
Abstract
Micropaleontological investigations on Permian successions in the NW of Iran based on algae led to the determination of taxa belonging to some families such as Ungdarellaceae (Ungdarella uralica), Gymnocodiaceae (Gymnocodium bellerophontis and Permocalculus sp.), Dasycladales (Mizzia cf. M. yabei), and some microproblematics (Vermiporella nipponica and Pseudovermiporella sodalica). These investigations are mainly focused on the Khachik Formation at the Ali Bashi Mountains. According to the algal community, a lagoon environment is offered for the studied successions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Fossil-rich Late Permian sections in the central Tethys, especially in Transcaucasia, Northwest and Central Iran, have been known for a long time (Abich 1878; Frech and Arthaber 1900; Stoyanow 1910; Ruzhentsev and Sarycheva 1965; Rostovtsev and Azaryan 1973; Kozur et al. 1980; Kotlyar et al. 1983; Stepanov et al. 1969; Teichert et al. 1973; Kozur 2005, 2007). They are among the best-known localities in which the dramatic faunal change of pelagic organisms between the Palaeozoic and Mesozoic can be studied.
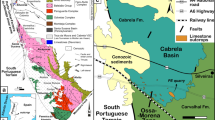
Permian–Triassic succession in the Ali Bashi Mountains, 9 km west of Julfa city, North West of Iran (Fig. 1), was discovered in the 1960s and described by Stepanov et al. (1969). This succession at Julfa area closely resembles that of Dorasham, which is located only 8 km toward the north. The Ali Bashi Mountains comprise one of the best exposures of the important Permian–Triassic boundary (PTB) interval worldwide and work as a standard for the transitional beds in the Tethyan realm. These exposures are known for less than 50 years but became extremely important for the study of the most severe extinction event in the Phanerozoic (Kozur 2005, 2007; Ghaderi et al. 2013, 2014a; Leda et al. 2014).

Location map and geological map of the study area. The star shows the location of the sections of the Ali Bashi Mountains, Julfa, NW Iran
In spite of this, the most famous studies were focused on lithostratigraphy, conodont biostratigraphy, event, and isotope stratigraphy around the PTB interval (e.g., Korte et al. 2004; Korte and Kozur 2005; Kozur 2004, 2005; Leda et al. 2014), and other older fossiliferous deposits, especially algae-rich Wuchiapingian (Dzhulfian) part of the sections, have been neglected. In this study, we will focus on the lagoonal carbonate sedimentary rocks to identify and systematically describe the Wuchiapingian algal flora and compare them with other equivalent materials from the region of Transcaucasica and Central Iran platform.
Geological and stratigraphical setting
Through the Late Palaeozoic, several Gondwanan terrains, including Sanandaj-Sirjan terrain, Alborz, and Central Iran, broke off the eastern Gondwanan margin due to the Permian Neotethyan opening (Stampfli and Borel 2002, 2004; Angiolini and Carabelli 2010) and moved together with other Cimmerian blocks to the north toward the equator (Sengor 1979; Stampfli et al. 1991, Stampfli and Borel 2002). Collision with the Eurasian active margin was the result of this movement in the Late Triassic. The studied sections here in Julfa region had a palaeogeographic position in the central Tethys near the equatorial humid belt in Late Permian and were situated on the NNW part of the Sanandaj–Sirjan Terrane of the Cimmerian microcontinent (Scotese and Langford 1995; Golonka 2000; Stampfli and Borel 2002, 2004; Torsvik and Cocks 2004).
Sedimentary successions of the Permian–Triassic, totally 1120.5 m thick, crop out in Ali Bashi Mountains, west of Julfa city. The base of Permian successions is characterized by a major discontinuity with Devonian–Carboniferous? volcano-sedimentary rocks. Permian deposits begin with 110-m red siliciclastic unit at the base, cropped at southwest of Julfa, equivalent to Doroud Formation (Assereto 1963) in Alborz Mountains, North of Iran. This succession is unconformably covered by marine carbonate sequences of Middle Permian–Triassic, which were discussed in the Stepanov et al. (1969) report. They recognized eight rock units (A–H) totally 1010.5 m thick from base to top (Table 1). Their units A (Genishik beds) and B (Khachik beds) belonged to Guadalupian and units C (Lower Julfa beds) and D (Upper Julfa beds) and lower part of E (transition beds) to Dzhulfian. They named unit F, the grayish red limestone unit which is about 3.6 m thick and contained Paratirolites, as the Paratirolites limestone and defined that as Induan. Units G and H were considered as equal to Induan Elikah Formation (Glaus 1964) in Alborz Range, north of Iran, and contain the Triassic vermiculate limestone full of bivalve subgenus Claraia.
Teichert et al. (1973) have described four sections at localities 1 to 4, about 500 m apart, and proposed the name Ali Bashi Formation instead of the units E and F of Stepanov et al. (1969). Ali Bashi 1 section according to Teichert et al. (1973) can serve as a standard for all lithostratigraphic PTB sections in NW Iran.
Partoazar (2002) revisited the section at Ali Bashi Mountains and resampled for studying the foraminiferal contents. In his revision, he changed the name of Gnishik and Khachik beds to new Julfa Formation; however, his compilation was not followed by other geologists. Also, he fairly revised the age of Khachik beds from Guadalupian to Dzhulfian (Wuchiapingian).
The term of “Khachik beds” has been used as “Khachik Formation” in Armenia (e.g. Kotlyar et al. 1983) and also in some reports from the Ali Bashi Mountains of Iran (e.g. Kozur 2005). In the present paper, we will follow them in usage of the term Khachik Formation.
In this research, we will discuss and describe the algal flora of the Khachik Formation (Khachik beds sensu Stepanov et al. 1969 and Shamar Member sensu Partoazar 2002) on the classic measured section by Stepanov et al. (1969) and the base of a new parallel section, the Main Valley section, which both are located at the Kuh-e-Ali Bashi (= Ali Bashi Mountains), 8 km west of Julfa (respectively in 38° 56′ 5.74″ N, 45° 31′ 20.53″ E and 38° 56′ 22.56″ N, 45° 30′ 40.94″ E). We grade the Khachik Formation into nine informal units (Fig. 2), in ascending order as follows:
-
Unit I:
Ungdarella limestone: Capitanian in age; 30-m thickness, medium- to thick-bedded limestone (bioclastic wackestone/packstone) in gray to yellow which conformably overlaid dark-gray limestone succession of Gnishik beds. The macrofauna consists of brachiopod and bryozoan debris, corals, crinoids, and bivalves. Ostracods, foraminifers, and some algae such as Ungdarella uralica and Gymnocodium sp. are the most important microfossils of this interval. The nominal species U. uralica is very common algae throughout the unit, and we named this interval based on its abundance.
-
Unit II:
Capitanian in age; 27.75-m thickness, dark- to light-gray shale with some intercalations of light-gray thin- to medium-bedded limestone (bioclastic wackestone/packstone). The macrofauna consists of crinoids and bivalves. Small foraminifers (abundant miliolids), ostracods, and some Gymnocodiacean algae are the microfossils of this unit.
-
Unit III:
Capitanian in age; 8.80-m thickness; light-gray medium-bedded limestone (bioclastic packstone) with few brachiopods; and small foraminifers (abundant miliolids), ostracods, and some algae and microproblematics such as Vermiporella sp.
-
Unit IV:
Capitanian in age; 15.2-m thickness, light-gray shale with many intercalations of light-gray thin- to medium-bedded limestone (bioclastic wackestone to compact bioclastic packstone). Abundant miliolids, rare fusulinids, Gymnocodium sp., Vermiporella sp., and Vermiporella nipponica are present at this unit.
-
Unit V:
Capitanian in age; 10.8-m thickness, gray thin- to thick-bedded limestone (bioclastic packstone). Bryozoans, ostracods, abundant miliolids, and algae such as Gymnocodium sp. are the most important fossils of this unit.
-
Unit VI:
Capitanian in age; 9.40-m thickness, light to dark-gray thin- to thick-bedded limestone (fossiliferous mudstone, bioclastic wackestone to bioclastic packstone), with black cherty nodules in upper beds and some gray to black shale intervals. Corals, belerophontid gastropods, miliolids, fusulinids, small lagenids, and algae and microproblematics such as V. nipponica, Vermiporella sp., and Mizzia sp. are the fossil content of this unit.
-
Unit VII:
Capitanian in age; 6.90-m thickness, yellowish thin to medium nodular limestone and very fine seams of shale and gypsum at the basal part, thick-bedded limestone at the middle part (fossiliferous mudstone to bioclastic wackestone), and a diabasic sill at the upper part. Bivalves, corals, goniatids, brachiopods, bryozoans, holothurian sclerites, crinoids, fusulinids, some algae such as gymnocodiacean debris, and Scolicia isp. are present in this unit.
-
Unit VIII:
Capitanian?–Wuchiapingian in age; 51.05-m thickness, thick-bedded cliff-forming limestone (mudstone, bioclastic wackestone, packstone, and Gymnocodium floatstone), dark gray in color (superficially red due to weathering), with some intercalations of gray to black shale, black cherty stripes at the lower parts, and cherty nodules at the middle and upper parts of the unit. Crinoids, holothurian sclerites, small lagenids and miliolids, ostracods, some algae, and microproblematics such as Gymnocodium bellerophontis, Gymnocodium sp., Mizzia sp., Permocalculus sp., Pseudovermiporella sodalica, V. nipponica, and Vermiporella sp. are present in this unit.
-
Unit IX:
Codonofusiella limestone: Wuchiapingian in age; 4.70-m thickness, gray, green to yellow medium- to thick-bedded limestone (mudstone, bioclastic wackestone, packstone, and Gymnocodium floatstone), without any cherty stripes or nodules. Codonofusiella limestone that is the uppermost unit of the Khachik Formation (Fig. 2) includes rich monogenetic fusulinid content full of Codonofusiella, especially Codonofusiella kwangsiana (Ghaderi et al. 2014b). Other foraminiferal contents of this unit are Agathammina sp.; Codonofusiella nana; Climacammina sp.; Frondina ex gr. permica; Frondina sp.; Globivalvulina cf. vonderschmitti; Glomomidiella nestellorum; Hemigordius spp.; Nankinella sp.; Palaeotextularia sp.; Paraglobivalvulina gracilis; Pseudobaisalina sp. bivalves; ostracods; some algae such as G. bellerophontis, Gymnocodium sp., Permocalculus sp., and V. nipponica are the other fossil contents of this unit.

Stratigraphic log of the Ali Bashi Section (sensu Stepanov et al. 1969) and the Codonofusiella limestone in Main Valley section, showing the range of the studied algae. Lower right: photo of the Ali Bashi Section pointing to NE
Codonofusiella limestone in the Main Valley section is 4.82 m thick with the same lithology and microfacies, full of G. bellerophontis, Permocalculus sp., and very rare V. nipponica.
The Julfa Formation (Julfa Beds sensu Stepanov et al. 1969), which is 35 m thick and Wuchiapingian in age, consists of gray, green to red shales with nodular limestone (bioclastic wackestone and ostracod- and crinoid-rich lime mudstone) and marlstone intercalations. This formation conformably overlays dark-gray Codonofusiella limestone. Julfa Formation macrofauna comprises brachiopods (particularly common at the lower part of the formation), ammonoids (Araxoceras in the lower part, Vedioceras in the upper part of the formation), nautiloids, rugose, and tabulate corals, etc.
The studied algal flora in this research has been obtained from the Khachik Formation, which ranges from the base of unit I (Ungdarella limestone) to the base of Lower Julfa beds (on top of Codonofusiella limestone). Abundance and diversification of algal contents abruptly decrease in the Lower Julfa beds through the Ali Bashi and Elikah Formation, which are full of open marine fossils. Algal reduction over two later formations is also associated with continuous gradual facies changes from bioclastic wackestone/packstone to lime mudstone and shale and confirms the deepening upward trend in the Uppermost Permian deposits of the Ali Bashi Mountains. However, the Julfa Formation records an overall deepening trend reaching outer ramp conditions, punctuated by several cycles.
Age and correlation
The related interval to the Khachik Formation is attributed to Wuchiapingian by Partoazar (2002). This contrasts with the age provided by Stepanov et al. (1969), which had considered the age of Khachik beds as Guadalupian. Jin et al. (2006) defined the base of Wuchiapingian by the first appearance of conodont subspecies Clarkina postbitteri postbitteri. This horizon is equal to the base of Codonofusiella–Reichelina Zone, which contains Post Extinction fauna and flora and is accentuated by Isozaki et al. (2007) and Saitoh et al. (2013). This new definition is in contrast with the previous age dating on the Kuh-e-Ali Bashi section by Stepanov et al. (1969) and Partoazar (2002).
The Codonofusiella-rich beds, which belong to the first foraminiferal biozone of the Wuchiapingian, are the last rock unit of the Khachik Formation. Thus, the uppermost part of the Khachik Formation certainly belongs to the Wuchiapingian; however, Stepanov et al. (1969) had considered Codonofusiella limestone as the uppermost Guadalupian. Partoazar (2002) had attributed this part properly to Wuchiapingian, but he had considered all of the preceding units as Wuchiapingian, too. This has disparity with the definition of the Lopingian base of Jin et al. (2006).
The abundance of frondose bryozoans, alatoconchid bivalves, and last large fusulinids in the carbonate successions of the unit VII is very similar to preextinction communities of the Late Capitanian (Wood 1998; Kofukuda et al. 2014). The first appearance of Clarkina postbitteri postbitteri is not recorded yet in the section. So, recognition of the Guadalupian–Lopingian is not precise, but if we consider the mentioned paleocommunities as Late Capitanian and the Codonofusiella assemblages as the index of Wuchiapingian base, the stage boundary will consider between the units VII and IX. Reduction of abundance and diversity of different fossil groups such as bivalves, brachiopods, fusulinids and growing of small foraminifers (as Lilliput Fauna after End Guadalupian Extinction) in the Unit VIII suggest that the Guadalupian–Lopingian Boundary is at the base of this unit or at least somewhere in this unit.
Systematic paleontology
One taxon belongs to Ungdarellaceae (U. uralica).
Two taxa belong to Gymnocodiaceae (G. bellerophontis and Permocalculus sp.)
One taxon belongs to Dasycladales (Mizzia cf. M. yabei).
Two taxa belong to microproblematica (Vermiporella nipponica and P. sodalica).
Abbreviations in descriptions: length of thallus (lt), diameter of thallus (dt), diameter of pores (dp), diameter of sporangia (ds), diameter of hollow (dh), outer diameter (ot), width of cells (wc), height of cell laminae (hcl), wall thickness (wt), number of branches (nb), and diameter of branches (db).
Family Ungdarellaceae Maslov 1956
Ungdarella (Maslov 1950) Maslov, 1956a
U. uralica Maslov, 1956a
Plate 1a, f, j

Scale bars equal to 200 μm. Ungdarella uralica Maslov, 1956a. a Longitudinal. section, FUM#J107.1; Khachik Formation, unit I, Ali Bashi Section. f Equatorial section, FUM#J107.3; Khachik Formation, unit I, Ali Bashi Section. j Longitudinal section, FUM#J108.3; Khachik Formation, unit I, Ali Bashi Section. Gymnocodium bellerophontis (Rothpletz) Accordi, 1956. b Longitudinal section, FUM#J155.1; Khachik Formation, unit VIII, Ali Bashi Section. c Oblique section, FUM#G111.14; Khachik Formation, unit IX, Main Valley section. g Transverse section, FUM#J160M.7; Khachik Formation, unit IX, Ali Bashi Section. h Transverse section, FUM#G119.8; Khachik Formation, unit IX, Main Valley section. i Oblique section, FUM#G111.35; Khachik Formation, unit IX, Main Valley section. Gymnocodium sp. d Longitudinal section, FUM#J112.1; Khachik Formation, unit I, Ali Bashi Section. e Longitudinal section, FUM#G118.5; Khachik Formation, unit IX, Main Valley section. Vermiporella nipponica (Endo in Endo etKanuma, 1954). k Longitudinal section, FUM#G120.42; Khachik Formation, unit IX, Main Valley section. l Transverse section, FUM#G120.47; Khachik Formation, unit IX, Main Valley section. Pseudovermiporella sodalica Elliott, 1958. m Tangential section, FUM#J150.8; Khachik Formation, unit VIII, Ali Bashi Section
Remarks: According to the Parvizi et al. (2013), the thallus is rod shaped, branched, with an apparently cellular construction. The thallus is almost parallel. Due to the generally strong recrystallization, only yellow or cream-colored indistinct bodies are seen. There is no distinct difference between hypotallus and peritallus in this taxon but the perithallus with long thin rows of subquadratic cells (Flugel 2004). This microorganism, occasionally assigned to rare algosponges (Parvizi et al. 2013), but is defined as red algae or rhodophyta accompanies with some taxon such as Permocalculus, Komia, and Gymnocodium (Shabanian et al. 2008) with a hypothallus and a perithallus (Mamet 1991), but the skeletal network of Ungdarella is generally sharply recrystallized, and when it is well preserved, it appears as perforated and sometimes filled by micrite so it differs from completely close cells of red algae (Vachard and Aretz 2004). Also, it is defined as microproblematic by Chuvashov et al. 1987 and pseudoalgae by Flugel, 2004
Dimensions: lt = 0.917–1.09 mm, ot = 0.052–0.074 mm, wc = 0.009–0.023 mm, and hcl = 0.014–0.028 mm.
Age: Late Visean (Vachard and Tahiri 1991; Gallagher 1996) to Late Permian (Vachard 1980), Murgabian (Middle Permian) (Parvizi et al. 2013), Midian, and Early Dzhulfian (Shabanian et al. 2008).
Occurrence: Carnic Alps, Russia, Turkey, Armenia, Iraq, Afghanistan, Himalaya, and Iran (e.g., Kolodka et al. 2012; Parvizi et al. 2013; Shabanian et al. 2008; Ghahramani et al. 2009). Ungdarella is cosmopolite (Mamet 1991).
Environment: Lagoon (Shabanian et al. 2008). Common in Middle Carboniferous shelf carbonates (Flugel 2004).
Family Gymnocodiaceae Elliot, 1955
Permocalculus Elliott, 1955
Permocalculus spp.
Plate 2d–k

Scale bars equal to 200 μm. Vermiporella nipponica (Endo in Endo etKanuma, 1954). a Longitudinal section, FUM#J150.10; Khachik Formation, unit VIII, Ali Bashi Section. b Transverse section, FUM#J150.9; Khachik Formation, unit VIII, Ali Bashi Section. Mizzia ? sp. c Transverse section, FUM#J133.3; Khachik Formation, unit VI, Ali Bashi Section. Permocalculus spp. d Transverse sections, some partly oblique, FUM#J153.6; Khachik Formation, unit VIII, Ali Bashi Section. e Transverse section, FUM#G116.23; Khachik Formation, unit IX, Main Valley section. f Longitudinal sections, FUM#G124.2; Khachik Formation, unit IX, Main Valley section. g Slab section, different sections, FUM#G124.1; Khachik Formation, unit IX, Main Valley section. h Tangential section, FUM#G124.3; Khachik Formation, unit IX, Main Valley section. i Tangential-oblique section, FUM#J160I.7; Khachik Formation, unit IX, Ali Bashi Section. j SEM photograph, FUM#G120.3; Khachik Formation, unit IX, Main Valley section. k SEM photograph, FUM#G120.3; Khachik Formation, unit IX, Main Valley section
Description: Thallus irregular, segmented. Segments of variable form: spherical, ovoid or barrel shaped, or elongated, irregularly finger-like, or with pinching and swelling units. Calcification varying from very thin to massive or solid. Pores small and cortical. Sporangia cortical or medullary.
Remarks: Very similar to Gymnocodium but commonly having larger units or segments, with finer (smaller) pores and more irregular calcification. This taxon is characterized by tightly bound segments with a wide, poorly calcified medulla and a narrow cortex intersected by numerous branched pores (Flugel 2004). Algal forms tentatively referred to Permocalculus are recorded by Golestaneh (1966) and Strong (1939) from ?Oligocene–Early Miocene rocks of South Iran.
Dimensions: lt = 0.765 mm, dt = 0.371–0.793 mm, wt = 0.107–0.215 mm, dp = 0.020–0.01 mm, and dh = 0.169–0.48 mm.
Age: Late Permian to Cretaceous (and Miocene). Permian and Cretaceous (Flugel 2004).
Occurrence: Texas, Iraq, Armenia, Turkey, the Carnic Alps, Saudi Arabia, Tunisia, Pakistan, Austria, and Iran (e.g., Okla 1992; Parvizi et al. 2013; Shabanian et al. 2008; Ghahramani et al. 2009; Schlagintweit 2010).
Environment: Lagoon (Shabanian et al. 2008), back reef facies (Flugel 2004).
Gymnocodium (Pin 1920) Elliott, 1955
G. bellerophontis (Rothpletz) Accordi, 1956
Plate 1b–c, g–i
Remarks: The wall is aragonitic in this taxon like as the other members of this family (Gymnocodiaceae) and calcified because of the digenetic process (Flugel 2004). There is regularly cylindrical thallus, which is weakly curved, with the files of cells smaller and less marked through the wall (Parvizi et al. 2013). Hollow calcareous segments, which are cylindrical, oval or cone shaped, and circular or oval in cross section, rarely bifurcating. The walls are perforated by pores, which radiate oblique distally and widen markedly outward. The sporangia are ovoid in terminal segments (Okla 1992). The cortex zone, where the pores are located, can be observed. In longitudinal sections, it is exhibiting the characteristic irregular outline of the thalli caused by thick peripheral filaments (Flugel 2004).
Dimensions: lt = 0.42–0.998 mm, dt = 0.011–0.217 mm, wt = 0.004–0.036 mm, dp = 0.017–0.025 mm, ds = 0.036–0.055 mm, and dh = 0.007–0.163 mm.
Age: Middle-Late Permian.
Occurrence: Slovenia, Serbia, Bosnia, Montenegro, Italy, Austria, Greece, Hungary, Tunisia, Urals, Turkey, Caucasus, Saudi Arabia, Oman, Iraq, Armenia, Iran, Pakistan, Afghanistan, Uzbekistan, Tibet, South China, Thailand, Malaysia, Japan, New Zealand, and ? Guatemala (e.g., Okla 1992; Lys et al. 1978; Partoazar 1995; Vaziri et al. 2005; Kolodka et al. 2012; Parvizi et al. 2012; Shabanian et al. 2008; Ghahramani et al. 2009; Schlagintweit 2010).
Environment: Lagoon (Shabanian et al. 2008), back reef facies (Flugel 2004).
Family Dasycladaceae Kutzing 1843
Mizzia (Schubert 1907) Rezak, 1959
Mizzia? sp.
Plate 2c
Remarks: Calcareous little bodies elongate elliptical, about twice longer than broad, with the largest diameter near the upper part of the member. Laterals non-ramified. Each member is clearly constricted at the base (Bilgutay 1959). Thallus is pear shaped or cylindrical in longitudinal view; branches are simple and without division, radially attached to the hollow zone.
Dimensions: dt = 0.401–0.171 mm, wt = 0.045–0.067 mm, dh = 0.084–0.238 mm, nb = 23–24 mm, and db = 0.042–0.43 mm.
Age: Permian (especially Murgabian and Midian), Late Permian (Bilgutay 1959).
Occurrence: Japan, South China, Guatemala, southwestern USA, Thailand, Malaysia, Pakistan, Afghanistan, Iran, Saudi Arabia, Tunisia, Turkey, Greece, Carnic Alps, Hungary, Slovenia, Croatia, and Montenegro (e.g., Bilgutay 1959; Bozorgnia 1973; Kolodka et al. 2012; Parvizi et al. 2013).
Environment: Lagoon (Shabanian et al. 2008).
Class Ulvophyceae
Vermiporella Stolley, 1883
V. nipponica (Endo in Endo etKanuma, 1954)
Remarks: Moderately thick wall; pits straight, of small diameter (Parvizi et al. 2013). Thallus is branched and irregular, central hollow is compressed, and pores are not connected to the central hollow from inside and just from one side are opened to the outside and can be observed in light and the wall in dark color.
Dimensions: lt = 0.061–0.438 mm, wt = 0.004–0.016 mm, dp = 0.003–0.008 mm, dh = 0.007–0.046 mm, and od = 0.014–0.075 mm.
Age: Cosmopolitan from the Sakmarian (Vachard and Krainer 2001; Krainer et al. 2003, 2009) to the Changsinghian, Murgabian (Parvizi et al. 2013).
Occurrence: Iran, Carnic Alps, and New Mexico (e.g., Vachard and Krainer 2001; Krainer et al. 2003, 2009; Gaillot 2006, unpublished; Gaillot and Vachard 2007; Ghahramani et al. 2009; Parvizi et al. 2013).
Environment: Lagoon and open marine (Shabanian et al. 2008). Lagoon facies (Dolenec et al. 2004).
Pseudovermiporella Elliott, 1958
P. sodalica Elliott, 1958
Plate 1m
Remarks: This taxon is characterized by attached tubes with perforated walls (Flugel 2004). This species includes some tubular skeletons with circular, oval, or irregular transverse sections. Pits are perpendicular to the central chamber and are polygonal or rounded in tangential sections (Parvizi et al. 2013). This taxon is significantly larger than V. nipponica (Vachard and Krainer 2001).
Dimensions: dt = 0.455 mm, wt = 0.033 mm, dp = 0.2 mm, and dh = 0.131 mm.
Age: Middle-Late Permian (Parvizi et al. 2013), Murgabian (Ghahramani et al. 2009), and Late Permian (Kalantari 1986).
Occurrence: Iraq, Oman, Afghanistan, Turkey, Hungary, Greece, Slovenia, Croatia, Armenia, North America, Tunisia, China, and Iran (e.g., Kolodka et al. 2012; Ghahramani et al. 2009; Parvizi et al. 2013).
Environment: Lagoon and open marine (Shabanian et al. 2008).
References
Abich H, 1878. Geologische Forschungen in den Kaukasischen Landern, I: Eine Bergkalkfauna aus der Araxesenge bei Djoulfa in Armenien. Holder Wien 128 S
Angiolini L, Carabelli L (2010) Upper Permian brachiopods from the Nesen Formation, North Iran. Palaeontology, 84:41–90
Assereto R (1963) The Paleozoic formations in Central Elburz (Iran) (preliminary note). Riv Ital Paleont Strat 69:503–543
Bilgutay U (1959) The Permian calcareous algae from southeastern Anatolia. Bulletin of the Mineral Research and Exploration Institute of Turkey, Ankara 52:48–58
Dolenec T, Ogorelec B, Dolenec M, Lojen S (2004) Carbon isotope variability and sedimentology of the upper Permian carbonate rocks and changes across the Permian-Triassic boundary in the Masore section (Western Slovenia). Facies 50:287–299
Flugel E (2004) Microfacies of Carbonate Rocks, Analysis Interpretation and Application. Springer-Verlage, Berline, Heidelberg. p. 976
Frech F, Arthaber GV (1900) Über das Paläozoicum in Hocharmenien und Persien. Mit einem Anhang über die Kreide von Sirab in Persien. Beiträge zur Paläontologie und Geologie von Österreich-Ungarn und des Orients 12:161–308
Gallagher SJ (1996) The stratigraphy and cyclicity of the Late Dinantian platform carbonates in parts of southern and western Ireland. In: Strogen P, Somerville ID, Jones GL (eds) Recent advances in Lower Carboniferous geology. Geological Society London, Special Publication, 107:239–251
Ghaderi A, Ashouri AR, Korn D, Mahmoudi Gharaie MH, Leda L (2013) New insight on stratigraphic correlation of the Permian-Triassic transitional beds in the Transcaucasus and northwest of Iran: problems and guidelines. Sedimentary Facies 5(2):221–224 (In Farsi)
Ghaderi A, Leda L, Schobben M, Korn D, Ashouri AR (2014a) High-resolution stratigraphy of the Changhsingian (Late Permian) successions of NW Iran and the Transcaucasus based on lithological features, conodonts and ammonoids. Fossil record 17:41–57
Ghaderi A, Garbelli C, Angiolini L, Ashouri AR, Korn D, Rettori R (2014b) Faunal changes near the end Permian extinction: the brachiopods of the Ali Bashi Mountains, NW Iran. Riv Ital Paleontol Stratigr 120(1):27–59
Ghahramani H, Shabanian R, Sartipzadeh S (2009) Introduction to Permian deposits calcareous algae in Qareh Ziodin area. 4th National Geological Congress of Payam-e-Noor University, pp. 43--48 [In Persian].
Glaus M (1964) Trias und Oberperm in zentralen Elburs (Persien). Eclogae Geol Helv 57(2):497–508
Golonka J (2000) Cambrian-Neogene plate tectonic maps. Wydawnictwa Uniwersytetu Jagiellońskiego, Kraków, pp. 1–125
Isozaki Y, Shimizu N, Yao J, Ji Z, Matsuda T (2007) End-Permian extinction and volcanism-induced environmental stress: the Permian–Triassic boundary interval of lower-slope facies at Chaotian, South China. Palaeogeogr Palaeoclimatol Palaeoecol 252:218–238
Jin Y, Shen S, Henderson CM, Wang X, Wang W, Wang Y, Cao C, Shang Q (2006) The Global Stratotype Section and Point (GSSP) for the boundary between the Capitanian and Wuchiapingian stage (Permian). Episodes 29(4):253–262
Kalantari A, 1986 Microfacies of carbonate rocks of Iran. Ministry of Oil, National Iranian Oil Company, Tehran, 287 p
Kofukuda D, Isozaki Y, Igo H (2014) A remarkable sea-level drop and relevant biotic responses across the Guadalupian–Lopingian (Permian) boundary in low-latitude mid-Panthalassa: irreversible changes recorded in accreted paleo-atoll limestones in Akasaka and Ishiyama, Japan. J Asian Earth Sci 82:47–65
Kolodka C, Vennin E, Vachard D, Trocme V, Goodarzi M (2012) Timing and progression of the end-Guadalupian crisis in the Fars province (Dalan Formation, Kuh-e-Gakhum, Iran) constrained by foraminifers and other carbonate microfossils. Facies, 58:131–153
Korte C, Kozur HW, Partoazar H (2004) Negative carbon isotope excursion at the Permian/Triassic boundary section at Zal, NW Iran. Hallesches Jahrbuch für Geowissenschaften, Reihe B, Beiheft, 18:69--71
Kotlyar GV, Zakharov YD, Koczyrkevicz BV, Kropatcheva GS, Rostovcev KO, Chedija IO, Vuks GP, Guseva EA, 1983 fPozdnepermskiy etap evolyutsii organicheskogo mira. Dzulficheskiy i dorashamskiy yarusy SSSR. In: Gramm MN, Rostovcev KO, (Eds) Permo-Triasovaya stadiya geologicheskoy evolyutsii. mezhdunarodnoy programmy geologicheskoy korrelatsii, 106: 1–70
Kozur HW (2004) Pelagic uppermost Permian and the Permian–Triassic boundary conodonts of Iran, part I: taxonomy. Hallesches Jahrbuch für Geowissenschaften B, Beiheft 18:39–68
Kozur HW (2005) Pelagic uppermost Permian and the Permian–Triassic boundary conodonts of Iran. Part II: investigated sections and evaluation of the conodont faunas. Hallesches Jahrbuch für Geowissenschaften B, Beiheft 19:49–86
Kozur H, Leven EY, Lozovskiy VR, Pyatakova MV (1980) Subdivision of Permian-Triassic boundary beds in Transcaucasia on the basis of conodonts. Int Geol Rev 22(3):361–368
Krainer K, Vachard D, Lucas SG (2003) Microfacies and microfossil assemblages (smaller foraminifers, algae, pseudo-algae) of the Hueco Group and Laborcita Formation (Upper Pennsylvanian-Lower Permian), South-Central New Mexico, USA. Rivista Italiana di Paleontologia e Stratigrafia, 109:3–36
Krainer K, Vachard D, Lucas SG (2009) Facies, microfossils (smaller foraminifers, calcareous algae) and biostratigraphy of the Hueco Group, Dona Ana Mountains, southern New Mexico, USA. Rivista Italiana di Paleontologia e Stratigrafia 115:3–26
Leda L, Korn D, Ghaderi A, Hairapetian V, Struck U, Reimold WU (2014) Lithostratigraphy and carbonate microfacies across the Permian–Triassic boundary near Julfa (NW Iran) and in the Baghuk Mountains (Central Iran). Facies 60:295–325
Lys M, Stampfli G, Jenny J (1978) Biostratigraphie du Carbonifere et du Permien de l’Elbourz oriental (Iran du NE). Notes Lab Paleont Univ Geneve 13:63--99
Maslov VP (1950) Importance of red algae for the stratigraphy of USSR. Doklady Akademii nauk SSSR 70(1):75–78. [in Russian]
Okla SM (1992) Permian Algae and Algal Microfacies from Unayzah, Quassim District, Saudi Arabia. Facies, 27:217--224
Partoazar H (1995) Permian deposits in Iran. Treatise on the geology of Iran. Geological Survey of Iran 22:1--340 [in Persian].
Partoazar H (2002) Permian-Triassic boundary conodonts from Jolfa-Abadeh Belt along Northwest and Central Iran. Permophiles, 41:34--40
Parvizi T, Rashidi K, Vachard D (2013) Middle Permian calcareous algae and microproblematica (Dalan Formation, Dena Mountain, High Zagros, SW Iran). Facies 59:149–177
Rostovtsev KO, Azaryan NR, 1973 The Permian-Triassic Boundary in Transcaucasia. In: The Permian and Triassic systems and their mutual boundary. Canadian Society of Petroleum Geologists Memoir 2: 89–99
Ruzhentsev VE, Sarycheva TG, (Eds.,), 1965. Razvitie i smena morskich organismov na rubezhe paleozoja i mezozoja. Trudy Paleont. Inst. AN SSSR (108). 431 pp
Saitoh M, Isozaki Y, Yao J, Ji Z, Ueno Y, Yoshida N (2013) The appearance of an oxygen-depleted condition on the Capitanian disphotic slope/basin in South China: Middle–Upper Permian stratigraphy at Chaotian in northern Sichuan. Glob Planet Chang 105:180–192
Scotese CR, Langford RP (1995) Pangea and the paleogeography of the Permian. In: Scholle PA, Peryt TM, Ulmer-Scholle DS (eds) The Permian of Northern Pangea. Springer, Berlin, pp. 3–19
Sengor AMC (1979) Mid-Mesozoic closure of Permo-Triassic Tethys and its complications. Nature 279:590–593
Shabanian R, Bagheri M, Hossein Yar Gh.R (2008) Introduction to Permian calcareous algae Northwest Iran. 12th congress of Geological society of Iran, pp. 28--32 [In Persian].
Stampfli GM, Borel GD (2002) A plate tectonic model for the Paleozoic and Mesozoic constrained by dynamic plate boundaries and restored synthetic oceanic isochrons. Earth Planet Sci Lett 196:17–33
Stampfli GM, Borel GD (2004) The TRANSMED transects in space and time. Constraints on the paleotectonic evolution of the Mediterranean domain. In: Cavazza W, Roure F, Spakman W, Stampfli GM, Ziegler P (eds) The TRANSMED atlas: the Mediterranean region from crust to mantle. Springer, Berlin, pp. 53–80
Stampfli GM, Marcoux J, Baud A (1991) Tethyan margins in space and time. Palaeogeogr Palaeoclimatol Palaeoecol 87:373–409
Stepanov DL, Golshani F, Stöcklin J, 1969. Upper Permian and Permian–Triassic Boundary in North Iran. Geological Survey of Iran, Report 12: 1–72.
Stoyanow AA (1910) On the character of the boundary of Palaeozoic and Mesozoic near Djulfa. Zap Imp S-Peterburgskago Miner Obshch, nov Ser 47:61–135
Teichert C, Kummel B, Sweet WC (1973) Permian-Triassic strata, Kuh-e-Ali Bashi, northwestern Iran. Bulletin of the Museum of Comparative Zoology 145:359–472
Torsvik TH, Cocks LRM (2004) Earth geography from 400 to 250 Ma: a palaeomagnetic, faunal and facies review. Journal of the Geological Society of London 161:555--572
Vachard D (1980) Téthys et Gondwana au Paléozoïque supérieur; les données afghanes: biostratigraphie, micropaléontologie, paléogéographie. Documents et Travaux Institut Géologique Albert de Lapparent 2:1–463
Vachard D, Krainer K (2001) Smaller foraminifers,characteristic algae and pseudo-algae of the latest Carboniferous/Early Permian Rattendorf Group, CarnicAlps (Austria/Italy). Rivista Italiana di Paleontologiae Stratigrafia, 107:169–195
Vachard D, Aretz M (2004) Biostratigraphical precisions on the Early Serpukhovian (Late Mississippian), by means of a carbonate algal microflora (cyanobacteria, algae and pseudo-algae) from La Serre (Montagne Noire, France). Geobios 37:43–666
Vachard D, Tahiri A (1991) Foraminifères, algues et pseudo-algues du Viséen de la region d’Oulmès (Maroc). Géologie Méditerranéenne 18:21–41
Vaziri SH, Akira Y, Kuwahara K (2005) Lithofacies and microbiofacies (foraminifers and radiolarians) of the Permian Sequence in the Shalamzar area, Central Alborz, North lran. J Geosci, Osaka City University 48:39--69
Wood R (1998) The ecological evolution of reefs. Annu Rev Ecol Syst 29:179–206
Acknowledgments
The authors thank the Ferdowsi University of Mashhad and the Aras Free Zone office for support during the field works. They kindly acknowledge the anonymous reviewers who improve the quality of this article by their constructive comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Ghaderi, A., Taherpour Khalil Abad, M., Ashouri, A.R. et al. Permian Calcareous algae from the Khachik Formation at the Ali Bashi Mountains, NW of Iran. Arab J Geosci 9, 699 (2016). https://doi.org/10.1007/s12517-016-2737-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-016-2737-7