
Abstract
In Egypt, Wadi Qena Middle Eastern Desert, Paleogene successions exposed at Hawashiya and Abu Had sections from north to south were measured and comprehensively investigated for their foraminiferal content. The Dakhla, Tarawan, and Esna formations make up the investigated sections. The studied successions are subdivided into six biozones; they are five zones of the Late Paleocene (Igorina albeari subzone (P3b), G. Globanomalina pseudomenardii/P. Parasubbotina variospira subzone (P4a), Acarinina subsphaerica subzone (P4b), A. soldadoensis/G. pseudomenardii subzone (P4c), Morozovella velascoensis zone (P5), and one zone of the Early Eocene (P. Pseudohastigerina wilcoxensis/M. velascoensis zone (E2). The paleoenvironmental interpretations and interpretation of sea level changes over the study area during the Paleocene-Eocene time interval have been carried out based on the lithostratigraphy, biostratigraphy, paleoecologic parameters (total foraminiferal number (TFN), diversity indices of foraminifera, life strategies of planktonic foraminifera, planktonic/benthonic (P/B) foraminiferal ratio, agglutinated/calcareous benthonic foraminiferal ratios, benthonic foraminiferal biofacies and total organic carbon (TOC), and calcium carbonate CaCo3%). Five sequences are identified in the studied Lower Paleogene successions in the Wadi Qena area. Four depositional sequences are determined in the Paleocene, and one in the Lower Eocene. A comprehensive description of these depositional sequences and their boundaries is provided. The findings of the current study are calibrated with sequence stratigraphy of the world sea-level variations.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
One of the most important periods of the Cenozoic is the Paleogene. The Late Paleocene/Early Eocene interval had witnessed significant worldwide climatic and environmental shifts, reflected in numerous lithological and paleontological events. These events coincided with the Paleocene/Eocene thermal maximum, a sudden warming occasion (Zachos et al. 2005). Kaiho et al. (2006) discovered that the extinction event, as well as the decrease in deep-water benthonic foraminifera test size, appears to be associated with a decrease in low oxygen levels. On the other hand, the increase in surface-water planktonic foraminifera test size could be attributed to a pronounced decrease in nutrient supply.
The extinction of many transitional and deep water benthonic foraminiferal species marks the P/E boundary)Kaiho 1994). While planktonic foraminifera did not extinct (Kelly et al. 1996). Nearly, all regions have seen major changes in foraminifera distribution and diversification. Thermocline inhabitants such as Subbotina vanished in the tropics, whereas Morozovella diversified with the presence of unusual taxa)Kelly et al.1996). The hyperthermals may have influenced marine phytoplankton in a variety of ways.
For a long time, many researchers have been interested in the stratigraphy of the Paleogene successions in Egypt. Among these are the following: Faris et al. (2000), Aubry et al. (2002), El-Deeb et al. (2000), Mandur and Bayoumi (2010), Mandur (2016). The studied successions in the center of Wadi Qena have not received enough attention due to the difficulty to reach. The Campanian–Maastrichtian succession of this region is studied by Hewaidy et al. (2019).
This research aims to build the sequence stratigraphic schemes in Wadi Qena with the aid of biostratigraphical and paleoenvironmental tools, depending on high-resolution analyses of the benthonic and planktonic foraminiferal content of the studied successions. To achieve the goal of this study, it is required to keep track of changes in the foraminifera parameters including the following: total foraminiferal number (TFN), diversity indices of foraminifera, life strategies of planktonic foraminifera, planktonic/benthonic (P/B) foraminiferal ratios, agglutinated/calcareous benthonic foraminiferal ratios, and total organic carbon (TOC) and calcium carbonate percentage (CaCO3%). In addition, determine the biofacies assemblages of the benthonic foraminiferal species using the statistical program (SPSS). The findings of the current study are calibrated with sequence stratigraphy and the world sea-level variations.
Geological setting and lithostratigraphy
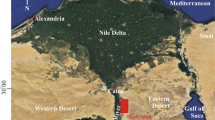
A notable feature of Egypt’s Eastern Desert is Wadi Qena. From north to south, it spans around 200 km, parallel to the Red Sea and the Nile River (Fig. 1), with a surface area of about 15,000 km2. Wadi Qana is located between latitudes 26°10′ and 28°00′ N and longitudes 32°15′ and 33°30′ E. It is a fault-bordered depression on the right bank of the Nile River. It stretches southward from the Nile Valley at Qena to the southern slopes of the Southern Galala. Vertical and lateral facies alterations from north to south distinguish the Lower Paleogene successions of the Eastern Desert. The lithostratigraphic divisions of the studied area are arranged from older to younger as follows:

Geological map of Wadi Qena area (modified after Conoco 1987), and the sites of the studied sections: (1) Hawashiya setion, (2) Abu Had section
Dakhla formation
Dakhla Formation consists of fissile shale which is the dominant rock type in this formation, with interbeds of siltstone and limestone. It typifies Dakhla Formation in its type locality in Gabal Gifata in Mut, Dakhla Oasis, Western Desert, as described by Said (1961). In north Wadi Qena, the Dakhla Formation unconformably overlies Sudr Formation and overlain unconformably by Tarawan Formation (Figs. 2, 4). It is extensively spread in central and southern Egypt. In the studied exposures of Hawashiya and Abu Had sections, Dakhla Formation is represented by Beida Shale Member. It measures around 16 m at the Hawashiya section and 7.5 m at the Abu Had section (Figs. 2, 4).

Planktonic foraminiferal distribution chart of Abu Had section
Tarawan formation
Tarawan Formation is primarily composed of chalk and chalky limestone; with occasional marl and shale intercalations. This lithology coincides with the description of the Tarawan Formation in the type section in Gabal Tarawan, Kharga Oasis that was identified by Awad and Ghobrial (1966). Tarawan Formation attains ≈7 m thick at Hawashiya and ≈7 m thick at Abu Had. It overlies unconformably Beida Shale Member and underlies Esna Formation in the studied area (Figs. 2, 4). It overlies unconformably with the Dakhla Formation and underlies the Esna Formation at the studied successions (Figs. 2, 4). Tarawan Formation has a wide geographic distribution; it is recorded in the oases of Farafra, Dakhla, and north Kharga in the southern and central Western Deserts, as well as the upper Nile Valley, the Red Sea region (Quseir-Safaga), and in Wadi Qena in the central Eastern Desert. This formation is therefore spread across central Egypt.
Esna formation
Said (1960) first described the Esna Formation in Gabal Owaina (Esna area) of the Nile Valley. Esna Formation conformably overlies Tarawan Formation and underlies Thebes Formation in the studied locality (Figs. 2, 4). It is distributed in the Sinai Peninsula, the Safaga-Quseir area of the Red Sea coastal plain, the Eastern Desert, the southern Western Desert, and the Nile Valley. In the study area, it is 12 m thick in the Hawashiya section and 10.5 m thick in the Abu Had section. This formation is made up of calcareous greenish-gray shale, which grades in the lower and upper parts into marl and argillaceous limestone respectively. Esna Formation is classified into two members by Abdel-Razik (1972), El Hanadi Member at the bottom and El-Shaghab at the top.
Materials and methods
Sixty-nine rock samples are collected from two Lower Paleogene successions at Hawashiya (lat. 28° 20′12″ N and long. 32° 22′75″ E) and Abu Had (lat. 28°53′12″ N and long. 32° 44′ 20″ E) in the northern and the southern parts the studied area respectively (Fig. 1). The rock samples are lithologically described before being prepared for micropaleontological study and microscopical examination. Dry rock samples weighing up to 10 g are soaked in hydrogen peroxide, run through a 63-mm sieve diluted in water, and dried in an oven. One of the foraminiferal metrics used in this study was the total estimated foraminiferal number (TFN) in 1 g of dry sediments. Using the indiscriminate square account in the selected dish, the planktonic/benthonic ratio (P/B %) is determined and expressed as P/(P + B) % in accordance with the planktonic foraminiferal number (PFN) and the benthonic foraminifera number (BFN). The number of individuals of one species in a sample divided by the total number of species in the same sample is used to calculate the Fisher Alpha Index, which measures species diversity. The SPSS application is used to conduct a factor analysis on the benthonic foraminiferal species to ascertain the relationship between the various connected groups and categorize them into biofacies assemblages. Benthonic foraminiferal biofacies, planktonic/benthonic ratio (P/B %) and diversity, and the total foraminiferal quantity are the paleoenvironmental factors identified in this study. LECO SC 632 equipment is used to measure total organic carbon (TOC).
Biostratigraphy
This research used the biostratigraphic zonation of planktonic foraminifera of the Paleocene-Eocene successions at Wadi Qena in order to enhance sequence stratigraphic interpretations. The planktonic foraminifers are useful tools for high-resolution biostratigraphy, as demonstrated in this study. The recognized biozones of planktonic foraminifera (Table 1) are correlated with those of the same interval of time, recorded in other parts of the world. Both the Paleocene/Eocene (P/E) border and the subchrones Selandian/Thanetian (S/T) boundary are discussed. The biozones are recognized by the first and last appearances of the marker species (FAD and LAD, respectively) or the lowest occurrence (LO) and the highest occurrence (HO) of the single taxon. In the present study, 87 species of planktonic foraminifera are identified and used to subdivide the Paleogene successions into their biozones. Some selected planktonic and benthonic foraminifera are illustrated. The standard biozonal scheme and estimated ages of Berggren and Pearson (2005) are used. The following is an in-depth explanation of the identified foraminiferal biozones, arranged from older to younger:
Igorina albeari subzone (P3b)
-
Author: (Berggren and Pearson 2005).
-
Definition: This zone is known as the interval from the FAD of Igorina albeari (Cushman and Bermudez) at its base to the FAD of Globanomalina pseudomenardii (Bolli) at the top.
-
Age: Early Late Paleocene (Selandian) (60.0–59.4 Ma, according to Luterbacher et al. 2004; Berggren and Pearson 2005).
-
Associated species: The most significant planktonic species found in this zone are as follows: Chiloguembelina midwayensis, Parasubbotina varianta, P. variospira, Subbotina triangularis, S. triloculinoides, Morozovella acuta, M. aequa, M. angulata, M. conicotruncata, Igorina pusilla, Praemurica praecursoria, P. uncinata, and Globanomalina imitate (Fig. 2). The benthonic foraminifera taxa which recorded in the limits of this subzone at the Abu Had succession are illustrated in Fig. 3.
-
Occurrence: Igorina albeari subzone occupies roughly 2 m thick of the Paleocene Dakhla Formation in the Abu Had section only (Fig. 2).
-
Remarks: This subzone corresponds to subzone P3b lowest-occurrence subzone (LO) (Berggren et al. 1995; Berggren and Pearson 2005; Wade et al. 2011). It correlates with Igorina albeari subzone recorded by El Deeb et al. (2000) in southwestern Sinai, and by Tantawy et al. (2001) in the Dakhla Oasis.

Benthonic foraminiferal distribution chart of Abu Had section
Globanomalina pseudomenardii/Parasubbotina variospira concurrent-range subzone (P4a)
-
Author: (Berggren and Pearson 2005).
-
Definition: Concurrent range of the naming taxa from the LO of Globanomalina pseudomenardii (Bolli) to the HO of Parasubbotina variospira (Belford) as defined by Berggren and Pearson (2005).
-
Age: Late Selandian (59.4–59.2 Ma) according to Cande and Kent (1995) and Berggren and Pearson (2005).
-
Associated species: The associated planktonic foraminiferal species in this subzone are as follows: Parasubbotina varianta, Morozovella angulata, M. conicotruncata, M. aequa, M. apanthesma, M. acuta, M. velascoensis, M. occlusa, M. pasionensis, Subbotina triangularis, S. velascoensis, Acarinina strabocella, Globanomalina chapmani, Igorina albeari, Acarinina nitida, and A. mckannai (Fig. 4). The recorded benthonic taxa in the limits of this subzone in Hawashiya section are illustrated in Fig. 5.
-
Occurrence: This subzone is roughly 3.5 m thick. It was found in the Paleocene of the Dakhla Formation in the Hawashiya section (Fig. 4), while absent in the Abu Had succession.
-
Remarks: It is equivalent to the lower portion of the Globanomalina pseudomenardii zone in Berggren and Pearson 2005; corresponding to the P4a concurrent-range subzone (Berggren et al. 1995; Faris et al 2000; El Deeb et al. 2000; Wade et al. 2011).

Planktonic foraminiferal distribution chart of Hawashiya section

Benthonic foraminiferal distribution chart of Hawashiya section
Acarinina subsphaerica partial-range subzone (P4b)
-
Author: (Berggren and Pearson 2005).
-
Definition: Acarinina subsphaerica subzone is a partial-range subzone of the naming taxon Acarinina subsphaerica as defined in Berggren and Pearson (2005). It spans the time interval from the HO of Parasubbotina variospira (Belford) to the LO of Acarinina soldadoensis (Bronnimann).
-
Age: Late Selandian/Early Thanetian (59.2–56.5 Ma according to Cande and Kent (1995) and Berggren and Pearson (2005).
-
Associated species: The dominant species of this zone are as follows: Globanomalina pseudomenardii, G. chapmani, Parasubbotina varianta, Ps. variospira, Morozovella angulata, M. conicotruncata, M. apanthesma, M. velascoensis, M. acuta, M. pasionensis, M. aequa, M. occlusa, Globanomalina chapmani, Subbotina cancellata, S. triangularis, S. velascoensis, S. triloculinoides, Igorina albeari, Acarinina strabocella, A. nitida, A. mckannai (Figs. 2, 4). The recorded benthonic foraminifera species that were recorded in the limits of this subzone in the studied successions are illustrated in Figs. 3 and 5.
-
Occurrence: Acarinina subsphaerica subzone reaches a thickness of 5.5 m in the Dakhla Formation Hawashiya section (samples 104 to 107) and 3.5 m in the Abu Had section (samples 30 to 32) (Figs. 2, 4).
-
Remarks: Acarina subsphaerica subzone is characteristic of the standard subzone (P4b) of (Berggren et al. 1995; Faris et al. 2000; El Deeb et al. 2000; Berggren and Pearson 2005; Wade et al. 2011).
Acarinina soldadoensis/Globanomalina pseudomenardii concurrent-range subzone (P4c)
-
Author: (Berggren and Pearson 2005).
-
Definition: Acarinina soldadoensis/Globanomalina pseudomenardii subzone is concurrent-range subzone of the naming taxa. It extends from the LO of Acarinina soldadoensis (Bronnimann) to the HO of Globanomalina pseudomenardii (Bolli).
-
Age: Late Thanetian (56.5–55.9 Ma according to Cande and Kent 1995; Berggren and Pearson 2005).
-
Associated species: The most important planktonic elements of this zone are as follows: Morozovella subbotinae, M. velascoensis, M. aequa, M. acuta, M. pasionensis, M. occlus, M. gracilis, Acarinina soldadoensis, A. nitida, A. mckannai, Igorina albeari, Subbotina velascoensis, S. triangularis, Parasubbotina variant, Globanomalina chapmani, and G.pseudomenardii (Figs. 2 and 4). Figures 3 and 5 illustrate the benthonic foraminiferal species that have been identified in the limits of this subzone in Hawashiya and Abu Had sections.
-
Occurrence: It is 11 m thick in the Dakhla and Tarawan formations at the Hawashiya section (samples 108 to 117), and 5.5 m thick at the Abu Had section in the same formations (samples 33 to 35). (Figs. 2, 4).
-
Remarks: This subzone characterizes the standard Late Thanetian subzone (P4c), corresponding to that described by Berggren et al. (1995), Faris et al. (2000), El Deeb et al. (2000), Berggren and Pearson (2005), and Wade et al. (2011).
Morozovella velascoensis partial-range zone (P5)
-
Author: (Berggren and Pearson 2005).
-
Definition: It is the biostratigraphic interval that defines the partial range of the naming taxon. It extends from the LAD of Globanomalina pseudomenardii (Bolli) to the FAD of Acarinina sibaiyaensis (El-Naggar).
-
Age: Latest Thanetian (55.9–55.5 Ma according to Cande and Kent 1995; Berggren and Pearson 2005).
-
Associated species: The most important planktonic foraminiferal elements of this zone are as follows: Morozovella subbotinae, M. aequa, M. acuta, M. pasionensis, M. occlus, M. gracilis, Acarinina soldadoensis, A. nitida, A. mckannai, A. sibaiyaensis, Igorina albeari, Subbotina velascoensis, S. triangularis, Parasubbotina variant, Globanomalina chapmani (Figs. 2, 4). Figures 3 and 5 depict the benthonic foraminiferal species of this zone at the Hawashiya and Abu Had sections.
-
Occurrence: It occupies Tarawan and Esna formations (8.5 m thick) at the Hawashiya section (samples 118 to 124) while at the Abu Had section, it is 9 m thick (samples 36 to 40) (Figs.2, 4).
-
Remarks: This zone was described by Berggren et al. (1995), Faris et al. (2000), El Deeb et al. (2000), and Berggren and Pearson (2005). Morozovella velascoensis is an index Latest Thanetian species characteristic for the standard zone (P5).
Pseudohastigerina wilcoxensis/Morozovella velascoensis concurrent-range zone (E2)
-
Author: (Berggren and Pearson 2005).
-
Definition: It covers an interval occupied by the concurrent ranges of the naming taxa, from the FAD of Pseudohastigerina wilcoxensis (Cushman and Ponton) to the LAD of Morozovella velascoensis (Cushman).
-
Age: Early Eocene (55. 35–54.5 Ma according to Cande and Kent (1995) and Berggren and Pearson (2005)
-
Associated species: It is dominated by representatives of the genus Morozovella as M. subbotinae, M. gracilis, and Acarinina soldadoensis (Figs. 2, 4). The benthonic foraminiferal species recorded in the limits of this zone are shown in Figs. 3 and 5.
-
Occurrence: This zone occupies the upper part of the Esna Formation measuring 6.5 m thick at the Hawashiya section (samples 125 to 127) and 5 m thick at the Abu Had section (samples 41 to 42) (Figs. 2, 4).
-
Remarks: Pseudohastigerina wilcoxensis/Morozovella velascoensis zone (E2) is concurrent-range zone as defined by Faris et al. (2000), El Deeb et al. (2000), Berggren and Pearson (2005), and Wade et al. (2011). It is characteristic of the standard Zone (E2). It is equivalent to Pseudohastigerina wilcoxensis/Morozovella velascoensis subzone of Berggren and Ouda (2003), and to the upper part of Acarinina sibaiyaensis/Morozovella velascoensis (P5b) subzone of Hamad (2009).
Stage boundaries
The Selandian/Thanetian (Se/Th) boundary
Along the outcropping coastal cliff at Itzurun Beach, in Zumaia, Basque Country, northern Spain, the global stratotype section and points (GSSP) for the base of the Selandian (Middle Paleocene) and Thanetian (Upper Paleocene) stages have been determined. According to Schmitz et al. (2011), the base of the Itzurun Formation of the Selandian Stage is located approximately 49 m above the K/P boundary, and that of the Thanetian Stage is located approximately 78 m above the K/P boundary. The Se/Th boundary can be tentatively located in the middle of the planktonic foraminiferal Acarinina subsphaerica subzone (P4b) (Berggren and Pearson 2005). Hawashiya and Abu Had successions of the current study comprise records of the Se/Th boundary. It is detected within the Tarawan Formation, between samples 116 and 117, at the Hawashiya section (Fig. 4), and the Abu Had section occurs between samples 36 and 37 (Fig. 2).
The Paleocene/Eocene (P/E) boundary
The global standard stratotype section and point (GSSP) of the P/E boundary or the base of the Eocene Series have been located at the base of the Dababiya Quarry = member, which occur in the lower part of Esna Formation in the abandoned quarry of Dababiya, near Luxor, Egypt (Aubry et al. 2007). It is an incredible track record of the carbon isotope excursion (CIE) and the P/E thermal maximum (PETM) (Aubry et al. 2007; Berggren et al. 2012). The P/E boundary is recorded in the Hawashiya and Abu Had sections within the Esna Formation between samples 124/125 at the first and between samples 40/41 at the second. It is dominated by flat-spired Acarinina, including A. sibuyanensis and related taxa typical of the Paleocene/Eocene thermal maximum (PETM) at the planktonic foraminiferal, while Mandur et al. (2022) recorded at nannoplankton; it is dominated by Tribrachiatus orthostylus, Tribrachiatus contortus, Fasciculithus tympaniformis, and Discoaster sp.
Paleoenvironment
A principal goal of this study is to provide paleoenvironmental interpretations during Paleocene-Lower Eocene interval in Wadi Qena in terms of foraminiferal parameters. The use of foraminifera by this way is well-known and valuable because it is extremely sensitive to changes in various ecological parameters (Van der Zwaan et al. 1990; Leckie and Olsson 2003). The parameters of foraminifera used in this research are as follows: total foraminiferal number (TFN) in 1 g of dry sediments, the planktonic/benthonic ratio (P/B %) is estimated in the selected tray using random square quantification and expressed as P/(P + B) % based on the planktonic foraminiferal number (PFN) and benthonic foraminifera number (BFN), Fisher Alpha Index is a quantitative measure of species diversity that is indicated by the proportion of the number of individuals of one species in a sample to the number of the total species within the same sample. To determine the relationship between the various associated groups and classify them into biofacies assemblages, the benthonic foraminifera species are subjected to factor analysis using the SPSS statistical program.
Total foraminiferal number (TFN)
The total number of foraminiferal individuals in 1 g of dry sediment is used to calculate the TFN. In the present study, it ranges from 1828 to 60,000(s/g) at the Hawashiya section (Fig. 6), and from 197 to 2616 (s/g) at the Abu Had section (Fig. 7).

Total foraminifera number (TFN), total planktonic foraminifera, total benthonic foraminifera, planktonic/benthonic (P/P + B) %, agglutinated/calcareous ratios (A/C), total agglutinated foraminifera (TA), total organic carbon (TOC), species richness, Fisher alpha index, and life strategy at Hawashiya section

Total foraminifera number (TFN), total planktonic foraminifera, total benthonic foraminifera, planktonic/benthonic (P/P + B) %, agglutinated/calcareous ratios (A/C), total agglutinated foraminifera (TA), total organic carbon (TOC), species richness, Fisher alpha index, and life strategy at Abu Had section
Diversity indices of foraminifera
A fraction of all species in the sample is used to assess the diversity using the Fisher alpha indicator. Fisher alpha index is computed using Fisher and Hay (past 3) statistical equation program. In the Paleocene Dakhla and Tarawan formations, the species richness at the Hawashiya section ranges between 40 and 60, with Fisher alpha ranging from 14.0 to 17.0 representing a high level of diversity. Also, at the Abu Had section, the species richness ranges between 30 and 45, with Fisher alpha ranging from 14.0 to 18.0 indicating high diversity (Figs. 6, 7). For the Early Eocene Esna Formation, the Hawashiya section has a species richness of 35–40 and Fisher alpha of 12.0–15.0 indicating high variety, whereas the Abu Had section has a species richness of 25–35 and Fisher alpha of 10.0–15.0, indicating low diversity (Figs. 6, 7).
Life strategies of planktonic foraminifera
Like modern planktonic foraminifera, the r- and k-strategy taxa are divisions of food communications that reflect their spectrum of environmental tactics (Table 2).
The intense thermocline and the k-strategy taxa characterized the oligotrophic regions, which have varied phytoplankton communities and specialized species.
The r-strategy taxa are influenced by shallow thermocline, well-mixed upper water column, low-diversity communities, and predatory organisms. The k/r strategy is mixed between r-strategy and k-strategy.
The morphology of planktonic foraminifera is a useful tool for detecting paleoenvironments (e.g., Silva and Sliter 1999; Keller et al. 2002; Petrizzo 2002; Krahl et al. 2017). There are three types of Paleogene planktonic foraminiferal groups. The r-strategy is distinguished by minor and straightforward morphotypes that are reminiscent of eutrophic marine ecosystems (e.g., Globanomalina, Pseudohastigerina, and Chiloguembelina). The k-strategy is assumed to comprise species with multifaceted morphologies as well as single and/or double keels (Acarinina, Morozovella, and Igorina). The k/r strategy is a combination of the r and k strategies, as denoted by Subbotina and Parasubbotina (Table 2).
Planktonic foraminiferal assemblage distribution of the Paleocene-Eocene interval at Hawashiya and Abu Had sections shows that Acarinina, Morozovella, and Igorina dominated large-sized planktonic foraminifera (k–strategy), ranging from 30 to 68% of total planktonic species. In the two studied sections, the r–strategy is frequently noted with fewer than 20% of total planktonic species, while the k/r strategy displays a survival strategy that is between 5 and 15% (Figs. 6, 7).
Planktonic/benthonic (P/B) foraminiferal ratio
The planktonic/benthonic ratio (P/B %) is calculated as P × 100/(P + B), where P is the total number of planktonic individuals in the samples examined and B is the total number of benthonic individuals. The P/B ratio was thought to be an effective value for determining the paleodepth (Van der Zwaan et al. 1990). Olsson and Nyong (1984) found that the inner shelf from 10 to 50 m depth is characterized by low planktonic percentages (1% and 5%), low diversity, and high benthonic fractions; the middle shelf depth between 50 and 100 m is characterized by planktonic foraminifera percentages from 8 to 25% and higher diversity; the outer shelf depth between 100 and 200 m has 30–70% planktonic foraminiferal accumulation, while the continental incline from 200 to 800 m is characterized by 90% of planktonic foraminifera.
In the Paleocene-Eocene interval, the P/B percent ratio is found with a dominantly low percentage of planktonic foraminiferal forms at the Hawashiya section, mostly around 20% and rarely reaching 70%. A generally high percentage of planktonic foraminiferal forms at the Abu Had section, ranging from 50 to 70% (Figs. 6, 7).
Agglutinated/calcareous benthonic foraminiferal ratios
With increasing depth, the agglutinated/calcareous benthonic ratio decreases (Saint-Marc 1992). The agglutinated/calcareous benthonic ratio in the current study ranges from low to high, pointing to a significant change in the paleodepth of deposition. In Hawashiya and Abu Had successions, the agglutinated/calcareous benthonic ratio (A/C) in the Dakhla Formation is significantly rising from 55 to 70%. In Tarawan Formation, it falls from 30 to 20%, and in the Esna Formation, it is rising from 20 to 50% (Figs. 6, 7).
Benthonic foraminiferal biofacies
The benthonic biofacies are ranked in terms of relative depth using foraminifera parameters including planktonic foraminifera percent and benthonic foraminifera percent and compared with recent foraminiferal distributions. Paleo-water depth curves for Hawashiya and Abu Had sections are created (Figs. 8, 9). To determine the biofacies assemblages, the benthonic foraminiferal taxa are statistically analyzed and categorized into seven biofacies, which are arranged from shallow to deep into the following:

Distribution of the benthonic foraminiferal biofacies at Hawashiya section

Distribution of the benthonic foraminiferal biofacies at Abu Had section
Lenticulina biofacies
It was found in the Abu Had section especially in the Late Paleocene Tarawan Formation (samples 34–37) (Fig. 9). It is defined by the frequent and widespread occurrence of Lenticulina midwayensis, L. obesus, L. pseudosecans, L. rotulata, Cibicidoides cookei, and Siphonina eocenica. It depicts the middle to outer shelf ecosystem at 50–100 m of sea depth (Nouradini et al. 2015; Székely et al. 2017).
Lagena biofacies
It is found in the Late Paleocene shales of the Esna Formation, occupying the limits of Morozovella velascoensis biozone at Abu Had succession only (Fig. 9). It is characterized by Lagena spp. and Oridorsalis plummerae, while Lenticulina spp. represents the second main component of this biofacies assemblage. This biofacies is commonly originated in an inner shelf environment (Gebhardt et al. 2013).
Pullenia biofacies
It was discovered in the Upper Paleocene limestone facies of the Tarawan Formation, occupying the stratigraphic range of Acarinina soldadoensis/Globanomalina pseudomenardii biozone at the Hawashiya section (Fig. 8). It is characterized by Pullenia coryelli, Pullenia americana, and Tappanina selmensis. Pullenia biofacies is commonly originated in inner to middle neritic environments (Jennings et al. 2014).
Alabamina biofacies
It originated in the Early Eocene shale facies of the Esna Formation and occupied the interval of the time of Pseudohastigerina wilcoxensis/Morozovella velascoensis biozone at the Hawashiya section (Fig. 8). It is distinguished by Factor 11 (2% of the overall explained variation by the SPSS statistically analyzed program) and known by the presence of Nonionella africana, Nodosaria radicula, Marginulina carri, and Lenticulina sp. Alabamina biofacies is commonly originated in middle to outer neritic environments (Sprong et al. 2012).
Angulogavelinella biofacies
It is recorded in the shale facies of the Dakhla Formation and occupies the interval of time that is defined by Globanomalina pseudomenardii/Parasubbotina variospira and Acarinina subsphaerica biozones at Hawashiya and Abu Had successions (Figs. 8, 9). It is distinguished by Osangularia plummerea, Gyroidinoides girardanus, G. subangulata, Cibicidoides sp., Bulimina sp., Praebulimina sp., and Angulogavelinella abudurbensis, and Angulogavelinella biofacies is commonly distributed in the outer neritic environment (Bice and Norris 2002; Guasti et al. 2006).
Loxostomoides biofacies
It is recorded in the shale facies of the Esna Formation and deposited during the Early Eocene and occupied by Pseudohastigerina wilcoxensis/Morozovella velascoensis biozone at the Abu Had section only (Fig. 9). It is distinguished by Lagena spp., Gyroidinoides girardanus, and Oridorsalis plummerae, whereas the Gyroidinoides spp. signified the main component of this biofacies. The Loxostomides biofacies is found in outer neritic to upper bathyal environments (Alegret and Thomas. 2005; El-Dawy et al. 2016).
Bulimina biofacies
It is recorded in the Lower Paleocene shale facies of the Dakhla Formation and occupies the time interval of Acarinina subsphaerica and Acarinina soldadoensis/Globanomalina pseudomenardii biozones at Hawashiya and Abu Had sections (Figs. 8, 9). It is characterized by Bulimina farafraensis, B. midwayensis, B. quadrata, B. strobili, B.pupoides, B.reussi, Cibicides sp., Quadrimorphina allomorphinoides, and Valvalabamina sp. The Bulimina biofacies is typically found in a lower bathyal environment (Baldanza et al. 2013; Ghourchaei et al. 2015).
Total organic carbon (TOC) and calcium carbonate (CaCo3%)
The total organic carbon (TOC) is calculated for tests that have been pretreated with concentrated hydrochloric acid, and it is measured using LECO SC 632 equipment using the LECO technique. The contents of TOC changed from low to high values from north to south in the Wadi Qena areas. The TOC values were increased at the Dakhla Formation and ranged between 0.25 and 0.29 at the Hawashiya and Abu Had sections. The TOC values were sharply decreased at the Tarawan Formation and ranged between 0.11 and 0.17 at the Hawashiya and Abu Had sections. The TOC values were increased at the Esna Formation and ranged between 0.25 and 0.27 at the Hawashiya and Abu Had sections (Figs. 6 and 7). The calcium carbonate (CaCo3) was determined by the mass of total inorganic carbon in 1 g of the total sample. It was noted that the CaCo3% in the Dakhla Formation was decreased in CaCo3% values ranging from 4.0 to 10.0% at Hawashiya and Abu Had sections. The Tarawan Formation was sharply increased in (CaCo3%) values ranging from 70.0 to 90.0% at Hawashiya and Abu Had sections, and in the Esna Formation at Hawashiya and Abu Had sections, CaCo3%values decreased from 2.0 to 7.0 (Figs. 6 and 7).
Sequence stratigraphy
Sequence stratigraphy is a useful tool for relating different environments to specific time intervals accompanied by regional and global correlation. This study focuses on the sequence stratigraphic classification of Paleocene to Lower Eocene successions exposed at the investigated areas. The transgressive systems tract (TST) and highstand systems tract (HST) can be identified in this classification. Because of the rapid relative sea level rise over the study area, the lowstand system tract is missing in the identified depositional sequences.
Five depositional sequences are recognized in the studied sections. These are the A, B, C, D, and E sequences. The sequences are precisely dated using marker and zonal species of the identified planktonic foraminifera. The various sequences are distinguished into systems tracts as the reflectance of certain relative sea level changes. The identified sequences are matched with the sea level curves from Haq et al. (1988), Haq and Schutter (2008), and Gradstein et al. (2012) as well as other regional and local charts. The detected depositional sequences and their boundaries are described below in detail.
The sequence boundary 1 (SB 1)
The sequence boundary 1(SB1) is a regional unconformity in southern Egypt that was reported in several publications (Tantawy 2003). It represents the Cretaceous-Paleogene (K/P) transition. At the Abu Had section, (SB1) is recorded within Dakhla Formation, between the Hamama and Beida members. It can be identified by the presence of a band of calcareous clay and marly clay in its dark gray shale. It is indicated by the disappearance of the Early Paleocene planktonic biozones (CF6) (Ayyat et al. 2013; Hewaidy, et al 2017) to (P3a).
Depositional sequence A (DS A)
It is denoted by a 2 m thick from the Lower Paleocene Dakhla Formation at the Abu Had section (Fig. 11). Only a transgression system tract (TST) is present. No lowstand or highstand system tracts are identified as a result of the rapid relative sea level rise and erosion.
Transgressive systems tract-A (TST-A)
At Abu Had, it reaches a thickness of 2 m of the Lower Paleocene Dakhla Formation and occupies the range of the Igorina albeari zone (P3b). It is collected of dark gray shale with k-strategy. It is marked by the following: (1) a high P/B% ratio fluctuating from 50 to 65%, with moderate preservation of planktonic foraminifera, (2) a visible rise in TFN oscillating from 700 to 1000 (s/g), (3) a universal uptick in the diversity of fauna (Fisher alpha index going from 18.787 to 22.8516 with species diversity 44–60, (4) a frequent occurrence of Angulogavelinella biofacies. These results point to a transgressive phase, an increase in relative sea level, and a predominance of an outer neritic environment during the deposition of this transgressive systems tract (Fig. 12).
The sequence boundary 2 (SB 2)
Only the Hawashiya section contains a recording of this sequence boundary. At the Cretaceous/Paleogene (K/P) transitions, it is signified by the unconformity surface between the Sudr and Dakhla formations. By having dark gray shale with marly clay intercalation, the sequence boundary 2 (SB 2) can be clearly seen.
Depositional sequence B (DS B)
At the Hawashiya section, it is mainly documented by 3.5 m of the Lower Paleocene Dakhla Formation. (Fig. 10). It consists of a transgressive systems tract (TST) only. No lowstand or highstand system tracts exist due to erosion or rapid sea level rise.

Sequence stratigraphy at Hawashiya section, Wadi Qena
Transgressive systems tract-B (TST-B)
It is represented by 3.5 m thick of the Paleocene Dakhla Formation at Hawashiya succession (Fig. 10). It occupies the range of Globanomalina pseudomenardii/Parasubbotina variospira subzone (P4a). It is composed of dark gray shale with k-strategy. It is characterized by the following: (1) high P/B% ratios ranging from 80 to 84%, with moderate preservation of planktonic foraminifera, (2) a strong upsurge in TFN ranging from 3580 to 60,000(s/g), (3) a common rise in faunal diversity (Fisher alpha index ranging from 12.3 to 15 with species richness 39–57, (4) a plentiful appearance of Angulogavelinella biofacies. These results demonstrate an increase in relative sea level, which denotes a transgression phase, and a predominance of an outer neritic environment during the deposition of this transgressive systems tract, (Fig. 12).

Sequence stratigraphy at Abu Had section, Wadi Qena
The sequence boundary 3 (SB 3)
This sequence boundary (SB 3) is present at Hawashiya and Abu Had successions, where it is marked by a ledge of ferruginous calcareous limestone within dark gray to green shale (Figs. 10, 11). The large eustatic sea-level fall at the Early Selandian correlated to Th-2 of Gradstein et al. (2012), Haq and Schutter (2008), and the present sequence boundary (SB3) (Fig. 12).

Sequence stratigraphic correlation of the studied Hawashiya and Abu Had sections, Wadi Qena and the standard sea-level charts
Depositional sequence C (DS C)
It is epitomized by 19.5-m-thick and 13-m-thick Dakhla and Tarawan formations at Hawashiya and Abu Had successions, respectively (Figs. 10, 11), occupying the interval from Acarinina subsphaerica zone (P4b) to Acarinina soldadoensis/Globanomalina pseudomenardii zone (P4c). A transgressive system tract (TST) and highstand system tract (HST) are distinguished in this depositional sequence.
Transgressive systems tract-C (TST-C)
(TST-C) reaches about 19.5 m thick at the Hawashiya section, and 13 m thick at Abu Had section. It covers the Acarinina subsphaerica zone (P4b), the Acarinina soldadoensis/Globanomalina pseudomenardii zone (P4c), and the lower part of Morozovella velascoensis biozone Zone (P5) of the Late Paleocene age and occupies upper Dakhla Formation and almost Tarawan Formation (Figs. 10, 11). K-strategy gray shale makes up its composition. It is distinguished by the following: (1) high P/B% ratios between 83 and 85% at Hawashiya succession and 50 to 56% at Abu Had succession with large and moderate preservation, (2) a TFN between 2750 and 36,140 (s/g) at Hawashiya section and 700 to 780 (s/g) at Abu Had section, (3) an increase in faunal diversity (Fisher alpha index between 10.3 and 12.15 at Hawashiya section, and 12.8516 to 13.787 at Abu Had section, with species richness 70–75 and 35–40 at Hawashiya and Abu Had sections, respectively), and (4) an abundant occurrence of Bulimina biofacies in Hawashiya and Abu Had sections (Figs. 10, 11). This demonstrates a transgressive phase, obvious rise in relative sea level, and dominance of an outer neritic environment and shallow to middle neritic during the formation of this transgressive systems tract (Fig. 12).
Highstand systems tract- C (HST-C)
It is denoted by 11 m thick of the Tarawan Formation at the Hawashiya section, and 5.5 m thick of the Tarawan Formation at the Abu Had section of the Late Paleocene Acarinina soldadoensis/Globanomalina pseudomenardii zone (P4c) and the lower part of Morozovella velascoensis zone (P5) (Figs. 10, 11). It is distinguished by the following: (1) a low P/B% ratio varying between 50 and 45% at the Hawashiya succession and 42–50% at Abu Had succession with small and high preservation, (2) a TFN fluctuating between 2500 and 2714 (s/g) at Hawashiya succession and 720–784 (s/g) at Abu Had succession, (3) an increase in faunal diversity (Fisher alpha index fluctuating between 10.3 and 12.15 or 7.8516. This suggests a decline in relative sea level, a regressive phase, and shallow to intermediate neritic habitats were predominant at the time of deposition (HST-C) (Fig. 12).
The sequence boundary 4 (SB 4)
At Hawashiya and Abu Had sections, (SB4) corresponds to an unconformity surface that separates Tarawan and Esna formations. This boundary is indicated by the presence of an uneven surface of the unconformity at the base of the chalky limestone and the dark calcareous shale of El Hanadi Member (Figs. 10, 11). This boundary matches SB5 of El-Azabi and El-Araby (2000) at the base of the Tarawan Formation at the Dakhla-Farafra stretch Western Desert.
Depositional sequence D (DS D)
The time interval covered by this depositional sequence is synchronous with the Morozovella velascoensis zone (P5) of the Upper Paleocene age. It is represented by the lower part of Esna Formation at both Hawashiya and Abu Had sections (Figs. 10, 11). At Abu Had, it is 5 m thick, whereas at the Hawashiya section, it is around 5.5 m thick. At the Hawashiya and Abu Had sections, a highstand system tract (HST) represents this depositional phase.
Highstand systems tract- D (HST-D)
At both Hawashiya and Abu Had sections, (HST-D) is synchronous with the Late Paleocene Morozovella velascoensis zone (P5). It measures 8.5 m thick and 9 m thick of the Esna Formation rock at Hawashiya and Abu Had sections, respectively. It is distinguished by the following: (1) a low P/B% ratios fluctuating between 25 and 35% at Hawashiya succession and 32 and 40% at Abu Had section with large and high preservation, (2) a decline in the TFN, which fluctuated between 643 and 200 (s/g) at Hawashiya sectionand between 471 and 500 (s/g) at Abu Had section, (3) a broad decline in (Fisher alpha index ranging from 4.3 to 7.15 and 6.8516 to 4.787 with species richness 40–43 and 58–44 at Hawashiya and Abu Had successions, respectively, and (4) abundant occurrence of Lenticulina biofacies in Hawashiya succession changed to Lagena biofacies in Abu Had succession (Figs. 10, 11). This suggests a regressive period with a decline in relative sea level, and the environment was shallow to moderate neritic at the time of deposition of (HST-D) (Fig. 12).
The sequence boundary 5 (SB 5)
The sequence boundary 5 (SB 5) is coeval the Paleocene/Early Eocene (P/E) transition. This boundary is comfirmed by the absence of the earliest Eocene planktonic biozone (E1). It is equivalent to the Early Eocene sequence boundaries YP-4 of Gradstein et al. (2012) and Haq and Schutter (2008) (Figs. 10 and 11).
Depositional sequence E (DS-E)
This depositional sequence is synchronous with the Early Eocene (Ypresian) Pseudohastigerina wilcoxensis/Morozovella velascoensis biozone zone (E2). It occupies El Shaghab Member of the Esna Formation at both Hawashiya and Abu Had sections (Figs. 10, 11). It is 5 m thick at Abu Had and 6.5 m thick at Hawashiya sections. This depositional sequence is represented by a transgressive system tract (TST) in the Hawashiya and Abu Had sections.
Transgressive systems tract- E (TST-E)
It is represented by a 6.5 m thick section at the Hawashiya section, and about 5 m thick at the Abu Had section, built up of Esna Formation rocks (El Shaghab Member). (TST-E) is coeval Pseudohastigerina wilcoxensis/Morozovella velascoensis zone of the Early Eocene age (Figs. 10, 11). It is made up mostly of gray shale with a high K-strategy content and described by the following: (1) a high P/B% ratio attaining from 85 to 90% at the Hawashiya section and 68–75% at the Abu Had section with large and moderately preserved forms, (2) a TFN ranging from 2530 to 2616 (s/g) at the Hawashiya section and 1234 to 1300 (s/g) at the Abu Had section, (3) a general rise in faunal diversity (Fisher alpha index fluctuating from 5.3 to 7.5 and 11.8516 to 12.787 with species richness 54–63 and 58–60 at Hawashiya and Abu Had sections, respectively, and (4) a plentiful incident of Alabamina biofacies in Hawashiya section changed to Loxostomides biofacies in Abu Had section (Figs. 11, 12). This suggests a transgression phase and an increase in relative sea level, with an outer neritic environment predominated at the time of deposition (TST-E) (Fig. 12).
Conclusions
The purpose of this research is to investigate the lithostratigraphy, biostratigraphy, paleoenvironmental interpretation, and sequence stratigraphy of the Lower Paleogene succession exposed at two sections in Wadi Qena, Eastern Desert, Egypt. These are Hawashiya and Abu Had sections. Lithostratigraphically, the Paleogene sediments are divided into three formations, from base to top: Dakhla (Beida Shale Member), Tarawan, and Esna. The studied planktonic foraminifera are used to identify six planktonic foraminiferal biozones, which are arranged from older to younger:
-
(1)
Igorina albeari Zone (P3b) Early Paleocene
-
(2)
G. pseudomenardii/P. variospira subzone (P4a) Late Paleocene
-
(3)
Acarinina subsphaerica subzone (P4b) Late Paleocene
-
(4)
A. soldadoensis/G. pseudomenardii subzone (P4c) Late Paleocene
-
(5)
Morozovella velascoensis zone (P5) Late Paleocene
-
(6)
P. wilcoxensis/M. velascoensis zone (E2) Early Eocene
In the Hawashiya section, the Se/Th boundary falls within the Tarawan Formation, between samples 116/117 at the top of Acarinina soldadoensis/Globanomalina pseudomenardii subzone (P4c), and in the Abu Had section, the Se/Th boundary falls within the Tarawan Formation, between samples 36/37 at the base of the Morozovella velascoensis biozone (P5).
The P/E boundary is detected in Esna Formation Hawashiya and Abu Had successions, between samples 123/124 and 40/41, respectively. Precisely, at the boundary between Morozovella velascoensis zone (P5) and Pseudohastigerina wilcoxensis/Morozovella velascoensis zone (E2). A detailed analysis of planktonic and benthonic foraminifera parameter (including the total number of foraminiferal species, species diversity, the P/B ratio, and the Aggl./Calc. ratio, Life strategies) and their abundance patterns is carried out. According to the statistical program SPSS analysis of benthonic foraminiferal species, the benthonic foraminiferal biofacies are divided into seven biofacies ranging in depth from shallow to deep. These are the following: Lenticulina biofacies, Lagena biofacies, Pullenia biofacies, Alabamina biofacies, Angulogavelinella biofacies, Loxostomoides biofacies, and Bulimina biofacies.
The results revealed that Dakhla Formation may be deposited in the inner to middle neritic environments, whereas the Tarawan Formation may be deposited in shallow to the middle neritic environment. The Esna Formation may be deposited in an outer neritic environment. Based on lithologic characters, detailed field examination for stratigraphic surfaces, and foraminiferal biostratigraphic studies, the studied Lower Paleogene successions in the Wadi Qena area are classified into five-order depositional sequences (DS) bounded by five sequence boundaries (SB). These are four of the Paleocene age and one of the Lower Eocene age. To show how the relative sea level fluctuation changed along the research region from north to south of Wadi Qena, a correlation was made between these five recorded depositional sequences and their sequence boundaries at the two analyzed sections. The recorded sequence boundaries in the study area correlate with those of previous studies both inside and outside Egypt.
Data Availability
The data were available at the Egyptian Petroleum Research Institute.
References
Abdel Razik TM (1972) Comparative studies on the Upper Cretaceous-Early Paleogene sediments on the Red Sea coast, Nile Valley and Western Desert, Egypt. 6th Arab Petroleum Congr. Algeria 71:1–23
Alegret L, Thomas E (2005) Cretaceous/Paleogene boundary bathyal paleo-environments in the central North Pacific (DSDP Site 465), the Northwestern Atlantic (ODP Site 1049), the Gulf of Mexico and the Tethys: the benthic foraminiferal record. Palaeogeogr Palaeoclimatol Palaeoecol 224:53–82
Aubry MP, Ouda K, Dupuis C, Berggren WA, Couvering JA (2007) The global standard stratotype-section and point (GSSP) for the base of the Eocene series in the Dababiya section (Egypt). Episodes 30(4):271–286
Aubry MP, Ouda K, Dupuis C, Van Couvering JA, Ali J, Berggren WA, Brinkhuis H, Gingerich PH, Heilman-Clausen C, Hooker J, Kenet DV, King C, Knox RWOB, Laga P, Molina E, Schmitz B, Steurbaut E,Ward DR (2002) Proposal: global standard stratotypesection and point (GSSP) at the Dababiya section (Egypt) for the base of the Eocene series. International Subcommission on the Paleogene Stratigraphy, Internal Report 58 pp
Awad G, Ghobrial M (1966) Zonal stratighraphy of Kharga Oasis. Geolog Surv Egypt, Cairo 34:1–77
Baldanza A, Bizzarri R, Famiani F, Garassino A, Hyžný M, Pasini G (2013) The bathyal decapod crustacean community from the Poggio i Sodi quarries (Siena Basin, Tuscany, Italy). Boletín De La Sociedad Geológica Mexicana 65(2):335–353
Berggren WA, Hilgen F, Langereis C, Kent DV, Obradovich J, Raffi I, Shackleton N (1995) Late Neogene chronology: new perspectives in high-resolution stratigraphy. Geol Soc Am Bull 107(11):1272–1287
Berggren WA, Ouda K (2003) Upper Paleocene-lower Eocene planktonic foraminiferal biostratigraphy of the Dababiya section, Upper Nile valley (Egypt). Micropaleontology 49:61–92
Berggren WA, Pearson PN (2005) A revised tropical to subtropical Paleogene planktonic foraminiferal zonation. J Foraminiferal Res 35(4):279–298
Berggren WA, Alegret L, Aubry M-P, Cramer BS, Dupuis C, Goolaerts S, Obaidalla N (2012) The Dababiya corehole, Upper Nile Valley, Egypt: preliminary results. The definitive version was published in Austrian. J Earth Sci 105(1):161–168
Bice KL, Norris RD (2002) Possible atmospheric CO2 extremes of the Middle Cretaceous (late Albian–Turonian). Paleoceanography and Paleoclimatology 17(4):1029–1050
Cande SC, Kent DV (1995) Revised calibration of the geomagnetic polarity timescale for the Late Cretaceous and Cenozoic. J Geophys Res: Solid Earth 100:6093–6095
Conoco C (1987) Geological map of Egypt, Scale 1: 500,000,-NH36SW-Beni Suef, Egypt. The Egyptian General Petroleum Corporation, Cairo (EGPC), Egypt
El Dawy M, Obaidalla NA, Mahfouz KH, Abdel Wahed SA (2016) Paleocene-Eocene transition at Naqb Assiut, Kharga Oasis, Western Desert, Egypt: stratigraphical and paleoenvironmental inferences. J Afr Earth Sc 117:207–222
El-Azabi M, El-Araby A (2000) Depositional cycles: an approach to the sequence stratigraphy of the Dakhla Formation, west Dakhla-Farafra stretch, Western Desert Egypt. J Afr Earth Sci 30(4):971–996
Ayyat El, Abdalla M, Obaidalla NA (2013) Stratigraphy, sedimentology and tectonic evolution of the Upper Cretaceous/Paleogene succession in north Eastern Desert Egypt. J Afr Earth Sci 81:35–59
El-Deeb WZ, Faris M, Mandur MM (2000) Foraminiferal and calcareous nannofossil biostratigraphy of the paleocene in north Central sinai area, Egypt. Annals Geol. Surv, Egypt, V.5:103-118
Faris M, El-Deeb WZ, Mandur MM (2000) Biostratigraphy of some Upper Cretaceous/Lower Eocene successions in Southwest Sinai Egypt. Ann Geol Surv Egypt 5:135–161
Gebhardt H, Ćorić S, Darga R, Briguglio A, Schenk B, Werner W, Sames B (2013) Middle to Late Eocene paleoenvironmental changes in a marine transgressive sequence from the northern Tethyan margin (Adelholzen, Germany). Austr J Earth Sci 106:45–72
Ghourchaei S, Ghasemi-Nejad E, Vahidinia M, Ashouri A (2015) Paleoenvironmental reconstruction of the Upper Cretaceous succession (Abtalkh Formation) of the Kopeh-Dagh Basin, northeastern Iran based on foraminiferal and palynological analyses. Arab J Geosci 8(4):2153–2168
Gradstein FM, Ogg JG, Schmitz MD, Ogg JM (2012) The geologic time scale Elsevier, 1: 1-1083
Guasti E, Speijer RP, Brinkhuis H, Smit J, Steurbaut E (2006) Paleoenvironmental change at the Danian-Selandian transition in Tunisia: foraminifera, organic-walled dinoflagellate cyst and calcareous nannofossil records. Mar Micropaleontol 59:210–229
Hamad MM (2009) Biostratigraphy of the Late Paleocene/Early Eocene deposits of Gebel El Bruk area, North-Central Sinai Egypt. Egypt J Paleontol 9:1–29
Haq BU, Hardenbol J, Vail PR (1988) Chronology of fluctuating sea levels since the Triassic. Science 235:1156–1167
Haq BU, Schutter SR (2008) A chronology of Paleozoic sea-level changes. Science 322:64–68
Hewaidy A, Farouk S, EL-Balkiemy A (2017) Foraminiferal biostratigraphy, stages boundaries and paleoecology of the uppermost Maastrichtian-Lower Eocene succession at Esh El-Mellaha area, North Eastern Desert, Egypt. J Am Sci, 13(5), 73-113
Hewaidy A, Farouk S, Mandur M, El Agroudy IS (2019) Planktonic foraminiferal and paleoenvironments of the Upper Campanian-Maastrichtian succession in Wadi Qena Egypt. Egypt J Petrol 28:47–59
Issawi B (1972) Review of Upper Cretaceous-Lower Tertiary stratigraphy in central and southern Egypt. AAPG Bull 56(8):1448–1463
Jennings AE, Walton ME, Ó Cofaigh C, Kilfeather A, Andrews JT, Ortiz JD, Dowdeswell JA (2014) Paleoenvironments during Younger Dryas‐Early Holocene retreat of the Greenland Ice Sheet from outer Disko Trough, central west Greenland. J Quat Sci, 29: 27-40
Kaiho K, Takeda K, Rose Petrizzo M, Zachos JC (2006) Anomalous shifts in tropical Pacific planktonic and benthic foraminiferal test size during the Paleocene-Eocene thermal maximum. Palaeogeogr Palaeoclimatol Palaeoecol 237:456–464
Kaiho K (1994) Benthic foraminiferal dissolved-oxygen index and dissolved-oxygen levels in the modern ocean. Geology 22:719–722
Keller G, Adatte T, Stinnesbeck W, Luciani V, Karoui-Yaakoub N, Zaghbib-Turki D (2002) b. Paleoecology of the Cretaceous-Tertiary mass extinction in planktonic foraminifera. Palaeogeogr Palaeoclimatol Palaeoecol 178:257–297
Kelly DC, Bralower TJ, Zachos JC, Premoli Silva I, Thomas E (1996) Rapid diversification of planktonic foraminifera in the tropical Pacific (ODP Site 865) during the late Paleocene thermal maximum. Geology 24:423–426
Krahl G, Koutsoukos EA, Fauth G (2017) Paleocene planktonic foraminifera from DSDP Site 356, South Atlantic: Paleoceanographic inferences. Mar Micropaleontol 135:1–14
Leckie RM, Olson HC (2003) Foraminifera as proxies of sea-level change on siliciclastic margins, in Micro Paleontologic proxies for sea level change and stratigraphic discontinuities, edited by H. C. Olson and R. M. Leckie, Spec. Publ. SEPM Society for Sedimentary Geology, 17:5 – 19
Luterbacher HP, Ali JR, Brinkhuis H, Gradstein FM, Hooker JJ, Monechi S, Ogg JG, Powell J, Rohl U, Sanfilippo A, Schmitz B (2004) The Paleogene period, in A geologic time scale, F. Gradstein, J. Ogg, and A. Smith (eds.), Cambridge University Press. Cambridge, pp. 384–408
Mandur MM )2016( Ypresian to Priabonian nannofossil biostratigraphy andPalaeoecology, Gulf of Suez, Egypt, J Palaeontol Soc India 61(1):19–28
Mandur MM, Bayoumi AH (2010) Calcareous nannofossil of the Upper Cretaceous /Lower Eocene succession in the Southwestern Sinai Area Egypt. World J Earth Sci 1(1):28–37
Mandur MM, Hewaidy AGA, Farouk S, El Agroudy IS (2022) Implications of calcareous nannofossil biostratigraphy, biochronology, paleoecology, and sequence stratigraphy of the Paleocene-Eocene of the Wadi Qena Egypt. J Afr Earth Sci 139:1–19
Nouradini M, Azami SH, Hamad M, Yazdi M, Ashouri AR (2015) Foraminiferal paleoecology and paleoenvironmental reconstructions of the lower Miocene deposits of the Qom Formation in Northeastern Isfahan, Central Iran. Boletín De La Sociedad Geológica Mexicana 67:59–73
Olsson RK, Nyong EE (1984) A paleoslope model for Campanian-lower Maestrichtian foraminifera of New Jersey and Delaware. J Foraminiferal Res 14(1):50–68
Petrizzo MR (2002) Palaeoceanographic and palaeoclimatic inferences from Late Cretaceous planktonic foraminiferal assemblages from the Exmouth Plateau (ODP Sites 762 and 763, eastern Indian Ocean). Mar Micropaleontol 45(2):117–150
Said R (1960) Planktonic foraminifera from the Thebes formation, Luxor Egypt. Micropaleontology 6(3):277–286
Said R (1961) Tectonic framework of Egypt and its influence on distribution of foraminifera. AAPG Bull 45(2):198–218
Saint-Marc P (1992) Biogeographic and bathymetric distribution of benthic foraminifera in Paleocene El Haria Formation of Tunisia. J Afr Earth Sc 15(3–4):473–487
Samir A (2002) Biostratigraphy and paleoenvironmental changes in the upper Cretaceous-Early Paleogene deposits of Gabal Samara section, Southwestern Sinai Egypt. Egypt J Paleontol 2:1–40
Schmitz B, Pujalte V, Eustoquio M, Monechi S, Orue-Etxebarria X, Speijer RP, Aubry MP (2011) The global stratotype sections and points for the bases of the Selandian (Middle Paleocene) and Thanetian (Upper Paleocene) stages at Zumaia, Spain, 4. Intl Union Geol Sci: Ottawa 34:220–245
Silva IP, Sliter WV (1999) Cretaceous paleoceanography: evidence from planktonic foraminiferal evolution. Geol Soc Am 332:301–328
Sprong J, Kouwenhoven TJ, Bornemann A, Schulte P, Stassen P, Steurbaut E, Speijer RP (2012) Characterization of the Latest Danian Event by means of benthic foraminiferal assemblages along a depth transect at the southern Tethyan margin (Nile Basin, Egypt). Mar Micropaleontol 86:15–31
Székely SF, Bindiu-Haitonic R, Filipescu S, Bercea R (2017) Biostratigraphy and paleoenvironmental reconstruction of the marine lower Miocene Chechiș Formation in the Transylvanian Basin based on foraminiferal assemblage. Carnets Geol 17:11–39
Tantawy A (2003) Calcareous nannofossil biostratigraphy and paleoecology of the Cretaceous-Tertiary transition in the central Eastern Desert of Egypt. Mar Micropaleontol 47:323–356
Tantawy A, Keller G, Adatte T, Stinnesbeck W, Kassab A, Schulte P (2001) Maastrichtian to Paleocene depositional environment of the Dakhla Formation, Western Desert, Egypt: sedimentology, mineralogy, and integrated micro-and macrofossil biostratigraphies. Cretac Res 22(6):795–827
Van der Zwaan G, Jorissen F, De Stigter H (1990) The depth dependency of planktonic/benthic foraminiferal ratios: constraints and applications. Mar Geol 95(1):1–16
Wade BS, Pearson PN, Berggren WA, Pälike H (2011) Review and revision of Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the geomagnetic polarity and astronomical time scale. Earth Sci Rev 104(1–3):111–142
Zachos JC, Rohl U, Schellenberg SA, Sluijs A, Hodell DA, Kelly DC, Thomas E, Nicolo M, Raffi I, Lourens L, McCarren H, Kroon D (2005) Rapid acidification of the ocean during the Paleocene- Eocene Thermal Maximum. Science 308:1611–1615
Acknowledgements
Grateful thanks are due to Editors-in-Chief Prof. Dr. Abdullah M. Al-Amri and the anonymous reviewers for their valuable scientific comments, which have greatly improved the original manuscript.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Responsible Editor: Attila Ciner
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mandur, M.M.M., Hewaidy, A.G.A., Farouk, S. et al. An approach to using foraminifera in sequence stratigraphic analysis of Wadi Qena, Central Eastern Desert, Egypt. Arab J Geosci 16, 190 (2023). https://doi.org/10.1007/s12517-023-11226-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-023-11226-2