
Abstract
The plant and animal components of Linearbandkeramik (LBK) subsistence systems were remarkably uniform with cattle, emmer and einkorn wheat providing the primary source of sustenance for Europe’s earliest agricultural communities. This apparent homogeneity in plant and animal use has been implicitly understood to indicate corresponding similarity in the types of husbandry practices employed by LBK farmers across the entire distribution of the LBK culture. Here, we examine the results from the stable (δ13C/δ15N) isotope analysis of animal bone and cereal grains from the site of Vráble-Veľké Lehemby (Slovakia), providing new information about Linearbandkeramik farming practices in the western Carpathians. Moderately high carbon isotope values from animal bone collagen show that all livestock were pastured in open areas with no evidence of forest pasturing, previously associated with LBK settlements in north-western Europe. High δ15N values measured from domesticated cereal grains suggest manuring took place at the site, while 15N enrichment in bone collagen suggest livestock fed on agricultural by-products and possibly grains. An integrated plant-animal management system was in use at Vráble where livestock grazed on cultivation plots post-harvest. Use of such strategy would have helped fatten animals before the lean winter months while simultaneously fertilising agricultural plots with manure. This study contributes to our growing understanding that although the building blocks of LBK subsistence strategies were remarkably similar, diversity in management strategies existed across central and north-western Europe.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Linearbandkeramik (LBK) is associated with the initial spread of farming and livestock herding across central and north-western Europe during the mid-late sixth millennium cal bc, a process that ultimately incorporated indigenous hunter-gatherers through acculturation or replacement (Gronenborn 2003; Lipson et al. 2017). LBK groups emerged from the Transdanubian linear pottery (TLP) groups in the northern Carpathian region around 5600 cal bc and rapidly expanded west, reaching central and eastern Europe by 5400 cal bc, and its north-western limit in the Paris basin by the mid-sixth millennium BC (Bickle and Whittle 2013; Gronenborn 2003; Pavlů 2005). LBK groups relied primarily on cattle, although pigs, sheep and goats were also exploited at varying intensities (Gillis et al. 2017, 2019; Lüning 2000; Manning et al. 2013b; Marciniak 2013). Cattle herds were largely comprised of adult animals, probably females, and exploited for their milk and meat (Gillis et al. 2017). Dairying was an important subsistence practice, indicated by some post-lactation slaughter of male calves when they were no longer needed for milk let-down. The importance of dairy in LBK societies is further demonstrated by the predominance of dairy fats in pottery, replacing the exploitation of fats from bone marrow and bone grease (Johnson et al. 2018), and the processing of milk into storable products, such as cheese (Salque et al. 2013). Beef was likely consumed as part of feasting events that brought communities together (Marciniak 2005). The focus on bringing most cattle into adulthood suggests considerable investment in their animals by LBK stockherders (Russell 1998), one that required careful planning to ensure large quantities of graze and browse were available as feed year round. LBK communities also farmed a reduced package of Near Eastern plant domesticates, greatly curtailed in taxonomic diversity due to cooler temperatures and wetter conditions in European temperate environments (Ivanova et al. 2018; Kreuz 2007; Kreuz and Schäfer 2011). The cultivation of hulled wheats, such as emmer (Triticum monococcum) and einkorn (Triticum dicoccum), may have been exploited by LBK farmers because the glume protected the grain from freezing and wet growing conditions as well as damp storage conditions (Colledge and Conolly 2007) and therefore better suited for temperate continental climates. Overall, einkorn and emmer wheats were raised on permanent garden plots and were the primary cultivars exploited by LBK groups (Bogaard 2004), while barley (Hordeum vulgare) and legumes (pea (Pisum sativum) and lentil (Lens culinaris)) were rarely farmed (Bogaard 2004; Kreuz 2007; Saqalli et al. 2014). Livestock herds also produce a key ingredient for crop cultivation: manure, which may have been deliberately collected and spread on fields or introduced through animals being penned on cultivation plots to graze on harvest residues. Although limited in number, stable isotopic analyses of carbonised seed remains are revealing that plant management strategies practised by LBK farmers were concerned with the fertilisation of crops in order to ensure production and possibly to increase yields (Fraser et al. 2013; Styring et al. 2017).
The plant and animal building blocks of LBK subsistence systems and the productive outcomes of those systems appear at first glance to have been remarkably homogenous across Europe (Manning et al. 2013b). Although recent studies have highlighted some difference between sexes, with males apparently consuming more protein than females, within the LBK sphere (Bickle and Whittle 2013), ancient DNA analysis has reinforced the traditional image of the LBK as the outcome of monolithic demic and cultural diffusion with little inherent diversity (Bramanti et al. 2009; Lazaridis et al. 2016). The apparent uniformity of LBK subsistence practices is surprising given that LBK groups inhabited assorted habitats ranging from riverine valleys with sparse woodland stands to dense mixed deciduous or alpine forests that bristled with swamps. Furthermore, these landscapes were set across a variety of climatic zones, which follow a general gradient of increased precipitation from east to west and decreased summer temperatures (Bogucki 1988; Kreuz 2007). It is likely that animal husbandry and agricultural practices, including pasturing and fodder provisioning, timing of animal breeding and crop planting, were shaped to local environments across the LBK cultural zone, but, to date, these key practices remain largely unexplored for LBK societies. Stable isotope analysis of animal and plants provides an important tool for characterising animal husbandry and agricultural practices and identifying potential articulation points between livestock herding and plant cultivation (Bogaard et al. 2013; Fraser et al. 2013; Styring et al. 2017). The data produced from stable isotope analysis can help us understand about how these practices were influenced by the size of farming population, their subsistence needs and the resource availability, including arable land as well as animal pasture (Ebersbach 2013) across time and space.
Here, we examine LBK animal pasturing and foddering strategies and their impact on agricultural activities at Vráble-Veľké Lehemby (Slovakia; 5250–4950 cal bc) through stable carbon and nitrogen isotope analyses of animal bone and cereal grain remains. Specifically, we examine the degree to which LBK agriculturalists grazed their animals in open and/or forested environments and whether cultivation and herding practices are detectable in bone collagen carbon and nitrogen isotopes of herbivorous species (Ambrose and Norr 1993; Jim et al. 2004). The results of this study will help build a picture about pasturing and foddering practices and its articulation with agricultural activities in the Western Carpathians, which will further our understanding of human−environment interactions during the LBK.
Background to the study
LBK subsistence practices and environment interactions
Mixed deciduous forests covered much of the central and northern European region during the sixth millennium BC (Kalis et al. 2003; Nielsen et al. 2012). These forested environments varied in density and composition depending on local topography, hydrology and soil conditions, making some areas, such as river valleys and floodplains, more attractive to settlement than thickly forested hills sides (Kalis et al. 2003; Kreuz 2007; Zanon et al. 2018). Openings within the canopy may have been caused by natural processes, such as lightning strikes, browsing by wild herbivores and rooting by wild boar (Vera 2000), and hunter-gatherer groups, who may have practised woodland management to encourage the proliferation of game and nut- and fruit-bearing plants (Caseldine and Hatton 1993; Innes and Blackford 2003; Simmons 1996). The earliest LBK communities were established in forest clearings as indicated by changes in woodland composition and weed seed assemblages recorded in pollen and archaeobotanical archives (Bogaard 2002, 2005; Kalis et al. 2003). Openings within the forests were exploited to establish LBK settlements, small-scale garden plots and animal herds (Bogaard et al. 2013; Kreuz 2007; Saqalli et al. 2014), and, eventually, farmers expanded these areas with selective harvesting of trees for fuel, construction material and tools. Small-scale garden plots and animal husbandry required high labour and resource inputs per unit of land to ensure success, which is a typical mode of production for family groups with a resilient subsistence base (Bogaard 2005; Ebersbach 2013). In addition to domesticated animals and plants, local environments were exploited for hunting and collection of wild foods during the LBK. In general, deer, wild pigs and aurochs were the most commonly exploited wild species with their frequencies in faunal assemblages usually no greater than 10% (Lüning 2000). There are exceptions, for example during early phases of the LBK in Austria and Hungary, and throughout the LBK period in Southern Germany and Alsace, wild fauna can account for as much as 50% (Döhle 1993; Tresset and Vigne 2001). Intensive hunting of wild prey may reflect both LBK settlers opening up and exploiting dense forested environments.
The introduction of domesticated livestock into forested environments would have further shaped their density and composition, and this, in turn, together with open pasture availability, likely influenced the range and characteristics of animal management strategies used by LBK herders. Historically, European forests have been important resources for animal pasture, pannage and fodder (Austad 1988; Read 2003) with the earliest direct botanical remains from trees found in animal dung from Swiss lake village contexts dating to the fifth millennium bc (Rasmussen 1993). To maintain herds and flocks throughout the winter, fodder would have likely been stockpiled throughout the year and collected from several sources including the forest and agricultural by-products (Gregg 1988; Saqalli et al. 2014). Detailed stable isotopic analysis of LBK faunal assemblages from Vaihingen an der Enz and Bischoffsheim, located in southern Germany and eastern France, respectively, has shown that cattle were pastured in forests and/or foddered on tree fodder or ‘leafy hay’ while small stock including pigs were kept within the settlement (Fraser et al. 2013; Gillis et al. submitted). Stable isotope analysis of carbonised cereals from Vaihingen and Viesenhäuser Hof (southern Germany) indicates that crops received a high input of manure (Fraser et al. 2013; Styring et al. 2017), which suggests use of penning strategy where animals are either allowed access to cultivation plots during fallow periods allowing for easy application of manure to fields or carolled facilitating the collection of animal dung. Penning animals on cultivation plots would have simultaneously provided animals with graze from the stubble of crops while ensuring the manuring of plots for the following growing season. Crop stubble or cereal by-products could be used as a source of fodder or post-harvest graze to cover shortages in open pasture due to over-grazing or snow cover (Saqalli et al. 2014).
We would expect LBK farmers to have adapted pasturing/foddering practices in response to local landscapes. The nature and extent of this interaction between LBK farmers and their animals with their surrounding environment would vary across central and north-western Europe given the variety of habitats encompassed by within the cultural zone. However, our present understanding of LBK crop cultivation and animal husbandry practices is one where there is homogeneity in management practices between communities (Kreuz et al. 2005; Manning et al. 2013a). Domesticated species, such as cattle, sheep and pigs, are often considered as a single group when in reality these species have different productive lifecycles and physiologies, which can result in diverse husbandry strategies. Coupled stable isotope analysis of botanical and faunal remains can help us understand the articulation between animal husbandry and plant cultivation. Furthermore, it can help generate detailed information about LBK land use and help us understand adaption of farming practices to forested environments.
The LBK settlement of Vráble and its paleo-environmental context (Fig. 1)

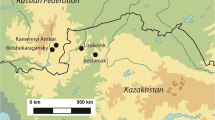
a Location of sites and regions mentioned in the text: north-western Europe: (1) Ensisheim-Ratfeld, (2) Bischoffsheim “AFUA du Stade”, (3) Vaihingen an der Enz, (4) Heilbronn-Neckargartach, (5) Stuttgart-Mühlhausen “Viesenhäuser Hof”; (6) Karsdorf, central Europe: (7) Chotěbudice, (8) Černý Vů, (9) Straubing-Lerchenhaid, (10) Rutzing/Haid, (11) Gnadendorf, (12) Vedrovice-Sídliště, (13) Těšetice-Kyjovice, (14) Aspen an der Zaya/Schletz; (15) Blatné, Carpathian basin: (16) Balatonszárszó Kis-erdei-dűlő (TLP), (17) Füzesabony-Gubakút (ALP), (18) Polgár-Ferenci-hát (ALP) (credit: Gillis, Stammen background map, ALP—Alford linear pottery; TLP—Transdanubian linear pottery). b Map of site with clearly defined neighbourhoods (credit: Müller-Scheeßel)
Vráble is one of the largest LBK settlements so far identified in central Europe, with over 313 house structures in an area of ca. 50 ha (Furholt et al. 2014, 2020; Müller-Scheeßel et al. 2020). It is located in Nitra region and lies on the northern edge of the Danube plain in southwestern Slovakia (Fig. 1). The site is partitioned by a small brook, Kováčovský potok, which flows into Žitava. This is one of several rivers connecting the mineral and stone raw material-rich northern Carpathians with the Danube, an important axis of communication in central Europe. The site consists of three spatially distinct neighbourhoods of similar size (named after their cardinal direction: North (N), South-east (SE) and South-west (SW)), separated from each other by the brook and a ditch-system that encircles the SW neighbourhood of the settlement. Radiocarbon determinations (n = 99) date the site from 5250 to 4950 cal bc, spanning the entire late LBK-Želiezovce phase, and indicate that the three neighbourhoods were more or less contemporaneous with each other (Meadows et al. 2019). The neighbourhoods consisted of the typical LBK-style post-built longhouses, which are grouped into individual yards, which measure an average of 0.5 ha and consist of several, non-contemporary houses and pits of different functions. The most important ones are the so-called lateral long pits, which usually flank both sides of each longhouse. These yards are interpreted as individual households, with a trans-generational transfer of the yard-space. The majority of material culture was obtained from the lateral long pits, which appear to have served as middens, and included pottery and burnt clay, chipped stone and bone artefacts as well as ground stone artefacts, such as millstones, rubbing stones or adzes.
The landscape of the Nitra region at present is characterised by plains, undulations and low foothills of the Western Carpathians and is dissected by the northern Danube tributaries (Fehér 2019: 2–3). These low-lying areas are characterised by thermophilous oak (Quercus pubescens) and maple (Acer tataricum) forests, locally with ash (Fraxinus ornus) or hornbeam (Carpinus) and sporadic patches of grassland. Alluvial zones are covered by elm (Ulmus) woodland and willow-poplar (Salix-Poplar) floodplain forests along the watercourses. Higher elevations and sub-mountain areas are occupied by beech forests, with meadows in forest clearances. Oak forests support rich understory composed of shrubs and herbs, whereas riparian vegetation also includes herbaceous flora, such as sedges (Carex sp.), grasses and floating-leaf plants (Fehér 2019; https://geo.enviroportal.sk/atlassr/). Nowadays, much of the area is deforested, open field landscape, with grain and root crops grown in the plains and vineyards in the foothills (Fehér 2019: 5, 9). Today, the overall climate is predominantly warm and dry with annual precipitation averaging 500–600 mm with mild winters (3 to − 2 °C), especially in the lowlands.
The anthracological archive retrieved from the three Vráble neighbourhoods (Schroedter in press) was composed mostly of the remains of halophilic oak (Quercus) and ash (Fraxinus), pointing to the presence of mixed deciduous forests with a more-or-less open canopy in the vicinity of Vráble. There were traces of light-demanding trees, such as hazel (Corylus), pine (Pinus) and cherry (Prunus), and also taxa that grow in moist areas, including alder (Alnus), poplar (Populus) and elm (Ulmus). The range of represented arboreal taxa at Vráble fits well into the picture of the vegetation of central Europe during the Atlantic period, which has been suggested to be a combination of relatively open mixed oak woodland and riparian vegetation (Magyari et al. 2010; Moskal-del Hoyo 2013). The landscape surrounding Vráble may have also consisted of dry grasslands as indicated by the continuous presence of non-arboreal pollen dating to around 7150 cal bp in the core taken near Santovka, 35 km east of Vráble (Šolcová et al. 2018) as well as by the evidence of persistent non-forest vegetation elsewhere in the Carpathian Basin (Magyari et al. 2010; Pokorný et al. 2015).
Sediment samples were floated from all features for paleobotanical remains. Botanical density (i.e. number of items per litre of soil) was extremely low, even from midden deposits from long pits. The low recovery of plant remains may be due to post-depositional disturbance and/or taphonomy (Filipović et al. in press). However, it is likely, based on their ubiquity in midden deposits, that the inhabitants of Vráble farmed cereal and pulse crops, primarily einkorn (Triticum monococcum) and emmer (T. dicoccum), which is concurrent with previous analysis of botanical remains from LBK sites (Bogaard 2004; Kreuz et al. 2005). Barley (Hordeum vulgare), free-threshing wheat (T. aestivum/durum/turgidum), lentil (Lens culinaris) and pea (Pisum sativum) were also cultivated albeit possibly in smaller quantities. Wild edible nuts and fruit, such as hazelnut (Corylus avellana), wild strawberry (Fragaria vesca) and Chinese lantern (Physalis alkekengi), were also exploited. The weed flora indicate that crop fields were intensively managed, as suggested from previous analysis of botanical remains from LBK sites (Bogaard 2004). The most common arable weeds in the assemblage, fat-hen (Chenopodium album), black bindweed (Fallopia convolvulus) and false cleavers (Galium spurium), indicate high soil disturbance as well as fertile conditions characteristic of intensively managed small permanent arable plots where the cultivation regime would have included tillage (probably by hand), weeding and manuring (Bogaard 2004, 2005). Previous analysis of botanical remains together from Vaihingen an der Enz demonstrated differences between ‘clans’ or neighbourhoods in terms of crop cultivation and to a lesser extent animal husbandry (Bogaard et al. 2016). However, no difference in terms of domesticated crops and weed seed frequencies has been detected between neighbourhoods at Vráble.
A total of 1741 animal bone specimens were recovered from pits associated with 22 separate LBK longhouses located in different areas of the site. Domesticated livestock formed the foundation of the animal-based subsistence economy at Vráble. Pigs were the most commonly identified species (NISP = 180), followed by sheep/goat (NISP = 154) and cattle (NISP = 149) (Table 1). This contrasts with other LBK settlements in Slovakia where cattle overwhelmingly dominate faunal assemblages, followed by sheep/goat and pigs (Bickle and Whittle 2013). Meat production appears to have been the primary focus of stockherders, with all domestic species slaughtered within the first and third year of life (Eckelmann et al. in press). The evidence for dairying is not conclusive, although some cattle survived past 6 years, which may represent slaughter of non-productive females. Overall, the material was poorly preserved and fragmented (Table 1).
Materials and methods
Carbon and nitrogen isotope mechanics
Carbon isotopes
The carbon isotopic compositions of flora ingested by livestock are influenced by photosynthetic pathway, water availability and forest canopy cover (Ambrose and Norr 1993; Jim et al. 2004). C3 plants exhibit a global δ13C mean of − 26.5 ± 2.0‰ (range − 38 to − 22‰; O'Leary 1988; Smith and Epstein 1971; Tieszen 1991; Vogel 1978) while C4 globally average − 12.5 ± 1.0‰ (range − 6 to − 19‰: O'Leary 1988; Smith and Epstein 1971; Tieszen 1991). The western Carpathian region was at the time of the site’s occupation characterised by a range of environments supporting primarily C3 biomes, including pine and mixed broad-leaf forests, while open forest-steppe conditions consisting of scattered woodland stands in areas covered by grasses and forbs may have also contained C4 plants, such as those from the grasses family (Poaceae sp.) (Collins and Jones 1986).
In C3 plants, the isotope fractionation of carbon isotopes largely takes place during the carboxylation process, which is directly affected by light intensity which impacts stomata conductance (Arens et al. 2000; Farquhar et al. 1989b; Van der Merwe and Medina 1991). In European forested temperate environments, canopy density substantially influences the stable carbon isotope composition of understory floral growth (Van der Merwe and Medina 1991; Vogel 1978), with understory plants exhibiting δ13C values of ca. − 31.5‰. This is due to a combination of 13C depletion of atmospheric CO2 under the canopy caused by CO2 respired by decaying organic matter (Tieszen 1991) and low light intensity at the forest floor decreasing photosynthesis efficiency (Farquhar et al. 1989b; Van der Merwe and Medina 1991). The carbon isotope ratios of C3 floral growth increases, between 1 and 6‰, with an opening up of the forest canopy depending on canopy density (Arens et al. 2000). Consequently, animals browsing and grazing under heavy forest canopies will exhibit low carbon isotope values in their skeletal tissues. Contemporary wild herbivores feeding in minimally managed mixed deciduous forest in temperate environments in both Poland and France exhibit bone collagen δ13C values ranging between − 26 and − 23‰ (Drucker et al. 2008). Forest type also appears to impact floral δ13C values. For example, red deer (Cervus elaphus) feeding in mixed deciduous English woodlands exhibit dietary values of − 23.9‰ ± 2.2‰ in comparison to a mean value of − 22.8‰ ± 1.3‰ for animals gazing in a coniferous plantation where undergrowth browse was likely limited and graze on the edge of the plantation was more readily available (Stevens et al. 2006).
The stable carbon isotope composition of plants is also influenced by water availability. During periods of water stress, low humidity and soil moisture, plants reduce stomatal conductance in order to conserve water resulting in lower discrimination against 13C (Arens et al. 2000; Farquhar et al. 1989a; Tieszen 1991). In temperate environments, seasonal variation in soil moisture conditions imparts a 1‰ enrichment in δ13C hair values of herbivores from winter to summer (cattle pastured in Germany, − 26 to − 25‰) (Schnyder et al. 2006). In water-inundated areas, such as swamps and river flood plains as well as riverbanks, the variable hydrological conditions can have different effects on the δ13C values of plant communities (Cloern 2002; Fan et al. 2018; Tieszen 1991). The foliar δ13C values in seasonally inundated areas will exhibit low values in comparison to dry areas. However, in frequently waterlogged areas, values have been observed to increase because of stomatal closure and less discrimination against 13C. During periods of prolonged flooding events, the δ13C values may decrease due to a combination of decreased photosynthetic ability as well as stomatal closure (Fan et al. 2018).
Nitrogen isotopes
Stable nitrogen isotope composition of livestock bones and teeth can provide insights into management strategies, such as weaning (Balasse and Tresset 2002; Gillis et al. 2013) and winter pasturing (Makarewicz 2014, 2015), while δ15N of cereal grains and pulses has been used to investigate prehistoric manuring practices (Bogaard et al. 2013; Fraser et al. 2013; Styring et al. 2014). Nitrogen isotope values in producers and consumers exhibit a c. 3‰ stepwise enrichment between diet and herbivore consumers (Sponheimer et al. 2003b; Vanderklift and Ponsard 2003) but this can vary between 1 and 6‰ in other organisms (DeNiro and Epstein 1981; Minagawa and Wada 1984; Schoeninger and DeNiro 1984; Sealy et al. 1987). This variation is in part due to the studied species trophic position, i.e. carnivore, omnivore or herbivore, as well as related to ecosystem (marine/terrestrial)(Schoeninger and DeNiro 1984), local climate and water availability (Heaton et al. 1986).
Soil nitrogen isotope ratios are influenced by physical and biological processing of organic and inorganic N compounds that take place in soils (Knoepp et al. 2015). Compounds containing 14N are lost through leaching, denitrification and volatilisation resulting in 15N enrichment in remaining organic matter (Knoepp et al. 2015). Environmental factors have the greatest influence on soil δ15values, such as mean annual rainfall and temperature, which are intrinsically linked to geography and topology. European temperate forest soils undisturbed by deforestation or farming activity, such as grazing and tillage, yield low δ15N soil values (− 6 to 0.6‰) reflecting wet and cool conditions where little 14N is lost through leaching and denitrification (Handley et al. 1999; Martinelli et al. 1999).
Nitrogen is fixed in plants from the atmosphere directly or indirectly. N2 fixing plants, such as legumes, convert atmospheric N2 to ammonia and in general exhibit δ15N values of around 0‰ reflecting atmospheric N2 (Ambrose and DeNiro 1989). In contrast, non-fixing plants, which include cereals and most tree species, rely solely on sources of fixed N derived from decomposed plant and animal matter, and activity of nitrogen-fixing microorganisms (Handley et al. 1999). Within these plants, δ15N values vary widely between and within species, reflecting physiological differences that impact the degree of N assimilation by plant roots or shoots and subsequent translocation and reduction processes, local differences in soil N and abiotic factors including aridity and salinity (Amundson et al. 2003; Liu et al. 2014). The latter factors can lead to 15N-enrichment due to increased denitrification favouring the loss of 14N (Ambrose and DeNiro 1989; Handley et al. 1999). The incorporation of manure into the soil N cycle can led to an increase in plant δ15N values by 4 to 9‰. This is because manure is enriched with 15N due to the loss of 14N by the animal during digestion and excretion (Robbins et al. 2005). Therefore, the nitrogen isotopes of cultivated plant remains can indicate quality of agricultural soils and manuring practices (Bogaard et al. 2013; Fiorentino et al. 2014; Fraser et al. 2011). There is a clear link between manure and high δ15N values in cereals, with the magnitude of this effect relative to the quantity of manure applied to fields (Bogaard et al. 2007; Fraser et al. 2011). For example, previous experimental work has demonstrated that crops grown without manure yield low values averaging 2.5‰ compared to crops subjected to under long-term manuring regimes receiving c. 35 to 37 t per hectare (t/ha) per annum of dung, yielding nitrogen isotope ratios averaging 6‰. Crops grown under low levels of manuring regimes (20 to 30 t/ha) produced δ15N values of between 2.5 and 6‰ (Fraser et al. 2011).
Sample selection and preparation for stable carbon and nitrogen isotope analysis
Faunal remains
To investigate animal dietary intake at Vráble, bone samples were taken from 45 domesticated animal bones (Bos taurus, n = 20; Ovis aries/Capra hircus, n = 13; Sus sp., n = 12 (SDATA 1)) from each of the three settlement areas. Only specimens exhibiting fused epiphyseal ends typical for older juvenile and mature animals were analysed in order to ensure suckling did not impact isotope values. Suckling can increase stable nitrogen isotope values and, to a lesser extent, stable carbon isotope values due to infants consuming a protein-rich diet via the mother’s milk, resulting in a trophic level positioned one step higher than that of the mother (Balasse and Tresset 2002; Gillis et al. 2013).
Bone collagen was extracted from the bone samples following Tuross et al. (1988) using EDTA (0.5 M; pH 7.5) solution. Small bone samples were soaked in 10 ml EDTA until complete decalcification, then the samples were rinsed in ultra-pure distilled water several times then rinsed in 0.1 M NaOH. Extracted collagen was left to soak overnight at 10 °C in distilled water and rinsed again five times in ultra-pure distilled water and then freeze dried. Bone collagen samples (800–1200 μg) were analysed for carbon and nitrogen isotopes on GVI IsoPrime and a Eurovector elemental analyser at Boston University. For 13CV-PDB, the CO2 gas was calibrated against NBS 20 (Solnhofen Limestone), NBS 21 (Spectrographic Graphite) and NBS 22 (Hydrocarbon Oil), and for 15Nair, the gas was calibrated against atmospheric N2 and IAEA standards N-1, N-2 and N-3 (all are ammonium sulphate standards). Two internal lab standards were used: peptone and glycine, which gave a mean analytical precision estimate (based on 3 peptone and 5 glycine standards) for 15N analyses, 0.08‰, and for 13C analyses, 0.02‰. Cereal grain samples were analysed at the Service de Spectrométrie de Masse isotopique du MNHN (SSMIM, Paris) using an elemental analyser (Flash 2000) interfaced with a mass spectrometer (DeltaV Advantage). The mean analytical precision estimated based on 13 Alanine standards for δ15N analyses was 0.06‰ and for δ13C analyses was 0.07‰.
The offset between diet and animal/human δ13C values depends on the tissue under study such as bone collagen, bioapatite (Jim et al. 2004; Sponheimer et al. 2003a), digestion physiology, the ratio between C4 and C3 plants in the diet (Codron et al. 2011), and the ratio of marine versus terrestrial protein in the diet (Schoeninger and DeNiro 1984). The enrichment between diet δ13C values and herbivore bone δ13C values collagen has been proposed to between 5.1 and 5.3‰ (Ambrose and Norr 1993; Jim et al. 2004; Sponheimer et al. 2003a). We will use 5.1‰ to be comparable to previous stable isotope studies of LBK faunal material (Berthon et al. 2018). The trophic enrichment factor between diet δ15N and animal δ15N values is 3‰ following (Schoeninger and DeNiro 1984; Sponheimer et al. 2003b; Vanderklift and Ponsard 2003).
Cereals
Cereal grains of emmer (T. dicoccum) and einkorn (T. monococcum) wheat selected for stable carbon and nitrogen isotope analysis were recovered from five houses: Houses 102 and 131 in the SE neighbourhood, houses 258 and 262 in the N neighbourhood and house 39 from the SW neighbourhood (SDATA 2). Grain δ15N values vary up to 3‰ within a single ear (Nitsch et al. 2015). Therefore, five or more grains for each species were selected from the houses and aggregated into a single bulk sample for each house. This was to reduce the potential over- or under-representation by single grains (Nitsch et al. 2015).
Pre-treatment of carbonised seeds using an acid−base−acid sequence or acid alone at applied at a variety of temperatures do not significantly impact stable isotope results, although acid–base–acid protocols lead to considerable sample loss (Brinkkemper et al. 2018; Vaiglova et al. 2014b). We chose a gentle acid protocol to remove exogenous carbonates as sampled grains exhibited no observable sediment contaminates under a microscope. Bulk samples were prepared by first soaking carbonised seeds in aqueous 0.5 M HCl for 30 min or until effervescence ceased at room temperature. Samples were then rinsed three times in ultra-pure distilled water and homogenised using a pestle.
Carbonisation causes loss and breakdown of proteins, lipids and carbohydrates (cellulose and starch) and shift in vivo δ15N and δ13C values in carbonised seeds. Charring experiments have proposed several offset values for δ15N values from 1.1‰ (± 0.4) (Fraser et al. 2011) to 0.3‰ (± 0.5) (Nitsch et al. 2015). This discrepancy is due to the variation in temperature and charring time applied to seeds in each of these experiments. We will use the most recent calculated offset value of 0.3‰ as it considers different possible variations in temperature and time and its impact on δ15N values. The offset between grain δ15N values and rachis values is 2.4‰ (± 0.8) (Fraser et al. 2011). An offset value of 0.1‰ (± 0.1) will be used for δ13C values (Nitsch et al. 2015).
Statistical analyses were carried out using PAST (Hammer et al. 2001), and where the total sample sizes were less than 30, we used non-parametric tests (Dytham 2003).
Results
Faunal isotopic results (Fig. 2)
Collagen yields from bone specimens ranged between 12.0 and 2.0% (Table 2). All samples with the exception of one sample (7154) yielded well-preserved collagen (20 to 0.5%), with CN ratio (3.1 to 3.5), %C (26 to 40%) and %N (11 to 16%) that fall within the ranges established by van Klinken (1999) for well-preserved collagen.

Stable isotope results for Vráble: a Mean values for Bos with outlier 7158 removed, Ovis/Capra and Sus sp.; Bos results for each settlement with outlier 7158 removed; b Ovis/Capra results for each settlement; c Sus sp. results for each settlement
The δ13C values for all sampled animals range from − 21.8 to − 16.0‰. Bos sp. exhibit an average δ13C of − 20.1 ± 0.4‰ (range, − 20.7 to − 18.9‰, n = 18) while the average for Ovis/Capra was − 20.3 ± 0.5‰ (range, − 21.3 to −19.3‰, n = 13) and for Sus sp. − 20.7 ± 0.5‰ (range, − 21.6 to − 19.8‰, n = 12). Bos was enriched on average − 0.6‰ in δ13C relative to Sus (Mann−Whitney, p = 0.002). There was no significant difference in the isotopic composition of livestock species between different neighbourhoods, probably due to the small sample size. Welsh’s t-test returned non-significant results suggesting that the variance between datasets to be the same. Altogether, these results suggest that all livestock from Vráble consumed a very similar diet composed of plants with a range of δ13C values of − 26.9 to − 21.1‰, using a dietary enrichment factor of 5.1‰ (Ambrose and DeNiro 1986).
Livestock δ15N values ranged from 6.8 to 11.7‰. Ovis/Capra samples exhibited an average value of 9.5 ± 1‰ (range, 8.1 to 11.7‰; Fig. 2c), while Sus sp. had a comparable average but smaller range in values (9.3 ± 0.5‰, range, 8.8 to 9.9‰; Fig. 2d). Although not significantly different from other species, cattle δ15N values (8.9 ± 1.2‰, range, 7.0 to 11.3‰; Fig. 2b) were on average 0.4 to 0.6‰ lower than those from Ovis/Capra and Sus sp., respectively. Assuming a fractionation factor of 3‰ between diet and bone collagen, stable nitrogen isotope values for livestock diets ranged from 3.8 to 8.7‰.
Cereal stable isotope results
The C to N ratios (24.1 to 18.2) and %C (59.7 to 52.6) and %N (3.6 to 2.8) values were consistent with previous reported values from carbonised archaeological cereal grains (Fraser et al. 2013; Vaiglova et al. 2014a) (Table 3). For the emmer samples, δ13C values averaged − 23.5 ± 0.5‰, and for the einkorn samples, − 23.6 ± 0.3‰. There is little carbon isotopic difference observed in charred seed remains recovered from different neighbourhoods nor are there isotopic differences between crop species.
The average δ15N values overall, adjusted for the charring effect, for emmer samples was 6.6 ± 1‰ and for einkorn samples was 6.9 ± 1‰ (Table 3). Einkorn grains from the northern neighbourhood have the highest δ15N values (7.9 ± 0.1‰ (n = 2)), which were between 1.5 and 2.1‰ higher than those from the SE and SW neighbourhoods. While emmer grains from the south-eastern neighbourhood, on the other hand, had the highest δ15N values (7 ± 1.4‰ (n = 2)). These are between 0.3 and 1.3‰ higher than those from the N and SW neighbourhoods. The estimated δ15N average for chaff for emmer, using the 2.4‰ offset, was on average 4.3 ± 1.0‰ and for einkorn was 4.6 ± 0.9‰.
Discussion
Vráble livestock foddering/pasturing from the perspective of carbon isotopes
The striking lack of difference in the carbon isotope ratios within and between sheep/goat, pigs and cattle at Vráble suggests all livestock ingested broadly plants growing in similar environments. This strongly suggests a highly generalised husbandry system was employed that did not apply species-specific pasturing or foddering regimes. Species-specific foddering regimes may have been practiced by LBK farmers in north-western Europe, with cattle pastured in forests while small stock remained within the settlement. This strategy was possibly developed to provide for large herds of cattle and/or the lack of available open pasture. In this area, there is strong stable isotopic evidence for forest pasturing and leafy hay based on cattle bone collagen δ13C values lower than − 22‰ (Gillis et al. submitted), based on a study of Scandinavian Aurochs browsing with forested environments (Noe-Nygaard et al. 2005). Using this baseline, Vráble livestock δ13C values (− 21.8 to − 16.0 ‰) do not appear to support closed canopy forest pasturing at the site. It is possible that livestock from Vráble were pastured within an open forested landscape or foddered with fodder resources from open forests. However, further compound-specific analysis of amino acids, which can directly demonstrate woody plant consumption (Kendall et al. 2018, 2019), is needed to confirm leafy hay foddering. Thus, we cannot confirm or rule out the use of forests as pasture or their resources for foddering at Vráble.
Comparison of stable carbon isotope results from Vráble with a dataset containing 17 roughly contemporaneous sites from across the LBK cultural zone (Fig. 1; SDATA 3) reveals little diversity in small stock diet in terms of δ13C values across these different regions. For example, sheep/goat from Vráble exhibit similar δ13C values with those reported from proto-LBK in the Carpathian Basin (TLP and Alföld linear pottery (ALP), − 19.9 ± 0.4‰, N = 3) and LBK settlements in other regions of central Europe (− 20.2 ± 0.6‰, N = 8) and north-western Europe (− 20.6 ± 0.4‰, N = 5) (Fraser et al. 2013; Gillis et al. submitted; Hedges et al. 2013; Oelze et al. 2011). This is also seen to a lesser extent in pigs (Carpathian Basin, − 20.3 ± 0.5‰; central Europe, − 20.5 ± 0.3‰), except Vráble pigs had values an average 0.4‰ higher than those from north-western Europe (− 21.1 ± 0.5‰; ANOVA p < 0.005; Vráble ~ north-western Europe: Tukey pairwise, p = 0.01). This may be a reflection of forests being used for pannage in this region.
Statistical analysis shows Vráble cattle δ13C values were significantly different than cattle from central and north-western Europe (ANOVA p < 0.005; Tukey test: central Europe ~ Vráble, p = 0.001; north-western Europe ~ Vráble, p < 0.005) whereas there was no difference when compared with proto-LBK (ALP/TLP) sites south of Vráble in the Carpathian Basin. This suggests Vráble cattle and those from the Carpathian Basin (average, − 19.9; range, − 21.6 to − 17.0‰) were pastured and foddered on plants within similar open environments. There is a clear 2.3‰ decrease in δ13C bone collagen values visible in LBK cattle from the riverine environments of Carpathian Basin to the more densely forested environments of eastern France and southern Germany. We propose that this clear environmental imprint is a direct reflection of the cattle pastures and resources used for fodder. Here, and in a large-scale regional analysis, the stable isotopic evidence suggests that there are variations in terms of cattle pasturing/foddering practices across the LBK cultural zone (Gillis et al. submitted), although it remains to be seen whether this is the result of local adaptive response to environment and availability of pasture or diversity within LBK cultural subsistence practices.
Evidence for manuring: Perspectives from the cereals
Cereal grains from Vráble exhibited high δ15N values suggesting fields were manured. In comparison to previous stable isotopic studies, cereal grains from Vaihingen (Fraser et al. 2013) were c. 2.3‰ (einkorn) and 2.4‰ (emmer) lower in δ15N than those from Vráble while those from at Viesenhäuser Hof (Styring et al. 2017) were 1 to 0.2‰ lower. Grains from long-term experiments (> 100 years) where manured plots received c. 20 t/ha of animal manure biennially, yielded δ15N values ranging between 2.7 and 5.7‰, whereas cereals under long-term or intensive manuring regimes yielded δ15N values greater than 6‰ (Bogaard et al. 2013). Consequently, at Vaihingen, cereal δ15N values were interpreted to be the result of low level manuring of about 10–15 t cattle manure per hectare using a conservative estimate of 5 t per animal (Fraser et al. 2013). In the case of Viesenhäuser Hof, it was proposed that cultivation plots were under a more intensive manuring regime, for example, 35 t/ha per annum (Styring et al. 2017). The considerable 15N enrichment in Vráble cereals suggests cultivation plots received high levels of manure. This may have been the result of manure being collected from pastures or by penning animals directly on cultivation plots. Penning animals on agricultural plots to feed upon crop stubble or by-products would have led to animal dung being contained within an area and reduced the need for collection. Small-scale garden plots requires high labour so penning animals on cultivation areas would have potentially reduced the labour output needed for collecting manure. Combined archaeobotanical and stable isotopic analysis at Vaihingen has indicated that some clans/households had access to manured fields (Bogaard et al. 2015) perhaps fertilised by animals kept within the settlement. While this differentiation between neighbourhoods is not evident at Vráble, future excavation and stable isotopic analysis of cereals may elucidate differences and similarities in animal-crop husbandry systems between communities.
Articulation between herding and cultivation practices
Livestock δ15N values ranged from 6.8 to 11.7 ‰ with ruminants exhibiting the highest values. When these results are examined within the regional context, Vráble cattle δ15N values were found to be statistically different than those from sites in other regions (ANOVA, p < 0.05). Furthermore, this is also seen for sheep/goat (ANOVA, p < 0.05) and pigs (ANOVA, p < 0.05). All livestock had higher δ15N values than those from other regions (Tukey tests between all regions and Vráble, p < 0.0001). High δ15N values in pigs reflect in part their omnivore diet, and access to food sources enriched with 15N, such as protein-rich food and human waste (Balasse et al. 2016). However, high δ15N values in herbivores are a direct reflection of the nitrogen isotopic composition of ingested plants. Soil and therefore plants can become enriched in 15N as a result of areas being used for long-term pasture (Makarewicz 2014, 2015). Moderately high δ15N values shared by all Vráble livestock species indicate animals consistently ingested 15N enriched plants within pastures. This may be the result of animals grazing long-term pastures but also may be caused by livestock feeding on crop stubble or by-products grown within manured fields. The δ15N values for animal fodder were estimated using the offset between diet and herbivores of 3‰. These were compared with the species-specific δ15N values from cereal grains and estimated for chaff (using an offset of 2.4‰ (Fiorentino et al. 2014)) (Fig. 3). The fodder estimates for individual species were not significantly different from the δ15N values from cereal grains (Kruskal–Wallis, p = 0.35). However, there was a significant difference between sheep/goat and pig fodder and estimated δ15N values for chaff (Kruskal–Wallis p < 0.001). Cattle fodder estimates were not significantly different than values estimated for emmer and einkorn chaff. These results suggest that spent grain or logged cereals were used as animal fodder. A previous analysis at Vaihingen indicated that the animals were mainly fed on wild browse and graze (Fraser et al. 2013) while at Viesenhäuser Hof, animals were fed on cereal chaff/spent grain (Styring et al. 2017). In these studies, the offset between diet and bone collagen was 4‰ in comparison to 3‰ used in this study. If we use the 4‰ offset, there is no difference between the chaff and animal fodder δ15N estimated values (Kruskal–Wallis, p = 0.1). There was a significant difference between δ15N values for cereals and animal fodder (Kruskal–Wallis, p = 0.002), with pig fodder estimates significantly different from δ15N values for emmer and einkorn grains (Mann–Whitney: ~ emmer, p = 0.05; ~ einkorn, p = 0.05). Further analysis is needed to determine which offset is the most accurate to use. However, what is clear is that cereal crops and possible by-products were used as animal fodder at the site.

Estimated δ15N values for Bos, Ovis/Capra and Sus sp. fodder (using 3‰ offset), δ15N results for carbonised emmer and einkorn cereal grains and estimated emmer and einkorn chaff values, using offset 2.4‰ (Fiorentino et al. 2014). δ15N values have been adjusted for charring (minus 0.3‰ (Fraser et al. 2011))
The intensive manuring regimes interpreted from cereal grain δ15N values suggest that Vráble livestock were penned on cultivation plots. The close articulation between crop cultivation and animal husbandry potentially to reduce labour demands is associated with manure collection as well as facilitating collection of other animal products, such as milk. Intensive articulated cultivation-herding strategy may be an indication that the availability of arable land for cultivation was low (Bogaard 2005; Ebersbach 2013), possibly due to geology, such as heavy soils, or differential access for individual clans or neighbourhoods as seen previously at Vaihingen (Bogaard et al. 2016). Over-specialisation may also have exposed LBK farmers to crop failure due to un-seasonable weather during the crop cycle. Small herds may not have provided populations within the three neighbourhoods with sufficient resources to cope with subsistence crisis caused by crop failure, potentially leading to inequality and strife between communities (Furholt et al. 2020). Trauma pathologies found within populations in later phases of the LBK has been proposed to be evidence of violent conflict over resources (Golitko and Keeley 2006), and diminished food supplies may have contributed to the eventual ‘collapse’ of LBK societies (Shennan et al. 2013).
Conclusions
The use of stable carbon and nitrogen isotope analysis in the investigation of prehistoric animal husbandry and agricultural practices is broadening our understanding of Neolithic subsistence strategies and how these evolved and adapted to different ecological niches and by cultural groups. The analysis of the Vráble faunal and cereal remains makes a significant contribution to enhancing our understanding of LBK crop cultivation and animal husbandry practices. The clear indication that agricultural plots at Vráble were likely supplied with manure demonstrates an articulation between plant cultivation and animal husbandry practices. It appears that all livestock species were pastured in open forest/open areas near the settlement, with cultivation plots used as pasture reducing labour needs for manure collection. The data presented here highlight the importance of stable isotope analysis of cereal grains to gain a nuanced picture of their growing conditions and to provide another dimension to archaeobotanical investigations. Our stable isotopic analysis of domesticated fauna was based on bone collagen, and the attenuation of stable isotope ratios in adult bone means that seasonal variation in foddering and pasture practices is lost. Further incremental analysis of livestock dentine and bioapatite is required to investigate whether plants with high δ15N values was a seasonal fodder suggesting a fodder source connected to crop cultivation or whether animals grazed on plants with high δ15N values all year round thus from long-term pasture use. In addition, detail analysis of livestock teeth from individual neighbourhoods would highlight similarities and variations between communities in terms of seasonality of husbandry practices.
The study contributes to the growing evidence for the existence of diversity as well as flexibility within LBK crop cultivation and stock herding practices. It is clear that Neolithic herding practices have been adapted since their introduction from the Near East into Europe; however, what is not clear is whether this was the result of adaptive response by farmers to local environments or cultural subsistence choices, or a combination of both. Future holistic stable isotopic analysis of plants and faunal from a single culture, such as the LBK, can help determine the important factors underlining this adaptive processes as well as build a more comprehensive picture of early Neolithic subsistence practices. This can be investigated by cross-correlation between detailed datasets of cultural attributes, such as frequencies of lithic technology and pottery styles, with environmental proxies and stable isotopic results from multiple sources including bone collagen, bioapatite and organic residues from ceramics. Together with a detailed chronological framework, this will generate the deep data sets required to trace the evolution of crop and animal husbandry systems, land use and subsistence practices over time and space, increasing our understanding of role played by domesticated plants and animals in shaping Neolithic societies.
Change history
31 October 2020
Springer Nature’s version of this paper was updated to present the missing electronic supplementary materials.
References
Ambrose SH, DeNiro MJ (1986) Reconstruction of African diet using bone collagen carbon and nitrogen isotopes ratios. Nature 319:321–324
Ambrose SH, DeNiro MJ (1989) Climate and habitat reconstruction using stable carbon and nitrogen isotope ratios of collagen in prehistoric herbivore teeth from Kenya. Quat Res 31:407–422
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of carbon isotope ratios of whole diet and dietary protien to those of bone collagen and carbonate. In: Lamber JB, Grupe G (eds) Prehistoric human bone: archaeology at the molecular level. Springer-Verlag, Berlin
Amundson R et al (2003) Global patterns of the isotopic composition of soil and plant nitrogen. Glob Biogeochem Cycles 17:31–10
Arens NC, Jahren AH, Amundson R (2000) Can C3 plants faithfully record the carbon isotope composition of atmospheric carbon dioxide? Paleobiology 26:137–164
Austad I (1988) Tree pollarding in western Norway. In: Birks HH, Birks HJB, Emil Kaland P, Moe D (eds) The cultural landscape- past, Present and Future. Cambridge University Press, Cambridge, pp 11–30
Balasse M, Tresset A (2002) Early weaning of Neolithic domestic cattle (Bercy, France) revealed by intra-tooth variation in nitrogen isotope ratios. J Archaeol Sci 29:853–859. https://doi.org/10.1006/jasc.2001.0725
Balasse M et al (2016) Wild, domestic and feral? Investigating the status of suids in the Romanian Gumelniţa (5th mil. cal BC) with biogeochemistry and geometric morphometrics. J Anthropol Archaeol 42:27–36. https://doi.org/10.1016/j.jaa.2016.02.002
Berthon R, Kovačiková L, Tresset A, Balasse M (2018) Integration of Linearbandkeramik cattle husbandry in the forested landscape of the mid-Holocene climate optimum: seasonal-scale investigations in Bohemia. J Anthropol Archaeol 51:16–27. https://doi.org/10.1016/j.jaa.2018.05.002
Bickle P, Whittle AWR (2013) LBK lifeways: a search for differences. In: Whittle AWR, Bickle P (eds) The first farmers of central Europe: diversity in LBK lifeways. vol Cardiff studies in Archaeology. Oxbow Books, Oxford, pp 1–28
Bogaard A (2002) Questioning the relevance of shifting cultivation to neolithic farming in the Loess belt of Europe. Veg Hist Archaeobotany 11:155–168. https://doi.org/10.1007/s003340200017
Bogaard A (2004) Neolithic farming in Central Europe. An archaeobotanical study of crop husbandry practices. Routledge, London
Bogaard A (2005) ‘Garden agriculture’ and the nature of early farming in Europe and the Near East. World Archaeol 37:177–196. https://doi.org/10.1080/00438240500094572
Bogaard A, Heaton THE, Poulton P, Merbach I (2007) The impact of manuring on nitrogen isotope ratios in cereals: archaeological implications for reconstruction of diet and crop management practices. J Archaeol Sci 34:335–343. https://doi.org/10.1016/j.jas.2006.04.009
Bogaard A et al (2013) Crop manuring and intensive land management by Europe's first farmers. Proc Natl Acad Sci U S A 110:12589–12594. https://doi.org/10.1073/pnas.1305918110
Bogaard A, Krause R, Strien H-C (2015) Towards a social geography of cultivation and plant use in an early farming community: Vaihingen an der Enz, south-west Germany. Antiquity 85:395–416. https://doi.org/10.1017/s0003598x00067831
Bogaard A et al (2016) The Bandkeramik settlement of Vaihingen an der Enz, Kreis Ludwigsburg (Baden-Württemberg): an integrated perspective on land use, economy and diet. Germania 94:1–60. https://doi.org/10.11588/ger.2016.39068
Bogucki P (1988) Forest farmers and stockholders. Early agriculture and its consequences in North-Central Europe. Cambridge Press, Cambridge
Bramanti B et al (2009) Genetic discontinuity between local hunter-gatherers and Europe’s first farmers. Science 326(5949):137–140. https://doi.org/10.1126/science.1176869
Brinkkemper O, Braadbaart F, van Os B, van Hoesel A, van Brussel AAN, Fernandes R (2018) Effectiveness of different pre-treatments in recovering pre-burial isotopic ratios of charred plants. Rapid Commun Mass Spectrom 32:251–261. https://doi.org/10.1002/rcm.8033
Caseldine C, Hatton J (1993) The development of high moorland on Dartmoor: fire and the influence of Mesolithic activity on vegetation change. In: Chambers FM (ed) Climate change and human impact on the landscape: studies in palaeoecology and environmental archaeology. Chapman and Hall, London, pp 119–131
Cloern JE (2002) Stable carbon and nitrogen isotope composition of aquatic and terrestrial plants of the San Francisco Bay estuarine system. Limnol Oceanogr 47:713–729. https://doi.org/10.4319/lo.2002.47.3.0713
Codron D, Codron J, Sponheimer M, Bernasconi SM, Clauss M (2011) When animals are not quite what they eat: diet digestibility influences 13C-incorporation rates and apparent discrimination in a mixed-feeding herbivore Canadian. J Zool 89:453–465. https://doi.org/10.1139/z11-010
Colledge S, Conolly J (2007) The origins and spread of domestic plants in Southwest Asia and Europe. Left Coast Press, Walnut Creek
Collins RP, Jones MB (1986) The influence of climatic factors on the distribution of C4 species in Europe. Vegetatio 64:121–129
DeNiro MJ, Epstein S (1981) Influence of diet on the distribution of nitrogen isotopes in animals. Geochim Cosmochim Acta 45:341–351
Döhle J (1993) Haustierhaltung und Jagd in der Linienbandkeramik-ein. ZfA 27:105–124
Drucker DG, Bridault A, Hobson KA, Szuma E, Bocherens H (2008) Can carbon-13 in large herbivores reflect the canopy effect in temperate and boreal ecosystems? Evidence from modern and ancient ungulates. Palaeogeogr Palaeoclimatol Palaeoecol 266:69–82. https://doi.org/10.1016/j.palaeo.2008.03.020
Dytham C (2003) Choosing and using statistics: a biologists guide, 2nd edn. Blackwell Science, Oxford.
Ebersbach R (2013) Quantitative approaches to reconstructing prehistoric stock breeding. In: Kerig T, Zimmermann A (eds) Economic archaeology: from structure to performance in European archaeology. Habelt, Bonn, pp 143–160
Eckelmann R, Schmölcke U, Makarewicz CA (in press) Chapter 5.4 The animal remains from the LBK and Želiezovce settlement site of Vráble. In: Furholt M, Cheben I, Müller J, Bistáková A, Wunderlich M & Müller-Scheeßel N (eds) Archaeology in the Zitava Valley 1 - the LBK settlement site of Vráble. SideStone Press, Leiden
Fan R, Morozumi T, Maximov TC, Sugimoto A (2018) Effect of floods on the δ13C values in plant leaves: a study of willows in Northeastern Siberia. PeerJ 6:e5374. https://doi.org/10.7717/peerj.5374
Farquhar GD, Ehleringer J, Hubick KT (1989) Carbon isotope discrimination and photosynthesis. Annu Rev Plant Biol 40:503–537
Fehér A (2018) Vegetation history and cultural landscapes case studies from South-west Slovakia. Springer Geography, Cham
Filipović D, Kroll H, Kirleis W (in press) Chapter 5.5 Archaeobotanical remains from the LBK and Želiezovce settlement site of Vráble. In: Furholt M, Cheben I, Müller J, Bistáková A, Wunderlich M & Müller-Scheeßel N (eds) Archaeology in the Zitava Valley 1 - the LBK settlement site of Vráble. SideStone Press, Leiden
Fiorentino G, Ferrio JP, Bogaard A, Araus JL, Riehl S (2014) Stable isotopes in archaeobotanical research. Veg Hist Archaeobotany 24:215–227. https://doi.org/10.1007/s00334-014-0492-9
Fraser RA, Bogaard A, Heaton T, Charles M, Jones G, Christensen BT, Halstead P, Merbach I, Poulton PR, Sparkes D, Styring AK (2011) Manuring and stable nitrogen isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary practices. J Archaeol Sci 38:2790–2804
Fraser RA, Bogaard A, Schäfer M, Arbogast R, Heaton THE (2013) Integrating botanical, faunal and human stable carbon and nitrogen isotope values to reconstruct land use and palaeodiet at LBK Vaihingen an der Enz, Baden-Württemberg. World Archaeol 45:492–517. https://doi.org/10.1080/00438243.2013.820649
Furholt M, Bátora J, Cheben I, Kroll H, Tóth P (2014) Vráble-Velké Lehemby: Eine Siedlungsgruppe der Linearkeramik in der Südwestslowakei. Vorbericht über die Untersuchungen der Jahre 2010 und 2012 und Deutungsansätze. SlovArch 62:227–266
Furholt M, Müller-Scheeßel N, Wunderlich M, Cheben I, Müller J (2020) Communality and discord in an early Neolithic settlement agglomeration: the LBK site of Vráble, Southwest Slovakia. Camb Archaeol J 30:469–489. https://doi.org/10.1017/S0959774320000049
Gillis R, Bréhard S, Bălăşescu A, Ughetto-Monfrin J, Popovici D, Vigne J-D, Balasse M (2013) Sophisticated cattle dairy husbandry at Borduşani-Popină (Romania, fifth millennium BC): the evidence from complementary analysis of mortality profiles and stable isotopes. World Archaeol 45:447–472. https://doi.org/10.1080/00438243.2013.820652
Gillis RE, Kovačiková L, Bréhard S, Guthmann E, Vostrovská I, Nohálová H, Arbogast RM, Domboróczki L, Pechtl J, Anders A, Marciniak A, Tresset A, Vigne JD (2017) The evolution of dual meat and milk cattle husbandry in Linearbandkeramik societies. Proc R Soc Lond B Biol Sci 284. https://doi.org/10.1098/rspb.2017.0905
Gillis RE, Gaastra JS, Vander Linden M, Vigne JD (2019) A species specific investigation into sheep and goat husbandry during the early European Neolithic. Environ Archaeol. https://doi.org/10.1080/14614103.2019.1615214
Gillis RE, Kendall IK, Balasse M, Evershed RP (in press) Conduite de l’élevage dans un environnement forestier à Bischoffsheim: une approche biogéochimique. In: Lefranc Ph, Arbogast R-M (eds) Bischoffsheim : un village du Néolithique ancien en basse Alsace (France). Archéologiques, Musée national d'Histoire et d'Art du Grand Duché du Luxembourg, Luxembourg
Golitko M, Keeley LH (2006) Beating ploughshares back into swords: warfare in the Linearbandkeramik. Antiquity 81:332–342. https://doi.org/10.1017/S0003598X00095211
Gregg S (1988) Foragers and farmers: population interaction and agriculture expansion in prehistoric Europe. University of Chicago, Chicago
Gronenborn D (2003) Migration, acculturation and culture change in western temperate Eurasia, 6500-5000cal BC. Doc 30:79–9. https://doi.org/10.4312/dp.30.3
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron 4:9. http://palaeo-electronica.org/2001_1/past/issue1_01.htm
Handley LL et al (1999) The 15N natural abundance (δ15N) of ecosystem samples reflects measures of water availability. Funct Plant Biol 26:185–199. https://doi.org/10.1071/pp98146
Heaton THE, Vogel JC, von al Chevallerie G, Collet G (1986) Climatic influence on the isotopic composition of bone collagen. Nature 322:822–823. https://doi.org/10.1016/0031-0182(94)90096-5
Hedges R et al (2013) The supra-regional perspective. In: Whittle AWR, Bickle P (eds) The first farmers of Central Europe: diversity in LBK lifeways. Oxbow, Oxford, pp 343–384
Innes JB, Blackford JJ (2003) The ecology of Late Mesolithic woodland disturbances: model testing with fungal spore assemblage data. J Archaeol Sci 30:185–194. https://doi.org/10.1006/jasc.2002.0832
Ivanova M, De Cupere B, Ethier J, Marinova E (2018) Pioneer farming in southeast Europe during the early sixth millennium BC: climate-related adaptations in the exploitation of plants and animals. PLoS One 13:e0197225. https://doi.org/10.1371/journal.pone.0197225
Jim S, Ambrose SH, Evershed RP (2004) Stable carbon isotopic evidence for differences in the dietary origin of bone cholesterol, collagen and apatite: implications for their use in palaeodietary reconstruction. Geochim Cosmochim Acta 68:61–72. https://doi.org/10.1016/s0016-7037(03)00216-3
Johnson EV, Timpson A, Thomas MG, Outram AK (2018) Reduced intensity of bone fat exploitation correlates with increased potential access to dairy fats in early Neolithic Europe. J Archaeol Sci 94:60–69. https://doi.org/10.1016/j.jas.2018.04.001
Kalis AJ, Merkt J, Wunderlich J (2003) Environmental changes during the Holocene climatic optimum in central Europe - human impact and natural causes. Quat Sci Rev 22:33–79. https://doi.org/10.1016/s0277-3791(02)00181-6
Kendall IP, Lee MRF, Evershed RP (2018) The effect of trophic level on individual amino acid δ15N values in a terrestrial ruminant food web. STAR 3:135–145. https://doi.org/10.1080/20548923.2018.1459361
Kendall IP, Woodward P, Clark JP, Styring AK, Hanna JV, Evershed RP (2019) Compound-specific δ15N values express differences in amino acid metabolism in plants of varying lignin content. Phytochemistry 161:130–138. https://doi.org/10.1016/j.phytochem.2019.01.012
Knoepp JD, Taylor RS, Boring LR, Miniat CF (2015) Influence of forest disturbance on stable nitrogen isotope ratios in soil and vegetation profiles. Soil Sci Soc Am J 79:1470–1481. https://doi.org/10.2136/sssaj2015.03.0101
Kreuz A (2007) Closed forest or open woodland as natural vegetation in the surroundings of Linearbandkeramik settlements? Veg Hist Archaeobotany 17:51–64. https://doi.org/10.1007/s00334-007-0110-1
Kreuz A, Schäfer E (2011) Weed finds as indicators for the cultivation regime of the early Neolithic Bandkeramik culture? Veg Hist Archaeobotany 20:333–348. https://doi.org/10.1007/s00334-011-0294-2
Kreuz A, Marinova E, Schäfer E, Wiethold J (2005) A comparison of early Neolithic crop and weed assemblages from the Linearbandkeramik and the Bulgarian Neolithic cultures: differences and similarities. Veg Hist Archaeobotany 14:237–258. https://doi.org/10.1007/s00334-005-0080-0
Lazaridis I et al (2016) Genomic insights into the origin of farming in the ancient Near East. Nature 536:419–424. https://doi.org/10.1038/nature19310
Lipson M et al (2017) Parallel palaeogenomic transects reveal complex genetic history of early European farmers. Nature 551:368–372. https://doi.org/10.1038/nature24476
Liu X-Y, Koba K, Makabe A, Liu C-Q (2014) Nitrate dynamics in natural plants: insights based on the concentration and natural isotope abundances of tissue nitrate. Front Plant Sci 5:1–14. https://doi.org/10.3389/fpls.2014.00355
Lüning J (2000) Steinzeitliche Bauern in Deutschland. Die Landwirtschaft im Neolithikum. Habelt, Bonn. 285p
Magyari EK, Chapman JC, Passmore DG, Allen JRM, Huntley JP, Huntley B (2010) Holocene persistence of wooded steppe in the Great Hungarian Plain. J Biogeogr 37:915–935. https://doi.org/10.1111/j.1365-2699.2009.02261.x
Makarewicz CA (2014) Winter pasturing practices and variable fodder provisioning detected in nitrogen (δ15N) and carbon (δ13C) isotopes in sheep dentinal collagen. J Archaeol Sci 41:502–510. https://doi.org/10.1016/j.jas.2013.09.016
Makarewicz CA (2015) Winter is coming: seasonality of ancient pastoral nomadic practices revealed in the carbon (δ13C) and nitrogen (δ15N) isotopic record of Xiongnu caprines. Archaeol Anthropol Sci 9:405–418. https://doi.org/10.1007/s12520-015-0289-5
Manning K, Downey SS, Colledge S, Conolly J, Stopp B, Dobney K, Shennan S (2013a) The origins and spread of stock-keeping: the role of cultural and environmental influences on early Neolithic animal exploitation in Europe. Antiquity 87:1046–1059
Manning K, Stopp B, Colledge S, Downey SS, Connolly J, Dobney K, Shennan S (2013b) Animal exploitation in the Early Neolithic of the Balkans and Central Europe. In: Colledge S, Conolly J, Dobney K, Manning K, Shennan S (eds) The origins and spread of domestic animals in Southwest Asia and Europe. Left coast press, Califonia, pp 237–252
Marciniak A (2005) Placing animals in the Neolithic: social zooarchaeology of prehistoric farming communities. University College London Press, London
Marciniak A (2013) Origin of stock-keeping and the spread of animal exploitation strategies in the early and middle Neolithic of the North European Plain. In: Colledge S, Conolly J, Dobney K, Manning K, Shennan S (eds) The origins and spread of domestic animals in Southwest Asia and Europe. Left Coast Press, Walnut Creek, pp 221–236
Martinelli LA et al (1999) Nitrogen stable isotopic composition of leaves and soil: tropical versus temperate forests. Biogeochemistry 46:46–65
Meadows J, Müller-Scheeßel N, Cheben I, Agerskov Rose H, Furholt M (2019) Temporal dynamics of Linearbandkeramik houses and settlements, and their implications for detecting the environmental impact of early farming. Holocene. https://doi.org/10.1177/0959683619857239
Minagawa M, Wada E (1984) Stepwise emirchment of 15N along food chains : further evidence and the relation between d15N and animal age. Geochim Cosmochim Acta 48:1135–1140
Moskal-del Hoyo M (2013) Mid-Holocene forests in eastern Hungary: new anthracological data. Rev Paleobot Palynol 193:70–81
Müller-Scheeßel N, Cheben I, Furholt M (2020) The LBK site of Vráble/Southwest Slovakia: results of the excavation season 2016. BerRGK 97:83–130
Nielsen AB et al (2012) Quantitative reconstructions of changes in regional openness in north-Central Europe reveal new insights into old questions. Quat Sci Rev 47:131–149. https://doi.org/10.1016/j.quascirev.2012.05.011
Nitsch EK, Charles M, Bogaard A (2015) Calculating a statistically robust δ13C and δ15N offset for charred cereal and pulse seeds. STAR 1:1–8. https://doi.org/10.1179/2054892315y.0000000001
Noe-Nygaard N, Price TD, Hede SU (2005) Diet of aurochs and early cattle in southern Scandinavia: evidence from 15N and 13C stable isotopes. J Archaeol Sci 32:855–871. https://doi.org/10.1016/j.jas.2005.01.004
Oelze VM, Siebert A, Nicklisch N, Meller H, Dresely V, Alt KW (2011) Early Neolithic diet and animal husbandry: stable isotope evidence from three Linearbandkeramik (LBK) sites in Central Germany. J Archaeol Sci 38:270–279. https://doi.org/10.1016/j.jas.2010.08.027
O'Leary MH (1988) Carbon isotopes in photosynethsis: fractionation techniques may reveal new aspects of carbon dynamics in plants. Bioscience 38:328–335
Pavlů I (2005) The Neolithisation of Central Europe. ArchRoz LVII:293–302
Pokorný P, Chytrý M, Juřičková L, Sádlo J, Novák J, Ložek V (2015) Mid-Holocene bottleneck for central European dry grasslands: did steppe survive the forest optimum in northern Bohemia, Czech Republic? Holocene 25:716–726. https://doi.org/10.1177/0959683614566218
Rasmussen P (1993) Analysis of goat/sheep faeces from Egolzwil 3 Switzerland: evidence for branch and twig foddering of livestock in the Neolithic. J Archaeol Sci 20: 479–502. https://doi.org/10.1006/jasc.1993.1030.
Read H (2003) A study of practical pollarding techniques in northern Europe. http://www.ancienttreeforum.co.uk/resources/other-publications/.Accessed Sept 2017
Robbins CT, Felicetti LA, Sponheimer M (2005) The effect of dietary protein quality on nitrogen isotope discrimination in mammals and birds. Oecologia 144:534–540. https://doi.org/10.1007/s00442-005-0021-8
Russell N (1998) Cattle as wealth in Neolithic Europe: where's the beef? In: Bailey D (ed) The archaeology of value: essays on prestige and the process of valuation, vol 730. Vol BAR international, Oxford, pp. 42-54.
Salque M, Bogucki P, Pyzel J, Sobkowiak-Tabaka I, Grygiel R, Szmyt M, Evershed RP (2013) Earliest evidence for cheese making in the sixth millennium BC in northern Europe. Nature 493(7433): 522-525. https://doi.org/10.1038/nature11698
Saqalli M, Salavert A, Bréhard S, Bendrey R, Vigne J-D, Tresset A (2014) Revisiting and modelling the woodland farming system of the early Neolithic linear pottery culture (LBK), 5600–4900 B.C. Veg Hist Archaeobotany 23:37–50. https://doi.org/10.1007/s00334-014-0436-4
Schnyder H, Schwertl M, Auerswald K, Schaufele R (2006) Hair of grazing cattle provides an integrated measure of the effects of site conditions and interannual weather variability on δ13C of temperate humid grassland. Glob Chang Biol 12:1315–1329. https://doi.org/10.1111/j.1365-2486.2006.01169.x
Schoeninger MJ, DeNiro MJ (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim Cosmochim Acta 48:625–639
Schroedter TM (in press) Chapter 5.6 little but worth it – Anthracological data and thoughts on forestation in the surroundings of the LBK and Želiezovce settlement site of Vráble. In: Furholt M, Cheben I, Müller J, Bistáková A, Wunderlich M & Müller-Scheeßel N (eds) Archaeology in the Zitava Valley 1 - the LBK settlement site of Vráble. SideStone Press, Leiden
Sealy J, Van der Merwe NJ, Lee Thorp JA, Lanham JL (1987) Nitrogen isotope ecology in southern Africa: implications for environment and dietray tracking. Geochim Cosmochim Acta 51:2707–2717
Shennan S, Downey SS, Timpson A, Edinborough K, Colledge S, Kerig T, Manning K, Thomas MG (2013) Regional population collapse followed initial agriculture booms in mid-Holocene Europe. Nat Commun 4:2486. https://doi.org/10.1038/ncomms3486
Simmons IG (1996) The environmental impact of later Mesolithic cultures. Edinburgh University Press, Edinburgh
Smith BN, Epstein HE (1971) Two categories of 13C/12C ratios for higher plants. Annu Rev Plant Biol 47:380–384
Šolcová A et al (2018) Early and middle Holocene ecosystem changes at the Western Carpathian/Pannonian border driven by climate and Neolithic impact. Boreas. https://doi.org/10.1111/bor.12309
Sponheimer M et al (2003a) An experimental study of carbon-isotope fractionation between diet, hair, and feces of mammalian herbivores Canadian. J Zool 81:871–876. https://doi.org/10.1139/z03-066
Sponheimer M et al (2003b) Nitrogen isotopes in mammalian herbivores: hair δ15N values from a controlled feeding study. Int J Osteoarchaeol 13:80–87. https://doi.org/10.1002/oa.655
Stevens RE, Lister AM, Hedges RE (2006) Predicting diet, trophic level and palaeoecology from bone stable isotope analysis: a comparative study of five red deer populations. Oecologia 149:12–21. https://doi.org/10.1007/s00442-006-0416-1
Styring AK, Fraser RA, Bogaard A, Evershed RP (2014) The effect of manuring on cereal and pulse amino acid δ15N values. Phytochemistry 102:40–45. https://doi.org/10.1016/j.phytochem.2014.02.001
Styring A, Rösch M, Stephan E, Stika H-P, Fischer E, Sillmann M, Bogaard A (2017) Centralisation and long-term change in farming regimes: comparing agricultural practices in Neolithic and Iron Age south-west Germany. Proc. Prehist. Soc. 83:357–381. https://doi.org/10.1017/ppr.2017.3
Tieszen LT (1991) Natural variations in the carbon isotope values of plants: implications for archaeology, ecology and paleoecology. J Archaeol Sci 18:227–248
Tresset A, Vigne JD (2001) La chasse, principal élément structurant la diversité des faunes archéologiques du Néolithique ancien, en Europe tempérée et en Méditerranée: tentative d’interprétation fonctionnelle Paper presented at the Rôle et statut de la chasse dans le Néolithique ancien danubien (5500-4900 av. J.-C.) In: Arbogast R-M Jeunesse C, Schibler J (Eds) Rôle et statut de la chasse dans le Néolithique ancien danubien (5500–4900 av. J.-C.), Marie Leidorf, Rahden/West. (2001), p. 129–151
Tuross N, Fogel ML, Hare PE (1988) Variability in the preservation of the isotopic composition of collagen from fossil bone. Geochim Cosmochim Acta 52:929–935. https://doi.org/10.1016/0016-7037(88)90364-x
Vaiglova P et al (2014a) An integrated stable isotope study of plants and animals from Kouphovouno, southern Greece: a new look at Neolithic farming. J Archaeol Sci 42:201–215. https://doi.org/10.1016/j.jas.2013.10.023
Vaiglova P, Snoeck C, Nitsch E, Bogaard A, Lee-Thorp J (2014b) Impact of contamination and pre-treatment on stable carbon and nitrogen isotopic composition of charred plant remains. Rapid Commun Mass Spectrom 28:2497–2510. https://doi.org/10.1002/rcm.7044
Van der Merwe NJ, Medina E (1991) The canopy effect carbon isotope ratios and foodwebs in amazonia. J Archaeol Sci 18:249–259
van Klinken GJ (1999) Bone collagen quality indicators for Palaeodietary and radiocarbon measurements. J Archaeol Sci 26:687–695
Vanderklift MA, Ponsard S (2003) Sources of variation in consumer-diet δ15N enrichment: a meta-analysis. Oecologia 136:169–182. https://doi.org/10.1007/s00442-003-1270-z
Vera F (2000) Grazing ecology and Forest history. Cabi publishing, Oxfordshire
Vogel JC (1978) Recycling of carbon in a forest environment. Acta Oecologica, Oecologia Plant 13:89–94
Zanon M, Davis BAS, Marquer L, Brewer S, Kaplan JO (2018) European Forest cover during the past 12,000 years: a palynological reconstruction based on modern analogs and remote sensing. Front Plant Sci 9:253. https://doi.org/10.3389/fpls.2018.00253
Acknowledgements
The analysis of the material was undertaken as a part of REG’s post-doc funded by the Deutsche Forschungsgemeinschaft (DFG) at the Graduate School, Kiel (GSC 208). The excavations directed by NM-S, MF and IC were funded by SFB 1266 Project ref. 2901391021 and VEGA-Project 2/0107/17. The analyses of botanical remains were conducted within the SFB 1266 project. Many thanks go to Dr. Marie Balasse and Denis Fiorillo at the SMIMMS platform (MNHN, Paris) and Dr. Robert Michener at the stable isotope lab University of Boston for assistance with mass spectrometry. Finally, we thank the two anonymous reviewers for their helpful comments in improving the manuscript.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
REG and CAM designed the study and wrote the manuscript with contributions from RE (zooarchaeology), DF (archaeobotany), NM-S, MF and IC (archaeology).
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gillis, R.E., Eckelmann, R., Filipović, D. et al. Stable isotopic insights into crop cultivation, animal husbandry, and land use at the Linearbandkeramik site of Vráble-Veľké Lehemby (Slovakia). Archaeol Anthropol Sci 12, 256 (2020). https://doi.org/10.1007/s12520-020-01210-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-020-01210-2