapuerca, Spain). Jass Reports 92:1e16
Chaix L (2000) A preboreal dog from the northern Alps (Savoie, France). BAR Int Ser 889:49–60
Chrószcz A, Janeczek M, Bielichová Z, Gralak T, Onar V (2013) Cynophagia in the Púchov (Celtic) culture settlement at Liptovská Mara Northern Slovakia. Int J Osteoarchaeol 22(2):145–157. https://doi.org/10.1002/oa.2320
Clark KM (1995) The later prehistoric and protohistoric dog: the emergence of canine diversity. Archaeozoologia 7:9–32
Clark G, Straus LG, Burton SS, Jackson-Clark V (1979) The North Burgos Archaeological Survey: an inventory of cultural remains, in CLARK, G. (Ed.). The North Burgos Archaeological Survey. Bronze and Iron Age archaeology on the Meseta del Norte (Province of Burgos, North-Central Spain) 19, 18 156. Arizona State University. Dept. of Anthropology.
Clutton-Brock J (1962) Near Eastern canids and the affinities of the Natufian dogs. Z Tierzuchtung Zuchtungsbiologie 76:326–333
Clutton-Brock J (1994) Hot dogs: comestible canids in Preclassic Maya culture at Cuello, Belize. J Archaeol Sci 21:819–826
Davis SJM (1978) Evidence for domestication of the dog 12.000 years ago in the Natufian of Israel. Nature 276:608–610. https://doi.org/10.1038/276608a0
Davis SJM (2006) Faunal remains from Alcáçova de Santarém, Portugal. Trabalhos de Arqueologia, 43. Instituto português de arqueologia. Lisboa.
Dayan T (1994) Carnivore diversity in the Late Quaternary of Israel. Quat Res 41:343–349. https://doi.org/10.1006/qres.1994.1039
Daza-Perea A (2011) Los depósitos de Perros. In: Blasco C, Liesau C, Ríos P (eds) Yacimientos calcolíticos con campaniforme de la región de Madrid. Patrimonio Arqueológico, 6. Universidad Autónoma Madrid 211–222
Daza-Perea A (2015) La fauna en el Calcolítico de la Región de Madrid: Los depósitos de canes. Másteres de la UAM. Año Académico 2011–2012 (Colección de Trabajos Fin de Máster para publicación digital), UAM Ediciones
Daza-Perea A (2017) Preliminary Studies of Late Prehistoric Dog (Canis lupus f. familiaris Linnaeus, 1758) Remains from the Iberian Peninsula: Osteometric and 2D Geometric Morphometric Approaches. Papers from the Institute of Archaeology 27(1):1–21. https://doi.org/10.5334/pia-487
Degerbol M (1961) Der Hund, das alteste Haustier Danemarks. Z Tierzuchtung Zuchtungsbiologie 76:334–341
Delaney-Rivera C, Plummer TW, Hodgson JA, Forrest F, Hertel F, Oliver JS (2009) Pits and pitfalls: taxonomic variability and patterning in tooth mark dimensions. J Archaeol Sci 36:2597–2608. https://doi.org/10.1016/j.jas.2009.08.001
Detry C, Cardoso JL (2010) On some remains of dog (Canis familiaris) from the Mesolithic shell middens of Muge, Portugal. J Archaeol Sci 37:2762–2774. https://doi.org/10.1016/j.jas.2010.06.011
Díaz del Río P (2001) La formación del paisaje agrario: Madrid en el III y II milenio B.C. Arqueología, Paleontología y Etnografía, 9.
Díaz-Andreu M, Liesau C, Castaño A (1992) El poblado calcolítico de Loma de Chiclana (Vallecas, Madrid). Excavaciones de urgencia realizadas en 1987. Arqueología, Paleontología y Etnografía 3, Madrid 31–116
Dimitrijević V, Vuković S (2015) Was the dog locally domesticated in the Danube Gorges? Morphometric study of dog cranial remains from four Mesolithic-Early Neolithic archaeological sites by comparison with contemporary wolves. Int J Osteoarchaeol 22:12–34. https://doi.org/10.1002/oa.2260
Domínguez-Rodrigo M, Piqueras A (2003) The use of tooth pits to identify carnivore taxa in tooth-marked archaeofaunas and their relevance to reconstruct hominid carcass processing behaviours. J Archaeol Sci 30:1385–1391. https://doi.org/10.1016/S0305-4403(03)00027-X
Driesch A (1976) A guide to the measurement of animal bones from archaeological sites: as developed by the Institut für Palaeoanatomie. Domestikationsforschung und Geschichte der Tiermedizin of the University of Munich, Peabody Museum of Archaeology and Ethnology, Harvard University
Driesch A Von Den, Boessneck J (1980) “Die Motillas von Azuer und Los Palacios (Prov. Ciudad Real) Untersuchung der Tierknochenfunde”. Studien über frühe Tierknochenfunde von der Iberischen Halbinsel, 7, München, pp. 84–121
Fabián García J, Blanco González A (2012) Cuatro enterramientos calcolíticos en hoyo del Cerro de la Cabeza (Ávila). Complutum 23(1):99–120
Fernández-Jalvo Y, Andrews P (2011) When humans chew bones. J Hum Evol 60:117–123. https://doi.org/10.1016/j.jhevol.2010.08.003
Francés-Negro M, Iriarte E, Galindo-Pellicena MA, Gerbault P, Carrancho A, Pérez-Romero A, Arsuaga JL, Carretero JM, Roffet-Salque M (2021) Neolithic to Bronze Age economy and animal management revealed using analyses lipid residues of pottery vessels and faunal remains at El Portalón de Cueva Mayor (Sierra de Atapuerca, Spain). J Archaeol Sci 131:105380. https://doi.org/10.1016/j.jas.2021.105380
Friesch K (1987) “Die Tierknochenfunde von Cerro de la Encina bei Monachil, provinz Granada (Grabungen 1977–1984)”. Studien über frühe Tierknochenfunde von der Iberischen Halbinsel, 11, München, 135
Galindo-Pellicena MA, Arsuaga JL, Carretero JM (2019) Pig management in the Chalcolithic and Bronze Age periods at the El Portalón site: integration in the Iberian context. Quatern Int 515:138–149. https://doi.org/10.1016/j.quaint.2019.02.041
Galindo-Pellicena MA, Martín-Francés L, Gracia A, De Gaspar I, Arsuaga JL, Carretero JM (2017) Evidences of the use of cattle as draught animals in Chalcolithic of El Portalón (Sierra de Atapuerca, Burgos). Quatern Int 438:1–10. https://doi.org/10.1016/j.quaint.2015.11.052
Galindo-Pellicena MA, Arsuaga JL, Pérez Romero A, Iriarte E, De Gaspar I, Carretero JM (2020) Metrical analysis of bovine bone remains from the Neolithic to the Bronze Age at the El Portalón site (Atapuerca, Burgos) in the Iberian context. Quatern Int 566–567:211–223. https://doi.org/10.1016/j.quaint.2020.06.001
García Blánquez LA, Martínez C, Ponce J (2002) Excavaciones arqueológicas en la Glorieta de San Vicente (Lorca). Resumen de las XIII Jornadas de Patrimonio Histórico y Arqueología Regional: 20-22. Dirección General de Cultura, Servicio de Patrimonio Histórico de Murcia
García Blánquez LA, Martínez Sánchez C (2004) Intervención arqueológica en Casa Noguera (Archivel, Caravaca de la Cruz). Memorias de Arqueología, 12. Murcia 235–522
García-Moncó J (2008) De Brennan a Bogart. Un mayor papel protagonista para el perro entre las primeras sociedades productoras de la Península Ibérica. IV Congreso Del Neolítico Peninsular (alicante. 2007. 411–417.
García RL, Sánchez F, Vicente A, Rojas JM, Pérez J (2008) El yacimiento Calcolítico “Valladares I (P.K. 0 + 150)” de Illescas (Toledo): Asentamiento Calcolítico en el valle medio del Tajo. ARSE: Boletín anual del Centro Arqueológico Saguntino 42:127–152
García-González R, Sánchez-Puente Z, Arsuaga JL, Carretero JM (2019) Dietary inferences from dental microwear patterns in Chalcolithic populations from the Iberian Peninsula: the case of El Portalón de Cueva Mayor (Sierra de Atapuerca, Burgos, Spain) and El Alto de la Huesera (Álava, Spain). Archaeol Anthropol Sci 11:3811–3823. https://doi.org/10.1007/s12520-018-0711-x
Gaudry A, Boule M (1892) Les oubliettes de Gargas. Mater Hist Temps Quaternaires 4:130–154
Gautier A (1990) La Domestication, et l’Homme créa ses Animaux. Errance, Paris
Geppert P (1992) Dog slaughtering in Germany in the 19th and 20th centuries with special consideration of the Munich area. Berl Munch Tierarztl Wochenschr 105(10):335–342
Germonpré M, Sablin MV, Stevens RE, Hedges RE, Hofreiter M, Stiller M, Despres VR (2009) Fossil dogs and wolves from Palaeolithic sites in Belgium, the Ukraine and Russia: osteometry, ancient DNA and stable isotopes. J Archaeol Sci 36:473–490. https://doi.org/10.1016/j.jas.2008.09.033
Germonpré M, Lazničkova-Galetova M, Sablin MV (2012) Palaeolithic dog skulls at the Gravettian Předmosti site, the Czech Republic. J Archaeol Sci 39:184–202. https://doi.org/10.1016/j.jas.2011.09.022
Germonpré M, Lazničkova-Galetova M, Losey RJ, Raikkonen J, Sablin MV (2015) Large canids at the Gravettian Předmosti site, the Czech Republic: the mandible. Quat Int 359:261–279. https://doi.org/10.1016/j.quaint.2014.07.012
Germonpré M, Fedorov S, Danilov P, Galeta P, Jiminez E-L, Sablin M, Losey R (2017) Palaeolithic and prehistoric dogs and Pleistocene wolves from Yakutia: identification of isolated skulls. J Archaeol Sci 43:1–19. https://doi.org/10.1016/j.jas.2016.11.008
Grandal-d’Anglade A, Albizuri S, Nieto A, Majó T, Agustí B, Alonso N, Antolín F, López JB, Moya A, Rodríguez A, Palomo A (2019) Dogs and foxes in Early-Middle Bronze Age funerary structures in the northeast of the Iberian Peninsula: human control of canid diet at the sites of Can Roqueta (Barcelona) and Minferri (Lleida). Archaeol Anthropol Sci 11:3949–3978. https://doi.org/10.1007/s12520-019-00781-z
Günther T, Valdiosera C, Malmström H, Ureña I, Rodríguez-Varela R, Osk Sverrisdóttir O, Daskalaki EA, Skoglund P, Naidoo T, Svensson EM, Bermúdez de Castro JM, Carbonell E, Dunnj M, Storåe J, Iriarte E, Arsuaga JL, Carretero JM, Götherström A, Jakobsson M (2015) Ancient genomes link early farmers from Atapuerca in Spain to modern-day Basques. PNAS. https://doi.org/10.1073/pnas.1509851112
Hain FH (1982) Kupferzeitliche Tierknochenfunde aus Valencina de la Concepción, Seville. In Studien über frühe Tierknochenfunde von der Iberischen Halbinsel, 8. München. Institut für Palaeoanatomie der Universität München
Hambleton E (2008) Review of Middle Bronze Age-Late Iron Age faunal assemblages from southern Britain. Research department report series N’71. Archaeological Science-English Heritage
Harrison D (1973) Some comparative features of the skull of wolves (Canis lupus Linn.) and pariah dogs (Canis familiaris Linn.) from the Arabian Peninsula and neighbouring lands. Bonner Zoologische Beitrage 185–191.
Hayden B (1990) Nimrods, piscators, pluckers, and planters: the emergence of food production. J Anthropol Archaeol 9:31e69. https://doi.org/10.1016/0278-4165(90)90005-X
Horard-Herbin M-P (2014) La viande de chien à l’Âge du Fer: Quels individus pour quelles consommations? CNRS, Revue Gallia
Hurtado Pérez V (1991) Informe de las excavaciones de urgencia en La Pijotilla. Campaña de 1990, Extremadura Arqueológica, 2 (Ejemplar dedicado a I Jornadas de Prehistoria y Arqueología en Extremadura): 45–68
Huxley TH (1880) On the cranial and dental characters of the Canida. Proc. Zool. Soc. Lond. Wiley Online Libr. 238–288.
Janssens L, Perri A, Crombé P, Van Dongen S, Lawler D (2019) An evaluation of classical morphologic and morphometric parameters reported to distinguish wolves and dogs. J Archaeol Sci Rep 23:501–533. https://doi.org/10.1016/j.jasrep.2018.10.012
Jeteiles L (1877) Die Stammvater unserer Hunderassen, Verlag der Wallishausser’schen Buchhandlung (Josef Klemm), Wien
Jolicoeur P (1959) Multivariate geographical variation in the wolf Canis lupus L. Evolution 283–299.
Klein RG, Uribe K (1984). The analysis of animal bones from archaeological sites. In: Butzer KW, Freeman LG (Eds.), Prehistoric archeology and ecology series. The University of Chicago press, Chicago
Kurtén B (1968) Pleistocene mammals of Europe. Weindefield and Nicolson, London
Larsson L (1990) Dogs in fraction - symbols in action. In: Vermeersch P, Van Peer P (eds) Fourth International Symposium “The Mesolithic in Europe.” Leuven University Press, Leuven, pp 153–160
Lawrence B, Reed CA (1983) The dogs of Jarmo, prehistoric archeology along the Zagros Flanks. University of Chicago Oriental Institute, Chicago, pp 485–489
Liesau C, Esparza A, Sánchez A (2014) ¿Huesos en la basura o depósito ritualizado? Los perros descuartizados de La Huelga (Dueñas, Palencia) Zephyrus, LXXIV: 89–115. https://doi.org/10.14201/zephyrus20147489115
Liesau C, Vega J, Daza A, Ríos P, Menduiña R, Blasco C (2013–2014) Manifestaciones simbólicas en el acceso Noreste del Recinto 4 del Foso en Camino de las Yeseras (San Fernando de Henares, Madrid). Saldvie: Estudios de prehistoria y arqueología 13–14:53–69
Lizcano Prestel R, Cámara Serrano JA (2003) Producción económica y sedentarización. El registro arqueológico del Polideportivo de Martos (Jaén). In: II Jornadas temáticas Andaluzas de Arqueología. Resúmenes. Ronda: 229–248.
López P, Molero G (1984) Análisis de los restos vegetales, faunísticos y polínicos del yacimiento de la Sarsa (Bocairent, Valencia) Trabajos de Prehistoria 4, 305–312.
Martín P, Saladié P, Nadal J, Vergés JM (2014) Butchered and consumed: small carnivores from the Holocene levels of El Mirador cave (Sierra de Atapuerca, Burgos, Spain). Quatern Int 353(5):153–169. https://doi.org/10.1016/j.quaint.2014.08.011
Martínez-Valle R (1993) La fauna de vertebrados. En Bernabeu, J. (ed.) El III milenio A.C. en el País Valenciano. Los poblados de Jovades (Cocentaina, Alacant) y Arenal de la Costa (Ontinyent, València). Saguntum (PLAV) 26:123–52
Martínez-Pillado V, Aranburu A, Arsuaga JL, Ruiz-Zapata B, Gil-García MJ, Stoll H, Yusta I, Iriarte E, Carretero JM, Edwards L, Cheng H (2014) Upper Pleistocene and Holocene palaeoenvironmental records in Cueva Mayor karst (Atapuerca, Spain) from different proxies: speleothem crystal fabrics, palynology, and archaeology. Int J Speleol 43:1–14
Meadow R. (1999) The use of size index scaling techniques for research on archaeozoological collections from the Middle East. In: Becker, C., Manhart, H., Peters, J., Schibler, J. (Eds.), Historiaanimalium ex ossibus. Festschrift für Angela von den Driesch. VerlagmarieLeidorf GmbH, Rahden/Westf, pp. 285–300
Méniel P (2006) Le chien en Gaule, Animali tra uomini e dei, archeozoologia del mondo preromano, ed. Ante Quem, Studi e Scavi 14:45–52
Milz H (1986) “Die Tierknochenfunde aus drei argarzeitlichen siedlungen in der provinz Granada (Spanien)”. Studien über frühe Tierknochenfunde von der Iberischen Halbinsel, 10, München, 149 p.
Moreno-García M (2003) Estudo dos restos faunísticos da Anta 3 da Herdade de Santa Margarida (Reguengos de Monsaraz). In: Gonçalves VS (ed) STAM-3, a Anta 3 da Herdade de Santa Margarida (Reguengos de Monsaraz). Instituto Português de Arqueologia, Lisbon, pp 432–439
Morey DF (2014) In search of Paleolithic dogs: a quest with mixed results. J. Archaeol. Sci. 52, 300–307 (300e307. 301). https://doi.org/10.1016/j.jas.2014.08.015
Murphy EM (2001) Medieval and Post-Medieval butchered dogs from Carrickfergus, Co., Antrim, Northern Ireland. Environ Archaeol 6:13–22
Musil R (2000) Domestication of wolves in Central European Magdalenian sites. BAR Int Ser 889:21–28
Napierala H, Uerpmann HP (2012) A ‘new’ Palaeolithic dog from Central Europe. Int J Osteoarchaeol 22:127–137. https://doi.org/10.1002/oa.1182
Nehring A (1888) Zur Abstammung der Hunde-Rassen, Zoologische Jahrbucher. Abt Syst Geogr Biol Tiere 3:51–96
Oliva M, Terrats N (2005) Cited by Albizuri 2011 El jaciment arqueológic de Can Roqueta/Torre Romeu (Sabadell, Vallès Occidental) Campanya 2002–2003. Excavation memory deposited in the Servei d’Arqueologia de la Generalitat de Catalunya. Unpublished
Ovodov ND, Crockford SJ, Kuzmin YV, Higham TF, Hodgins GW, van der Plicht J (2011) A 33,000-year-old incipient dog from the Altai Mountains of Siberia: evidence of the earliest domestication disrupted by the Last Glacial Maximum. PLoS One 6:e22821. https://doi.org/10.1371/journal.pone.0022821
Pales L, Garcia MA (1981) Atlas osteologiques pour servir a l’identification des mammiferes du Quaternaire. CNRS, Paris
Pérez-Romero A, Carretero JM, Alday A, Arsuaga JL (2013) La Cerámica Protohistórica e Histórica en el yacimiento de El Portalón de Cueva Mayor, Sierra de Atapuerca, Burgos. Boletín de la Sociedad Española de Cerámica y Vidrio 52(4):183e193
Pérez-Romero A, Carretero JM, Juez L, Ortega AI, Ortega MC, Arsuaga JL (2010) Una dobla almohade del Siglo XIII en el yacimiento de El portalón de Cueva Mayor, Sierra de Atapuerca (Burgos). Nvmisma 254:85e106
Pérez-Romero A, Carretero JM, Alday A, Galindo-Pellicena MA, Adán G, Juez L, Arsuaga JL (2015) La gestión del utillaje óseo de la Edad del Bronce en el yacimiento de El Portalón de Cueva Mayor, sierra de Atapuerca, Burgos. Complutum 26:113–131
Pérez-Romero A, Alday A, Iriarte E, Francés-Negro M, Galindo-Pellicena MA, Álvarez-Fernández A, Juez L, Arsuaga JL, Carretero JM (2016) La cerámica de la Edad del Bronce en el yacimiento de El Portalón de Cueva Mayor (Sierra de Atapuerca, Burgos, España). Munibe 67:105–126. https://doi.org/10.21630/maa.2016.67.06
Pérez-Romero A, Iriarte E, Galindo-Pellicena MA, García-González R, Rodríguez L, Castilla M, Francés M, Santos E, Valdiosera C, Arsuaga JL, Alday A, Carretero JM (2017) An unusual Pre-Bell Beaker Copper Age cave burial context from El Portalón de Cueva Mayor site (Sierra de Atapuerca, Burgos). Quat Int 433:142–155. https://doi.org/10.1016/j.quaint.2015.06.063
Pickering TR, Domínguez-Rodrigo M, Heaton JL, Yravedra J, Barba R, Bunn HT, Musiba C, Baquedano E, Diez-Martín F, Mabulla A, Brain CK (2013) Taphonomy of ungulate ribs and the consumption of meat and bone by 1.2-million-year-old hominins at Olduvai Gorge, Tanzania. J Archaeol Sci 40:1295–1309. https://doi.org/10.1016/j.jas.2012.09.025
Pidoplichko I, Allsworth-Jones P, Djindjian F (2001) Upper Paleolithic dwellings of mammoth bones in the Ukraine. In: British Archeological Reports (BAR). International Series 712 JSTOR.
Piña A, Saña M (2004) Cited by Albizuri 2011 Análisi arqueozoològica de restes de fauna recupertas al jaciment de Can Roqueta/Torre-Romeu (Sabadell, Vallès Occidental). Campanya 2002/2003. Excavation memory deposited in the Servei d’Arqueologia de la Generalitat de Catalunya. Unpublished.
Pionnier-Capitan M (2010) La domestication du chien en Eurasie: etude de la diversite passee, approches osteoarcheologiques, morphometriques et paleogenetiques. Ecole normale superieure, Lyon (Sciences).
Pionnier-Capitan M, Bemilli C, Bodu P, Celerier G, Ferrie J-G, Fosse P, Garcia M, Vigne J-D (2011) New evidence for Upper Palaeolithic small domestic dogs in South-Western Europe. J Archaeol Sci 38:2123–2140. https://doi.org/10.1016/j.jas.2011.02.028
Quero Castro S (1982) El poblado del Bronce Medio de Tejar del Sastre (Madrid) Estudios de prehistoria y arqueología madrileñas, 1: 183–248.
Rodríguez-Hidalgo A, Saladié P, Ollé A, Carbonell E (2015) Hominin subsistence and site function of TD10.1 bone bed level at Gran Dolina site (Atapuerca) during the late Acheulean. J Quat Sci 30:679–701. https://doi.org/10.1002/jqs.2815
Román Rodríguez JM, Conlin Hayes E (2001) Excavaciones arqueológicas de urgencia en la calle Dolores Quintanilla nº 6. Carmona (Sevilla), Anuario arqueológico de Andalucía 1997, 3: 529–535
Romero AJ, Diez C, Saladié P (2016) Mammal bone surface alteration during human consumption: An experimental approach. Journal of Archaeological Science: Reports 8:82–89
Rütimeyer L (1861) Die Fauna der Pfahlbauten der Schweiz. Geschichte der Wilden und der Haus-Saugetieren. Neue Denkschriften der Algemeinen Schweizerischen Geselschaft fur die Gesammten. Naturwissenschaften 19:97–143
Sablin M, Khlopachev G (2002) The earliest ice age dogs: evidence from Eliseevichi 11. Curr. Anthropol. 43, 795–799 ( 10.1086/34)
Sala N, Arsuaga JL (2018) Regarding Beasts and Humans: a review of taphonomic works with living carnivores. Quatern Int 466:131–140. https://doi.org/10.1016/j.quaint.2016.03.011
Sala N, Conard NJ (2016) Taphonomic analysis of the hominin remains from Swabian Jura. Implications for the mortuary practices during the Upper Paleolithic in Southwestern Germany. Quatern Sci Rev 150:278–300. https://doi.org/10.1016/j.quascirev.2016.08.018
Sala N, Arsuaga JL, Haynes G (2014a) Taphonomic comparison of bone modifications caused by wild and captives wolves (Canis lupus). Quatern Int 330:126–135. https://doi.org/10.1016/j.quaint.2013.08.017
Sala N, Arsuaga JL, Martínez I, Gracia-Téllez A (2014b) Carnivore activity in the Sima de los Huesos (Atapuerca, Spain) hominin sample. Quatern Sci Rev 97:71–83. https://doi.org/10.1016/j.quascirev.2014.05.004
Sala N, Arsuaga JL, Martínez I, Gracia-Téllez A (2015) Breakage patterns in Sima de los Huesos (Atapuerca, Spain) hominin sample. J Archaeol Sci 55:113–121. https://doi.org/10.1016/j.jas.2015.01.002
Saladié P, Huguet R, Rodríguez-Hidalgo A, Cáceres I, Esteban-Nadal M, Arsuaga JL, Bermúdez de Castro JM, Carbonell E (2012) Intergroup cannibalism in the European Early Pleistocene: the range expansion and imbalance of power hypotheses. J Hum Evol 63:682–695. https://doi.org/10.1016/j.jhevol.2012.07.004
Saladié P, Rodríguez-Hidalgo A, Díez C, Martín-Rodríguez P, Carbonell E (2013) Range of bone modifications by human chewing. J Archaeol Sci 40:380–397. https://doi.org/10.1016/j.jas.2012.08.002
Sánchez A, Rueda C, Bellón JP (2005) Nuevos datos sobre la Zona Arqueológica de Marroquíes Bajos: el quinto foso. Trab Prehist 62(2):151–164
Sanchis A, Sarrión I (2004) Restos de cánidos (Canis familiaris ssp.) en yacimientos valencianos de la Edad del Bronce. Archivo De Prehistoria Levantina 25:161–198
Schmid E (1972) Atlas of animal bones: for prehistorians, archaeologists and quaternary geologists. Elsevier Science Ltd., Amsterdam
Selvaggio MM, Wilder J (2001) Identifying the involvement of multiple carnivore taxa with archaeological bone assemblages. J Archaeol Sci 28:465–470. https://doi.org/10.1006/jasc.2000.0557
Shipman P, Rose J (1983) Early hominid hunting, butchering, and carcass-processing behaviors: approaches to the fossil record. J Anthropol Archaeol 2:57–98. https://doi.org/10.1016/0278-4165(83)90008-9
Simoons FJ (1991) Food in China: a cultural and historical inquiry. CRC Press, Boston
Simoons FJ (1994) Eat not this flesh: food avoidances from prehistory to the present, 2nd edn. University of Wisconsin Press, Madison
Simpson GG, Roe A, Lewontin RC (1960) Quantitative zoology. Harcourt Brace, New York
Stiner MC, Weiner S, Bar-Yosef O, Kuhn SL (1995) Differential burning, recrystallization, and fragmentation of archaeological bone. J Archaeol Sci 22:223–237. https://doi.org/10.1006/jasc.1995.0024
Street M (2002) Ein wiedersehen mit dem hund von Bonn-Oberkassel. Bonner Zool Beitr 50:269–290
Studer T (1901) Die prahistorischen Hunde in ihrer Beziehung zu den gegenwartig lebenden Rassen. Zurcher und Furrer, Zurich
Sutcliffe AJ (1970) Spotted hyaena: crusher, gnawer, digester and collector of bones. Nature 227:1110–1113
Tchernov E, Valla FF (1997) Two new dogs, and other Natufian dogs, from the southern Levant. J Archaeol Sci 24:65–95. https://doi.org/10.1006/jasc.1995.0096
Valera AC (2008) O recinto calcolítico dos Perdigões: fossos e fossas do Sector I. Apontamentos de Arqueologia e Património 3:19–27
Villa P, Mahieu E (1991) Breakage patterns of human long bones. J Hum Evol 21:27–48. https://doi.org/10.1016/0047-2484(91)90034-S
Vretemark M , Sten S (2010) Skeletal manipulations of dogs at the Bronze Age site of Százhalombatta-Földvár in Hungary. In: D. Campana, P. Crabtree, S.D. de- France, J. Lev-Tov, and A.M. Choyke, editors, Anthropological approaches to zooarchaeology: colonialism, complexity and animal transformations, Oxford, Oxbow 274:2010–2012.
White T (1992) Prehistoric cannibalism at Mancos 5Mtumr-2346. Princeton University Press, Princeton
Wolfgram A (1894) Die Einwerkung der Gefangenschaft auf die Gestaltung des Wolfschadels. Zool Jahrb (abt Syst) 7:773–822
Acknowledgements
We extend our sincerest gratitude to the team at the UCM-ISCIII Centre of Human Evolution and Behaviour, Fundación Atapuerca, and at the Laboratory of Human Evolution at the University of Burgos (UBU) for their technical support throughout our research and excavation work. Special thanks are also due to the Portalón team for their support and efforts throughout the course of the fieldwork. We are very grateful to Marta Negro Cobo, Director of the Museum of Burgos, for providing us access to the archaeological samples from the prior Portalón excavations.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This study was financed by the Ministerio de Economía y Competitividad, Spain Project PGC2018-093925-B-C33 (MCIU/AEI/ FEDER, UE) and Quaternary Ecosystems-UCM Research Group. NS has received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation program (Grant agreement No. 949330) and from Ministerio de Ciencia e Innovación under the Ramón y Cajal program (RYC2020-029656-I). RBO has received funding from the Atapuerca Foundation. The field work at the Atapuerca sites is financed by the Junta de Castilla y León and the Atapuerca Foundation.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Galindo-Pellicena, M., Sala, N., De Gaspar, I. et al. Long-term dog consumption during the Holocene at the Sierra de Atapuerca (Spain): case study of the El Portalón de Cueva Mayor site. Archaeol Anthropol Sci 14, 84 (2022). https://doi.org/10.1007/s12520-022-01522-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-022-01522-5