
Abstract
Herbivore teeth are a valuable source of information for inferring the hunting season of past hunter-gatherers, the spatial–temporal organization of their activities, their socio-economic organization, and their adaptation to the seasonal fluctuation of the resources. Numberless of studies have been conducted on Neanderthals across Eurasia, but only few of them rely on the application of cementochronology and tooth wear analyses combined to obtain information about the ungulate paleodiet, paleoenvironments, and the time range of the mortality events. In this study, we present the results achieved though the combination of these two high-resolution techniques applied to large and medium-sized herbivore teeth yielded by two Middle Paleolithic sites in the north-east of Italy. We combined the two methodologies with the aim to overcome any possible lack of information, due to the use of a single method. This study addressed to analyze the material coming from two caves in the Venetian region, De Nadale and San Bernardino, and to produce data supporting the interpretation of the origin of the two archaeofaunal assemblages as a result of seasonal hunting events that took place mainly in winter. In this specific geographic and environmental context, our data gain a better understanding of Neanderthal subsistence strategies and occupational patterns.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The spatial–temporal organization of the activities within the territory of Neanderthals is a key proxy to discuss their socio-economic organization and the responses they developed to cope with the seasonal fluctuation of the resources (Conard and Prindiville 2000; Rendu 2010; Delagnes and Rendu 2011; Rosell et al. 2012; Chacón et al. 2015, among others). Depending on the region and the time considered, different mobility patterns were developed by these past human populations for the exploitation of its biotope. In this context, the Mousterian record in the Italian Alpine range and foreland could disclose insights into the adaptation of Neanderthals, their scheduling capacity, and the flexibility of their economy: by exploiting at the same time mountainous, low altitude and plain environments, these natives were induced to adapt their ecological relations through designing their seasonal mobility patterns. In this scenario, zooarchaeological data play a fundamental role in inferring the seasonality of prey procurement by hominids.
Nowadays both seasonality and duration of occupation can be estimated through some valuable analytical methods applied to teeth, such as the analysis of carbon and oxygen stable isotopes (e.g., Feranec et al. 2009; Balasse et al. 2012; Julien et al. 2012; Blumenthal et al. 2014), the study of tooth eruption and replacement patterns (e.g.Wilson et al. 1982; Mariezkurrena 1983; Carter 1998; Bunn and Pickering 2010), the dental micro- and mesowear analyses (e.g., Rivals and Deniaux 2005; Rivals et al. 2009a, b; Rivals and Semprebon 2012; Rivals and Semprebon 2012; Sánchez-Hernández et al. 2016; Uzunidis 2020), and the cementochronology technique (e.g., Klevezal 1988; Pike-Tay 1991a,b; Lieberman 1994; Burke and Castanet 1995; Rendu 2007, 2010; Naji et al. 2015). Nevertheless, using one of these methods alone may not be always reliable, since when used independently, a certain technique may provide too low-resolution data.
In this paper, we present a twofold high-resolution approach, combining cementum increment and dental wear analyses on ungulate teeth (Cervus elaphus, Megaloceros giganteus, Bos/Bison, and Capreolus capreolus) from two Middle Paleolithic multi-layered sites, San Bernardino Cave (Grotta Maggiore di San Bernardino) and De Nadale Cave, located in the Berici Hills in north-eastern part of Italy. Our objectives are to provide a better understanding of the seasonality and the length of occupations of these sites in order to contribute to the reconstruction of mobility patterns and exploitation of the territory by Neanderthals during the late Middle Paleolithic between the Alpine fringe and foreland.
Dental wear analysis, both meso- and microwear, is easily affected by dietary changes, ruled in nature by the annual changing of seasons, but on different time scales (Grine 1986; Fortelius and Solounias 2000). Mesowear analysis records a macroscopic wear, such as the shape and the highness of the cusps of the molars, revealing the average annual diet of the last months or a whole year (Fortelius and Solounias 2000; Ackermans et al. 2018). By contrast, microwear analysis investigates microscopic features which are the recording of the diet of the last days or weeks before the death of the animal (Merceron et al. 2004; Semprebon et al. 2004; Rivals and Semprebon 2012). So, the combination of these two dental wear techniques gives access not only to the degree of attrition and abrasiveness of food in an annual cycle (mesowear), but also to any possible variations due to seasonal environmental influences (microwear) (Sánchez-Hernández et al. 2019). Moreover, microwear analysis is also useful to assess the duration of accumulation events and, consequently, the extent of the human occupation. This approach helps to differentiate assemblages which were accumulated in a seasonal or shorter event, events longer than a season, and separated events occurring in different seasons (Rivals et al. 2015a, b, 2018).
In addition to that, the cementochronology technique, which is based on the observation of the microscopic incremental tissue of the teeth (see “Materials and methods”) enables us to estimate the age at death of each individual and the season when the animal was killed (Lieberman 1991; Pike-Tay 1991a; Gourichon 2004; Rendu 2007; Naji et al. 2015, among the others). This allows us to correlate the seasonal feeding pattern with a specific period of the annual cycle (Sánchez-Hernández et al. 2019, 2020).
The sites
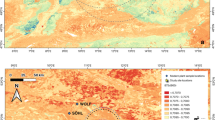
The region where De Nadale and San Bernardino caves are located is dominated by three geomorphological units: the Po and the Adige alluvial plains in the south, the pre-Alps in the north, and two small sub-alpine massifs (the Berici and the Euganean Hills) in the south-east (Fig. 1). The present-day physical landscape of the Berici Hills is an ensemble of markedly different morphological zones. Above both De Nadale and San Bernardino caves, at an average elevation of 250 m, the karst plateau forms a gentle honeycomb with sinkholes and various depressions (including ponors and limestone pavements) succeeding one another, delineating an extremely uneven topography with peaks and block karst affected by surface dissolution. In De Nadale surroundings, the plateau is dissected by the Calto valley bottom, a depressed system with pocket-valleys, a swampy environment, and steep slopes all around. To the east, the Pozzolo ancient karst surface is a wide trench cutting through the plateau in a NW–SE direction at an elevation of 150 m, ending at both the SE and NW (Sauro 2002). The San Bernardino Cave opens on the Eastern slope of the Berici Hills at dominant position onto the Bacchiglione river alluvial plain, facing the southern side of the Euganean Mounts. The Berici Hills area produced an important amount of paleolithic evidence, both as open-air sites and as caves and shelters (Leonardi and Broglio 1962; Bertola and Peresani 2000; Peresani 2001a, 2015) used by Neanderthal foragers as part of a settlement system extended to the Euganean Hills, the Alpine foreland, and the southern slope of the Alps (e.g., Peresani et al. 2011, Peresani 2013).

A The geographical location of De Nadale and San Bernardino Caves in the north-east of Italy; B the stratigraphic sequence of De Nadale Cave (dotted lines show the roof and the bedrock of the cave; Unit 7 is colored in green); C the entrance of De Nadale Cave; D the entrance of San Bernardino Cave; E the stratigraphic sequence of San Bernardino Cave (Unit II is colored in yellow and Unit IV in pink)
De Nadale Cave
De Nadale cave is a small cavity at 80 m a.s.l. above the narrow Calto valley. It was first reported in 2006 and extensively excavated since 2014. The field campaigns carried out from 2014 to 2017 and still ongoing exposed a short stratigraphic sequence composed of eight different stratigraphic units (SU), including one single anthropic layer (SU 7) embedded between Pleistocene sterile sediments (SU 6 and SU 8) (Jéquier et al. 2015). SU 7 extends on almost entirely the cavity. It yielded a cultural assemblage attributed to the Quina Mousterian (Jéquier et al. 2015; Livraghi et al. 2021) dated to 70.2 + 1/ − 0.9 ka BP by U/Th (Jéquier et al. 2105). The anthropic layer yielded thousands of fragmented bones, charcoals, and a Neanderthal deciduous tooth (Arnaud et al. 2016). The anthracological assemblage from Unit 7 of De Nadale cave is characterized by the strong presence of spruce—larch woodland and cryophilous pine forests—and indicates the important role that montane and alpine flora played in this region during MIS 4 (Vidal-Matutano et al. 2022). The frequentation at De Nadale is framed in a landscape dominated by open woodland formations and dry meadows at the very beginning of MIS 4 (López-García et al. 2018), a period still quite unknown in the north of Italy.
Large and medium-sized ungulates—red deer (Cervus elaphus), giant deer (Megaloceros giganteus), and large bovids (Bison priscus and Bos primigenius)—dominate the faunal spectrum, both according to NISP and MNI. Smaller ungulates, such as roe deer (Capreolus capreolus), chamois (Rupicapra rupicapra), wild boar (Sus scrofa), and ibex (Capra ibex), have also been recovered although in lower quantity. Carnivore remains yielded by Unit 7 are scarce, and they have been identified mainly as belonging to cave bear (Ursus spelaeus) and other non-identifiable bear species, with lower percentages of wolf (Canis lupus), fox (Vulpes vulpes), and badger (Meles meles) (Livraghi et al. 2021).
San Bernardino Cave
The San Bernardino Cave opens at 135 m a.s.l. Several systematic archaeological excavations were carried out in the second half of the last century and focused on the atrial area: the first began in 1959, while the second stage went on from 1986 to 1994. The excavations unearthed a 4.5-m-thick stratigraphic sequence composed by eight layers spanning from the Late Middle Pleistocene to the Late Pleistocene and containing Mousterian lithic assemblages (Leonardi and Broglio 1961; Peresani 1995; Fiore et al. 2004).
Sediment composition and stratigraphy of units from VIII to I identify three main cycles (Cassoli and Tagliacozzo 1994; Peresani 2001a, b; López-García et al. 2017), dated on the basis of U/Th and electron spin resonance (ESR) and radiocarbon from MIS 7 to MIS 3 (Gruppioni 2004; Picin et al. 2014; Terlato et al. 2021): the 1st cycle is referable to the Late Middle Pleistocene and characterized by a phase of wet and temperate climatic conditions (unit VIII) with broadleaf, wooded landscapes, followed by a phase of slightly colder oscillations (unit VII). The 2nd cycle is referable to the Late Pleistocene and characterized by temperate climatic conditions, generally forested environment with some open spaces and wetlands (unit VI). This period was followed by a colder climatic phase (units V–IV) which resulted in the formation of a steppe environment. The 3rd cycle is referable to the Late Pleistocene and characterized by a humid phase (unit III) correlated to a more wooded landscape. In particular, unit II shows an increasing in anthropogenic remains.
So far, zooarchaeological studies have been carried out in detail only on unit II. The faunal record is dominated by ungulates, among which the roe deer (Capreolus capreolus) is the most common species, followed by the red deer (Cervus elaphus) in a lower quantity. The wild boar (Sus scrofa), the elk (Alces alces), the chamois (Rupicapra rupicapra), and large bovines are present but scarce. The evidence of giant deer (Megaloceros giganteus), ibex (Capra ibex), and rhinoceros (Stephanorhinus sp.) (Romandini et al. 2018; Terlato et al. 2019; 2021) is rare. This trend seems to be common also in other units, where preliminary studies pointed out the predominance of red and roe deer over chamois and ibex, with a stable presence of few bovines, giant deer, wild boar, and elk (Cassoli and Tagliacozzo 1994; Fiore et al. 2004).
Materials and methods
The sample analyzed in this study is composed of 74 teeth, sorted among the whole faunal material yielded by the two deposits during the excavations. The material was studied at taxonomical and taphonomical levels in previous works (Cassoli and Tagliacozzo 1991; Livraghi et al. 2021; Terlato et al. 2019, 2021) and stored at the Department of Humanities of the University of Ferrara, Italy.
Among the 74 specimens, 23 out of 25 gave interpretable results when studied with the cementochronology technique and 36 out of 59 with the dental wear analyses. Despite the general good macroscopic appearance, two teeth out of the 25 specimens selected for the cementochronology were discarded. They were affected by a microbiological attack which caused extensive demineralization of the hydroxyapatite, followed by collagen lysis and, subsequently, the complete loss of the structure of the cementum itself (Geusa et al. 1999). Among the 59 teeth selected for dental wear analyses, 23 teeth were discarded since they did not present the optimal features for the study. To allow a good evaluation, we selected only the molars and the fourth premolars, and we discarded young and senile individuals—to avoid any bias due to their unworn or, on the contrary, heavily blunt surface. Moreover, some of these 23 discarded teeth had a badly preserved enamel that did not allow any observation. Both techniques have been performed on ten specimens, but only four gave positive results with both.
The San Bernardino Cave sample is composed of 47 specimens (Table 1) coming from units II (NR: 38), IV (NR: 8), and VI (NR: 1), mostly belonging to roe deer, the most common taxon, followed by red deer, elk, giant deer, and bovids. Dental wear analysis has been carried out on 39 teeth and cementochronology on 12; among them, four remains were suitable for a directly combined study obtained applying the two techniques.
The De Nadale Cave sample is composed of 27 (Table 1) teeth coming from Unit 7 (NR: 19) and 13 (NR: 8) identified as belonging to giant deer, red deer, and roe deer. No bovid teeth were found. We carried out dental wear analysis on 20 specimens, while cementochronology was applied on 13 remains; among them, six teeth were suitable for a directly combined study obtained applying both techniques. We were not able to sample four large-sized ungulate teeth since they have been chosen for new U/Th dating, and they are currently under study.
Cementochronology
As pointed out by several biological studies, in the teeth of most temperate, sub-arctic, and arctic mammal species, the cementum surrounding the roots grows regularly, according to a predictable seasonal rhythm and starting from the complete eruption of the tooth until the death of the animal (Klevezal and Kleinenberg 1969; Gordon 1984; Klevezal 1988; Pike-Tay 1991a, b; Lieberman and Meadow 1992). This incremental tissue appears, under transmitted cross-polarized light, as a stratified deposit: somewhat regular bands are organized in pairs which correlate with a year timespan. Every couple of layers is namely the result of the annual deposition of cementum and consists of a thicker and translucent band (TB, accretion line, or growth layer), which is formed during periods of more substantial and fast tissue growth (i.e., from spring to autumn) and a thinner and opaque band (OB, the “line of arrested growth”—LAG—or annulus), which deposits during periods of reduced tissue growth, such as winter (Pike-Tay 1991a, b, 1995; Lieberman 1994). Therefore, the age at death of the animal was deduced adding the number of pairs of layers to the time of tooth eruption, while the season of death was pointed out by the nature of the band observed.
This regularity in cementum growth has been observed both in cellular and acellular cementum, but only the latter seems to be reliable and suitable for this kind of approach since its deposition is more regular and rarely biased by mechanical and/or biological stress (Pike-Tay 1991a, b; Lieberman and Meadow 1992; Lieberman 1993a, 1994; Stutz 2002a; Gourichon 2004; Rendu 2007).
Among the 74 teeth we sampled, 25 (i.e., 12 specimens from San Bernardino Cave and 13 from De Nadale Cave) were analyzed with the cementochronology approach, following the well-established protocol for the archaeological application of this method (Saxon and Higham 1968; Spiess 1976; Gordon 1988; Pike-Tay 1991a, b; Lieberman and Meadow 1992; Lieberman 1993a, b, 1994; Burke and Castanet 1995; Rendu and Armand 2009; among others).
Following the literature (Lieberman and Meadow 1992; Gourichon 2004; Rendu 2007; Naji et al. 2015), we sampled as many teeth as possible, coming from different individuals, still encased in the alveolar bone or, in case of loose teeth, not showing any post-mortem damage on the roots, any presence of manganese stains, weathering cracks, or concretions. In some cases, and it will be specified, we chose more than one tooth per individual, to validate the results and to avoid any uncertainty in the observations.
We analyzed the upper half of the root, in correspondence of the cervix of the tooth, where the cementum is clearly readable. The sample was processed following the techniques for ground thin sections applied in archaeology contexts (Rendu 2007; Naji et al. 2015; Gourichon and Parmegiani 2016):
-
Extraction (if necessary) of the tooth from the alveolar bone.
-
Cleaning of the specimen with ethyl alcohol.
-
Embedding of the roots of the tooth in translucent epoxy resin. The specimen is located vertically in a plastic mold, filled with epoxy resin, and put into a vacuum pump for 12–24 h, in order to remove bubbles and let the resin completely permeate the tooth tissue. There, the polymerization process begins.
-
Cutting of the block of resin containing the tooth, using a slow-speed diamond saw. Sections are made both transversally and longitudinally, after the removal, when possible, of the tooth crown to preserve it for further analyses. To avoid optical superimposition of the cementum layers within the section, the transverse cuts are made orthogonally, and the longitudinal cuts are made as much parallel as possible to the major axis.
-
Gluing the obtained slice (0.5–1 mm) on glass-slide with epoxy glue. The slice is heated and pressed for 8 h so that it can adhere perfectly to the glass.
-
Abrading of the upper face of the slice until it reaches the thickness of 30–50 μm. The obtained thin section is also polished with the diamond powder to erase scratches that can hinder the observation under the microscope.
We examined the thin sections with a transmitted light polarizing microscope (Leica DM2500P) at × 10, × 20, × 40, and × 50 magnifications, in order to recognize the best regions of interest for the observation. We counted the incremental bands, and we identified the last deposits through optical images by using three distinct light filters: plane-polarized light, cross-polarized light, and full-wave retardation plate (λ plate). The use of the λ plate allowed us to identify the eventual presence of microbiological alterations (fungi or bacteria), diagenetic alterations of the cementum (recrystallization, false increments, collagen leaching), or the effect of weathering (Stutz 2002b; Rendu et al. 2009). The whole process was carried out at PACEA laboratories of the University of Bordeaux.
The results of the observations and the pictures taken with a high-resolution camera connected to the microscope were digitally reworked and enhanced with the support of the software Image J, following the standard protocol (Lieberman et al. 1990). This allowed us to achieve a more accurate estimation when the last cementum accretion was a growth layer, avoiding potential subjective mistakes due to the competence and the experience of the observer (Lubinski and O'Brien 2001). By comparing the mean thickness of the proceeding and fully formed bands, the “good season” to which the growth layer corresponds can be divided into three sub-periods: the beginning (up to one-third of the mean thickness, 1–33.3%), the middle (from one- to two-thirds, 33.4–66.6%), and the end (more than two-thirds, 66.7–100%) (Gourichon 2004; Rendu 2007; Sánchez-Hernández et al. 2020).
Tooth mesowear
We carried out the mesowear technique through evaluating the relief and the degree of sharpness of the molars’ cusps, by observing the buccal side of the upper molars and lingual side of the lower molars, with the naked eye. The sharpness and the morphology of the cuspids point out different degrees of attritive or abrasive dental wear, which are the result of different kinds of diets, registered within the animal’s lifespan (Ackermans et al. 2020). In general, by applying this method, herbivores can be grouped in three main feeding categories:
-
Browsers: feeding on leaves from shrubs and trees. The molars present high relief and sharp apices, due to the low degree of abrasion and the high degree of attrition.
-
Grazers: feeding on grass. The molars present low relief and blunt apices, due to the high degree of abrasion and the low degree of attrition.
-
Mixed feeders: the molars present intermediate values of abrasion and attrition, with a variable morphology of the tooth outline, according with the feeding preference of an individual.
Tooth mesowear analysis was applied to 19 specimens, 9 of which from De Nadale Cave and 10 from San Bernardino Cave.
Following Fortelius and Solounias (2000) and Mihlbachler et al. (2011), each tooth was scored with a 0 to 6 value, where stage 0 corresponds with a high and sharp cusps type and stage 6 to a completely blunt with no relief profile molar. The averaged value of the mesowear measurements taken on teeth from an assemblage corresponds to the mesowear score (MWS). To avoid biased results, this technique was applied to non-fractured teeth, in which the crown and the occlusal surface do not show any damage or taphonomical alterations (Fortelius and Solounias 2000; Kaiser and Fortelius 2003). Teeth that do not present significant wear as well as those that are heavily worn, depending on the age of the individual, were not suitable for the evaluation and were discarded (Rivals and Semprebon 2006; Rivals et al. 2007).
Tooth microwear
The microwear technique describes and analyzes a pattern of microscopic features readable on the occlusal surface enamel, which provides information about the diet of an individual at the time of its death (Grine 1986; Solounias and Semprebon 2002; Semprebon et al. 2004). These marks are indeed left by the abrasive particles present in food, which may leave scratches and pits during the masticatory process, with a rapid overprint of these marks within each food intake (Grine 1986).
We prepared and described the sample to be analyzed following the well-established protocol (Solounias and Semprebon 2002; Semprebon et al. 2004; Rivals and Semprebon 2011, 2012; Rivals et al. 2007, 2009a, b, c a, b, Rivals et al. 2015a, b, Rivals et al. 2018; Sánchez-Hernández et al. 2019, 2020, among the others). The occlusal surface of each specimen was cleaned with acetone and then 96% ethanol, and once dry, it is molded with vinylpolysiloxane, a high-resolution dental silicone. The molds obtained were filled with transparent epoxy resin, in order to create highly detailed casts. Every cast was carefully screened under the transmitted light of the stereomicroscope (a Zeiss Stemi 2000C) at × 35 magnification with an ocular reticle delimitating a 0.16 mm2 square area. The microscopic features of the enamel were easily observed and quantified, thanks to the refractive properties of the clear epoxy cast. These microfeatures were classified into three categories: pits (circular or sub-circular scars), scratches (elongated scars with a straight direction), and gouges (larger and deeper pits with irregular outline). We quantified the micro-features on the enamel of the paracone of the upper molars and the protoconid of the lower molars. We sampled two different areas on each specimen, in order to average the microwear features per tooth. The results were compared with a database containing information on extant and wild ungulate taxa (Solounias and Semprebon 2002). The number of scratches (from now on, Nscr) and the number of pits (Npit) are strongly linked to the dietary habit of the ungulates. In modern populations, browsers show a wear pattern with a low Nscr, while grazers display a high Nscr. As predictable, mixed feeders are characterized by the overlap of the two other patterns, since they switch seasonally (and/or regionally), between diets based either on browse or on grass. To better discriminate mixed feeders from browsers or grazers, we applied the well-established method developed by Semprebon and Rivals (2007) which gave significative results when applied both to extant and to fossil samples (e.g., Rivals et al. 2018; Sánchez-Hernández et al. 2019). Thus, we calculated the percentage of individuals in a population with scratch numbers that fall between 0 and 17 scratches in the 0.16 mm2 area (i.e., the LSR, low scratch range): the browsers have LSR values that fall between 0 and 22.2%, browsers show values comprised in the range of 72.73–100%, and the mixed feeders overlap partially with the other two categories, being comprised between 20.93 and 70% (Solounias and Semprebon 2002; Semprebon and Rivals 2007; Rivals and Semprebon 2010). Qualitative features were evaluated too: the scratch width score (SWS) defines the thickness of the scratches using a scoring system from 0 to 4 (from “fine” to “mixed coarse/hyper-coarse”). This scale varies according to the abrasive properties of food consumed by the individual. Moreover, the frequency of cross scratches (%XS) refers to the presence of scratches with different directions relative to the main orientation.
Beside this, microwear analysis gives information about the relative duration of the occupation through the estimation of the duration of the mortality event(s) of the ungulates (Rivals et al. 2009b, 2015b). Following Rivals et al. (2015b), we calculated the coefficient of variation (CV) and the standard deviation (SD) of a species’ scratch variability, and we plotted the values into a heat map which was divided in three areas, corresponding to different durations of event(s): (A) a season-long (or shorter) period, (B) a timespan longer than a season; and (C) at least two separated events that occurred in different non-contiguous seasons (Rivals et al. 2015b). Taxa with a minimum of four individuals suitable for the analysis were selected to get a picture of each population’s variability. As some of the samples used here are too small to detect the true CV and SD values of the larger population that they represent, we applied a joint bootstrapped function of CV and SD (n = 500, with replacement) using the R code by Domínguez-Rodrigo et al. (2019).
Results
Cementochronology
De Nadale Cave
When analyzed with the cementochronology method, 12 (92.3%) of the 13 teeth we selected gave positive results (Table 2), presenting at least one suitable region of interest to perform the observation. Fungi or bacterial alteration and recrystallization are present, affecting especially the dentine, but they do not bias the analysis of the cementum. The sole exception consists of a giant deer’s molar (CN661), which does not show any presence of cementum, probably due to the heavy microbiological alteration that affected the dentine and lead to the collapse of the structure of the layers.
The cementum bands are clearly readable and measurable, and the dental tissue is generally well preserved. This made possible the digital enhancement to be performed on each specimen, except for a red deer incisor (CN929): in this case, it was not possible to recognize the end of the last band (LCB), making the measurement not reliable.
Among the three taxa sampled, the results are homogeneous for Cervus elaphus and Capreolus capreolus, but they show a certain degree of intraspecific variability for Megaloceros giganteus. Namely, the only sampled tooth of roe deer presents an opaque band (OB) as LCB; the same was observed for all the specimen of red deer, except for two teeth, presenting a very thin translucid band (TB) in one case and a complete TB on the other case.
Giant deer shows some significative discrepancies from the pattern of the two taxa mentioned: in two cases, the LCB is an opaque band (bad season), two other cases are halfway through the translucid band development, and one tooth shows a nearly complete translucid band, which corresponds to the end of the good season.
San Bernardino Cave
Cementochronology gave positive results from 11 (91.6%) out of 12 teeth (Table 3). The structure of the cementum is visible and reliable in most of the sample, even though the dentine often presents microbiological and diagenetic alterations. Only one specimen, a roe deer lower third molar, gave no readable data, due to the recrystallization of the cementum itself.
For unit VI, only one tooth was suitable for the analysis, a C. capreolus lower third molar, which clearly shows an opaque band as LCB.
For unit IV, the sample consist of only one C. elaphus tooth, with clear cementum stratification, including the LCB which was recognized as a very thin opaque band, corresponding to the beginning of the bad season.
The largest sample came from unit II, and it is composed of one lower molar of M. giganteus, one premolar from a Bovidae, four teeth from C. capreolus, and four from C. elaphus. Both the giant deer’s and the bovine’s specimens show a very thin opaque band as LCB, which points out a bad season mortality. The data for C. capreolus are quite homogeneous, too: three teeth show a last opaque band (bad season), while one specimen ends with a halfway grown translucid band (middle of the good season).
The red deer, instead, shows a slightly higher intraspecific variability: two teeth show a very thin translucid band (very beginning of the good season), one ends with an almost fully grown TB, and one ends with half of a full TB (Fig. 2).

A Schematic representation of hunting events for Megaloceros giganteus, Cervus elaphus, and Capreolus capreolus at De Nadale Cave. B Schematic representation of hunting events for Megaloceros giganteus, Cervus elaphus, Capreolus capreolus, and bovids at San Bernardino Cave. (black, sample from Unit II; yellow, sample from Unit IV; red, sample from Unit VI). Percentages correspond to the degree of development of the last translucent cementum band (TB)
Meso- and microwear analyses
De Nadale Cave
Unit 7 yielded very few specimens suitable for the mesowear analysis: among the totality of the teeth sampled from the site, only 9 of them (Table 4) gave positive results, since the cusps were not always well preserved, fractured, or biased by post-depositional events.
The three taxa analyzed show a quite homogeneous trend, with mesowear scores (MWS) ranging from 0.75 for C. capreolus (N = 4) to 2 for M. giganteus (N = 2). C. elaphus (N = 3) has a MWS of 1, falling between the two above-mentioned species. Accordingly, data suggest that the roe deer and the red deer have a browser diet, while giant deer tends toward a browse-dominated mixed feeder diet. Nevertheless, the number of specimens that allowed us to apply the methodology is too modest to give a meaningful interpretation of the data.
Microwear analysis was applied to a broader sample: 16 teeth, out the 20 specimens sampled from De Nadale Cave, gave positive results when observed under the microscope.
The average numbers of pits (Npit) and scratches (Nscr) are very close for all the three species considered, and, when plotted, they fall within the limits of the confidence ellipse for modern browsers (Fig. 3A). This result is consistent with the microwear score (LSR) that indicates pure browser values since no individual presents a Nscr higher than 17. From a qualitative point of view, the three taxa have high rates of individuals showing large pits (LP) and significative percentages of gouges, which are features of a browse dietary preference with the possibility of fruit and seed consumption. No giant deer or roe deer individuals show cross scratches (XS), which are present only on a low percentage of red deer specimens. The scratch width score (SWS) observed shows a predominance of fine and mixed fine-coarse scratches, related to the high level of attrition typical of the browse diet and consistent with all the above-mentioned data.

Above: bivariate plot of the average number of pits and scratches in the selected taxa from De Nadale Cave: Megaloceros giganteus (Mg); Cervus elaphus (Ce), Capreolus capreolus (Cc). Error bars correspond to standard deviation (± 1 SD) for the fossil samples. Plain ellipses correspond to the Gaussian confidence ellipses (p = 0.95) on the centroid for the extant leaf browsers and grazers from Solounias and Semprebon (2002). Below: boundary lines with the error probability (heat map) based on SD and CV values of microwear data used for the classification of samples into short events (region A), long-term events (region B), or two separated short events (region C)
When plotted into the heat map (Fig. 3B), data available give significative results for all the taxa sampled: all the three populations have low standard deviations (SD) and low coefficient of variation (CV) of the numbers of scratches (Table 4). They plot in area [A] of the heatmap, indicating a short duration of the accumulation event(s).
San Bernardino Cave
Only a few specimens (Table 4) from Unit II of San Bernardino Cave were suitable for the mesowear analyses, mainly because of the not optimal state of preservation that characterizes the archaeological material. The most common damage that affected the sample was the presence of several broken tips that could bias the evaluation of the MWS and that were discarded. Nevertheless, the trend is homogeneous for the three species taken into account, with a MWS ranging from 0.40 of C. capreolus (N = 5), to 1.25 of C. elaphus (N = 4), to 2 of A. alces (N = 1). Once again, data show that the roe deer and the red deer have a pure browser diet, while elk tends toward a browse-dominated mixed feeder diet; however, for the latter, the sample size is too small to get a definite interpretation.
The number of specimens sampled for microwear analysis, however, is more consistent, with a total of 20 teeth out of 39 that gave positive results. Most of them were yielded by Unit II (N = 16), while only 4 teeth came from Unit IV, which was less rich in archaeological evidence than Unit II.
The material sampled from Unit IV, which belongs to red deer (N = 2), roe deer (N = 1), and bovids (N = 1), is characterized by a low number of scratches (Nscr from 6.5 to 9.5 per counting unit) and a low number of pits (Npit from 4.5 to 9.75 per counting unit). Large pits are present on the bovid and on the C. elaphus specimens but completely absent on the C. capreolus molar. In the three taxa, gouges and cross scratches are absent, except for one of the red deer’s molars examined which presents some gouges. The SWS indicates a mix of fine and coarse scratches in the sample. The bivariate distribution of the numbers of scratches and pits, as well as the LSR, classifies the three taxa among the pure browsers (Fig. 4A).

Above: bivariate plot of the average number of pits and scratches in the selected taxa from San Bernardino Cave: Alces alces (Aa); Cervus elaphus (Ce), Capreolus capreolus (Cc), and bovids (BB). Roman numbers next to the abbreviations of the taxa indicate the unit of provenance (II, Unit II; IV, Unit IV). Error bars correspond to standard deviation (± 1 SD) for the fossil samples. Plain ellipses correspond to the Gaussian confidence ellipses (p = 0.95) on the centroid for the extant leaf browsers and grazers from Solounias and Semprebon (2002). Below: boundary lines with the error probability (heat map) based on SD and CV values of microwear data used for the classification of samples into short events (region A), long-term events (region B), or two separated short events (region C)
The evidence yielded by Unit II is more robust: 5 teeth from C. capreolus, 4 from C. elaphus, and one from A. alces show a Nscr ranging from 8 to 8.86 and a Npit from 4.5 to 6.5. Large pits are frequent on elk and red deer teeth. The range of specimens showing gouges is significant for, again, elk and red deer. Large pits and gouges are present on roe deer molars as well, but the values are significantly lower. Cross scratches were observed only on few red deer’s specimens. The SWS points out the presence of a mixture of fine and coarse scratches on the surface of the teeth, related to the high level of attrition of the browse diet. The bivariate plot shows that the three species fall within the limits of the confidence ellipse for modern browsers (Fig. 4A).
When plotted into the heat map (Fig. 4B), data available for Unit IV gave positive results only for Cervus elaphus, despite the smallness of the sample. The microwear values (Table 4) show a low intraspecific variability and lie within zone [A]. The same scenario emerged for Unit II: all the three species taken into account—Alces alces, Cervus elaphus, and Capreolus capreolus—fall within the boundary of zone [A], attesting a low intraspecific variability.
Discussion
According to cementochronology and meso- and microwear analyses on the teeth of the most abundant taxa at De Nadale and San Bernardino Caves, we estimated the extent and the seasonality of mortality event(s) for each population. Consequently, knowing that the two deposits have an anthropogenic origin (Cassoli and Tagliacozzo 1994; Peresani 2001a, b; Jéquier et al. 2015; Livraghi et al. 2021; Terlato et al. 2021), we inferred the duration and the seasonality of the occupation of the Neanderthal groups that exploited them.
The three taxa dominating Unit 7 of De Nadale Cave—C. elaphus, M. giganteus, and C. capreolus—show a homogeneous dietary pattern characterized by the high consumption of attritive resources (mainly leaves and shrubs). This low variability, which is highlighted both by the MWS and by the LRS, places them within the limits of the confidence ellipse of the extant browsers (Fig. 3A) and seems to indicate that the three cervid species fed on similar vegetation. Due to competition for the same ecological niche, they probably partitioned the resources occupying different habitats with similar vegetation.
The presence of red deer and roe deer relates to a landscape with a predominance of woodlands over open spaces under relatively cold-temperate climatic conditions and in proximity of a humid area. This view is not in conflict with the massive presence of giant deer (Chritz et al. 2009), and it is consistent with data inferred from the study on micromammals (López-García et al. 2018).
The low standard deviation and coefficient of variation, falling into zone [A] of the heatmap, indicate that the accumulation event(s) lasted for a limited period in a timespan of a year, less than or equal to a season (Fig. 3B).
Moreover, the cementochronology data confirm the presence of more than a single occupation: thin sections indicate that most of the game animals were killed during the bad season, namely winter (at least four red deer, two giant deer, and one roe deer), but other minor hunting events occurred in the middle and at the end of the good season (Fig. 2A). No pattern of seasonal prey selection can be recognized, since the three taxa were exploited homogeneously during each short occupational event (i.e., red deer, giant deer, and roe deer were hunted during winter and the good season as well).
Combining the results coming from the two methodologies, thus, De Nadale Cave takes the form of a short-term occupational site, with multiple accumulation events that took place mostly during winter and some minor events during the good season. Results are consistent with the presence of 38 bone fragments of large-sized ungulates which were recognized as infants (0–5 months) or early juvenile, not older than 1 year, that were probably part of the herds exploited by the human groups (Livraghi et al. 2021).
A similar situation can be depicted for the teeth sampled from Unit II at San Bernardino Cave: the three cervid taxa—A. alces, C. elaphus, and C. capreolus—have the same patterns of dietary traits, specific of the pure browsers (Table 4, Fig. 4A). This result fits well with the palaeoecological reconstruction that underlines the presence of generally temperate conditions in a forested landscape, interspersed by wetlands and humid areas (Cassoli and Tagliacozzo 1994; Peresani 2001a, b; López-García et al. 2017; Terlato et al. 2021). Also in this case, the low SD and CV point out one or more short mortality events, plotting in zone [A] of the heatmap (Fig. 4B).
Cementochronology results indicate the presence of a major mortality event during the bad season and a very small one, represented by an individual of red deer and an individual of roe deer, in the middle of the good season. Two other teeth slightly deviate from the standard results, but they can be placed at the very end of the bad season (Fig. 2B). There is no evidence of a preferential or seasonal exploitation of a single species rather than the others.
For Unit IV, data are too exiguous to be reliable: only one incisor of a red deer was suitable to be cut for the cementochronology, while tooth wear analyses were applied to four teeth, one recognized as Bovidae, one as roe deer, and two as red deer. The small sample is aligned with the results from Unit II, plotting into the limits of the extant browsers, but only the two molars from red deer were suitable for the evaluation of the extent of the occupation, falling into zone [A].
Interpreted as a whole, all the data point out a high mobility pattern of the human groups that inhabited the region. This scenario fits well with other studies based on dental wear analyses on Middle Paleolithic materials from Western European sites. They suggest similar conclusions related to high and diversified mobility of the human groups in relation to seasons and environmental context. For example, similar evidence is reported in France at Payre, level F (MIS 8–7) interpreted as a short-term occupation associated with other activities (Rivals et al. 2009c; Moncel and Rivals 2011), and at the specialized reindeer hunting camp of Les Pradelles (MIS 4–3) (Costamagno et al. 2006), in Belgium at Scladina Cave (Moncel et al. 1998; Patou-Mathis and Bocherens 1998), and in Germany at Salzgitter Lebenstedt (MIS 5–3), where the accumulation of reindeer remains corresponds to seasonal (or shorter) events (Gaudzinski and Roebroeks 2000).
Unlike the data given by De Nadale and San Bernardino Caves, some studies underline a different scenario: at Portel-Ouest (France), the samples, corresponding to four taxa from the same level, fall in two distinct areas of the heatmap. The accumulation of reindeer, red deer, and large bovids is linked to seasonal or shorter events, while the horse corresponds to a longer event. This prey procurement pattern can be easily related to known human hunting strategies for different prey at different moments of the year (Rivals et al. 2015b).
Moreover, another different occupational pattern for Neanderthal groups was recognized at Taubach (Germany), where the remains of bison show a long-term event of accumulation (Rivals et al. 2015b), and at Covalejos Cave (Spain), which is therefore characterized as either long-term occupations or as a succession of short-term occupations throughout the year (Sánchez-Hernández et al. 2019).
Conclusion
Despite the difficulty to assess the nature of Neanderthal occupations and to identify the type of settlement (i.e., butchery halt, unspecialized short or long occupation camp, etc.), both De Nadale Cave and San Bernardino Cave shed new light on the organization of the territory.
Both sites attest to a high mobility pattern of the human groups that occupied the caves for short periods of time, during a timespan shorter or equal to a season. From the study, the tendency to settle in the two sites during winter emerged, with some brief occupations during the good season.
From the methodological point of view, the integrated application of cementochronology and tooth wear analyses allowed us to confirm the significance of combining these two high-resolution methods. The usefulness of this combination is in the possibility to obtain more detailed data and to avoid any lack of information due to the application of a single methodology. Specifically, the combination of both approaches suggests that the lack of variability in the microwear signal does not necessarily indicate a long-term or a succession of short-term prey procurement events.
References
Ackermans NL, Martin LF, Codron D, Hummel J, Kircher PR, Richter H, Kaiser TM, Clauss M, Hatt J (2020) Mesowear represents a lifetime signal in sheep (Ovis aries) within a long-term feeding experiment. Palaeogeogr Palaeoclimatol Palaeoecol 553:109793. https://doi.org/10.1016/j.palaeo.2020.109793
Ackermans NL, Winkler DE, Schulz-Kornas E, Kaiser TM, Müller DWH, Kircher PR, Hummel J, Clauss M, Hatt J-M (2018) Controlled feeding experiments with diets of different abrasiveness reveal slow development of mesowear signal in goats (Capra aegagrus hircus). J Exp Biol 221(21):jeb186411. https://doi.org/10.1242/jeb.186411
Arnaud J, Benazzi S, Romandini M, Livraghi A, Panetta D, Salvadori PA, Volpe L, Peresani M (2016) A Neanderthal deciduous human molar with incipient carious infection from the Middle Palaeolithic De Nadale cave. Italy Am J Phys Anthropol 162(2):370–376
Balasse M, Obein G, Ughetto-Monfrin J, Mainland I (2012) Investigating seasonality and season of birth in past herds: a reference set of sheep enamel stable oxygen isotope ratios. Archaeom 54(2):349–368. https://doi.org/10.1111/j.1475-4754.2011.00624.x
Bertola S, Peresani M (2000) Variabilità tecnologica in due insiemi litici di superficie dei Colli Berici. Quad Archeol Veneto XVI 1:92–96
Blumenthal SA, Cerling TE, Chritz KL, Bromage TG, Kozdon R, Valley JW (2014) Stable isotope time-series in mammalian teeth: in situ δ 18O from the innermost enamel layer. Geochim Cosmochim Acta 124:223–236
Bunn HT, Pickering TR (2010) Methodological recommendations for ungulate mortality analyses in paleoanthropology. Quat Res 74(3):388–394. https://doi.org/10.1016/j.yqres.2010.07.013
Burke A, Castanet J (1995) Histological observations of cementum growth in horse teeth and their application to archaeology. J Archaeol Sci 22:479–493
Chacón MG, Bargalló A, Gabucio MJ, Rivals F, Vaquero M (2015) Neanderthal behaviors from a spatio-temporal perspective: an interdisciplinary approach to interpret archaeological assemblages. In: Conard NJ, Delagnes A (eds) Settlement Dynamics of the Middle Paleolithic and Middle Stone Age, vol IV. Tübingen Publications in Prehistory. Kerns Verlag, Tübingen, pp 253–294
Carter RJ (1998) Reassessment of seasonality at the Early Mesolithic site of Star Carr, Yorkshire based on radiographs of mandibular tooth development in red deer (Cervus elaphus). J Archaeol Sci 25(7):851–856
Cassoli PF, Tagliacozzo A (1994) I resti ossei di macromammiferi, uccelli e pesci della Grotta Maggiore di San Bernardino sui Colli Berici (VI): considerazioni paleoeconomiche, paleoecologiche e cronologiche. Bul Paletnol Ital 85:1–71
Chritz KL, Dyke GJ, Zazzo A, Lister AM, Monaghan NT, Sigwart JD (2009) Palaeobiology of an extinct Ice Age mammal: stable isotope and cementum analysis of giant deer teeth. Palaeogeogr Palaeoclimatol Palaeoecol 282:133–144
Conard NJ, Prindiville TJ (2000) Middle Palaeolithic hunting economies in the Rhineland. Int J Osteoarchaeol 10:286–309
Costamagno S, Meignen L, Beauval C, Vandermeersch B, Maureille B (2006) Les Pradelles (Marillac-le-Franc, France): a Mousterian reindeer hunting camp? J Archaeol Anthropol 25:466–484
Delagnes A, Rendu W (2011) Shifts in Neandertal mobility, technology, and subsistence strategies in western France. J Archaeol Sci 38:1771–1783
Domínguez-Rodrigo M, Sánchez-Flores AJ, Baquedano E, Arriaza MC, Aramendi J, Cobo-Sánchez L, Organista E, Barba R (2019) Constraining time and ecology on the Zinj paleolandscape: Microwear and mesowear analyses of the archaeofaunal remains of FLK Zinj and DS (Bed I), compared to FLK North (Bed I) and BK (Bed II) at Olduvai Gorge (Tanzania). Quat Int 526:4–14
Feranec RS, Hadly EA, Paytan A (2009) Stable isotopes reveal seasonal competition for resources between late Pleistocene bison (Bison) and horse (Equus) from Rancho La Brea, southern California. Palaeogeogr Palaeoclimatol Palaeoecol 271:153–160
Fiore I, Gala M, Tagliacozzo A (2004) Ecology and subsistence strategies in the Eastern Italian Alps during the Middle Palaeolithic. Int J Osteoarchaeol 14:273–286
Fortelius M, Solounias N (2000) Functional characterization of ungulate molars using the abrasion-attrition wear gradient: a new method for reconstructing paleodiets. Am Mus Novit 3301:1–36
Gaudzinski S, Roebroeks W (2000) Adults only. Reindeer hunting at the Middle Palaeolithic site Salzgitter Lebenstedt. Northern Germany J Hum Evol 38:497–521
Geusa G, Bondioli L, Capucci E, Cipriano A, Grupe G, Savoré C, Macchiarelli R (1999) Dental Cementum Annulations and Age at Death Estimates. In: L. Bondioli and R. Macchiarelli (eds) Osteodental Biology of the People of Portus Romae (Necropolis of Isola Sacra, 2nd–3rd Cent. AD), 2, Roma: Museo Nazionale "L. Pigorini".
Gordon BC (1984) Selected bibliography of dental annular studies on various mammals. Zooarchaeol Res News Supplement 2:1–24
Gordon BC (1988) Of men and reindeer herds in French Magdalenian prehistory. BAR International Series, Oxford, p 390
Gourichon L (2004) Faune et saisonnalité: L’organisation temporelle des activités de subsistance dans l’Épipaléolithique et le Néolithique précéramique du Levant nord (Syrie). Unpublished thesis (PhD). Université Lumière-Lyon 2.
Gourichon L, Parmegiani V (2016) Preliminary analysis of dental cementum of Lama guanicoe for the estimation of age and season at death: studies of modern specimens and further archaeological applications. J Archaeol Rep 6:856–861
Grine FE (1986) Dental evidence for dietary differences in Australopithecus and Paranthropus: a quantitative analysis of permanent molar microwear. J Hum Evol 15:783–822
Gruppioni G (2004) Datation par les méthodes uranium-thorium (U/Th) et résonance paramagnétique électronique (RPE) de deux gisements du Paléolithique moyen et supérieur de Vénétie: la grotte de Fumane (Monts Lessini- Vérone) et la grotte majeure de San Bernardino (monts Berici-Vicence). Unpublished thesis (PhD). University of Ferrara.
Jéquier C, Peresani M, Romandini M, Delpiano D, Joannes-Boyau R, Lembo G, Livraghi A, López-García JM, Obradovíc M, Nicosia C (2015) The de Nadale cave, a single layered Quina Mousterian site in the North Italy. Quartär 62:7–21
Julien M-A, Bocherens H, Burke A, Drucker DG, Patou-Mathis M, Krotova O, Péan S (2012) Were European steppe bison migratory? 18O, 13C and Sr intra-tooth isotopic variations applied to a paleoethological reconstruction. Quat Int 271:106–119
Kaiser TM, Fortelius M (2003) Differential mesowear in occluding upper and lower molars opening mesowear analysis for lower molars and premolars in hypsodont horses. J Morphol 258:67–83
Klevezal GA (1988) Recording structures of mammal: determination of age and reconstruction of life history. (Russian title: Registriruyushchie structury mlekopitayushchikh v zoologicheskikh issledovaniyakh.). A. A. Balkema, Rotterdam
Klevezal GA, Kleinenberg SE (1969) Age determination of mammals from annual layers in teeth and bones. Acad Sci USSR, Israel Program for Scientific Translations Series, Jerusalem
Leonardi P, Broglio A (1961) Paleolitico superiore in situ nel deposito pleistocenico della Grotta di S. Bernardino nei colli Berici orientali (Vicenza). Atti Istituto Veneto Scienze, Lettere ed Arti CXlX:435–450
Lieberman DE (1993a) The rise and fall of seasonal mobility among hunter-gatherers: the case of the Southern Levant. Curr Anthropol 34:569–598
Lieberman DE (1993b) Life history variables preserved in dental cementum microstructure. Sci 261(5125):1162–1164. https://doi.org/10.1126/science.8356448
Lieberman DE (1994) The biological basis for seasonal increments in dental cementum and their application to archaeological research. J Archaeol Sci 21(4):525–539
Lieberman DE, Meadow RH (1992) The biology of cementum increments (with an archaeological application). Mam Rev 22:57–77
Lieberman DE, Deacon TW, Meadow RH (1990) Computer image enhancement and analysis of cementum increments as applied to teeth of Gazella gazella. J Archaeol Sci 17:519–533
Livraghi A, Fanfarillo G, Dal Colle M, Romandini M, Peresani M (2021) Neanderthal ecology and the exploitation of cervids and bovids at the onset of MIS4: a study on De Nadale cave, Italy. Quat Int 586:24–41. https://doi.org/10.1016/j.quaint.2019.11.024
López-García JM, Luzi E, Peresani M (2017) Middle to Late Pleistocene environmental and climatic reconstruction of the human occurrence at Grotta Maggiore di San Bernardino (Vicenza, Italy) through the small-mammal assemblage. Quat Sci Rev 168:42–54
López-García JM, Livraghi A, Romandini M, Peresani M (2018) The de Nadale cave (Zovencedo, Berici hills, North-eastern Italy): a small-mammal fauna from near the onset of marine isotope stage 4 and its paleoclimatic implications. Paleogeogr Palaeoclimatol Palaeoecol 506:196–201
Lubinski PM, O’Brien CJ (2001) Observations on seasonality and mortality from a recent catastrophic death Assemblage. J Archaeol Sci 28:833–842
Mariezkurrena K (1983) Contribución al conocimiento del desarrollo de la dentición y el esqueleto postcraneal de Cervus elaphus. Munibe 35:149–202
Merceron G, Blondel C, Brunet M, Sen S, Solounias N, Viriot L, Heintz E (2004) The Late Miocene paleoenvironment of Afghanistan as inferred from dental micro-wear in artiodactyls. Palaeogeogr Palaeoclimatol Palaeoecol 207:143–163
Mihlbachler MC, Rivals F, Solounias N, Semprebon GM (2011) Dietary change and evolution of horses in North America. Sci 331:1178–1181
Moncel M-H, Rivals F (2011) On the question of short-term Neanderthal site occupations: Payre, France (MIS 8–7), and Taubach/Weimar, Germany (MIS 5). J Anthropol Res 67(1):47–75
Moncel M-H, Patou-Mathis M, Otte M (1998) Halte de chasse au chamois au Paléolithique moyen: la couche 5 de la grotte Scladina (Sclayn, Namur, Belgique). In: Brugal J-P, Meignen L, Patou-Mathis M (eds) Économie préhistorique: les comportements de subsistance au Paléolithique, APDCA, Antibes, pp 291–309
Naji S, Gourichon L, Rendu W (2015) La cémentochronologie. In: Balasse M, Brugal J-P, Dauphin Y, Geigl E-M, Oberlin C, Reiche I (eds) Messages d’os. Archéométrie du squelette animal et humain, éditions des Archives contemporaines, Paris, pp 217–240
Patou-Mathis M, Bocherens H (1998) Comportements alimentaires des hommes et des animaux a` Scladina. In: Otte M, Patou-Mathis M, Bonjean D (eds) Recherches aux grottes de Sclayn, L'Archéologie, vol. 2, ERAUL, Liège, pp 329–336
Peresani M (1995) Sistemi tecnici di produzione litica nel Musteriano d’Italia. Studio tecnologico degli insiemi litici delle unità VI e II della Grotta di San Bernardino (Colli Berici, Veneto). Riv Sci Preist XLVII:79–167
Peresani M (2001a) An overview of the middle palaeolithic settlement system in North-Eastern Italy. In: Conard NJ (ed) Settlement dynamics of the Middle Palaeolithic and Middle Stone Age, Publications in Prehistory, Introductory Volume. Kerns Verlag, Tubingen, pp 485–506
Peresani M (2001) The Middle Palaeolithic settlement system of the Eastern Italian Alps in ecological context. Atti del XIII Congresso degli Antropologi Italiani. Riv Antropol 78(Suppl):19–23
Peresani M (2012) Fifty thousand years of flint knapping and tool shaping across the Mousterian and Uluzzian sequence of Fumane cave. Quat Int 247:125–150
Peresani M (2013) Contesti, risorse e variabilità della presenza umana nel Paleolitico e nel Mesolitico nei Colli Euganei. Preist Alp 47:109–122
Peresani M (2015) I Neandertaliani e il Musteriano nei Colli Berici. Insediamenti e sfruttamento delle materie prime litiche. Archeol Veneta XXXVIII 1:10–27
Peresani M, Chrzavzez J, Danti A, De March M, Duches R, Gurioli F, Muratori S, Romandini M, Trombino L, Tagliacozzo A (2011) Fire-places, frequentations and environmental setting of the final Mousterian at Grotta di Fumane: a report from the 2006–2008 research. Quartär 58:131–151
Picin A, Peresani M, Falguéres C, Gruppioni G, Bahain JJ (2014) San Bernardino Cave (Italy) and the appearance of Levallois technology in Europe: results of a radiometric and technological reassessment. PLoS ONE 8(10):e76182
Pike-Tay A (1991a) L’analyse du cément dentaire chez les cerfs: l’application en préhistoire. Paleo 3(1):149–169
Pike-Tay A (1991b) Red deer hunting and the upper Palaeolithic of Southwestern France: a study in seasonality. BAR International Series, 569, Oxford
Rendu W (2007) Planification des activités de subsistance au sein du territoire des derniers Moustériens Cémentochronologie et approche archéozoologique de gisements du Paléolithique moyen (Pech-de-l’Azé I, La Quina, Mauran) et Paléolithique supérieur ancien (Isturitz). Ph.D., Université Bordeaux 1
Rendu W (2010) Hunting behavior and Neanderthal adaptability in the Late Pleistocene site of Pech-de-l’Azé I. J Archaeol Sci 37:1798–1810
Rendu W, Armand D (2009) Saisonnalité de prédation du Bison du gisement moustérien de La Quina (Gardes-le-Pontaroux, Charente), niveau 6c. Apport à la compréhension des comportements de subsistance. Bul Soc Préhist Fr 106(4):679–690
Rendu W, Armand D, Pubert É, Soressi M (2009) Approche taphonomique en cémentochronologie: réexamen du niveau 4 du Pech-de-l’Azé I (Carsac, Dordogne, France). Paléo 21:223–236
Rivals F, Deniaux B (2005) Investigation of human hunting seasonality through dental microwear analysis of two Caprinae in late Pleistocene localities in Southern France. J Archaeol Sci 32:1603–1612
Rivals F, Semprebon G (2006) A comparison of the dietary habits of a large sample of the Pleistocene pronghorn Stockoceros onusrosagris from the Papago Springs Cave in Arizona to the modern Antilocapra americana. J Vertebr Paleontol 26:495–500
Rivals F, Semprebon GM (2010) What can incisor microwear reveal about the diet of ungulates? Mamm 74:401–406
Rivals F, Semprebon GM (2011) Dietary plasticity in ungulates: insight from tooth microwear analysis. Quat Int 245:279–284
Rivals F, Semprebon GM (2012) Paleoindian subsistence strategies and late Pleistocene paleoenvironments in the northeastern and southwestern United States: a tooth wear analysis. J Archaeol Sci 39:1608–1617
Rivals F, Mihlbachler MC, Solounias N (2007) Effect of ontogenetic-age distribution in fossil samples on the interpretation of ungulate paleodiets using the mesowear method. J Vertebr Paleontol 27:763–767
Rivals F, Schulz E, Kaiser TM (2009a) Late and middle Pleistocene ungulates dietary diversity in Western Europe indicate variations of Neanderthal paleoenvironments through time and space. Quat Sci Rev 28:3388–3400
Rivals F, Schulz E, Kaiser TM (2009b) A new application of dental wear analyses: estimation of duration of hominid occupations in archaeological localities. J Hum Evol 56:329–339
Rivals F, Moncel M-H, Patou-Mathis M (2009c) Seasonality and intra-site variation of Neanderthal occupations in the Middle Palaeolithic locality of Payre (Ardèche, France) using dental wear analyses. J Archaeol Sci 36:1070–1078
Rivals F, Julien M-A, Kuitems M, Van Kolfschoten T, Serangeli J, Drucker DG, Bocherens H, Conard NJ (2015a) Investigation of equid paleodiet from Schoeningen 13 II-4 through dental wear and isotopic analyses: archaeological implications. J Hum Evol 89:129–137
Rivals F, Prignano L, Semprebon GM, Lozano S (2015b) A tool for determining duration of mortality events in archaeological assemblages using extant ungulate microwear. Sci Rep 5:17330
Rivals F, Kitagawa K, Julien MA, Patou-Mathis M, Bessudnov AA, Bessudnov AN (2018) Straight from the horse’s mouth: high-resolution proxies for the study of horse diet and its relation to the seasonal occupation patterns at Divnogor’ye 9 (Middle Don, Central Russia). Quat Int 474B:146–155
Romandini M, Thun Hohenstein U, Fiore I, Tagliacozzo A, Perez A, Lubrano V, Terlato G, Peresani M (2018) Late Neandertals and the exploitation of small mammals in northern Italy: fortuity, necessity or hunting variability? Quaternaire 29:61–67
Rosell J, Cáceres I, Blasco R, Bennàsar M, Bravo P, Campeny G, Esteban-Nadal M, Fernández-Laso MC, Gabucio MJ, Huguet R, Ibáñez N, Martín P, Rival F, Rodríguez-Hidalgo A, Saladié P (2012) A zooarchaeological contribution to establish occupational patterns at Level J of Abric Romaní (Barcelona, Spain). Quat Int 247:69–84
Sánchez-Hernández C, Rivals F, Blasco R, Rosell J (2016) Tale of two timescales: combining tooth wear methods with different temporal resolutions to detect seasonality of Palaeolithic hominin occupational patterns. J Archaeol Sci Rep 6:790–797
Sánchez-Hernández C, Gourichon L, Pubert E, Rendu W, Montes R, Rivals F (2019) Combined dental wear and cementum analyses in ungulates reveal the seasonality of Neanderthal occupations in Covalejos Cave (Northern Iberia). Nat Sci Rep 9:14335. https://doi.org/10.1038/s41598-019-50719-7
Sánchez-Hernández C, Gourichon L, Soler J, Soler N, Blasco R, Rosell J, Rivals F (2020) Dietary traits of ungulates in northeastern Iberian Peninsula: did these Neanderthal preys show adaptive behavior to local habitats during the Middle Palaeolithic? Quat Int 557:47–62
Sauro U (2002) The Monti Berici: a peculiar type of karst in the southern Alps. Acta Carsologica 31(3–6):99–114
Saxon A, Higham C (1968) A new research method for economic prehistorians. Am Antiq 34(3):303–311
Semprebon GM, Rivals F (2007) Was grass more prevalent in the pronghorn past? An assessment of the dietary adaptations of Miocene to Recent Antilocapridae (Mammalia: Artiodactyla). Palaeogeogr Palaeoclimatol Palaeoecol 253:332–347
Semprebon GM, Godfrey LR, Solounias N, Sutherland MR, Jungers WL (2004) Can low-magnification stereomicroscopy reveal diet? J Hum Evol 47:115–144
Spiess A (1976) Determining season of death of archaeological fauna by analysis of teeth. Arctic 29:23–55
Stutz AJ (2002a) Pursuing past seasons: a re-evaluation of cementum increment analysis in paleolithic archaeology. Unpublished thesis (PhD). University of Michigan.
Stutz AJ (2002b) Polarizing microscopy identification of chemical diagenesis in archaeological cementum. J Archaeol Sci 29:1327–1347
Terlato G, Livraghi A, Romandini M, Peresani M (2019) Large bovids on the Neanderthal menu: exploitation of Bison priscus and Bos primigenius in northeastern Italy. J Archaeol Sci Rep 25:129–143
Terlato G, Lubrano V, Romandini M, Marin-Arroyo AB, Benazzi S, Peresani M (2021) Late Neanderthal subsistence at San Bernardino cave (Berici hills – Northeastern Italy) inferred from zooarchaeological data. Alp Mediterr Quat. https://doi.org/10.26382/AMQ.2021.10
Uzunidis A (2020) Large ungulates mobility and Neanderthal subsistence behaviours: a preliminary tooth microwear analysis. J Archaeol Sci Rep 29:102084. https://doi.org/10.1016/j.jasrep.2019.102084
Vidal-Matutano P, Livraghi A, Peresani M (2022) New charcoal evidence at the onset of MIS 4: first insights into fuel management and the local landscape at De Nadale cave (northeastern Italy). Rev Palaeobot Palynol 298:104594. https://doi.org/10.1016/j.revpalbo.2021.104594
Wilson B, Grigson C, Payne S (1982) Ageing and sexing animal bones from archaeological sites. Archaeopress, Oxford
Acknowledgements
Research at the De Nadale and at San Bernardino Caves is coordinated by the University of Ferrara (M.P.) in the framework of a project supported by the Ministry of Culture-Veneto Archaeological Superintendence (SABAP – Verona, Vicenza e Rovigo) and the municipalities of Zovencedo and Mossano. The project has been financed by the Hugo Obermaier Society, local private companies (R.A.A.S.M. and Saf), and local promoters. The authors thank all the students and anyone who took part in the excavation, Carlos Sánchez-Hernández for helping with the digital enhancement of cementum images, Davide Delpiano for providing inputs to the paper, Diana Marcazzan and Mirka Govoni for the digital elaboration and map creation, and anonymous reviewers for contributing to ameliorate the manuscript. Special thanks go to Eric Pubert and the PACEA laboratory and the cementochronology department at the University of Bordeaux for helping during the cementum analysis.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature.
Author information
Authors and Affiliations
Contributions
All authors have contributed to this manuscript and have approved the final version.
Alessandra Livraghi: Conceptualization, investigation, formal analysis, validation, writing (original draft), and writing (review and editing). Florent Rivals: Conceptualization, investigation, methodology, validation, formal analysis, writing (review and editing), and supervision. William Rendu: Investigation, validation, and writing (review and editing). Marco Peresani: Supervision, conceptualization, funding acquisition, resources, project administration, and writing (review and editing).
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Livraghi, A., Rivals, F., Rendu, W. et al. Neanderthals’ hunting seasonality inferred from combined cementochronology, mesowear, and microwear analysis: case studies from the Alpine foreland in Italy. Archaeol Anthropol Sci 14, 51 (2022). https://doi.org/10.1007/s12520-022-01514-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-022-01514-5