
Abstract
Tana del Barletta is an upland cave used from the Late Neolithic to the Middle Bronze Age, located in the vicinity of the coast in Liguria (NW Italy). The excavation revealed the presence of a faunal assemblage dominated by caprine and cattle remains. In order to gain new data on late prehistoric farming strategies (e.g. seasonal mobility, coastal grazing, animal diet), intra-tooth series of stable oxygen and carbon isotopes have been obtained from cattle and sheep/goat tooth enamel, along with intra-tooth series of nitrogen and carbon isotopes from cattle dentine collagen. Due to the prevalence of maxillary teeth, a modern calf has also been analysed to assess intra-individual isotopic differences between the maxillary and mandibular dentition. Modern data on oxygen isotope values of meteoric water from different altitudes around the area of the site were used as a reference for interpretation. The results indicate that the water ingested by the herd was mostly characterised by particularly low δ18O values, highlighting the importance of the uplands for the late prehistoric farmers of the region. However, the input of water sourced from lower elevations, especially during the winter months, cannot be dismissed. In addition, the nitrogen isotopic composition of cattle collagen rules out the ingestion of salt-tolerant vegetation or seaweed, suggesting that grazing did not occur directly on the coastal plain.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Numerous works of environmental archaeology have focused on the rugged territory overlooking the sea on the northwestern Italian coast (Liguria). These contributions include geoarchaeological (Macphail et al. 1997), archaeobotanical (Arobba et al. 2018) and zooarchaeological (Rowley-Conwy 2000; Rowley-Conwy et al. 2020) investigations, as well as studies on archaeometallurgy (Maggi and Pearce 2005), ceramic petrography (Capelli et al. 2008), osteology (Marchi et al. 2006; Sparacello and Marchi 2008; Sparacello et al. 2016, 2017; Dori et al. 2020; Orellana-González et al. 2020; Varalli et al. 2020) and stable isotope analyses of human remains (Pettitt et al. 2003; Goude et al. 2011, 2020; Varalli et al. 2015).
In Liguria, there is abundant evidence supporting the importance of animal husbandry since the beginning of the Neolithic, with a range of zooarchaeological, geoarchaeological and archaeobotanical data suggesting that during the Late Neolithic mobile pastoralism became a paramount subsistence strategy for the communities settled in the region. Most of the archaeological sites consist of caves and rock shelters located in the area of Finale Ligure and in the Pennavaira Valley (Fig. 1). The prevalence of domestic species in the faunal assemblages, the occurrence of deciduous teeth and the presence of dung layers and thick spherulite-rich fumier deposits (Macphail et al. 1997; Rowley-Conwy 1997; Barker et al. 1990; Rowley-Conwy et al. 2020) suggest that these sites were largely used as stables since the Early Neolithic (c. 5800–5200 BC) (Rowley-Conwy et al. 2020) and especially during the Middle Neolithic (Square-Mouthed Pottery: c. 5000–4500 BC), when some of them (i.e. Cave of Arene Candide) were occupied all the year round and animals were fed with fodder collected in the surroundings (Maggi and Nisbet 2000). At the Cave of Arene Candide, during the Impressa phase of the Early Neolithic (c. 5800–5600 BC), animal farming seems to be mostly aimed at milk production (Rowley-Conwy et al. 2020), while with the onset of the Middle Neolithic, the strategy switched towards meat (Rowley-Conwy 1997). The data from the other Ligurian sites are insufficient to draw any firm conclusions but, on a larger geographical scale, the situation in the Liguro-Provençal milieu and Languedoc is rather varied (Rowley-Conwy et al. 2013).

Location of Ligurian cave sites that provided evidence for animal husbandry between the Neolithic and the Early Bronze Age. 1: Grotta del Pertuso; 2: Tana della Volpe; 3: Arma della Grà di Marmo; 4: Grotta Cornarea; 5: Grotta del Pertusello; 6: Arma dello Stefanin; 7: Arma di Nasino; 8: Grotta dell’Olivo; 9: Caverna delle Arene Candide; 10: Arma dell’Aquila; 11: Arma delle Anime; 12: Caverna del Frate; 13: Bric Tana. The grey star marks the position of Tana del Barletta, and the black circle marks the city of Genoa
The subsequent onset of the Chassey (c. 4300–3600 BC) and Copper Age (c. 3500–2000) cultures saw a decrease in tree taxa, together with evidence for controlled use of fire (Nisbet 1997) and changes in stratigraphic and sedimentological units (Cruise et al. 2009; Macphail et al. 1997), suggesting that herds were taken to graze in newly created pastures (Maggi and Nisbet 1990; Maggi and Campana 2008; Branch and Marini 2014; Branch et al. 2014; Morandi and Branch 2018). The relevance of pastoral activities for the communities settled in the Liguro-Provençal arc is also shown by the Late Neolithic-Early Bronze Age anthropogenic indicators (Huet 2017a: 21–27) associated with the well-known rock engravings of Mt. Bégo, in the Maritime Alps (De Lumley 1995; Barfield and Chippindale 1997; Huet 2017b).
In this regard, while on the coast there are examples of open-air sites with multiple burials, at higher elevations only multiple burials in small natural caves are known (Maggi and Pearce, in press, with bibliography). Three cases in particular are worth mentioning: the Grotta del Pertuso in the upper Argentina valley (1330 m a.s.l.) (Fig. 1, no. 1), which hosted the remains of a minimum of 18 individuals of all ages and both sexes from the Copper Age to the Early Bronze Age (Del Lucchese and Ricci 1987; Formicola and Balestri 1987; Maggi and Nisbet 1990; Varalli et al. 2015), Tana della Volpe (750 m a.s.l.) (Fig. 1, no. 2), hosting no less than 68 individuals distributed over a thousand years throughout the Copper Age (Del Lucchese and De Pascale 2011) and Arma della Grà di Marmo (1000 m a.s.l.) (Fig. 1, no. 3), with no less than 30 individuals (Ricci 1998). According to Maggi and Nisbet (1990), this and other evidence, including cases of endogamy (Bareschino et al. 2013; Maggi and Pearce, in press), may indicate that whole family groups managed the pastoral activities over a long period of time, rather than individual herders guiding their livestock.
The system of presumed short-distance transhumance towards summer highland pastures, suggested by the occurrence of stable-caves at different altitudes and small burial caves in the uplands, besides archaeological and environmental evidence, continues until the Early Bronze Age (2200–1800 BC).
Later on, an increase in open-air sites occurs, the location of which suggests a more structured arrangement of routes and resources (Maggi and Nisbet 1990; Maggi 1999, 2004). However, even though understanding the nature of animal husbandry, including seasonal movements, is of paramount importance to better interpret subsistence strategies at the time, isotopic investigations of faunal remains—and oxygen isotope data in particular—are very scant so far (Le Bras-Goude et al. 2006; D’Angela and Longinelli 1993).
In order to address this and assess the potential of stable isotope analyses for contributing to key research questions for the later prehistory of the region, we conducted a pilot study, analysing cattle and caprine tooth enamel and dentine collagen from Tana del Barletta, a small high-altitude cave in the Pennavaira Valley (western Liguria) which was used mainly from the Neolithic to the Bronze Age. The context appears particularly suitable to assess herd mobility in the territory, not only because of the relatively close proximity of mountain pastures and sea shore, which can be expected to produce markedly different oxygen isotope values, but also because of the availability of coastal grazing grounds, which should be traceable by higher δ15N values (Britton et al. 2008).
The Tana del Barletta tooth assemblage consists almost entirely of maxillary teeth. However, nearly all isotopic studies of sequentially formed tissues make use of mandibular molars, whether investigating archaeological material (e.g. Towers et al. 2014; Buchan et al. 2016) or analysing modern samples for reference datasets (e.g. Balasse et al. 2012a). Therefore, due to the scarcity of knowledge regarding the variation of the oxygen isotopic composition in maxillary teeth and their relationship with mandibular teeth (Balasse et al. 2021), we used a modern cattle specimen to assess the reliability of the upper dentition to record intra-tooth variations.
Isotopic background (δ18O, δ13C, δ15N) and current interpretative models for animal mobility
Along with the potential for determining birth seasonality, the application of stable isotopes to detect seasonal mobility in archaeological herds has attracted the attention of the scholars since the early 2000s (Pederzani and Britton 2019) and remains a very frequent approach (Gerling et al. 2017; Chazin et al. 2019; Tomczyk et al. 2019; Bishop et al. 2020; Trentacoste et al. 2020; Ventresca Miller et al. 2020), due to the prominence of the debate revolving around ancient forms of mobile pastoralism (Arnold and Greenfield 2006; Jourdain-Annequin and Duclos 2006).
Isotopic fractionation involves a change in the ratio between light and heavy isotopes during temperature-controlled phase transitions. Condensation preferentially removes heavier 18O from atmospheric water, resulting in a 18O depletion of cloud masses. At high- and mid-latitudes, this creates seasonal temperature fluctuations, which are responsible for cyclic variations in the oxygen isotopic composition of meteoric water (temperature effect) (Dansgaard 1964), and the farther atmospheric water travels inland, away from source water bodies (e.g. oceans), the more it becomes depleted in 18O, with a ~ 1.8‰ depletion every 1000 km (continental effect) (Rozanski et al. 1993). Furthermore, as clouds rise along an altitudinal gradient, the oxygen isotopic composition of meteoric water in the uplands becomes 18O-depleted relative to warmer lowlands (altitude effect) (Rozanski et al. 1993; Pederzani and Britton 2019). Due to this mechanism, a ~ 0.15–0.3‰ decrease in δ18O values occurs every 100-m increase in altitude in the Italian peninsula, where the rugged geomorphology particularly accentuates the altitude effect (Longinelli and Selmo 2003; Giustini et al. 2016).
The δ18O value of mammal body water is largely determined by the δ18O of ingested water, and reflected within the δ18O of phosphate and structural carbonate components of skeletal tissues (biogenic apatite and collagen). When the δ18O of precipitation fluctuates throughout the year, incrementally formed tissues, such as tooth enamel, record these seasonal variations, albeit with some attenuation (Fricke and O’Neil 1996).
So far, examinations of archaeological assemblages making use of stable oxygen isotopes have attempted several approaches to infer or rule out herd mobility. These are the amplitude of intra-tooth δ18O values (Mashkour et al. 2005), the relationship between the pattern of variations in the δ18O and δ13C curves (Makarewicz 2017a; Makarewicz et al. 2017; Tejedor-Rodríguez et al. 2021), variations in δ13C intra-tooth series (Balasse and Ambrose 2005) and the combination of δ18O and 87Sr/86Sr ratios (Balasse et al. 2002; Pellegrini et al. 2008; Valenzuela-Lamas et al. 2016). However, especially where stable oxygen isotopes were considered on their own, the multitude of different interpretative strategies used by researchers has highlighted the need for a more rigorous methodological framework to detect past seasonal mobility (see Makarewicz 2017a; Hermes et al. 2018). Analyses of modern animals with known life histories from different environments play thus an important role in the intra- and inter-individual variations that are to be expected (e.g. Britton et al. 2009; Makarewicz and Pederzani 2017; Tornero et al. 2018).
Based on the current knowledge of precipitation/enamel δ18O relationships at mid-latitudes, a general trend valid for cases of vertical mobility seems to be best described by Britton et al. (2009) (see also Henton 2012 and Hermes et al. 2018). A model of wider applicability can be hence extrapolated, where movement to 18O-depleted elevations in summer and to 18O-enriched lowlands in winter leads to a flattening of the pronounced sinusoidal pattern typical of sedentary animals (e.g. Fricke and O’Neil 1996; Fricke et al. 1998), although local variables can play a role in masking this trend (see e.g. Knockaert et al. 2018). Nevertheless, in domesticates, such a pattern may also be produced by certain herding practices, e.g. the supply of 18O-enriched leaf water through winter foddering or 18O-depleted groundwater in summer (Makarewicz and Pederzani 2017; Hermes et al. 2018).
The relative abundance of 13C and 12C (usually expressed as δ13C) in vegetation varies according to the photosynthetic pathway of the plants and is further modified by the environment in which plants grow (Lambers et al. 2008). In Mediterranean settings, the great majority of taxa follows the Calvin-Benson cycle (C3 plants) (Deines 1980), which tends to result in lower δ13C values (~ − 34 to − 24‰) (Smith and Epstein 1971; Tauber 1981), although these also moderately fluctuate seasonally and along the altitudinal gradient (Körner et al. 1991; Smedley et al. 1991). A group of native plants following the Hatch-Slack cycle (C4 plants) also occurs in temperate Europe (Pyankov et al. 2010), although they are generally too rare to register a significant impact on consumers’ δ13C values. In the case of Tana del Barletta, it is also relevant to highlight that seaweeds—which obtain their carbon primarily from dissolved CO2 in the oceans—and seaside plants can also exhibit high δ13C values (~ − 18 to − 10‰), falling within a range similar to that of terrestrial C4 plants (Tauber 1981). It is likely that in antiquity the coast of western Liguria was home to a sand-dune habitat favourable to the growth of salt-tolerant C4 Mediterranean halophytes, such as Salsola spp. and Euphorbia peplis (Arobba et al. 2001; Acosta and Ercole 2015). To enable comparison of modern plant δ13C with archaeological datasets, a 1.5‰ difference has also to be taken into account, due to the fossil fuel effect (Francey et al. 1999).
When considered jointly, oxygen and carbon stable isotopes can also provide indications of mobility, as in modern specimen’s lowland/highland seasonal migration results in inverted δ18O and δ13C seasonal curves (Tornero et al. 2018). However, the distinction between vertical movement and winter foddering can be difficult to discern (Makarewicz 2017a; Makarewicz et al. 2017).
A number of studies have established a correlation between coastal flora and higher δ15N values (Virginia and Delwiche 1982; Heaton 1987; Page 1995). This is evident in seaweeds, halophytes and even in non-halophytic coastal vegetation, due to nitrogen enrichment of the soil in proximity of the sea. These elevated δ15N values are passed on through the food chain and can be observed in the bone and dentine collagen of the consumers (Britton et al. 2008). Due to the absence of remodelling of primary dentine, this latter allows observation of nitrogen levels during discrete phases of tooth formation. However, instrumental and natural averaging of the signal still occurs to some degree, due to a c. 1.5–2-month equilibration period of the metabolites within the body reservoir (Balasse et al. 2001), as well as to the sampling of growth increments oblique to the axis of the tooth, which causes some mixing of dentine layers deposited over different periods (Balasse et al. 2001; Zazzo et al. 2006). Studies of archaeological bone and dentine collagen from Britain and north-central Europe have found significantly higher δ15N, and sometimes also δ13C values in cattle and caprine specimens from salt-marsh sites, and these isotopic thresholds have been taken as indicative of coastal grazing (Britton et al. 2008; Müldner et al. 2014).
It is important to note that the carbon and nitrogen isotopic composition of both enamel bioapatite and dentine collagen in cattle also reflects the pre-weaning intake of maternal milk and the post-weaning dietary change along first and second molars. As prior to weaning mother and calf occupy two different trophic levels, a c. 3–3.5‰ offset in collagen δ15N values occurs, the mother’s signal being lower (Balasse et al. 2001; Gillis et al. 2013), whereas M1 enamel δ13C values reflect a progressive enrichment in 13C, due to rumen development (Towers et al. 2014). In addition, also the oxygen isotopic composition of enamel in cattle and sheep first molars is influenced by the pre-weaning signal, reflecting the composition of mother’s body water, which has already undergone fractionation (Fricke and O’Neil 1996).
Archaeological context
Tana del Barletta is a small cave located at c. 900 m a.s.l. on the steep calcareous slope of Mt. of Gettine, c. 14 km from the coast, in the northern side of the Pennavaira Valley (western Liguria) (N 44°07′57.7″, E 8°03′51.7″) (Figs. 1 and 2). Archaeological investigations during the mid-1980s identified a superimposition of three layers (c. 20 cm apart) allowing radiocarbon dating of the occupations between the Late Neolithic and the Middle Bronze Age, possibly followed by episodic visits in the Iron Age (Del Lucchese et al. 1987; Maggi and Nisbet 1990; Barker et al. 1990). Few artefacts were uncovered by the excavation, including stone tools and potsherds, along with animal bones and very few human remains (Del Lucchese et al. 1987; Barker et al. 1990). Out of a total of 1713 identifiable zooarchaeological remains, caprines prevail in the assemblage (822 fragments, from 54 to 74% according to the phase of occupation), followed by cattle (314 fragments, from 21 to 32%) and pigs (127 fragments, from 7 to 20%), while deer and hare only occur in very low amounts (Barker et al. 1990).

Entrance of the cave of Tana del Barletta on the slope of Mt. of Gettine (top left) and view of the late prehistoric horizons brought to light by the 1984 archaeological fieldwork (bottom left). Stratigraphic section of the cave deposit (top right) showing the site phases and the radiocarbon-dated hearths (the fireplace at the base of level 2, not intercepted by the section, is projected on the sequence with a dashed line). Plan of the cave showing the location of the hearths within the area where bovid teeth were found (bottom right)
The cave of Tana del Barletta is of special interest for its location near the ridge, while most of the contemporary sites in the Pennavaira Valley typically occur in the valley floor (Leale Anfossi 1962; Del Lucchese et al. 1987; Maggi and Nisbet 1990). Animal husbandry is documented in the cave since the Late Neolithic, suggesting the exploitation of the high pastures of Mt. Galero (1708 m a.s.l.), just c. 4.5 km from the site.
It has been proposed that the Pennavaira Valley was probably involved in a system of family-run short-range transhumance, connecting hypothetical main settlements located in the lowlands of the Albenga plain with the mountain pastures above 1000 m a.s.l., c. 20 km inland (Maggi and Nisbet 1990), possibly on a seasonal basis (Barker et al. 1990).
Materials and methods
Archaeological assemblage and chronology
The faunal assemblage from Tana del Barletta was examined for the presence of bovine and caprine teeth for isotopic analysis. Although the overall number of bovid remains found in the cave amounts to more than a thousand fragments (Barker et al. 1990), teeth are rare and all of them were analysed. In total, ten teeth were recovered: six bovine maxillary teeth (enamel bioapatite and dentine collagen analysis), two caprine mandibular teeth and two caprine maxillary teeth (enamel bioapatite analysis) (Fig. 3). Five of the teeth are dated to phase 3 (representing at least three different Late Neolithic cattle specimens), four to phase 2 (representing two Late Copper Age sheep/goat specimens), one to phase 1 (representing one Early Bronze Age cattle specimen) (Table 1).

Cattle teeth (from TDB-1 to TDB-6) and sheep/goat teeth (from TDB-7 to TDB-10) from Tana del Barletta. Jaw bones associated with TDB-4–5, TDB-7–8 and TDB-9–10 were sampled for bone collagen analysis. A = anterior side (mesial); P = posterior side (distal). Scale bars = 2 cm. TDB-3 was photographed after removal of a portion for dentine collagen extraction
As the adjacent teeth TDB-4–5, TDB-7–8 and TDB-9–10 were still inserted in the maxilla/mandible, a fragment from each of the associated jaw bones was also sampled for collagen analysis. Molars have been indicated as M1/3, as no reliable discrimination could be made between isolated cattle upper molars on a purely morphological basis. The difficulty in differentiating loose cattle M1 and M2 is well known to zooarchaeologists (Beasley et al. 1993). When it comes to maxillary teeth, the issue is further complicated by the absence of the typically three-lophed shape of third molars, which makes them readily identifiable in mandibular rows. The small size of the assemblage also impeded any sound assumptions based on tooth size.
Both couples of caprine teeth were still in the jaw and allowed for confident identification of M1, M2 and P4 (fourth premolar) (Table 1).
For cattle teeth, the distinctive distally-pointing curvature of the roots was used to separate left- from right-side dentition. Although Payne’s (1973) and Grant’s (1982) tooth wear stages are not normally reported for maxillary teeth, an approximate indication is here provided (Table 1).
Previous zooarchaeological works on Tana del Barletta are limited to a list of taxa and quantification of remains (Barker et al. 1990), without providing data on the minimum number of individuals (MNI) of cattle and caprine specimens. The MNI can at least be estimated as regards the tooth assemblage, on the basis of chronology, tooth laterality, wear and shape (Table 1). In light of these parameters, the assemblage used for this study is likely to represent a minimum of four cattle specimens (TDB-1; TDB-2; TDB-3–4–5; TDB-6) and two sheep/goat specimens (TDB-7–8; TDB-9–10). In the published report for the site, caprine remains included both sheep and goat and were grouped as Ovis/Capra (Barker et al. 1990), while overall, in the late prehistory of the region sheep remains are far more abundant (Rowley-Conwy et al. 2020).
In order to achieve a better determination of the material analysed, specific criteria based on morphological differences in lower dentition were considered, following Halstead et al. (2002). The diagnostic features observable in the assemblage are summarised in Table 2. However, further experimental works on caprine mandibles (Zeder and Pilaar 2010; Gillis et al. 2011) have shown that goat is more difficult to identify than sheep, some of the criteria do not prove particularly robust (e.g. the M1.2 and M1.3 criteria) and there can be a number of potential sources of misidentification (Helmer 2000).
The time span of formation of bovid teeth relevant to this study is as follows: cattle M1, from the last c. 4.5 months in utero to 3–2 months of life; cattle M2, from 1 to 12–13 months; cattle M3, from 9–10 to 23–24 months (Brown et al. 1960); sheep/goat M1 starts to form in utero and reaches completion by 9 months; sheep/goat M2, from 2 to 12 months; sheep/goat P4, from c. 12 to 18–24 months (Weinreb and Sharav 1964; Milhaud and Nézit 1991; Hillson 2005). The timing of tooth development in goat may not overlap exactly, and delays might occur in maxillary teeth relative to mandibular teeth (Vigal and Machordom 1985). However, the rate of tooth growth does not reflect the timing of tooth mineralisation, due to a delay caused by the process of enamel maturation. This phase, where most of the carbonate fraction is deposited, takes c. 6–7 months in cattle (Towers et al. 2014) and c. 5–6 months in sheep/goat (Balasse et al. 2012a), and the apposition of mineralisation fronts makes it virtually impossible to sample discrete growth increments, decreasing the time resolution of sequential sampling (Balasse 2002).
The chronology of the remains is based on the 14C dates available for the superimposed hearths excavated in the 1980s (Del Lucchese et al. 1987; Barker et al. 1990), produced by the laboratory of the Harwell Atomic Energy Research Establishment, along with three new radiometric dates purposely obtained for this study at the Oxford Radiocarbon Accelerator Unit (Table 3). For the new dating programme, we selected a tooth from the lowest hearth (TDB-5), a tooth from the upper hearth (TDB-6) and a tooth from the caprine jaws originally collected in a reworked deposit on the surface of the cave (TDB-8). TDB-5, still in the jaw with TDB-4, was found in the same stratigraphic unit as TDB-1–2–3 and therefore provides a date for them. TDB-8, still in the jaw with TDB-7, was found associated with TDB-9–10. Direct dating of the teeth analysed isotopically was necessary not only to check the accuracy of previous radiocarbon results from charcoal samples, but also to include in the study the caprine teeth, now shown to be Late Copper Age (TDB-7–8–9–10).
All dates were recalibrated in OxCal 4.3 (Bronk Ramsey 2009), using the latest available curve (IntCal13; Reimer et al. 2013). The previous chronology of Barletta phases 3 and 2 (Late Neolithic and Copper Age) was confirmed by the new results and supported by the artefacts recovered during the excavation, which included plain and cordoned ware, a flint arrowhead and a serpentinite axe (Del Lucchese et al. 1987).
Modern assemblage
Given the large prevalence of maxillary teeth in the archaeological assemblage, modern remains of cattle were also sampled, in order to investigate whether the isotopic signals measured in mandibular and maxillary teeth are similar and comparable. The animal belonged to a specimen of Piedmontese breed and was always raised in the same location, a calcareous meadow in Paruzzaro (eastern Piedmont, Italy). In the farm, adults were fed with a mixture of hay, maize, beet, soy, bran and sugarcane, while calves were given milk (90%) and a “microfibre” composed of maize, barley, soy, pea and straw (10%). The head was obtained in September, defleshed by burying it in a biologically active topsoil and recovered one year later. The teeth were then extracted using a Dremel cutting wheel. On the mandible and maxilla of the specimen examined, deciduous premolars (p2, p3, p4) appeared to be still in place, while the permanent first molar (M1) had erupted, and the permanent second molar (M2) had become visible.
The precise age at death of the animal was unknown; therefore, it was estimated with reference to the tooth development stage. Based on permanent molars’ eruption timing (M1: between 6 and 12 months; M2: between 12 and 16 months; Brown et al. 1960; Koarai et al. 2016) and wear stage (Jones and Sadler 2012), the animal was likely to be an immature individual about 7–8 months of age, within Jones-Sadler’s early stage C. In order to increase the reliability of the results and compare the maxillary/mandibular signal twice, both the left and right sides of the jaws were sampled. Due to the incomplete stage of formation of second molars, only first molars were sampled for stable carbon and oxygen isotope analysis.
Sampling and analysis of enamel bioapatite (δ18O, δ13C)
Bucco-mesial lobes were selected as sampling points due to enamel thickness and easiness of sampling (Towers et al. 2014). A conventional sampling method drilling the enamel with a diamond-tipped abrasive drill bit perpendicularly to the tooth growth axis was adopted. Depending on the length of the crown, from 7 to 12 (cattle) and from 3 to 7 (sheep/goat) samples per tooth were taken from archaeological material, and from 9 to 11 from modern material. Around c. 8–12 mg (cattle) and c. 4–6 mg (sheep/goat) of enamel powder were collected. Following Balasse et al. (2002), samples were reacted with acetic acid 0.1 M (approximately 0.1 mL solution/0.1 mg powder), at room temperature for 4 h, then rinsed five times and freeze-dried. All samples were reacted with 100% phosphoric acid at 70 °C for 1 h. An amount varying between 0.6 and 0.8 mg of enamel powder was used for measurements. The CO2 produced was analysed on a Thermo Delta V Advantage isotope ratio mass spectrometer coupled to a Thermo gas chromatography-based GasBench II system via a Conflo IV interface at the Chemical Analysis Facility (CAF) of the University of Reading, UK.
Values were calibrated with the international standards NBS 19 (limestone; accepted values: δ18O = − 2.20‰, δ13C = + 1.95‰) and LSVEC (lithium carbonate; values used in this study: δ18O = − 26.40‰, δ13C = − 46.60‰) using a two-point calibration curve with drift correction. All values are reported in per mil relative to the Vienna Pee Dee Belemnite (VPDB) scale. Over the period of analysis, measured values for NBS 19 were − 2.19 ± 0.14‰ (δ18O) and 1.97 ± 0.10‰ (δ13C), while for LSVEC these were − 26.44 ± 0.16‰ (δ18O) and − 46.57 ± 0.11‰ (δ13C). The accuracy (percent error), based on repeat analysis of an internal carbonate standard (Reading limestone calcite—RLC; accepted values: δ18O = − 1.77‰, δ13C = + 2.04‰), was 4.91% for δ18O and 1.05% for δ13C. The analytical precision (1σ std. dev.), based on repeat analysis of an internal standard of modern horse enamel prepared at the Reading stable isotope laboratory (Reading horse enamel—RHE), was around 0.20‰ for δ18O and 0.12‰ for δ13C (see Online Resources 1–2).
Sampling and analysis of dentine and bone collagen (δ15N, δ13C)
Before sampling, all material was cleaned using abrasive dental drill bits. Collagen extraction followed Longin (1971), with modifications according to Collins and Galley (1998). Using a Dremel diamond cutting wheel, a whole tooth loph was cut perpendicularly to the axis into c. 2-mm thick slices (c. 100–500 mg), and when still inserted in the jaw a portion of the bone (c. 250–450 mg) was also sampled. Tooth slices and bone fragments were dissolved for 2–4 days in refrigerated 0.5 HCl, rinsed in acidic (pH 3) deionised water and heated up to 70 °C. Residues were recovered using a 60-90-μm Ezee® filter. The samples were then freeze-dried for 48 h, prior to weighting into tin capsules and duplicate analysis with a Sercon Elemental Analyzer coupled to a Europa Geo 20–20 isotope ratio mass spectrometer at the University of Reading, UK. Values were calibrated using a two-point calibration curve with drift correction, using internal working standards calibrated to internationally certified reference materials including the amino acid methionine (Elemental Microanalysis/MethR), powdered Bovine Liver Standard (NIST1577a-BLS) and a batch of pork gelatine and fish skin prepared at the Reading stable isotope laboratory (Reading pork gelatine—RPG; accepted values: δ15N = 5.04‰, δ13C = − 21.54‰; Reading fish skin—RFS; accepted values: δ15N = 13.99‰, δ13C = − 15.64‰). All δ15N values are reported in per mil relative to the atmospheric N2 (AIR), and δ13C values in per mil relative to the VPDB scale. Analytical precision (1σ std. dev.) over the period of analysis, determined by repeat analysis of internal standards (RPG and RFS), was ± 0.19‰ for δ15N and ± 0.06‰ for δ13C (see Online Resources 3–5).
Reference values of environmental water
In the absence of direct isotope measurements of water sources from around the site, reference data on 18O abundance in meteoric water from atmospheric precipitation (henceforth δ18Omw) were obtained using the Online Isotopes in Precipitation Calculator (OIPC 3.1; Bowen and Wilkinson 2002; Bowen 2019). Following the algorithms by Bowen and Revenaugh (2003) and Bowen et al. (2005), monthly and mean annual δ18Omw values were calculated for three locations: the cave site, the coastal plain of Albenga (c. 12 km from the site, 10–20 m a.s.l.) and the upland pasture of Mt. Galero (c. 4.5 km from the site, 1700 m a.s.l.) (Table 4). The values interpolated have been checked against direct measurements of environmental water available from the nearest pluviometric stations included in the database of the Global Network for Isotopes in Precipitation (GNIP) (IAEA/WMO 2015): Genoa and Monaco for the coast and Bossea for the upland area (Table 4). Most of these instrumental data were collected before the global rise of temperatures following the first years of the twenty-first century. A slight offset is noticeable, OIPC estimates for the coast being ~ 3‰ lower in winter months relative to the average winter values from the IAEA stations of Genoa and Monaco, and ~ 2‰ higher in summer months relative to the average summer values from the station of Genoa, while the offset is slightly reduced for winter values at high elevations (~ 1–2‰, Mt. Galero and Bossea IAEA station). Summer values are not available for Bossea, as the warmest month for which data were recorded is March (− 8.8‰), and among the winter months the lowest values occur in November (but data are lacking for January).
Interpolations from the three locations show a large intra-annual range, spanning from 8.1 (coastal plain) to 9.4‰ (upland pastures). However, the decade-spanning IAEA records available for Genoa and Monaco indicate a smaller seasonal range (~ 3‰ for Genoa, from − 6.5 to − 3.4‰ and ~ 5.5–6‰ for Monaco, from − 6.9 to − 1.2‰).
Relationship between oxygen isotopic composition of structural carbonate and environmental water
Specific formulas for conversion of enamel δ18O values (δ18Oen) into δ18Omw were applied to the dataset from Tana del Barletta. Although not appropriate for every geographical location (Janzen et al. 2020), these formulas, used before in key studies of herd mobility (Pellegrini et al. 2008; Britton et al. 2009), and routinely applied also in later Quaternary and pre-Quaternary works (e.g. Pushkina et al. 2020; Owocki et al. 2020; Pederzani et al. 2021), have received criticism by Pollard et al. (2011). However, in the Alpine region, a significant correlation between water isotopic composition derived from archaeological cattle bones, altitude and modern water was found by Mayr et al. (2016). Moreover, there is a paucity of comparable δ18O(PO3-4) values from the study region where the site is located (the choice suggested by Pollard et al. 2011); therefore, conversion from δ18Oen to δ18O(H2O) represents the only applicable approach to tackle issues of ancient mobility. Following such considerations, we decided to use these equations, with the baseline assumption of their rough reliability (Table 5).
After conversion of values from PDB to SMOW scales according to Werner and Brand (2001) (1) and estimations of phosphate bioapatite δ18O values from bioapatite structural carbonate δ18O values according to Iacumin et al. (1996) (2), oxygen isotope ratios in bone phosphate were converted into oxygen isotope ratios in precipitation, as calculated using the inverse form of the equations for cattle (3) and sheep (4) by D’Angela and Longinelli (1990):
The last step was also performed according to the inverse form of the equation for mammals (5) by Amiot et al. (2004):
However, using Eq. (5), the changes in the values appeared to be negligible in the case of cattle (≤ 0.4) but up to 2‰ lower in the case of caprines. Therefore, the data given below will only refer to the values obtained using the genus-specific Eqs. (3) and (4) by D’Angela and Longinelli (1990).
Results
Stable oxygen and carbon isotopes of modern cattle enamel
In the modern cattle, δ18O values range between − 5.4 and − 11.1‰ and δ13C values between − 9.9 and − 11.5‰. The intra-tooth variation ranges between 4.4 and 4.9‰ for the δ18O signal and is equivalent to or less than 1‰ for the δ13C signal (Table 6). Overall, the pattern of intra-tooth variations in both mandibular M1 and in the left maxillary M1 is very similar: it follows an increasing slope for the δ18O signal, while the δ13C sequences appear rather flat (Fig. 4). However, the upper right M1, despite its general increasing trend towards the ERJ, shows two drops, one of which is within the most recent part of the sampled enamel.

Plots showing the δ18O and δ13C variation in bioapatite enamel from maxillary and mandibular first molars of a modern calf
The maxillary M1 from both sides exhibit δ18O values comprised between − 5.4 and − 10.7‰/ − 10.8‰. Mean δ18O values are − 8.1‰ and − 8.0‰, and the intra-tooth range 5.4 and 5.3‰ for the left and right maxillary M1, respectively. The corresponding mandibular δ18O maxima and minima from both sides are between − 6.0‰/ − 6.3‰ and − 10.6‰/ − 11.1‰. Mean δ18O values are − 8.7‰ and − 8.3‰, and the intra-tooth range is 4.8 and 4.6‰.
In the maxillary molars, δ 13C values are comprised between − 11.5 and − 10.2‰ (averaging − 10.6‰ and − 10.8‰). In mandibular molars, δ 13C values are comprised between − 11.1 and − 9.9‰ (averaging − 10.5 and − 10.4‰).
Stable oxygen and carbon isotopes of archaeological cattle enamel
Bovine teeth display δ18O values comprised between − 8.6 and − 3.9‰ (Table 1; Fig. 5). Although some reduced intra-tooth variations are visible, the intra-tooth range between maximum and minimum δ18O values is rather narrow, even in teeth representing large time spans such as the M2/3 (from 0.9 to 2.9‰, averaging only 1.6‰). TDB-1 and TDB-6 (M1/3) show a similar, decreasing and finally newly increasing pattern of δ18O towards the ERJ, while in TDB-4 (M1/2) δ18O values increase towards the ERJ, and in TDB-2–3–5 (M1/3 and M2/3) the δ18O sequences are relatively flat.

Plots showing the δ18O and δ13C variation in bioapatite enamel within cattle and sheep/goat teeth from Tana del Barletta (* = sample possibly contaminated by dentine)
δ 13C values are relatively homogenous (ranging from − 14.6 to − 11.3‰), and intra-tooth variations are small (averaging 1.2‰).
Stable oxygen and carbon isotopes of archaeological caprine enamel
Caprine δ18O values range from − 7.8 to − 1.2‰ (Table 1; Fig. 5). Overall, apart from TDB-10, sheep/goat teeth exhibit higher δ18O values than cattle teeth. TDB-7 and TDB-10 (both M1) show a similar, decreasing, pattern of δ18O variations. The intra-tooth range between maximum and minimum δ18O values is quite limited (from 0.8 to 3.2‰, averaging only 1.9‰).
δ 13C values are relatively homogenous (from − 14.6 to − 11.9‰), with small intra-tooth variations (averaging 1.4‰).
Stable nitrogen and carbon isotopes of archaeological dentine and bone collagen
The C:N ratios (range 3.1 − 3.5), C content (range 19.6 − 43.6%) and N nitrogen content (range 6.7 − 15.7%) of all dentine samples fall within the range of reliable values (DeNiro 1985; Ambrose 1990). Collagen yield percentages in dentine (between 2.7 and 9.3%, averaging 6.5%) are also sufficient to produce reliable results (Van Klinken 1999) (Online Resource 5). The isotopic composition of cattle collagen appears extremely stable across tooth dentine. All cattle teeth exhibit δ15N values between 3.5 and 5.9‰, with an intra-tooth range often very limited (≤ 0.3‰). δ 13C values are equally homogeneous (from − 22.1 to − 21.2‰), with most of the teeth having particularly small intra-tooth variations (in five cases ≤ 0.4‰) (Table 7; Fig. 6).

Plots showing the δ15N and δ13C variation in dentine collagen within cattle teeth from Tana del Barletta
The C:N ratios (range 3.2–3.3), C content (range 39.1–43.7%) and N content (range 14.1–15.7%) of the archaeological bone fragments (cattle maxilla enclosing TDB-4–5 and upper and lower caprine jaws associated with the contiguous teeth TDB-7–8 and TDB-9–10) fall in the accepted range for well-preserved collagen (DeNiro 1985; Ambrose 1990), except for the jaw related to TDB-9–10, which therefore cannot be taken into account (Online Resource 5). Bone samples exhibit δ15N and δ13C values very close to those recorded in dentine samples (Table 7).
Discussion
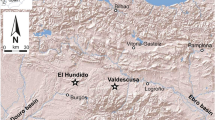
To shed light on the subsistence strategies of late prehistoric societies settled in Liguria, it will be of paramount importance to assess whether caves containing animal remains were permanently occupied or used as seasonal shelters (Barker et al. 1990; Rowley-Conwy 1997, 2000; Rowley-Conwy et al. 2020). As regards Tana del Barletta, the cave had previously been interpreted as a pastoral shed aimed at the seasonal exploitation of the nearby pastures of Mt. Galero (Fig. 7, triangle), while the plain of Albenga may have been home to one or more permanent villages (Maggi and Nisbet 1990) (Fig. 7, circle).

Map showing the location of Tana del Barletta (star), the mountain pastures of Mt. Galero (triangle), the coastal plain of Albenga (circle) and the IAEA (International Atomic Energy Agency) pluviometric stations considered in this study (1 = Genoa; 2 = Bossea; 3 = Monaco)
Stable oxygen isotopes are useful indicators to gain insights into the environment grazed by the livestock penned in the cave, as the δ18O of tooth enamel broadly reflects the oxygen isotopic composition of ingested water, without undergoing remodelling once mineralisation has occurred (Kennedy 2018). In obligate drinking animals (e.g. Bos sp.), the relationship between the isotopic composition of the water intake and enamel bioapatite is particularly strong, as the composition of the latter largely reflects the δ18O of the dominant water source (Pederzani and Britton 2019; Pederzani et al. 2021). Therefore, cattle teeth from Tana del Barletta can offer precious information on the isotopic composition of ingested water, which is in turn controlled by environmental and geographical parameters.
Mandibular and maxillary isotopic composition of cattle first molars
Just a handful of studies have specifically reported the use of maxillary teeth to investigate the isotopic composition of incrementally formed tissues (Balasse et al. 2003, 2017, 2021; Towers et al. 2011; Makarewicz 2017a; Makarewicz and Pederzani 2017), and only very recently archaeological and modern upper molars (from sheep) have been specifically examined (Balasse et al. 2020). In turn, having a majority of studies based on mandibular dentition encourages analysts to select solely archaeological mandibular teeth to obtain comparable data, thus enlarging the gap.
Using a modern skull, the isotopic pattern recorded in maxillary and mandibular cattle M1 was compared twice (sampling left-sided and right-sided molars). Although the time span of formation of M1 crowns does not embrace a full annual cycle (Brown et al. 1960), there is a very good match between the range of maximum and minimum δ18O values recorded in cattle lower molars of both sides and their upper counterparts (less than 1‰ offset). The similarity of the intra-tooth pattern is also evident, despite a slight difference between the right maxillary M1 and the other M1s. The former shows two drops in the values (Fig. 4), which are not seen in the left maxillary M1 and are likely due to an incomplete mineralisation towards the enamel-root junction, noted during sampling, or to the drilling procedure (e.g. deep sampling causing a partial inclusion of dentine).
It is possible that the increasing pattern of variation observed in modern M1s was influenced by the pre-weaning intake of maternal milk. However, the δ18O values of this latter are strongly related to the same geographical and seasonal parameters determining the δ18O values of drinking water (Crittenden et al. 2007), which in the teeth analysed appear to equally affect both mandibular and maxillary tooth apatite.
The results suggest that the oxygen isotopic composition of cattle mandibular and maxillary first molars does not differ substantially (cf. Makarewicz and Pederzani (2017) and Balasse et al. (2020) on lower and upper sheep molars). However, potential variations, especially in other molars (Balasse et al. 2021), should be investigated with larger datasets, to assess the degree of inter-individual variability (Robinson et al. 1995; Towers et al. 2014). This would be highly relevant for isotopic studies of cattle dentition from archaeological contexts, in the event of absence of mandibular teeth in the faunal assemblage (see e.g. Balasse et al. 2021) and availability only of maxillary teeth for intra-tooth sequential analysis.
Low values and narrow amplitude of variation in stable oxygen isotope series
As regards stable oxygen isotopes from the archaeological assemblage, due to the presence of both worn and unworn molars which cannot be precisely identified (M1/3), it is possible that at least in some teeth the results are influenced by the lactation signal, reflecting in M1 the composition of mother’s body water rather than that of meteoric water (Fricke and O’Neil 1996; Franz-Odendaal et al. 2003; Henton et al. 2010). However, the contiguous maxillary molars TDB-4 (M1/2) and TDB-5 (M2/3) are particularly promising to observe the isotopic composition of cattle water intake over a long period, as these teeth represent the longest time span observable for an individual cow in the dataset obtained. According to the known data on cattle tooth development, the series TDB-4–5 is likely to represent approximately one year of life or slightly more (Brown et al. 1960). However, this can only be taken as an approximation, as measured δ18O values refer to the bioapatite composition after mineralisation, which mostly occurs during the maturation phase (Robinson et al. 1995), lasts at least 6–7 months and may not always progress at a perfectly uniform rate (Towers et al. 2014). The highest and the lowest values in the series, − 4.6‰ and − 7.5‰, were selected to calculate the range of δ18O values of the water ingested by the specimen. By applying conversion equations from δ18Oen to δ18Omw, we obtained − 7.7‰ (warmest period) and − 10.6‰ (coldest period) (Table 5). If only the c. 1-year time span of TDB-5 is considered, the extremes range from − 9.5 to − 10.6‰. Taking into consideration also the other available cattle teeth, among the Late Neolithic molars, the lowest value still occurs within TDB-5, corresponding to − 10.6‰ of ingested water, and the highest one in TDB-1, corresponding to − 7.0‰ of ingested water. The minimum value recorded in the only Early Bronze Age molar (TDB-6) is even lower, corresponding to − 11.7‰ of ingested water. These results allow us to stress two points which appear evident and require an attempt of explanation: markedly low values and unusually narrow range of amplitude of δ18Oen values, even in teeth formed over 1 year (Fig. 5).
It is apparent that the δ18Omw values obtained from δ18Oen, ranging in obligate drinking cattle from − 11.7 to − 7.0‰, are consistently lower than expected, especially as regards the supposedly summer values. The lowest δ18Omw values (− 11.7‰ and − 10.6‰, TDB-6 and TDB-5) are as low as the modelled winter values from the site and winter instrumental data from the weather station of Bossea (Table 4). Considering a certain degree of dampening affecting isotopic intra-tooth records (Passey and Cerling 2002; Hoppe et al. 2004; Blumenthal et al. 2014; Green et al. 2018), largely due to the sampling method (Zazzo et al. 2005; Pederzani et al. 2021), the actual value of the water intake corresponding to the δ18Oen minima may have been even lower, compatible with δ18Omw modelled values from mountain pastures located above the site.
On the other hand, the highest enamel-derived δ18Omw values (− 7.0‰ and − 7.4‰, TDB-1 and TDB-2) are too low to appear compatible with the modelled (− 1.5‰) and instrumentally recorded (~ 3‰) composition of meteoric water from the coastal plain (especially during warm months), even allowing for dampening effect (Fig. 8). As a confirmation, the series TDB-4/TDB-5, formed over at least one year, never reaches values higher than − 7.7‰. The potential impact of lactation on the data could not fully explain this anomaly, as it would have probably resulted in slightly higher pre-weaning values (Fricke and O’Neil 1996; Franz-Odendaal et al. 2003), due to the presence of δ18O-enriched plant water in cow’s milk (Camin et al. 2008).

Amplitude of the calculated δ18O range of water ingested by cattle (horizontal rectangle) and ranges of each tooth (vertical white rectangles). Grey vertical rectangles show the seasonal amplitude of meteoric water interpolated for the cave site, mountain pastures and coastal plain, and the seasonal amplitude of meteoric water measured at the weather stations of Genoa and Monaco, along with November-March values from Bossea (instrumental data from this location are lacking for the summer period)
It is difficult to provide a straightforward interpretation for the results of stable oxygen isotope analysis, as direct measurements of water across the altitudinal gradient would be needed, and the number of samples should be increased including more sites. However, at least, as regards Tana del Barletta, a couple of possible scenarios can be advanced.
Livestock management scenarios inferred from stable oxygen isotope series
According to the currently available data, the total oxygen isotope amplitude of water ingested by cattle mostly overlaps with the annual range of values expected from relatively high elevations (Fig. 8), suggesting isotopically depleted inputs all-year-round. This fact would have relevant implications in terms of herd management and use of the site, as it would point to a more structured occupation of the highlands, where high-altitude caves do not serve only as seasonal or occasional sheds, but as stable outposts integrated into a pastoral system intensively exploiting upland territories. This view may be contextualised by the presence of human remains pertaining to c. 20 individuals of all ages recovered at the cave of Grotta del Pertuso (1330 m a.s.l.), to explain which Maggi and Nisbet (1990) hypothesised the existence in the area of a blood-tied clan engaged in herding activities.
On the other hand, the isotopic range of the calculated cattle water intake (probably reduced due to signal averaging from amelogenesis) also partly overlaps with winter values from the lowlands (Fig. 8), leaving open the possibility that a certain seasonal mobility occurred, which may have included grazing at low altitudes during cold months.
In addition, more subtle factors may be also responsible for low values of ingested water. For example, particularly negative measurements may be due to local environmental parameters, such as the impact of snow melting on the drinking sources accessed by the animals or the intrusion of isotopically lighter groundwater (Rozanski et al. 1993; Geyh et al. 2000; Hermes et al. 2018). The ingestion of water from buffered sources such as large lakes and rivers, isotopically different from local precipitation (Gonfiantini 1986; Pederzani et al. 2021), can be ruled out, given their absence in the study area.
Also, human behaviour can affect the signal, such as herders provisioning water to the animals from 18O-depleted sources (Makarewicz 2017b). In this regard, available meteorological records from the station of Mt. Settepani (1375 m a.s.l.) show the presence of c. 50–100 cm of snow nowadays on the area of Mt. Galero during harsh winters (ARPA 2020). Similar environmental variables, as well as frequent grazing at high elevations, may also have affected the unexpectedly low oxygen isotope ratio of meteoric water calculated from cattle and caprine bone at the coastal cave of Arene Candide (D’Angela and Longinelli 1993).
Moreover, the effect of percent error and variation in analytical precision observed in δ18O measurements is too reduced to determine such low values (see above, and Online Resource 1), and even the use of equations to estimate the oxygen isotopic composition of meteoric water, which according to Pollard et al. (2011) and Pryor et al. (2014) should bear an uncertainty of c. ± 1–2‰, may not be enough to account for these results. Finally, the inferred low isotopic values of precipitation cannot be attributed to cooler climatic conditions, as the dates of all samples analysed in this study fall within the end of the Atlantic and the full Subboreal periods, when temperatures were broadly similar to and not cooler than in the current Subatlantic (Marcott et al. 2013). Mean July air temperatures reconstructed for the northern Apennine area also appear rather uniform over the last 5000–6000 years (Samartin et al. 2017), and the Abies forests were already declining, paving the way to the spread of Fagus woodland, which still is the dominant biome in the lower montane belt (Branch and Marini 2014; Branch et al. 2014; Arobba et al. 2018).
Whichever scenario best describes the situation occurring at Tana del Barletta, the flatness of the δ18Oen trends (Fig. 5) contrasts with the typically sinusoidal pattern usually observed in bovid enamel from high- and mid-latitudes (e.g. Balasse et al. 2012a, b; Towers et al. 2014, 2017; Knockaert et al. 2018), often showing a large (≥ 3.5‰) intra-tooth variation due to seasonality (e.g. Buchan et al. 2016; Makarewicz et al. 2017; Tornero et al. 2018), unlike records from low latitudes (Janzen et al. 2020). In this study, four cattle teeth exhibit a maximum intra-tooth range below 1.5‰, and no clear and pronounced S-shaped patterns are identifiable in the dataset, with the exception of TDB-1, along perhaps with attenuated curves in TDB-4 and TDB-6. The same situation also concerns the only sheep/goat second molar (TDB-8), characterised by a flat curve and a small intra-tooth range (1.0‰). Some seasonal fluctuations may be captured by caprine first molars TDB-7 and TDB-10, but their record does not span a complete annual cycle (Weinreb and Sharav 1964; Milhaud and Nézit 1991; Zazzo et al. 2010). Truncated annual sequences may also be partly responsible for apparently reduced intra-tooth δ18O ranges in cattle enamel, as, unlike in teeth attached to the jaws, identification as M1, M2 and M3 is not possible for loose two-lophed maxillary molars.
However, if flattened δ18O patterns reflect reality, as it is likely at least in the case of the TDB-4–5 series, this anomaly may be accounted for within the model pushed forward by Britton et al. (2009), according to which migration from cooler summer locations to warmer winter locations results in a buffered pattern lacking the seasonal extremes (cf. Henton 2012). Seasonal movement from the mid-slope zone around the site and the mountain-top pastures may have played a role in evening out the δ18O gradient observable in the analysed samples, especially reduced in cattle molars. Further intra-tooth sequences spanning long periods from Ligurian upland sites will be crucial to reach a firmer conclusion.
The difference between evaporation-insensitive (cattle) and evaporation-sensitive (caprines) taxa appears clearly in the c. 1-year-spanning sheep/goat contiguous series TDB-7–8 (M1 + M2). The oxygen isotope maxima and minima recorded for this specimen are ~ 2–2.5‰ higher than the corresponding extremes in cattle samples, probably as a result of 18O-enriched leaf water in the ingested browse caused by evaporative fractionation (Kohn 1996; Kohn et al. 1998). A different picture results from the sheep/goat series TDB-9–10 (P4 + M1), characterised by a particularly low minimum value (− 7.8‰), corresponding to − 9.0‰ of atmospheric precipitation, which suggests for this specimen access to the same open water sources ingested by cattle during the cold season. However, in winter, a smaller offset between obligate and semi-obligate drinkers should be expected (Makarewicz 2017a). Further inferences are not possible, as this series does not represent a full seasonal cycle, due to the timing of formation of caprine first molar crowns (c. 6–9 months; Weinreb and Sharav 1964; Milhaud and Nézit 1991; Zazzo et al. 2010).
Assessing coastal grazing through nitrogen stable isotopes
The nitrogen isotopic composition of collagen has been used in previous studies to track the inclusion of 15N-enriched coastal vegetation in the diet, but to date the approach had not been applied to Mediterranean settings. All samples from Tana del Barletta exhibit δ15N values with reduced intra-tooth range (~ 1‰ or less) consistently comprised between 3.5 and 5.9‰, falling within the expected range for herbivores feeding exclusively on inland plants (Schoeninger et al. 1983) (Table 7; Fig. 6). The limited amplitude of intra-tooth variations may partly be an effect of sampling through the whole dentine thickness, in addition to further complications due to the formation of secondary dentine later in life (Frank and Nalbandian 1989; Balasse 2003). Interestingly, the M1/2 and M2/3 associated with maxillary bone show decreasing mean values (TDB-4: 4.4 ‰; TDB-5: 3.9 ‰; bone: 3.2 ‰), possibly reflecting the cessation of milk consumption due to the weaning process (Balasse et al. 2001). In the same sample sets, an opposite trend is visible in the δ13C values, which in case of an M1 + M2 series could reflect the development of the rumen (Towers et al. 2014).
Considering a 3–5‰ enrichment up the food chain (Schoeninger and DeNiro 1984; Bocherens and Drucker 2003), the extremes of the collagen δ15N values (+ 3.5 in TDB-1 and + 5.9 in TDB-3; Table 7) point to ingestion of plants having a total δ15N range included c. between − 1.5 and + 3‰. Such nitrogen isotopic composition is standard for inland C3 vegetation, with a certain degree of variation due to differential net soil nitrogen mineralisation rates (Wada et al. 1975; Kahmen et al. 2008; Kristensen et al. 2011). The values obtained are well below the 8–9‰ threshold suggestive of coastal grazing identified in northwest European case studies (Britton et al. 2008; Müldner et al. 2014). Accordingly, the light carbon isotopic composition of cattle dentine from Tana del Barletta rules out any relevant dietary contribution of halophytic and marine taxa. In spite of its near-coast position, a similar situation has been recorded at the cave of Arene Candide, where relatively low δ15N values from herbivore bone collagen (~ 4–5‰) were found (Francalacci and Borgognini Tarli 1988; Le Bras-Goude et al. 2006).
Dietary and environmental insights from carbon stable isotopes
The carbon isotopic composition of both enamel and collagen (δ13Cen, δ13Ccoll) appears extremely steady throughout tooth development in all bovine and caprine specimens (from ~ − 14.5 to ~ − 11.5‰ for δ13Cen and from ~ − 22 to ~ − 21‰ for δ13Ccoll, with a ~ 9‰ spacing between δ13Cen and δ13Ccoll, close to the offset reported by Towers et al. 2011). Following fractionation (~ 12–15.5‰ enrichment relative to diet for enamel and ~ 5‰ enrichment for collagen; van der Merwe 1982; Passey et al. 2005; Zazzo et al. 2010), the obtained δ13Cen and δ13Ccoll values reflect a dietary signature between ~ − 30‰ and ~ − 23.5‰. After correction for the fossil fuel effect, which causes a ~ 1.5‰ depletion in modern atmospheric CO2 (Francey et al. 1999), the dietary signature ranges between − 31.5 and − 25‰. These 13C/12C ratios are comprised within the range of plants following the C3 photosynthetic pathway, and allow us to rule out the ingestion of isotopically heavier 13C-enriched vegetation, such as expected in case of consumption of halophytes (e.g. Salsola sp.) and seaweeds (Tauber 1981). The lowest extreme would fall on the threshold value (~ − 31.5‰) indicating a closed canopy according to Kohn (2010) and slightly beyond the one considered by Balasse et al. (2012b). This might be indicative of animal grazing within the understorey of a densely forested environment, in agreement with the charcoal analysis suggesting a mixed deciduous woodland around the site (Del Lucchese et al. 1987), or point to the intake of 13C-depleted leafy fodder (Balasse et al. 2012b).
Conclusions
Given the lack of knowledge regarding the reliability of maxillary cattle teeth to record oxygen and carbon isotope fluctuations, we assessed intra-tooth variations in four upper and lower M1s from a modern specimen. The measured values show similar patterns and very close minima, maxima and range of amplitude. No shifts between lower and upper M1s were observed. However, a c. 5-month discrepancy has been estimated between maxillary and mandibular M3 isotopic records (Balasse et al. 2021). Further research is needed to clarify this issue and better understand the differences in the isotopic signal between upper and lower dentition. This is particularly relevant for archaeological assemblages, where mandibular teeth may not always be available (see e.g. Balasse et al. 2021).
According to the available estimates for the oxygen isotopic composition of environmental water, both maximum and minimum δ18O values of cattle enamel appear particularly low. This may point to the exploitation of high elevations throughout the year, which may have involved a prolonged presence of the herd between the site and upland pastures. In this regard, it should be stressed that, between the 4th and 3rd Millennium BC, palaeobotanical and sedimentological records from the Ligurian mountains suggest anthropogenic disturbance associated with woodland clearance, in order to increase the availability of pastureland (Maggi and Campana 2008; Cruise et al. 2009; Branch and Marini 2014). However, according to the estimates for the isotopic composition of precipitation, for the specimens of Tana del Barletta the input of water from the lowlands in the summer months cannot be ruled out, suggesting the possibility of seasonal animal mobility. Such a scenario would be compatible with a view proposed by previous works, which indicated the transition to the Late Neolithic (marked in Liguria by the Chassey culture) as a possible onset for transhumant pastoralism (Binder et al. 2008; Maggi and Campana 2008).
It is nevertheless unlikely that grazing occurred directly on the coastal plain, as shown by the nitrogen isotopic composition of cattle dentine collagen, the values of which rule out the ingestion of salt-tolerant vegetation or seaweed. This is confirmed by the carbon isotopic composition of cattle dentine and enamel, which falls within the range of inland C3 plants.
The amplitude of the δ18O intra-tooth variation from both cattle and sheep enamel often appears particularly narrow. This pattern contrasts with the characteristic sinusoidal curve observed repeatedly in archaeological and modern domestic herds at temperate latitudes. Such a rather homogeneous trend can be partly explained by assuming some form of seasonal vertical movement of the specimens between the site and a potential lowland settlement (Barker et al. 1990; Maggi and Campana 2008). This would reduce the observable fluctuation, due to the use of drinking sources from higher altitudes in the hot season (18O-depleted), and drinking sources from lower altitudes in the cold season (18O-enriched) (Britton et al. 2009; Henton 2012). However, it should be borne in mind that these considerations are based on the reduced number of samples available from the cave, and larger assemblages from a variety of sites should be analysed in order to reach more secure conclusions which can be generalised on a regional scale.
Finally, the diverse geology characterising the area surrounding the cave in a range of few kilometres appears particularly promising for a future application of strontium isotope analysis to the tooth assemblage from Tana del Barletta. Along this, we aim to obtain direct δ18O measurements from different water sources at various elevations, assessing whether localised environmental variables may have contributed to the markedly low δ18O maximum values observed in the dataset from the site.
Further investigations of contemporary transhumant herds with known feeding histories from different geographical and environmental contexts will also be helpful to resolve certain equifinalities and provide more robust interpretative frameworks. In particular, there appears to be a paucity of isotope studies on southern European and Mediterranean settings, where only in recent years research has been intensified (e.g. Valenzuela-Lamas et al. 2016; Knockaert et al. 2018; Tornero et al. 2018; Trentacoste et al. 2020) and transhumance is a central topic (Maggi et al. 1990a, b; Barker 2005; Arnold and Greenfield 2006; Carrer and Migliavacca 2019).
Data availability
Not applicable.
Code availability
Not applicable.
Change history
14 August 2022
Missing Open Access funding information has been added in the Funding Note.
19 July 2022
A Correction to this paper has been published: https://doi.org/10.1007/s12520-022-01612-4
References
Acosta ATR, Ercole S (eds.) (2015) Gli habitat delle coste sabbiose italiane: ecologia e problematiche di conservazione. ISPRA, Roma
Ambrose SH (1990) Preparation and characterization of bone and tooth collagen for isotopic analysis. J Archaeological Sci 17(4):431–451
Amiot R, Lécuyer C, Buffetaut E, Fluteau F, Legendre S, Martineau F (2004) Latitudinal temperature gradient during the Cretaceous Upper Campanian-Middle Maastrichtian: δ18O record of continental vertebrates. Earth Planet Sc Lett 226(1–2):255–272
Arnold ER, Greenfield HJ (2006) The origins of transhumant pastoralism in temperate southeastern Europe. In: Robertson EC, Seibert JD, Fernandez DC, Zender MU (eds) Space and Spatial Analysis in Archaeology. Univ of Calgary Pr, Calgary, pp 243–251
Arobba D, Caramiello R, Firpo M (2001) Studio paleobotanico e sedimentologico di un sondaggio alla foce del fiume Centa (Albenga, Liguria occidentale). Allionia 38:147–158
Arobba D, Caramiello R, Firpo M, Mercalli L, Morandi LF, Rossi S (2018) New evidence on the earliest human presence in the urban area of Genoa (Liguria, Italy): a multi-proxy study of a mid-Holocene deposit at the mouth of the Bisagno river. Holocene 28(12):1918–1935
ARPA (2020) ARPA Piemonte. Banca dati meteorologica. https://www.arpa.piemonte.it/rischinaturali/accesso-ai-dati/annali_meteoidrologici/annali-meteo-idro/banca-dati-meteorologica.html. Accessed July 2021
Balasse M (2002) Reconstructing dietary and environmental history from enamel isotopic analysis: time resolution of intra-tooth sequential sampling. Int J Osteoarchaeol 12(3):155–165
Balasse M (2003) Potential biases in sampling design and interpretation of intra-tooth isotope analysis. Int J Osteoarchaeol 13(1–2):3–10
Balasse M, Ambrose SH (2005) Mobilité altitudinale des pasteurs néolithiques dans la vallée du Rift (Kenya): premiers indices de l’analyse du δ C de l’émail dentaire du cheptel domestique. Anthropozoologica 40(1):147–166
Balasse M, Bocherens H, Mariotti A, Ambrose SH (2001) Detection of dietary changes by intra-tooth carbon and nitrogen isotopic analysis: an experimental study of dentine collagen of cattle (Bos taurus). J Archaeolo Sci 28(3):235–245
Balasse M, Ambrose SH, Smith AB, Price TD (2002) The seasonal mobility model for prehistoric herders in the south-western Cape of South Africa assessed by isotopic analysis of sheep tooth enamel. J Archaeol Sci 29(9):917–932
Balasse M, Smith AB, Ambrose SH, Leigh SR (2003) Determining sheep birth seasonality by analysis of tooth enamel oxygen isotope ratios: the Late Stone Age site of Kasteelberg (South Africa). J Archaeol Sci 30(2):205–215
Balasse M, Obein G, Ughetto-Monfrin J, Mainland I (2012a) Investigating seasonality and season of birth in past herds: a reference set of sheep enamel stable oxygen isotope ratios. Archaeometry 54(2):349–368
Balasse M, Boury L, Ughetto-Monfrin J, Tresset A (2012b) Stable isotope insights (δ18O, δ13C) into cattle and sheep husbandry at Bercy (Paris, France, 4th millennium BC): birth seasonality and winter leaf foddering. Environ Archaeol 17(1):29–44
Balasse M, Renault-Fabregon L, Gandois H, Fiorillo D, Gorczyk J, Bacvarov K, Ivanova M (2020) Neolithic sheep birth distribution: results from Nova Nadezhda (sixth millennium BC, Bulgaria) and a reassessment of European data with a new modern reference set including upper and lower molars. J Archaeol Sci 118. https://doi.org/10.1016/j.jas.2020.105139
Balasse M, Tresset A, Bălăşescu A, Blaise E, Tornero C, Gandois H, Fiorillo D, Nyerges ÉÁ, Frémondeau D, Banffy E, Ivanova M (2017) Animal Board Invited Review: Sheep birth distribution in past herds: a review for prehistoric Europe (6th to 3rd millennia BC). Animal 11(12):2229–2236
Balasse M, Gillis R, Živaljević I, Berthon R, Kovačiková L, Fiorillo D, Arbogast RM, Bălăşescu A, Bréhard A, Nyerges ÉÁ, Dimitrijević V, Bánffy E, Domboróczki L, Marciniak A, Oross K, Vostrovská I, Roffet-Salque M, Stefanović S, Ivanova M (2021) Seasonal calving in European Prehistoric cattle and its impacts on milk availability and cheese-making. Sci Rep 11(1):1–11
Bareschino A, Del Lucchese A, Formicola V (2013) Condizioni di vita e comportamento funerario in Valle Argentina (IM) tra il IV ed il I millennio a.C. In: Campana N, Del Lucchese A, Gardini A (eds) Archeologia in Liguria, Nuova Serie. Sagep, Genova 4, vol. 4, pp 278–280
Barfield L, Chippindale C (1997) Meaning in the later prehistoric rock-engravings of Mont Bégo, Alpes-Maritimes, France. Proc Prehist Soc 63:103–128
Barker G (2005) Agriculture, pastoralism, and Mediterranean landscapes in prehistory. In: Blake E, Knapp AB (eds) The archaeology of Mediterranean prehistory. Blackwell, Malden-Oxford, pp 46–76
Barker G, Biagi P, Clark C, Maggi R, Nisbet R (1990) From hunting to herding in the Val Pennavaira (Liguria - northern Italy). In: Biagi P (ed) The Neolithisation of the Alpine region. Monografie di Natura Bresciana 13. Brescia, Museo Civico di Scienze Naturali di Brescia, pp 99–121
Beasley MJ, Brown WAB, Legge AJ (1993) Metrical discrimination between mandibular first and second molars in domestic cattle. Int J Osteoarchaeol 3(4):303–314
Binder D, Lepère C, Maggi R (2008) Épipaléolithique et Néolithique dans l’arc Liguro-Provençal: bilan et perspectives de recherche. Bulletin du Musée d’Anthropologie Préhistorique de Monaco, Suppl.:49–62
Bishop KG, Garvie-Lok S, Haagsma M, MacKinnon M, Karapanou S (2020) Mobile animal management in the Mediterranean: investigating Hellenistic (323–31 BCE) husbandry practices in Thessaly, Greece using δ13C, δ18O, and 87Sr/86Sr recorded from sheep and goat tooth enamel. J Archaeol Sci Rep 31:102331
Blumenthal SA, Cerling TE, Chritz KL, Bromage TG, Kozdon R, Valley JW (2014) Stable isotope time-series in mammalian teeth: in situ δ18O from the innermost enamel layer. Geochim Cosmochim Ac 124:223–236
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13(1–2):46–53
Bowen GJ (2019) The online isotopes in precipitation calculator, version 3.1. https://wateriso.utah.edu/waterisotopes/pages/data_access/oipc.html. Accessed July 2021
Bowen GJ, Revenaugh J (2003) Interpolating the isotopic composition of modern meteoric precipitation. Water Resour Res 39(10). https://doi.org/10.1029/2003WR002086
Bowen GJ, Wilkinson B (2002) Spatial distribution of δ18O in meteoric precipitation. Geology 30(4):315–318
Bowen GJ, Wassenaar LI, Hobson KA (2005) Global application of stable hydrogen and oxygen isotopes to wildlife forensics. Oecologia 143(3):337–348
Branch NP, Marini NA (2014) Mid-Late Holocene environmental change and human activities in the northern Apennines, Italy. Quat Int 353:34–51
Branch NP, Black S, Maggi R, Marini NA (2014) The Neolithisation of Liguria (NW Italy): an environmental archaeological and palaeoenvironmental perspective. Environ Archaeol 19(3):196–213
Britton K, Müldner G, Bell M (2008) Stable isotope evidence for salt-marsh grazing in the Bronze Age Severn Estuary, UK: implications for palaeodietary analysis at coastal sites. J Archaeol Sci 35(8):2111–2118
Britton K, Grimes V, Dau J, Richards MP (2009) Reconstructing faunal migrations using intra-tooth sampling and strontium and oxygen isotope analyses: a case study of modern caribou (Rangifer tarandus granti). J Archaeol Sci 36(5):1163–2117
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51(1):337–360
Brown WAB, Christofferson DVM, Massler M, Weiss MB (1960) Postnatal tooth development in cattle. Am J Vet Res 21(80):7–34
Buchan M, Müldner G, Ervynck A, Britton K (2016) Season of birth and sheep husbandry in late Roman and Medieval coastal Flanders: a pilot study using tooth enamel δ18O analysis. Environ Archaeol 21(3):260–270
Camin F, Perini M, Colombari G, Bontempo L, Versini G (2008) Influence of dietary composition on the carbon, nitrogen, oxygen and hydrogen stable isotope ratios of milk. Rapid Commun Mass Sp 22(11):1690–1696
Capelli C, Cabella R, Del Lucchese A, Piazza M, Starnini E (2008) Archaeometric analyses of Early and Middle Neolithic pottery from the Pian del Ciliegio rock shelter (Finale Ligure, NW Italy). Archeosciences-Rev A 32:115–124
Carrer F, Migliavacca M (2019) Prehistoric transhumance in the Northern Mediterranean. In: Sabatini S, Bergerbrant S (eds) The textile revolution in Bronze Age Europe: production, specialisation, consumption. Cambridge Univ Pr, Cambridge, pp 217–238
Chazin H, Gordon GW, Knudson KJ (2019) Isotopic perspectives on pastoralist mobility in the Late Bronze Age South Caucasus. J Anthropol Archaeol 54:48–67
Collins MJ, Galley P (1998) Towards an optimal method of archaeological collagen extraction: the influence of pH and grinding. Anc Biomol 2:209–222
Crittenden RG, Andrew AS, LeFournour M, Young MD, Middleton H, Stockmann R (2007) Determining the geographic origin of milk in Australasia using multi-element stable isotope ratio analysis. Int Dairy J 17(5):421–428
Cruise GM, Macphail RI, Linderholm J, Maggi R, Marshall PD (2009) Lago di Bargone, Liguria, N Italy: a reconstruction of Holocene environmental and land-use history. Holocene 19(7):987–1003
D’Angela D, Longinelli A (1990) Oxygen isotopes in living mammal’s bone phosphate: further results. Chem Geol 86(1):75–82
D’Angela D, Longinelli A (1993) Oxygen isotopic composition of fossil mammal bones of Holocene age: palaeoclimatological considerations. Chem Geol 103(1–4):171–179
Dansgaard W (1964) Stable isotopes in precipitation. Tellus 16(4):436–468
Deines P (1980) The isotopic composition of reduced organic carbon. In: Fritz P, Fontes JCh (eds) Handbook of environmental isotope geochemistry, vol 1. The Terrestrial Environment. A. Elsevier, Amsterdam, pp 329–406
De Lumley H (1995) Le Grandiose e le Sacré, Charly-Yves Chaudoreille, Edisud. La Calade, Aix-en-Provence
Del Lucchese A, De Pascale A (2011) Nuovi dati sulle sepolture in grotta nella Liguria di Ponente. In: L’Età del rame in Italia. Atti della XLIII Riunione Ssientifica, in memoria di Gianni Bailo Modesti, Bologna, 26–29 novembre 2008. Istituto Italiano di Preistoria e Protostoria, Firenze, pp 727–732
Del Lucchese A, Maggi R, Nisbet R (1987) Tana del Barletta. In: Del Lucchese A, Melli P (eds) Archeologia in Liguria III.1. Scavi e scoperte 1982–1986. Tormena, Genova, pp 165–168
Del Lucchese A, Ricci M (1987) Il Pertuso. In: Del Lucchese A, Melli P (eds) Archeologia in Liguria III.1. Scavi e scoperte 1982–1986. Tormena, Genova, pp 169–172
DeNiro MJ (1985) Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317(6040):806–809
Dori I, Varalli A, Seghi F, Moggi-Cecchi J, Sparacello VS (2020) Environmental correlates of growth patterns in Neolithic Liguria (northwestern Italy). Int J Paleopathol 28:112–122
Formicola V, Balestri M (1987) Il materiale scheletrico umano. In: Del Lucchese A, Melli P (eds) Archeologia in Liguria III.1. Scavi e scoperte 1982–1986. Tormena, Genova, pp 172–176
Francalacci P, Borgognini Tarli SB (1988) Multielementary analysis of trace elements and preliminary results on stable isotopes in two Italian prehistoric sites. Methodological aspects. In: Grupe G, Herrmann B (eds) Trace elements in environmental history. Springer, Berlin-Heidelberg, pp 41–52
Francey RJ, Allison CE, Etheridge DM, Trudinger CM, Enting IG, Leuenberger M, Langenfelds RL, Michel E, Steele LP (1999) A 1000-year high precision record of δ13C in atmospheric CO2. Tellus B 51(2):170–193
Frank RM, Nalbandian J (1989) Structure and ultrastructure of dentine. In: Berkovitz BKB, Boyde A, Frank RM, Höhling HJ, Moxham BJ, Nalbandian J, Tonge CH (eds) Handbook of microscopic anatomy, vol. 5/6. Teeth. Berlin-Heidelberg, Springer, pp 173–247
Franz-Odendaal TA, Lee-Thorp JA, Chinsamy A (2003) Insights from stable light isotopes on enamel defects and weaning in Pliocene herbivores. J Biosci 28(6):765–773
Fricke HC, O’Neil JR (1996) Inter-and intra-tooth variation in the oxygen isotope composition of mammalian tooth enamel phosphate: implications for palaeoclimatological and palaeobiological research. Palaeogeogr Palaeoclimatol Palaeoecol 126(1–2):91–99
Fricke HC, Clyde WC, O’Neil JR (1998) Intra-tooth variations in δ18O (PO4) of mammalian tooth enamel as a record of seasonal variations in continental climate variables. Geochim Cosmochim Ac 62(11):1839–1850
Gerling C, Doppler T, Heyd V, Knipper C, Kuhn T, Lehmann MF, Pike AWG, Schibler J (2017) High-resolution isotopic evidence of specialised cattle herding in the European Neolithic. PLoS ONE 12(7):e0180164
Geyh M, D’Amore F, Darling G, Paces T, Pang Z, Šilar J (2000) Environmental isotopes in the hydrological cycle: principles and applications, Vol 4. Groundwater Saturated and Unsaturated Zone. IAEA-UNESCO, Vienna-Paris
Gillis R, Chaix L, Vigne JD (2011) An assessment of morphological criteria for discriminating sheep and goat mandibles on a large prehistoric archaeological assemblage (Kerma, Sudan). J Archaeol Sci 38(9):2324–2339
Gillis R, Bréhard S, Bălăşescu A, Ughetto-Monfrin J, Popovici D, Vigne JD, Balasse M (2013) Sophisticated cattle dairy husbandry at Borduşani-Popină (Romania, fifth millennium BC): the evidence from complementary analysis of mortality profiles and stable isotopes. World Archaeol 45(3):447–472
Giustini F, Brilli M, Patera A (2016) Mapping oxygen stable isotopes of precipitation in Italy. J Hydrol Reg Stud 8:162–181
Gonfiantini R (1986) Environmental isotopes in lake studies. In: Fritz P, Fontes JCh (eds) Handbook of environmental isotope geochemistry, vol 2. The Terrestrial Environment. B. Elsevier, Amsterdam, pp 113–168
Goude G, Müller K, Buscaglia F, Reiche I (2011) Détermination de l’état de conservation, du consolidant et dosages isotopiques (δ13C, δ15N) d’ossements d’anciennes collections anthropologiques. Le cas de la grotte La Pollera (Ligurie, Italie). Archeosciences-Rev A 35:223–233
Goude G, Dori I, Sparacello VS, Starnini E, Varalli A (2020) Multi-proxy stable isotope analyses of dentine microsections reveal diachronic changes in life history adaptations, mobility, and tuberculosis-induced wasting in prehistoric Liguria (Finale Ligure, Italy, northwestern Mediterranean). International J Paleopathol 28:99–111
Grant A (1982) The use of tooth wear as a guide to the age of domestic ungulates. In: Wilson B, Grigson C, Payne S (eds) Ageing and sexing animal bones from archaeological sites. Brit Archaeol Rep Brit Ser 109:91–108
Green DR, Smith TM, Green GM, Bidlack FB, Tafforeau P, Colman AS (2018) Quantitative reconstruction of seasonality from stable isotopes in teeth. Geochim Cosmochim Ac 235:483–504
Halstead P, Collins P, Isaakidou V (2002) Sorting the sheep from the goats: morphological distinctions between the mandibles and mandibular teeth of adult Ovis and Capra. J Archaeol Sci 29(5):545–553
Heaton TH (1987) The 15N/14N ratios of plants in South Africa and Namibia: relationship to climate and coastal/saline environments. Oecologia 74:236–246
Helmer D (2000) Discrimination des genres Ovis et Capra à l’aide des prémolaires inférieures 3 et 4 et interpretation des âges d’abattage: l’exemple de Dikili Tash (Grèce). Ibex - J Mount Ecol 5:29–38
Henton E (2012) The combined use of oxygen isotopes and microwear in sheep teeth to elucidate seasonal management of domestic herds: the case study of Çatalhöyük, central Anatolia. J Archaeol Sci 39(10):3264–3276
Henton E, Meier-Augenstein W, Kemp HF (2010) The use of oxygen isotopes in sheep molars to investigate past herding practices at the Neolithic settlement of Çatalhöyük, central Anatolia. Archaeometry 52(3):429–449
Hermes T, Pederzani S, Makarewicz CA (2018) Ahead of the curve? Implications for isolating vertical transhumance in seasonal montane environments using sequential oxygen isotope analyses of tooth enamel. In: Ventresca Miller AR, Makarewicz CA (eds) Isotopic investigations of pastoralism in prehistory. Themes in contemporary archaeology. Routledge, London, vol. 4, pp 57–76
Hillson S (2005) Teeth. Cambridge Univ Pr, Cambridge
Hoppe KA, Stover SM, Pascoe JR, Amundson R (2004) Tooth enamel biomineralization in extant horses: implications for isotopic microsampling. Palaeogeogr Palaeoclimatol Palaeoecol 206(3–4):355–365
Huet T (2017a) Les gravures piquetées du mont Bego (Alpes-Maritimes) Organisation spatiale et sériation (VIe-IIe millénaire av. J.-C.). Mémoires de la Société Préhistorique Française, 63. Société Préhistorique Française, Paris
Huet T (2017b) New perspectives on the chronology and meaning of Mont Bégo rock art (Alpes-Maritimes, France). Camb Archaeol J 27(2):199–221
Iacumin P, Bocherens H, Mariotti A, Longinelli A (1996) Oxygen isotope analyses of co-existing carbonate and phosphate in biogenic apatite: a way to monitor diagenetic alteration of bone phosphate? Earth Planet Sc Lett 142(1–2):1–6
IAEA/WMO (2015) Global Network of Isotopes in Precipitation. The GNIP Database. https://nucleus.iaea.org/wiser. Accessed July 2021.
Janzen A, Balasse M, Ambrose SH (2020) Early pastoral mobility and seasonality in Kenya assessed through stable isotope analysis. J Archaeol Sci 117. https://doi.org/10.1016/j.jas.2020.105099
Jones GG, Sadler P (2012) Age at death in cattle: methods, older cattle and known-age reference material. Environ Archaeol 17(1):11–28
Jourdain-Annequin C, Duclos JC (eds) (2006) Aux origines de la transhumance. Les Alpes et la vie pastorale d’hier et d’aujourd’hui. Picard, Paris
Kahmen A, Wanek W, Buchmann N (2008) Foliar δ15N values characterize soil N cycling and reflect nitrate or ammonium preference of plants along a temperate grassland gradient. Oecologia 156(4):861–870
Kennedy LF (2018) Mobility and oxygen isotopes. In: López-Varela SL (ed) The encyclopedia of archaeological sciences. Wiley, Hoboken. https://doi.org/10.1002/9781119188230.saseas0386
Knockaert J, Balasse M, Rendu C, Burens A, Campmajo P, Carozza L, Bousquet D, Fiorillo D, Vigne JD (2018) Mountain adaptation of caprine herding in the eastern Pyrenees during the Bronze Age: a stable oxygen and carbon isotope analysis of teeth. Quat Int 484:60–74
Koarai K, Kino Y, Takahashi A, Suzuki T, Shimizu Y, Chiba M, Osaka K, Sasaki K, Fukuda T, Isogai E, Yamashiro H, Oka T, Sekine T, Fukumoto M, Shinoda H (2016) 90Sr in teeth of cattle abandoned in evacuation zone: record of pollution from the Fukushima-Daiichi Nuclear Power Plant accident. Sci Rep 6. https://doi.org/10.1038/srep24077
Kohn MJ (1996) Predicting animal δ18O: accounting for diet and physiological adaptation. Geochim Cosmochim Ac 60(23):4811–4829
Kohn MJ (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proc Natl Acad Sci USA 107(46):19691–19695
Kohn MJ, Schoeninger MJ, Valley JW (1998) Variability in oxygen isotope compositions of herbivore teeth: reflections of seasonality or developmental physiology? Chem Geol 152(1–2):97–112
Körner C, Farquhar GD, Wong SC (1991) Carbon isotope discrimination by plants follows latitudinal and altitudinal trends. Oecologia 88(1):30–40
Kristensen DK, Kristensen E, Forchhammer MC, Michelsen A, Schmidt NM (2011) Arctic herbivore diet can be inferred from stable carbon and nitrogen isotopes in C3 plants, faeces, and wool. Can J Zool 89(10):892–899
Lambers H, Chapin FS III, Pons TL (2008) Plant physiological ecology. Springer, Berlin
Le Bras-Goude G, Binder D, Formicola V, Duday H, Couture-Veschambre C, Hublin JJ, Richards M (2006) Stratégies de subsistance et analyse culturelle de populations néolithiques de Ligurie: approche par l’étude isotopique (δ13C et δ15N) des restes osseux. B Mem Soc Anthro Par 18(1–2):43–53
Leale Anfossi M (1962) Ritrovamenti archeologici e giacimenti preistorici nelle grotte di Val Pennavaira. Rassegna Speleologica Italiana 14(2):1–7
Longin R (1971) New method of collagen extraction for radiocarbon dating. Nature 230:241–242
Longinelli A, Selmo E (2003) Isotopic composition of precipitation in Italy: a first overall map. J Hydrol 270(1–2):75–88
Macphail RI, Courty MA, Hather J, Wattez J, Ryder M, Cameron N, Branch NP (1997) The soil micromorphological evidence of domestic occupation and stabling activities. In: Maggi R (ed) Arene Candide: a functional and environmental assessment of the Holocene sequence (Excavations Bernabò Brea-Cardini 1940–50). Il Calamo, Roma, pp 53–88
Maggi R (1999) Coasts and uplands in Liguria and northern Tuscany from the Mesolithic to the Bronze Age. In: Tykot RH, Morter J, Robb JE (eds) Social dynamics of the prehistoric central Mediterranean, specialist studies on the Mediterranean, 3. Accordia Research Institute, London, pp 47–65
Maggi R (2004) L’eredità della Preistoria e la costruzione del paesaggio. In: de Marinis RC, Spadea G (eds) I Liguri. Un antico popolo europeo tra Alpi e Mediterraneo. Skira, Milano, pp 35–50
Maggi R, Campana N (2008) Archeologia delle risorse ambientali in Liguria: estrazione e sussistenza fra IV e III millennio BC. Bull Musée D’anthropol Préhistorique Monaco, Suppl 1:65–74
Maggi R, Nisbet R (1990) Prehistoric pastoralism in Liguria. In: Maggi R, Nisbet R, Barker G (eds) Archeologia della pastorizia nell’Europa meridionale. Atti della Tavola Rotonda, Chiavari 1989, Rivista di Studi Liguri 56. Istituto Internazionale di Studi Liguri, Bordighera, pp 265–296
Maggi R, Nisbet R (2000) Alberi da foraggio e scalvatura neolitica: nuovi dati dalle Arene Candide. In: Pessina A, Muscio G (eds.) La Neolitizzazione tra Oriente e Occidente. Edizioni del Museo Friulano di Storia Naturale, Udine, pp 289–308
Maggi R, Nisbet R, Barker G (eds) (1990) Archeologia della pastorizia nell’Europa meridionale. Atti della Tavola Rotonda, Chiavari 1989, Rivista di Studi Liguri 56. Istituto Internazionale di Studi Liguri, Bordighera
Maggi R, Pearce M (in press) L’età del Rame in Liguria. In: Preistoria e Protostoria in Liguria, Atti della LIII Riunione Scientifica, Genova, 16–20 ottobre 2018. Rivista di Scienze Prestoriche
Maggi R, Pearce M (2005) Mid fourth-millennium copper mining in Liguria, north-west Italy: the earliest known copper mines in Western Europe. Antiquity 79(303):66–77
Makarewicz CA (2017a) Sequential δ13C and δ18O analyses of early Holocene bovid tooth enamel: resolving vertical transhumance in Neolithic domesticated sheep and goats. Palaeogeogr Palaeoclimatol Palaeoecol 485:16–29
Makarewicz CA (2017b) Stable isotopes in pastoralist archaeology as indicators of diet, mobility, and animal husbandry practices. In: Ventresca Miller AR, Makarewicz CA (eds) Isotopic investigations of pastoralism in prehistory. Themes in contemporary archaeology. Routledge, London, vol. 4, pp 149–166
Makarewicz CA, Pederzani S (2017) Oxygen (δ18O) and carbon (δ13C) isotopic distinction in sequentially sampled tooth enamel of co-localized wild and domesticated caprines: complications to establishing seasonality and mobility in herbivores. Palaeogeogr Palaeoclimatol Palaeoecol 485:1–15
Makarewicz CA, Arbuckle BS, Öztan A (2017) Vertical transhumance of sheep and goats identified by intra-tooth sequential carbon (δ13C) and oxygen (δ18O) isotopic analyses: evidence from Chalcolithic Köşk Höyük, central Turkey. J Archaeol Sci 86:68–80
Marchi D, Sparacello VS, Holt BM, Formicola V (2006) Biomechanical approach to the reconstruction of activity patterns in Neolithic Western Liguria, Italy. Am J Phys Anthropol 131(4):447–455
Marcott SA, Shakun JD, Clark PU, Mix AC (2013) A reconstruction of regional and global temperature for the past 11,300 years. Science 339(6124):1198–1201
Mashkour M, Bocherens H, Moussa I (2005) Long distance movement of sheep and of Bakhtiari nomads tracked with intra-tooth variations of stable isotopes (13C and 18O). In: Davies J, Fabis M, Mainland I, Richards M, Thomas R (eds) Diet and health in past animal populations. Current Research and Future Directions. Oxbow Books, Oxford, pp 113–122
Mayr C, Grupe G, Toncala A, Lihl CM (2016) Linking oxygen isotopes of animal-bone phosphate with altimetry: results from archaeological finds from a transect in the Alps. In: Grupe G, McGlynn GC (eds) Isotopic landscapes in bioarchaeology. Springer, Berlin, pp 157–172
Milhaud G, Nézit J (1991) Développement des molaires chez le mouton. Etude morphologique, radiographique et microdurométrique. Recl Med Vet 167(2):121–127
Morandi LF, Branch NP (2018) Long-range versus short-range prehistoric pastoralism. Potential of palaeoecological proxies and a new record from western Emilia, northern Apennines, Italy. In: Pelisiak A, Nowak M, Astaloş C (eds) People in the mountains: current approaches to the archaeology of mountainous landscapes. Archaeopress, Oxford, pp 47–59
Müldner G, Britton K, Ervynck A (2014) Inferring animal husbandry strategies in coastal zones through stable isotope analysis: new evidence from the Flemish coastal plain (Belgium, 1st–15th century AD). J Archaeol Sci 41:322–332
Nisbet R (1997) Arene Candide: charcoal remains and prehistoric woodland use. In: Maggi R (ed) Arene Candide: a functional and environmental assessment of the Holocene sequence (Excavations Bernabò Brea-Cardini 1940–50). Il Calamo, Roma, pp 103–112
Orellana-González E, Sparacello VS, Bocaege E, Varalli A, Moggi-Cecchi J, Dori I (2020) Insights on patterns of developmental disturbances from the analysis of linear enamel hypoplasia in a Neolithic sample from Liguria (northwestern Italy). Int J Paleopathol 28:123–136
Owocki K, Kremer B, Cotte M, Bocherens H (2020) Diet preferences and climate inferred from oxygen and carbon isotopes of tooth enamel of Tarbosaurus bataar (Nemegt Formation, Upper Cretaceous, Mongolia). Palaeogeogr Palaeoclimatol Palaeoecol 537. https://doi.org/10.1016/j.palaeo.2019.05.012
Page HM (1995) Variation in the natural abundance of 15N in the halophyte, Salicornia virginica, associated with groundwater subsidies of nitrogen in a southern California salt-marsh. Oecologia 104(2):181–188
Passey BH, Cerling TE (2002) Tooth enamel mineralization in ungulates: implications for recovering a primary isotopic time-series. Geochim Cosmochim Ac 66(18):3225–3234
Passey BH, Robinson TF, Ayliffe LK, Cerling TE, Sponheimer M, Dearing MD, Ehleringer JR (2005) Carbon isotope fractionation between diet, breath CO2, and bioapatite in different mammals. J Archaeol Sci 32(10):1459–1470
Payne S (1973) Kill-off patterns in sheep and goats: the mandibles from Asvan Kale. Anatol Stud 23:281–303
Pederzani S, Britton K (2019) Oxygen isotopes in bioarchaeology: principles and applications, challenges and opportunities. Earth-Sci Rev 188:77–107
Pederzani S, Aldeias V, Dibble HL, Goldberg P, Hublin JJ, Madelaine S, McPherron S, Sandgathe D, Steele TE, Turq A, Britton K (2021) Reconstructing Late Pleistocene paleoclimate at the scale of human behavior: an example from the Neandertal occupation of La Ferrassie (France). Sci Rep 11(1):1–10
Pellegrini M, Donahue RE, Chenery C, Evans J, Lee-Thorp J, Montgomery J, Mussi M (2008) Faunal migration in late-glacial central Italy: implications for human resource exploitation. Rapid Commun Mass Sp 22(11):1714–1726
Pettitt PB, Richards M, Maggi R, Formicola V (2003) The Gravettian burial known as the Prince (“Il Principe”): new evidence for his age and diet. Antiquity 77(295):15–19
Pollard AM, Pellegrini M, Lee-Thorp JA (2011) Some observations on the conversion of dental enamel δ18Op values to δ18Ow to determine human mobility. Am J Phys Anthropol 145(3):499–504
Pryor AJ, Stevens RE, O’Connell TC, Lister JR (2014) Quantification and propagation of errors when converting vertebrate biomineral oxygen isotope data to temperature for palaeoclimate reconstruction. Palaeogeogr Palaeoclimatol Palaeoecol 412:99–107
Pushkina D, Saarinen J, Ziegler R, Bocherens H (2020) Stable isotopic and mesowear reconstructions of paleodiet and habitat of the Middle and Late Pleistocene mammals in south-western Germany. Quat Sci Rev 227. https://doi.org/10.1016/j.quascirev.2019.106026
Pyankov VI, Ziegler H, Akhani H, Deigele C, Lüttge U (2010) European plants with C4 photosynthesis: geographical and taxonomic distribution and relations to climate parameters. Bot J Linn Soc 163(3):283–304
Reimer PJ, Bard E, Bayliss A, Beck JW, Blackwell PG, Bronk Ramsey C, Grootes PM, Guilderson TP, Haflidason H, Hajdas I, Hatté C, Heaton TJ, Hoffmann DL, Hogg AG, Hughen KA, Kaiser KF, Kromer B, Manning SW, Niu M, Reimer RW, Richards DA, Scott EM, Southon JR, Staff RA, Turney CSM, Van Der Plicht J (2013) IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 55(4):1869–1887
Ricci M (1998) Alta Valle Argentina. In: Del Lucchese A, Maggi R (eds): Dal Diaspro al Bronzo, Quaderni della Soprintendenza Archeologica della Liguria 5. Luna Editore, La Spezia, pp 57–62
Robinson C, Kirkham J, Brookes SJ, Bonass WA, Shore RC (1995) The chemistry of enamel development. Int J Dev Biol 39(1):145–152
Rowley-Conwy P (1997) The animal bones from Arene Candide. Final report. In: Maggi R (ed) Arene Candide: a functional and environmental assessment of the Holocene sequence (Excavations Bernabò Brea-Cardini 1940–50). Il Calamo, Roma, pp 153–277
Rowley-Conwy P (2000) Milking caprines, hunting pigs: the Neolithic economy of Arene Candide in its west Mediterranean context. In: Rowley-Conwy P (ed) Animal bones, human societies. Oxbow Books, Oxford, pp 124–132
Rowley-Conwy P, Gourichon L, Helmer D, Vigne J-D (2013) Early domestic animals in Italy, Istria, the Tyrrhenian Islands and southern France. In: Colledge S, Conolly J, Dobney K, Manning K, Shennan S (eds) The origin and spread of domestic animals in Southwest Asia and Europe. Routledge, New York, pp 161–194
Rowley-Conwy P, Panelli C, Rossi S, Nisbet R, Maggi R (2020) First farmers in Liguria, north-western Italy: new evidence from Arene Candide and nearby sites. In: Gron KJ, Sorensen L, Rowley-Conwy P (eds) Farmers at the frontier: a pan European perspective on Neolithisation. Oxbow Books, Oxford, pp 125–144
Rozanski K, Araguás-Araguás L, Gonfiantini R (1993) Isotopic patterns in modern global precipitation. In: Swart PK, Lohmann KC, Mckenzie J, Savin S (eds) Climate Change in Continental Isotopic Records, Geophysical Monograph Series, vol 78. Wiley, Hoboken, pp 1–36
Samartin S, Heiri O, Joos F, Renssen H, Franke J, Brönnimann S, Tinner W (2017) Warm Mediterranean mid-Holocene summers inferred from fossil midge assemblages. Nat Geosci 10(3):207–212
Schoeninger MJ, DeNiro MJ (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim Cosmochim Ac 48(4):625–639
Schoeninger MJ, DeNiro MJ, Tauber H (1983) Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diet. Science 220(4604):1381–1383
Smedley MP, Dawson TE, Comstock JP, Donovan LA, Sherrill DE, Cook CS, Ehleringer JR (1991) Seasonal carbon isotope discrimination in a grassland community. Oecologia 85(3):314–320
Smith BN, Epstein S (1971) Two categories of 13C/12C ratios for higher plants. Plant Physiol 47(3):380–384
Sparacello V, Marchi D (2008) Mobility and subsistence economy: a diachronic comparison between two groups settled in the same geographical area (Liguria, Italy). Am J Phys Anthropol 136(4):485–495
Sparacello VS, Roberts CA, Canci A, Moggi-Cecchi J, Marchi D (2016) Insights on the paleoepidemiology of ancient tuberculosis from the structural analysis of postcranial remains from the Ligurian Neolithic (northwestern Italy). Int J Paleopathol 15:50–64
Sparacello VS, Roberts CA, Kerudin A, Müller R (2017) A 6500-year-old Middle Neolithic child from Pollera Cave (Liguria, Italy) with probable multifocal osteoarticular tuberculosis. Int J Paleopathol 17:67–74
Tauber H (1981) 13C evidence for dietary habits of prehistoric man in Denmark. Nature 292(5821):332–333
Tejedor-Rodríguez C, Moreno-García M, Tornero C, Hoffmann A, García-Martínez de Lagrán Í, Arcusa-Magallón H, Garrido-Pena R, Ignacio Royo-Guillén J, Díaz-Navarro S, Peña-Chocarro L, Alt KW, Rojo-Guerra M (2021) Investigating Neolithic caprine husbandry in the Central Pyrenees: insights from a multi-proxy study at Els Trocs cave (Bisaurri, Spain). PLoS ONE 16(1):e0244139
Tomczyk W, Giersz M, Sołtysiak A, Kamenov G, Krigbaum J (2019) Patterns of camelid management in Wari Empire reconstructed using multiple stable isotope analysis: evidence from Castillo de Huarmey, northern coast of Peru. Archaeol Anthropol Sci 11(4):1307–1324
Tornero C, Aguilera M, Ferrio JP, Arcusa H, Moreno-García M, Garcia-Reig S, Rojo-Guerra M (2018) Vertical sheep mobility along the altitudinal gradient through stable isotope analyses in tooth molar bioapatite, meteoric water and pastures: a reference from the Ebro valley to the Central Pyrenees. Quat Int 484:94–106
Towers J, Jay M, Mainland I, Nehlich O, Montgomery J (2011) A calf for all seasons? The potential of stable isotope analysis to investigate prehistoric husbandry practices. J Archaeol Sci 38(8):1858–1868
Towers J, Gledhill A, Bond J, Montgomery J (2014) An investigation of cattle birth seasonality using δ13C and δ18O profiles within first molar enamel. Archaeometry 56(S1):208–236
Towers J, Mainland I, Montgomery J, Bond J (2017) Calving seasonality at Pool, Orkney during the first millennium AD: an investigation using intra-tooth isotope ratio analysis of cattle molar enamel. Environ Archaeol 22(1):40–55
Trentacoste A, Lightfoot E, Le Roux P, Buckley M, Kansa SW, Esposito C, Gleba M (2020) Heading for the hills? A multi-isotope study of sheep management in first-millennium BC Italy. J Archaeol Sci Rep 29. https://doi.org/10.1016/j.jasrep.2019.102036
Valenzuela-Lamas S, Jiménez-Manchón S, Evans J, López D, Jornet R, Albarella U (2016) Analysis of seasonal mobility of sheep in Iron Age Catalonia (north-eastern Spain) based on strontium and oxygen isotope analysis from tooth enamel: first results. J Archaeol Sci Rep 6:828–836
Van der Merwe NJ (1982) Carbon isotopes, photosynthesis, and archaeology: different pathways of photosynthesis cause characteristic changes in carbon isotope ratios that make possible the study of prehistoric human diets. Am Sci 70(6):596–606
Van Klinken GJ (1999) Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J Archaeol Sci 26(6):687–695
Varalli A, Goude G, Del Lucchese A (2015) Dal Neolitico all’Età del Ferro: indagine paleonutrizionale su alcune popolazioni della Liguria attraverso un approccio isotopico. In: Conventi M, Del Lucchese A, Gardini A (eds) Archeologia in Liguria, Nuova Serie. Sagep, Genova, vol. 5, pp 11–19
Varalli A, Villotte S, Dori I, Sparacello VS (2020) New insights into activity-related functional bone adaptations and alterations in Neolithic Liguria (Northwestern Italy). B Mem Soc Anthro Par 31(1–2):34–58
Ventresca Miller AR, Haruda A, Varfolomeev V, Goryachev A, Makarewicz CA (2020) Close management of sheep in ancient Central Asia: evidence for foddering, transhumance, and extended lambing seasons during the Bronze and Iron Ages. STAR 6(1):41–60
Vigal CR, Machordom A (1985) Tooth eruption and replacement in the Spanish wild goat. Acta Theriol 30(19):305–320
Virginia RA, Delwiche CC (1982) Natural 15N abundance of presumed N2-fixing and non-N2-fixing plants from selected ecosystems. Oecologia 54(3):317–325
Wada E, Kadonaga T, Matsuo S (1975) 15N abundance in nitrogen of naturally occurring substances and global assessment of denitrification from isotopic viewpoint. Geochem J 9(3):139–148
Weinreb MM, Sharav Y (1964) Tooth development in sheep. Am J Vet Res 25:891–908
Werner RA, Brand WA (2001) Referencing strategies and techniques in stable isotope ratio analysis. Rapid Commun Mass Sp 15(7):501–519
Zazzo A, Balasse M, Patterson WP (2005) High-resolution δ13C intratooth profiles in bovine enamel: implications for mineralization pattern and isotopic attenuation. Geochim Cosmochim Ac 69(14):3631–3642
Zazzo A, Balasse M, Patterson WP (2006) The reconstruction of mammal individual history: refining high-resolution isotope record in bovine tooth dentine. J Archaeol Sci 33(8):1177–1187
Zazzo A, Balasse M, Passey BH, Moloney AP, Monahan FJ, Schmidt O (2010) The isotope record of short-and long-term dietary changes in sheep tooth enamel: implications for quantitative reconstruction of paleodiets. Geochim Cosmochim Ac 74(12):3571–3586
Zeder MA, Pilaar SE (2010) Assessing the reliability of criteria used to identify mandibles and mandibular teeth in sheep, Ovis, and goats Capra. J Archaeol Sci 37(2):225–242
Acknowledgements
Dr. Daniele Arobba provided expertise in the regional flora, Prof. Nick Branch supported the funding application, Dr. Paola Calegari provided references on bovid drinking habits. Thanks are also due to Mariavittoria Parravicini and Fabio Fornoni for providing accommodation during the initial stage of the research. The authors are indebted to two anonymous reviewers for their valuable comments on the manuscript.
Funding
Open Access funding enabled and organized by Projekt DEAL. Radiocarbon dating of teeth from Tana del Barletta was supported by the National Radiocarbon Facility of the Natural Environment Research Council and Arts and Humanities Research Council (UK), grant NF/2017/2/15.
Author information
Authors and Affiliations
Contributions
L.F.M. devised the project, produced the main conceptual ideas and interpretations, wrote the paper with inputs from D.F. and created the figures. L.F.M. and D.F. carried out laboratory work on enamel samples. L.F.M. and G.M. carried out laboratory work on collagen samples. D.F. and G.M. helped supervise the project. R.M. directed the excavation of the site, provided expertise in the archaeology of the region and assistance to obtain faunal samples.
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this article was revised:
1) During the proof check, author had updated the affiliation of co-author “D. Frémondeau” to affiliations 2 and 3. However, the published version still has only affiliation 3, while she should have both “University of Reading” and “UCL London”.
2) During the correction of the proof, author had slightly modified a sentence in the section “Archeaeological context”, and this modification does not appear in the published version. Author’s correction to delete “just below 1000 m a.s.l” was not carried out.
The original article has been corrected.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Morandi, L.F., Frémondeau, D., Müldner, G. et al. Sequential analyses of bovid tooth enamel and dentine collagen (δ18O, δ13C, δ15N): new insights into animal husbandry between the Late Neolithic and the Early Bronze Age at Tana del Barletta (Ligurian Prealps). Archaeol Anthropol Sci 13, 147 (2021). https://doi.org/10.1007/s12520-021-01418-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-021-01418-w