
Abstract
There are few archaeological projects incorporating archaeobotanical sampling and even fewer published archaeobotanical studies in Thailand. Available data show that rice was the ubiquitous cereal in prehistory and particularly during the Metal/Iron Age. This either signifies the importance of rice as a crop or signals a preservation bias; both topics are considered in this paper. The site Khao Sam Kaeo in Peninsular Thailand (ca. 400–100 BCE) is strategically located between the Indian Ocean and the South China Sea providing evidence of Indian, Han Chinese and locally produced cultural material. The archaeobotanical assemblage attests to South Asian and East Asian influence as well: the mungbean and horsegram of Indian origin and the northern Chinese cereal foxtail millet. But the site has also yielded the greatest amount of rice from Thai archaeology and provides information on the domestication of rice and the cultivation practices during this Late Prehistoric period.
Similar content being viewed by others


Introduction
This paper is intended to review what we know so far about rice in Thailand during prehistory using an archaeobotanical approach. There are very few archaeobotanical studies published from Thai archaeological sites making it difficult to have a clear picture of this history. I use my work in three sites (Khao Sam Kaeo [KSK], Phukhao Thong [PKT] and Ban Non Wat [BNW]) and the published data from four other sites (Khok Phanom Di [KPD], Non Pa Wai [NPW], Non Mak La [NML] and Nil Kham Haeng [NKH]) to interpret the evolution of rice in prehistoric Thailand through cultivation systems and the type of rice cultivated. Furthermore, I provide preliminary information on charring experiments of cereals to show that preservation biases exist and should be considered in discussions.
Results
Rice finds in Thailand
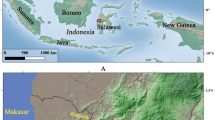
During the course of my research, I have identified 29 sites in Thailand (Fig. 1) which report rice finds dating from the Hoabinhian to the Late Prehistoric period (Fuller et al. 2010 online supplement). There are differences in the data quality dependent on the accuracy of interpretation and these were taken into account. From these sites, only nine have evidence of rice resulting from flotation and an additional eight from phytolith analysis. In the case of rice, the presence of rice spikelet bases allows for the identification of the domestication status of rice.

Map showing sites with evidence of rice. 1 Banyan Valley Cave; 2 Phu Lon; 3 Ban Chiang; 4 Nong Han Kumphawapi, Ban Na Di; 5 Ban Chiang Hian, Non Noi, Ban Kho Noi; 6 Non Nok Tha; 7 Non Dua, Don Taphan; 8 Ban Non Wat, Phimai, Noen U-Loke, Non Muang Kao, Ban Tamyae; 9 Khok Phanom Di, Nong Nor; 10 Khok Charoen; 11 Non Pa Wai, Nil Kham Haeng, Non Mak La, Lopburi, Ban Tha Kae; 12 Ban Don Ta Phet, Ban Na Khun Saen 2; 13 Khao Sam Kaeo; 14 Phukhao Thong; 15 Non Khao Wong.
The earliest sites where rice has been found show rice may have possibly been cultivated as early as the mid-Holocene in north and central Thailand (Kealhofer 2002). These inferences have been based on phytolith studies. The phytoliths from these sites were taken from sediment sequences in lake cores and alluvial deposits and the rice cannot be considered domesticated. Evaluating the status of domestication using phytoliths remains problematic (Fuller and Qin 2009; Fuller et al. 2010), though there are several scholars who believe it is possible (Saxena et al. 2006; Zhao et al. 1998). In order to assess the domestication status of rice, archaeobotanists examine the abscission scars found in rice spikelet bases, which can only be done with macroremains (Thompson 1997; Fuller et al. 2009). If one were to strictly adhere to the examination of rice spikelet bases, there would only be a handful of sites in Thailand that would positively yield evidence for domesticated rice. Most rice reports come from rice temper or impressions in pottery consisting mainly of husks and, like phytoliths, these data do not provide information on the domesticated status of the cereal.
The first evidence of domesticated rice in Thailand using macroremains dates to 2000–1500 BCE from the Neolithic period in the coastal site of KPD (Thompson 1996). KPD provides rice finds in the form of domesticated-type rice spikelet bases and weeds of cultivation. Higham (2002) originally proposed that rice agricultural expansion followed major riverine routes and would be archaeologically visible in interior sites, an idea previously put forth for Austroasiatic language expansion by Blust (1996). However, Ban Tha Kae and Ban Chiang are the earliest interior sites dating to the Neolithic and are reported to have rice cultivation, but the evidence is based on rice-tempered pottery, so it may be open to doubt. The first inland sites that provide reliable domesticated rice finds are NPW, NKH and NML in Lopburi. The rice finds at these sites date to the first millennium BCE and not earlier. Interestingly, these sites provide evidence that millets were cultivated before rice. The primary crop found in all three sites in the second millennium BCE was foxtail millet (Setaria italica) and the mode of cultivation was dryland farming (Weber et al. 2010). NPW has evidence of Setaria during the third millennium BCE signifying the introduction of millet cultivation at least a thousand years before rice at this site. This cereal originates from the north of China, though in the third millennium BCE, it was also evident in south China bordering Vietnam together with rice remains (Fuller et al. 2010).
In the Late Prehistoric period, more evidence for domesticated rice comes from samples from the Iron Age site BNW. These were floated and rice grain, spikelet bases, husk and weeds of cultivation have been identified. The Metal Age (400–200 BCE) sites KSK and PKT in the southern Peninsula have also yielded a large number of rice remains and associated weeds, as well as the Indian pulses Vigna radiata and Macrotyloma uniflorum. All three sites have domesticated japonica-type rice and possibly dryland and rainfed rice cultivation systems (Castillo and Fuller 2010).
Origins of rice
The widely held view is that rice in Southeast Asia came from China and that it was Oryza sativa spp. japonica. The linguistic evidence indicates that the original domesticators of rice, depending on the author of the hypothesis, were the Miao-Yao coming from south and central China (Blench 2005), the Austroasiatic speakers (Sagart 2005) or Austric speakers (Blust 1996; Higham 1996) coming from the Yangzi Valley. Movements of agriculturalists have also been proposed such as the Austronesians from Taiwan to the Philippines and further southwards (Bellwood 2007) and the Tibeto-Burman into northern China (van Driem 1998). The archaeological evidence consistently points to the Yangzi valley as the area where rice was first domesticated (Fuller et al. 2007, 2010; Nakamura 2010; Zhao 2010). However, which group of people brought rice cultivation to Thailand remains a matter of debate. Unfortunately, archaeological work in the region does not assist due to the lack of archaeobotanical sampling (Castillo and Fuller 2010). There are not enough rice finds to permit geographic and chronological resolution for a clear picture of the diffusion of rice cultivation to emerge.
Genetic studies remain divided as to whether rice domestication had a single origin or multiple origins (He et al. 2011; Molina et al. 2011; Sang and Ge 2007). The multiple-origins model proposes two centres of domestication, one in China ca. 4000 BCE and the other in South Asia ca. 2000 BCE (Fuller 2007). The single-origin model considers indica to be a hybrid of japonica rice and therefore, the origin lies in China even though its development or expansion occurred in India (Molina et al. 2011). The archaeobotanical evidence in Thailand does not corroborate one or the other. It does, however, point towards a largely japonica-type variety in prehistory and using either model ultimately shows that rice in prehistoric Thailand until at least the Iron Age has its origins in China.
The morphometric analyses of rice grains from four Thai archaeological sites (KSK, PKT, BNW and Noen U-Loke [NUL]) suggest that rice in prehistoric Thailand was Oryza sativa japonica (Fig. 2). The length–width (L/W) ratios of these rice grains were compared with those of modern populations of domesticated and wild rice. According to Ahn (1993), L/W ratios are not affected by charring so ancient and modern rice should therefore be comparable. Indica rice normally has a L/W ratio above 2.5, whereas japonica rice is below 2.3 (Fuller et al. 2009). The prehistoric rice measured all come from Iron and Late Metal Age sites from the NE region and southern Peninsula of Thailand (map 1) indicating that ca. 400–200 BCE, it was the Chinese rice subspecies japonica that was being consumed and cultivated. However, genetic studies are needed to confirm this view as morphometrics is just a first step towards identification. At present, there has been no DNA fingerprinting of prehistoric rice in Thailand.

Comparison of L/W ratios of rice from four prehistoric sites in Thailand to modern populations of domesticated and wild rice. Modern and wild population measurements courtesy of Fuller.
The first rice samples from the sites of BNW, NUL and KSK were sent to Japan in July 2011 and will be analysed using DNA chloroplast and nuclear genome markers. The analysis will, hopefully, provide information on rice variety (indica, tropical japonica, temperate japonica) and whether it was the waxy or sticky type of rice.
The morphometric analysis above indicates that at the four sites (KSK, BNW, PKT and NUL) during the late prehistoric period, japonica was the type of rice found across Thailand. But several questions arise. When did indica become the dominant rice variety; what agricultural regime was practised in prehistory (wetland vs. dryland cultivation); was the cultivation technique also brought in with the introduction of rice or was it a local innovation?
Cultivation systems and weeds of cultivation
Today, indica is the dominant rice type and rainfed cultivation is the main agricultural system practised in Thailand. Cultivation today uses bunded fields inundated by retaining rainwater and allowed to dry naturally. Rainfed systems of cultivation were most likely practised during prehistoric times as well. White (1995) proposes that in Thailand, both wetland and dryland rice cultivation evolved from inundated rice cultivation, a less labour demanding technique than irrigated cultivation. This is true in the case of the low-lying coastal site KPD during the Neolithic where rice cultivation is believed to have been dependent on natural flooding at a nearby swamp (Thompson 1996).
To define systems of land use and cultivation practices, archaeobotanists have relied on the weed flora associated with economic crops because weed species occur in certain ecological zones, are displaced travelling with particular crop packages and help identify crop processing stages (Bogaard et al. 1999; Colledge 1994; Colledge et al. 2005; Fuller and Qin 2009; Jones 2002; Kealhofer and Piperno 1994). Unfortunately, macroremains from the Khao Wong Prachan valley sites (NKH, NPW and NML) dating to the Bronze Age do not contain sufficient numbers of weed seeds to define the rice agricultural regime and such weed seeds as are found provide ambiguous results. Sedges normally associated with wetland rice as well as dryland weeds such as chenopods are found in the samples (Weber et al. 2010). Furthermore, prior to rice cultivation, millets were being cultivated in dryland conditions, potentially signifying a continuum in the cultivation practice for rice in the area.
During the Metal Age at KSK, the majority of the weed assemblage comes from dryland habitats. The predominant weed is Spilanthes acmella belonging to the Asteraceae family (Fig. 3). It is significant in that 94% of the samples with rice contained this weed representing a high level of co-occurrence. Furthermore, S. acmella is reported to be a weed of rice throughout Indonesia, Bangladesh, India, Philippines, Sri Lanka and Thailand (Moody 1989) found in rainfed and upland fields (Soerjani et al. 1987). The other crops found at KSK, such as foxtail millet and the mungbean, are also indicative of dryland cultivation systems and are drought-resistant and found mainly in upland cultivation systems. So it is clear from the weeds and other cultivars that the rice cultivation system at KSK was dryland.

SEM image of the weed Spilanthes acmella from Khao Sam Kaeo.
A similar assemblage of pulses and weeds associated with rice at KSK is also found at PKT, another Metal Age site in the southern Peninsula. The weed assemblage at the Iron Age site BNW in northeast Thailand also indicates dryland cultivation. It appears that rice cultivation in Thailand during the Metal Age was rainfed and upland. As a point of comparison, a geomorphological study in Kedah situated in the Thai–Malay Peninsula hypotheses communities in the first millennium CE being dependent on dryland cereal cultivation and not irrigated rice agriculture (Allen 1991).
The cultivation practices inferred from prehistoric sites in Thailand and one in the Thai–Malay Peninsula discussed above differ from the lowland paddy field agricultural system that was in place at the centre of origin in the Lower Yangtze when rice spread outwards to other regions ca. 4000 BCE (Fuller and Qin 2009). This difference may be because wetland paddy field agriculture in Thailand developed later. Although the earliest paddy field agriculture is found in China, it is during the first millenium CE that indica together with wetland systems of cultivation may have been introduced into Southeast Asia from India as a result of exchange networks. In India, the expansion of rice agriculture occurs during the Iron Age and is linked to labour-intensive irrigated rice cultivation (Castillo and Fuller 2010; Fuller and Qin 2009; Shaw et al. 2007). It seems likely that during the early contact period with South Asia (300 BCE onwards), Thailand already had an established rice agricultural regime primarily focused on dry cropping in low-lying areas and the rice grown was japonica. KSK attests to this conclusion. It was after continuous contact with India that wetland systems of agriculture were developed.
Discussion
Khao Sam Kaeo: the point of contact with India
KSK lies in the narrowest stretch of the Thai–Malay Peninsula known as the Kra Ithmus. Thus, it is in a strategic location for several spheres of influence and contact from South Asia to the west and East Asia and Insular Southeast Asia to the east. The evidence of these contacts is well-documented (Bellina 2007; Bellina and Silapanth 2006; Bellina-Pryce and Silapanth 2008; Glover and Bellina 2011). Excavations from 2006 to 2009 at this late prehistoric site included an archaeobotanical agenda, and the macroremains from the site mirror the material culture having both South Asian and East Asian originated crops. South Asian influence is not evident before the Iron Age in Thailand.
The archaeobotanical remains include the mungbean (V. radiata) and the horsegram (M. uniflorum), both originating from India. There are no earlier reported finds of either horsegram or mungbean in Thailand before the Iron Age. In principle, the mungbean could have been domesticated in Thailand since the wild progenitor is found all across Thailand. However, genetic studies have shown that the modern Thai domesticated mungbean is more closely related to wild progenitors found in India than the native Thai wild progenitor (Castillo and Fuller 2010). This may indicate that the mungbean was brought into Thailand by Indian people as a domesticated pulse probably through the southern Peninsula via entrepôts like KSK. The cereal foxtail millet is one of the two Chinese native crops found at KSK, the other, is the rice. This suggests that there was a package of South Asian crops which included mungbean and horsegram brought into Thailand. This late prehistoric package may not have included rice as it was probably already in cultivation. Cultivation in KSK or its hinterlands is probable due to the large size of this 34-ha urban settlement. In order to maintain the social network composed of specialised craftsmen and travellers, there must have been an agricultural base to sustain them. Although rice fields have not been identified yet, the geomorphological study at KSK (Allen 2009; Allen and Silapanth unpublished report) indicates probable cultivation was on gently sloping plateau land and hill slopes in the hinterlands.
Caveats: preservation bias
Rice is the most commonly reported archaeobotanical find in Thailand. This is not surprising because unlike other macroremains, rice is easy to recognise. It also has distinctive phytoliths (e.g. bulliforms) and is large enough to be found with the naked eye, whereas the retrieval of finer fraction (e.g. millets) requires the use of flotation. Also being now the most important economic crop in Southeast Asia, scholars have dedicated more effort in the search of prehistoric rice to clarify its history and that of the people that consumed and produced it and have therefore reported more finds than for other cereals.
At several excavations in Thailand where I have worked as the on-site archaeobotanist, I have found that rice is the most common crop in the samples floated. This pattern leads one to assume that rice was the most important economic crop in prehistory, just as it is today. However, preservation biases must be considered. The lack of substantial evidence of millets in Southeast Asia has been attributed to preservation issues, for example (Weber et al. 2010; Weber and Fuller 2008). Very preliminary interpretations of charring experiments using real fire instead of a muffler furnace suggest that whilst rice grain does not necessarily preserve better than some other crops, it does have greater visibility when the husk is taken into account. In comparison to the small millets, Panicum milliaceum (broomcorn millet) and Setaria italica (foxtail millet), hulled rice is more easy to recognise than each of the millets in charring experiments (Table 1).
Although details of these charring experiments are discussed elsewhere (Castillo unpublished), the main conclusions are as follows: All three cereals preserve better when hulled, and rice disintegrates to an unidentifiable state when naked grains are charred. In two out of the first three experiments, the preservation of rice (excluding husk but including spikelet bases) compared to the millets was higher, though it was especially high when the rice had been in the fire for the longest duration of the experiment (Ex1). In all the three experiments where hulled cereal was used (Ex1, Ex2 and Ex3), rice husks preserved better than the husks from the millets. These experiments lead me to believe that if hulled rice or the waste products after dehusking happened to come into contact with fire in prehistory, they would have a higher chance of preserving than remains of the two millets referred to above. The archaeological record shows that ca. 67% of reported rice in Thailand are rice husks or rice impressions (Fuller et al. 2010 online supplement). Similarly, in my own work at KSK, rice spikelet bases comprise 78% of the rice assemblage (Castillo unpublished).
It appears that at least in the Bronze Age in the Khao Wong Prachan Valley, millets were the dominant cereals and were cultivated before rice (Weber et al. 2010). Perhaps the reason why millets in this area are archaeologically visible is because flotation was used. In order to verify this conclusion what is needed is to continue gathering samples for flotation from more sites spanning hunter-gatherer sites to the proto-historic periods across the whole of Thailand.
Conclusion
There is no doubt that more archaeological and archaeobotanical work needs to be carried out across Thailand as well as in the neighbouring countries in order to refine the history of rice agriculture in the region. In this paper, I have outlined what is known so far about rice in Thailand and used an archaeobotanical approach to qualify what information can actually be inferred from these data. The presence of rice, its domestication status, variety and cultivation practices all contribute to the understanding of the origins and movements of rice. However, we should also take into consideration that other domesticates, including millets, may have been the precursors to rice and how this will contribute to our understanding of the people that settled or migrated to Thailand.
- BNW:
-
Ban Non Wat
- KPD:
-
Khok Phanom Di
- KSK:
-
Khao Sam Kaeo
- NKH:
-
Nil Kham Haeng
- NML:
-
Non Mak La
- NPW:
-
Non Pa Wai
- PKT:
-
Phukhao Thong
References
Ahn S-M. Origin and differentiation of domesticated rice in Asia. Unpublished PhD. thesis, Institute of Archaeology, University of London; 1993.
Allen J. Trade and site distribution in early historic-period Kedah: geoarchaeological, historic, and locational evidence. Indo-Pacific Prehistory Assoc Bull. 1991;10:307–19.
Allen J. Etude geomorphologique: paleo-drainage, erosion, gestion des sols et agriculture. In: Bellina B, editor. La Campagne 2009 de la Mission Franco-Thai en Peninsule Thai-Malaise. Paris: CNRS; 2009. p. 20–45.
Allen J, Silapanth P. Soil profiles and interpretative summaries, five excavation units, Khao Sam Kaeo; Unpublished report.
Bellina B. Cultural exchange between India and Southeast Asia: production and distribution of hard stone ornaments (VI c. BC-VI c. AD). Paris: Éditions de la Maison des sciences de l’homme; Éditions Épistèmes; 2007.
Bellina B, Silapanth P. Khao Sam Kaeo and the Upper Thai Peninsula: understanding the mechanisms of early trans-Asiatic trade and cultural exchange. In: Bacus EA, Glover IC, Pigott VC, editors. Uncovering Southeast Asia’s past: selected papers from the 10th International Conference of the European Association of Southeast Asian Archaeologists. Singapore: National University of Singapore Press; 2006. p. 379–92.
Bellina-Pryce B, Silapanth P. Weaving cultural identities on trans-Asiatic networks: Upper Thai-Malay Peninsula – an early socio-political landscape. Bull de l’Ecole Fr d’Extrême-Orient. 2008;93(2006):257–93.
Bellwood P. Prehistory of the Indo-Malaysian Archipelago. 2nd ed. Canberra: ANU Press; 2007.
Blench RM. From the mountains to the valleys: understanding ethnolinguistic geography. In: Sagart L, Blench RM, Sanchez-Mazas A, editors. SE Asia, in perspectives on the phylogeny of East Asian languages. London: Curzon Press; 2005. p. 31–50.
Blust R. Beyond the Austronesian homeland: the Austric hypothesis and its implications for archaeology. In: Goodenough WH, editor. Prehistoric settlement of the Pacific. Philadelphia: American Philosophical Society; 1996. p. 117–40.
Bogaard A, Palmera C, Jones G, Charles M, Hodgson JG. A FIBS approach to the use of weed ecology for the archaeobotanical recognition of crop rotation regimes. J Archaeol Sci. 1999;26:1211–24.
Castillo C, Fuller DQ. Still too fragmentary and dependent upon chance? Advances in the study of early Southeast Asian archaeobotany. In: Bellina-Pryce B, Pryce TO, Bacus E, Wisseman-Christie J, editors. 50 Years of archaeology in Southeast Asia: essays in honour of Ian Glover. Bangkok: River Books; 2010. p. 90–111.
Colledge S. Plant exploitation on epipalaeolithic and early neolithic sites in the Levant, PhD thesis, Department of Archaeology and Prehistory. Sheffield: University of Sheffield; 1994.
Colledge S, Conolly J, Shennan S. The evolution of neolithic farming from SW Asian Origins to NW European limits. Eur J Archaeol. 2005;8(2):137–56.
Fuller DQ. Non-human genetics, agricultural origins and historical linguistics in South Asia. In: Petraglia MD, Allchin B, editors. The evolution and history of human populations in South Asia. Netherlands: Springer; 2007. p. 393–443.
Fuller DQ, Qin L. Water management and labour in the origins and dispersal of Asian rice. World Archaeol. 2009;41(1):88–111.
Fuller DQ, Qin L, Harvey EL. A critical assessment of early agriculture in East Asia, with emphasis on Lower Yangtze Rice Domestication. Pradghara; 2007: 18.
Fuller DQ, Qin L, Harvey EL. An evolutionary model for Chinese rice domestication: reassessing the data of the Lower Yangtze region. In: Ahn S-M, Lee J-J, editors. New approaches to prehistoric agriculture. Seoul: Sahoi Pyoungnon; 2009. p. 312–45.
Fuller DQ, Sato Y-I, Castillo C, Qin L, Weisskopf AR, Kingwell-Banham EJ, Song J, Ahn S-M, van Etten J. Consilience of genetics and archaeobotany in the entangled history of rice. Archaeol Anthropol Sci. 2010;2(2):115–31.
Glover IC, Bellina B. Ban Don Ta Phet and Khao Sam Kaeo: the earliest Indian contact reassessed, in Proceedings of the International Conference Early Indian Influences in Southeast Asia: reflections on cross-cultural movements. Singapore: Institute of Southeast Asian Studies; 2011.
He Z, Zhai W, Wen H, Tang T, Wang Y, Lu X, Greenberg AJ, Hudson RR, Wu C-I, Shi S. Two evolutionary histories in the genome of rice: the roles of domestication genes. PLoS Genet. 2011;7(6):e1002100. doi:10.1371/journal.pgen.1002100.
Higham CFW. The Bronze Age of Southeast Asia Cambridge. Cambridge: Cambridge University Press; 1996.
Higham CFW. Languages and farming dispersals: austroasiatic languages and rice cultivation. In: Bellwood P, Renfrew C, editors. Examining the farming/language dispersal hypothesis. Cambridge: McDonald Institute for Archaeological Research; 2002. p. 223–32.
Jones G. Weed ecology as a method for the archaeobotanical recognition of crop husbandry practices. Acta Palaeobotanica. 2002;42:185–93.
Kealhofer L. Changing perceptions of risk: the development of agro-ecosystems in Southeast Asia. Am Anthropol. 2002;104(1):178–94.
Kealhofer L, Piperno D. Early agriculture in Southeast Asia: phytolith evidence from the Bang Pakong Valley, Thailand. Antiquity. 1994;68(260):564–72.
Molina J, Sikora M, Garud N, Flowers JM, Rubinstein S, Reynolds A, Huang P, Jackson S, Schaal BA, Bustamante CD, Boyko AR, Purugganan MLD. Molecular evidence for a single evolutionary origin of domesticated rice. PNAS. 2011;108(20):8351–6.
Moody K. Weeds reported in rice in South and Southeast Asia. Los Baños: International Rice Research Institute; 1989.
Nakamura S-I. The origin of rice cultivation in the Lower Yangtze Region, China. Archaeol Anthropol Sci. 2010;2(2):107–13.
Sagart L. Tai-Kadai as a subgroup of Austronesian. In: Sagart L, Blench RM, Sanchez-Mazas A, editors. Perspectives on the Phylogeny of East Asian Languages. London: Curzon Press; 2005. p. 177–81.
Sang T, Ge S. The puzzle of rice domestication. J Integr Plant Biol. 2007;49:760–8.
Saxena A, Prasad V, Singh IB, Chauhan MS, Hasan R. On the Holocene record of phytoliths of wild and cultivated rice from Ganga Plain: evidence for rice-based agriculture. Curr Sci. 2006;90(11):1547–52.
Shaw J, Sutcliffe JV, Lloyd-Smith L, Schwenninger J-L, Chauhan MS. Ancient irrigation and Buddhist history in Central India: optically stimulated luminescence dates and pollen sequences from the Sanchi Dams. Asian Perspect. 2007;46(1):166–201.
Soerjani M, Kostermans AJGH, Tjitrosoepomo G. Weeds of rice in Indonesia. Jakarta: Balai Pustaka; 1987.
Thompson GB. The excavation of Khok Phanom Di, a prehistoric site in central Thailand, vol IV: subsistence and environment: the botanical evidence (the biological remains, part II). London: The Society of Antiquaries of London; 1996.
Thompson GB. Archaeobotanical indicators of rice domestication - a critical evaluation of criteria. In: Ciarla R, Rispoli F, editors. South-east Asian Archaeology 1992. Proceedings of the Fourth International Conference of the European Asssociation of South-east Asian Archaeologists. Rome, 28th September - 4th October 1992. Rome: Istituto Italiano per L’Africa e L’Oriente; 1997. p. 159–73.
Van Driem G. Neolithic correlates of ancient Tibeto-Burman migrations. In: Blench R, Spriggs M, editors. Archaeology and language II: archaeological data and linguistic hypotheses. New York: Routledge; 1998. p. 67–102.
Weber SA, Fuller DQ. Millets and their role in early agriculture. Pragdhara. 2008;18:69–90.
Weber S, Lehman H, Barela T, Hawks S, Harriman D. Rice or millets: early farming strategies in prehistoric central Thailand. Archaeol Anthropol Sci. 2010;2(2):79–88.
White J. Modeling the development of early rice agriculture: ethnoecological perspectives from Northeast Thailand. Asian Perspect. 1995;34(1):37–68.
Zhao Z. New data and new issues for the study of origin of rice agriculture in China. Archaeol Anthropol Sci. 2010;2(2):99–105.
Zhao Z, Pearsall D, Benfer Jr R, Piperno D. Distinguishing rice (Oryza sativa Poaceae) from wild Oryza species through phytolith analysis, II: finalized method. Econ Bot. 1998;52(2):134–45.
Acknowledgements
I thank Dr. Dorian Fuller and Prof. Vincent Pigott for their comments, Dr. Berenice Bellina for giving me the opportunity to work at Khao Sam Kaeo, John Watson for help in the editing of the manuscript and the anonymous reviewer for providing useful comments. This work was funded in part by the Arts and Humanities Research Council and the Royal Thai Embassy in London.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Castillo, C. Rice in Thailand: The Archaeobotanical Contribution. Rice 4, 114–120 (2011). https://doi.org/10.1007/s12284-011-9070-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12284-011-9070-2