
Abstract
Anti-inflammatory transcriptional effects of nineteen compounds (1–19) from the soft coral Sinularia maxima were evaluated using NF-κB luciferase and reverse transcriptase polymerase chain reaction. Compounds 1, 2, 4, 8, 15, 17, and 18 significantly inhibited TNFα-induced NF-κB transcriptional activity in HepG2 cells in a dose-dependent manner, with IC50 values ranging from 15.81 ± 2.29 to 29.10 ± 1.54 μM. Furthermore, the transcriptional inhibitory function of these compounds was confirmed by a decrease in intercellular adhesion molecule-1 and inducible nitric oxide synthase gene expression levels in HepG2 cells. These results provide a scientific rationale for the use of the soft coral S. maxima warrant further studies to develop new agents for the prevention and treatment of inflammatory.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Activation of nuclear factor kappa B (NF-κB) represents a family of Rel domain-containing proteins including five NF-κB units that can form 15 transcription factors through homo- and heterodimerization (Baldwin 2001a). NF-κB plays an important role in the transcriptional regulation of numerous cytokines and adhesion molecules. It is arguably the most important transcription factor for the initiation or progression of numerous human diseases (Mattson and Camandola 2001). As a ubiquitous transcription factor governing the expression of viruses or a variety of inflammatory cytokine genes, NF-κB was first implicated in the pathogenesis of human immunodeficiency virus-1 (HIV-1) infection (Nabel and Baltimore 1987). NF-κB, a nuclear transcription factor, regulates the expression of various genes, including cytokines, iNOS, COX-2 and ICAM-1, which play critical roles in apoptosis, autoimmune diseases, and inflammation (Albert and Baldwin 1996). Further studies suggest that activation of NF-κB is responsible for the pathological progression of neurological disorders, carcinogenesis, immune deficiency, rheumatoid arthritis, atherogenesis, Crohn’s disease, cystic fibrosis, asthma, osteopetrosis, ischemic reperfusion, etc. (Chen et al. 1999).
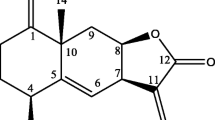
Among the Alcyonacean soft corals, genus Sinularia is one of the most widely distributed soft coral genera, constituting a dominant portion of the biomass in the tropical reef environment (Lakshmi and Kumar 2009). Previous studies have indicated that diterpenes, a main constituent of the genus Sinularia, exhibit various biological activities, such as anti-inflammatory (Chao et al. 2011; Cheng et al. 2010; Lu et al. 2010; Su and Wen 2011), antiviral (Cheng et al. 2010), and cytotoxic (Grote et al. 2008; Kamel et al. 2007; Lo et al. 2009; Su et al. 2009) effects. As a part of our ongoing investigations on screening active compounds from Vietnamese Sinularia soft corals towards anti-inflammatory effects (Thao et al. 2012, 2013a, b), we recently reported the isolation, structure elucidation, and inhibitory effects on lipopolysaccharide-stimulated production of proinflammatory cytokines in bone marrow-derived dendritic cells of 12 diterpenoids [sinumaximol A (1), sinumaximol B (2), sinumaximol C (3), sethukarailin (4), sinumaximol D (5), sinumaximol E (6), sinumaximol F (7), sinumaximol G (8), sinumaximol H (9), (1S,2E,4S,6E,8S,11R)-2,6,12(20)-cembratriene-4,8,11-triol (10), isomandapamate (11), and sinumaximol I (12)] (Thao et al. 2012) and 7 norditerpenoids [scabrolide A (13), 12-hydroxy-scabrolide A (14), yonarolide (15), ineleganolide (16), 5-epinorcembrene (17), 13-epi-scabrolide C (18), and norcembrene 5 (19)] (Thao et al. 2013a) from Sinularia maxima (see Fig. 1). The current study provides new insight into the ways by which diterpenoids and norditerpenoids modulate TNFα-induced NF-κB activity in human HepG2 cells.

Structure of compounds 1–19 from the soft coral Sinularia maxima
Materials and methods
Biological material
The sample of soft coral S. maxima was collected at Nhatrang Bay, in November in 2010 and identified by Prof. Do Cong Thung (Institute of Marine Environment and Resources, VAST). A voucher specimen (SM112010_01) was deposited at the Institute of Marine Biochemistry and Institute of Marine Environment and Resources, VAST.
Cell culture and reagents
Human hepatocarcinoma HepG2 cells were maintained in Dulbecco’s modified Eagle’s medium (Invitrogen, Carlsbad, CA, USA) containing 10 % heat-inactivated fetal bovine serum, 100 units/mL penicillin, and 10 μg/mL streptomycin at 37 °C and 5 % CO2. Human TNFα was purchased from ATgen (Seoul, Korea). Cells were counted with a hemocytometer, and the number of viable cells was determined through trypan blue dye exclusion.
Cytotoxicity assay
A 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt (MTS) assay (CellTiter 96® AQueous One Solution Assay, Promega, Madison, WI, USA) was performed to analyze the effect of the different compounds on cell viability. Cells were cultured overnight in 96-well plates (1 × 104 cells per well). Cell viability was assessed after the incubation with the compounds at a concentration of 10 μM for 24 h. The number of viable cells was determined by measuring the absorbance at 490 nm of the dissolved formazan product after addition of MTS for 30 min as described by the manufacturer.
NF-κB and iNOS-luciferase assay
The luciferase vector was first transfected into HepG2 cells. After a limited amount of time, the cells were lysed and luciferin, the substrate of luciferase, was introduced into the cellular extract along with Mg2+ and an excess of ATP. Under these conditions, luciferase enzymes expressed by the reporter vector could catalyze the oxidative carboxylation of luciferin. Cells were seeded at 1.5 × 105 cells per well in a 12-well plate and grown for 24 h. All cells were transfected using Lipofectamine™ LTX (Invitrogen) according to the manufacturer’s protocol. Luciferase (Luc) activity was assayed using an LB 953 Autolumat (EG&G Berthold, Nashua, NH, USA) as described previously (Kim et al. 2010). NF-κB-Luc was kindly provided by Dr. Kyoon E. Kim (Chungnam National University, Daejeon, Korea). The transfected HepG2 cells were pretreated for 1 h with either vehicle (DMSO) and compounds, followed by 1 h of treatment with 10 ng/mL TNFα. Unstimulated HepG2 cells were used as a negative control (−). Cells were then harvested, and luciferase activity was assayed. All experiments were performed in triplicate.
RNA preparation and RT-PCR
HepG2 cells were pretreated in the absence and presence of compounds for 1 h, then exposed to 10 ng/mL TNFα for 6 h. Total mRNA was prepared from the cell pellets using Easy-blue. The levels of mRNA were assessed by reverse transcriptase polymerase chain reaction (RT-PCR) (Quang et al. 2012).
Total RNA was extracted from cells using easy-BLUE™ (iNtRON Biotechnology, Seoul). Approximately 2 μg of total RNA was reverse-transcribed using Moloney murine leukemia virus reverse transcriptase and oligo (dT) primers (Promega) for 1 h at 42 °C. The resulting cDNA was polymerase chain reaction-amplified using Taq polymerase premixture (TaKaRa, Shiga, Japan). Polymerase chain reaction products were subjected to electrophoresis on 1 % agarose gels and stained with ethidium bromide. Polymerase chain reaction was conducted with the following primer pairs: iNOS sense 5′-TCATCCGCTATGCTGGCTAC-3′, iNOS antisense 5′-CTCAGGGTCACGGCCATTG-3′, ICAM-1 sense 5′-GCCCAGCACTTCACGCATCAG-3′, ICAM-1 antisense 5′-GACCAGGCACCAGACCAAAGACC-3′, glyceraldehyde 3-phosphate dehydrogenase sense 5′-TGTTGCCATCAATGACCCCTT-3′, and glyceraldehyde 3-phosphate dehydrogenase antisense 5′CTCCACGACGTACTCAGCG-3′. β-actin sense 5′-TCACCCACACTGTGCCCATCTACG-3′, and β-actin antisense 5′-CAGCGGAACCGCTCATTGCCAATG-3′.
Statistical analysis
All results are expressed as mean±SD values. Data were analyzed by one-factor analysis of variance. Quantification of polymerase chain reaction products was performed using Image Lab™ software (Bio-Rad). If a statistically significant effect was found, the Newman–Keuls test was performed to isolate the difference between the groups. P < 0.5 was considered to be significant.
Results
To investigate cellular toxicity of the compounds 1–19, they were applied at various concentrations to HepG2 cells for 24 h, after which cell viability was measured in an MTS assay as described in Materials and methods. None of the compounds displayed any cellular toxicity at the concentration of 10 μM (data not shown). They were therefore used in subsequent experiments for further evaluation of their effects on NF-κB activation, iNOS and ICAM-1 expressions at concentrations of 0.1, 1.0 and 10 μM (see Fig. 4).
To evaluate the anti-inflammatory activity of nineteen compounds listed above, we first examined their inhibitory effects on NF-κB transcriptional activation in HepG2 cells (see Figs. 2, 3). Cells were treated with compounds at various concentrations prior to stimulation with TNFα (10 ng/mL). Among compounds tested, 1, 2, 4, 8, 15, 17, and 18 were found to have effect on the inhibition of NF-κB activation with 50 % inhibition concentration (IC50) values ranging from 15.81 ± 2.29 to 29.10 ± 1.54 μM. Other compounds exhibited moderate, weak or not determined activities (Table 1).

Effects of compounds 1–9 on tumor necrosis factor-α (TNFα)-induced nuclear transcription factor κB activation in HepG2 cells. HepG2 cells transiently transfected with pNF-κB-luciferase were pretreated for 1 h with vehicle (dimethyl sulfoxide-DMSO) or one of the compounds, prior to 1 h of treatment with TNFα (10 ng/mL). Unstimulated HepG2 cells acted as a negative control. Cells were then harvested, and luciferase activities were assessed. Results are expressed as relative luciferase activity. Sulfasalazine was used as a positive (Pos.) control. Data are mean–SD values (n = 3). P < 0.5 versus control

Effects of compounds 10–19 on tumor necrosis factor-α (TNFα)-induced nuclear transcription factor κB activation in HepG2 cells. HepG2 cells transiently transfected with pNF-κB-luciferase were pretreated for 1 h with vehicle (dimethyl sulfoxide-DMSO) or one of the compounds, prior to 1 h of treatment with TNFα (10 ng/mL). Unstimulated HepG2 cells acted as a negative control. Cells were then harvested, and luciferase activities were assessed. Results are expressed as relative luciferase activity. Sulfasalazine was used as a positive (Pos.) control. Data are mean–SD values (n = 3). P < 0.5 versus control
Discussion
NF-κB was first described in 1986 as a nuclear transcription factor required for immunoglobulin kappa light chain transcription in B-cells. Since its discovery, it has been demonstrated that NF-κB is constitutively expressed in all cell types and plays a central role as a regulator of the cellular stress response. The NF-κB-mediated signaling pathway has been considered both pro-inflammatory and anti-apoptotic in character, and therefore, has been implicated in the pathogenesis of a wide variety of diseases, including inflammatory disorders and tumor development (Robinson and Mann 2010). As previously demonstrated, activation of NF-κB has been linked to multiple pathophysiological conditions such as cancer, arthritis, asthma, inflammatory bowel disease, and other inflammatory conditions (Baldwin 2001b). It can be activated by various stimuli, such as microbial and viral products, cytokines, DNA damage, and noxious chemicals. The induction of several pro-inflammatory mediators occurs as a result of increased inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) activities (Surha et al. 2001). NF-κB and the signaling pathways that regulate many physiological processes, including innate and adaptive immune responses, cell death, and inflammation, have become a focal point for intense drug discovery and development efforts (Chung et al. 2007; Perkins 2007). Indeed, increasing evidence has validated NF-κB as a target for anti-inflammatory and anticancer agents.
To date, the inhibition of components from S. maxima on NF-κB transcriptional activation has not been evaluated. In this study, the effects of compounds 1–19 on TNFα-induced NF-κB transcriptional activity in HepG2 cells were evaluated using a NF-κB luciferase assay. To confirm their inhibitory effects of the compounds on NF-κB transcriptional activity, the effects of the isolated compounds on the upregulation of the pro-inflammatory proteins iNOS and ICAM-1 were assessed in TNFα-stimulated HepG2 cells by RT-PCR.
HepG2 cells were first transfected with NF-κB luciferase reporter plasmids. After treatment with 10 ng/mL TNF-α, luciferase activity increased fivefold, demonstrating an increase in transcriptional activity compared to untreated cells. Compounds were pretreated with transfected HepG2 cells at various concentrations, followed by stimulation with TNFα. The results showed that compounds 1, 2, 4, 8, 15, 17, and 18 significantly inhibited TNFα-induced NF-κB transcriptional activation in a dose-dependent manner with IC50 values of 21.35 ± 3.21, 29.10 ± 1.54, 25.81 ± 1.38, 15.81 ± 2.29, 25.1 ± 2.58, 28.19 ± 2.65, 20.13 ± 0.29 μM, respectively (see Figs. 2, 3). Compounds 3, 10–14, and 19 exhibited moderate or weak inhibitory effects with IC50 values ranging from 40.42 ± 2.11 to 325.71 ± 4.77 μM, whereas other compounds were inactive at the indicated concentrations, compared with the positive control (see Table 1).
NF-κB activation is known to be involved in the upregulation of inflammatory NF-κB target gene expression, including iNOS and ICAM-1, which play important roles in the inflammatory response. iNOS is highly expressed in macrophages, leading to organ destruction in some inflammatory and autoimmune diseases (Kleinert et al. 2004). ICAM-1 (CD54) is a 90-kDa inducible cell-surface glycoprotein that promotes leukocyte adhesion in inflammatory conditions (Rothlein et al. 1986; Springer 1990). Analysis of the ICAM-1 complementary deoxyribonucleic acid sequence has revealed it to be a member of the immunoglobulin gene superfamily (Staunton et al. 1988). ICAM-1 is expressed basally at low levels on many cell types, including endothelial cells, macrophages, myocytes, and vascular smooth muscle cells, but can be induced to high levels by stimulation with LPS, phorbol ester, or inflammatory cytokines, such as TNF-α or interleukin-1β (Colic and Drabek 1991; Ruetten et al. 1999).
To confirm the transcriptional inhibitory function of compounds 1, 2, 4, 8, 15, 17, and 18; we further investigated their effects on ICAM-1 and iNOS gene expression in TNFα-stimulated HepG2 cells using RT-PCR. Consistent with their inhibitory activity toward NF-κB, compounds 1, 2, 4, 8, 15, 17, and 18 significantly inhibited the induction of ICAM-1 and iNOS mRNA in a dose-dependent manner (see Fig. 4), indicating that these compounds reduced transcription of these genes. Moreover, the housekeeping protein β-actin was unchanged by the presence of compounds 1, 2, 4, 8, 15, 17, and 18 at the same concentration (see Fig. 4). Our data suggest that compounds 1, 2, 4, 8, 15, 17, and 18 isolated from the soft coral S. maxima have therapeutic potential as anti-inflammatory, anti-atherosclerotic, and anti-arthritic substances. However, elucidation of the detailed mechanisms underlying the inhibition of the TNFα-induced NF-κB pathway and subsequent decreases in ICAM-1 and iNOS gene expression by compounds 1, 2, 4, 8, 15, 17, and 18 requires further investigation.

Inhibitory effects of compounds 1–19 on TNFα-induced expression of inducible nitric oxide synthase (iNOS) and intercellular adhesion molecule-1 (ICAM-1) mRNAs in HepG2 cells. Effects of compounds 1, 2, 4, 8, 15, 17 and 18 on ICAM-1 and iNOS mRNA expression in HepG2 cells were assessed. HepG2 cells were pretreated with one of the listed compounds for 1 h and then treated with TNFα (10 ng/mL) for 6 h. Total mRNAs were prepared from the cell pellets using easy-BLUE™ (iNtRON Biotechnology, Seoul)
Consideration of the structure–activity relationship of these compounds indicated that the ketone groups at C-3 and/or C-6, the hydroxyl group at C-7 and/or C-8 are necessary for the anti-inflammatory activity of diterpenoids and norditerpenoids. This finding was confirmed by comparing the structure and activity of compounds 1, 2, 4, 8, 15, 17, and 18, therefore this information may facilitate identification of anti-inflammatory lead compounds from diterpenoids and norditerpenoids. This primary finding provides support for further studies on these compounds for the development of anti-inflammatory agents.
References
Albert, S., and J. Baldwin. 1996. The NF-κB and IκB Proteins: new discoveries and insights. Annual Review of Immunology 14: 649–681.
Baldwin, A.S. 2001a. Control of oncogenesis and cancer therapy resistance by the transcription factor NF-κB. The Journal of Clinical Investigation 107: 241–246.
Baldwin, A.S. 2001b. Series introduction: the transcription factor NF-κB and human disease. The Journal of Clinical Investigation 107: 3–6.
Chao, C.H., K.J. Chou, C.Y. Huang, Z.H. Wen, C.H. Hsu, Y.C. Wu, C.F. Dai, and J.H. Sheu. 2011. Bioactive cembranoids from the soft coral Sinularia crassa. Marine Drugs 9: 1955–1968.
Chen, F., V. Castranova, X. Shi, and L.M. Demers. 1999. New insights into the role of nuclear factor-κB, a ubiquitous transcription factor in the initiation of diseases. Clinical Chemistry 45: 7–17.
Cheng, S.Y., C.T. Chuang, S.K. Wang, Z.H. Wen, S.F. Chiou, C.H. Hsu, C.F. Dai, and C.Y. Duh. 2010. Antiviral and anti-inflammatory diterpenoids from the soft coral Sinularia gyrosa. Journal of Natural Products 73: 1184–1187.
Chung, W.Y., J.H. Park, M.J. Kim, H.O. Kim, J.K. Hwang, S.K. Lee, and K.K. Park. 2007. Xanthorrhizol inhibits 12-O-tetradecanoylphorbol-13-acetate-induced acute inflammation and two-stage mouse skin carcinogenesis by blocking the expression of ornithine decarboxylase, cyclooxygenase-2 and inducible nitric oxide synthase through mitogen-activated protein kinases and/or the nuclear factor-κB. Carcinogenesis 28: 1224–1231.
Colic, M., and D. Drabek. 1991. Expression and function of intercellular adhesion molecule 1 (ICAM-1) on rat thymic macrophages in culture. Immunology Letters 28: 251–257.
Grote, D., H.M. Dahse, and K. Seifert. 2008. Furanocembranoids from the soft corals Sinularia asterolobata and Litophyton arboreum. Chemistry and Biodiversity 5: 2449–2456.
Kamel, H.N., D. Ferreira, L.F.G. Fernandez, and M. Slattery. 2007. Cytotoxic diterpenoids from the hybrid soft coral Sinularia maxima × Sinularia polydactyla. Journal of Natural Products 70: 1223–1227.
Kim, K.K., K.S. Park, S.B. Song, and K.E. Kim. 2010. Up regulation of GW112 gene by NF-κB promotes an antiapoptotic property in gastric cancer cells. Molecular Carcinogenesis 49: 259–270.
Kleinert, H., A. Pautz, K. Linker, and P.M. Schwarz. 2004. Regulation of the expression of inducible nitric oxide synthase. European Journal of Pharmacology 500: 255–266.
Lakshmi, V., and R. Kumar. 2009. Metabolites from Sinularia species. Natural Product Research 23: 801–850.
Lo, K.L., A.T. Khalil, Y.H. Kuo, and Y.C. Shen. 2009. Sinuladiterpenes A-F, new cembrane diterpenes from Sinularia flexibilis. Chemistry and Biodiversity 6: 2227–2235.
Lu, Y., J.H. Su, C.Y. Huang, Y.C. Liu, Y.H. Kuo, Z.H. Wen, C.H. Hsu, and J.H. Sheu. 2010. Cembranoids from the soft corals Sinularia granosa and Sinularia querciformis. Chemical and Pharmaceutical Bulletin 58: 464–466.
Mattson, M.P., and S. Camandola. 2001. NF-κB in neuronal plasticity and neurodegenerative disorders. The Journal of Clinical Investigation 107: 247–251.
Nabel, G., and D. Baltimore. 1987. An inducible transcription factor activates expression of human immunodeficiency virus in T cells. Nature 326: 711–713.
Perkins, N.D. 2007. Integrating cell-signalling pathways with NF-κB and IKK function. Nature Reviews Molecular Cell Biology 8: 49–62.
Quang, T.H., N.T.T. Ngan, C.V. Minh, P.V. Kiem, B.H. tai, N.P. Thao, S.B. Song, and Y.H. Kim. 2012. Anti-inflammatory and PPAR transactivational effects of secondary metabolites from the roots of Asarum sieboldii. Bioorganic and Medicinal Chemistry Letters 22: 2527–2533.
Robinson, S.M., and D.A. Mann. 2010. Role of nuclear factor κB in liver health and disease. Clinical Science 118: 691–705.
Rothlein, R., M.L. Dustin, S.D. Marlin, and T.A. Springer. 1986. A human intercellular adhesion molecule (ICAM-1) distinct from LFA-1. Mediators of Inflammation 137: 1270–1274.
Ruetten, H., C. Thiemermann, and M. Perretti. 1999. Upregulation of ICAM-1 expression on J774.2 macrophages by endotoxin involves activation of NF-κB but not protein tyrosine kinase: comparison to induction of iNOS. Mediators of Inflammation 8: 77–84.
Springer, T.A. 1990. Adhesion receptors of the immune system. Nature 346: 425–434.
Staunton, D.E., S.D. Marlin, C. Stratowa, M.L. Dustin, and T.A. Springer. 1988. Primary structure of ICAM-1 demonstrates interaction between members of the immunoglobulin and integrin supergene families. Cell 52: 925–933.
Su, J.H., Y.F. Lin, Y. Lu, H.C. Yeh, W.H. Wang, T.Y. Fan, and J.H. Sheu. 2009. Oxygenated cembranoids from the cultured and wild-type soft corals Sinularia flexibilis. Chemical and Pharmaceutical Bulletin 57: 1189–1192.
Su, J.H., and Z.H. Wen. 2011. Bioactive cembrane-based diterpenoids from the soft coral Sinularia triangular. Marine Drugs 9: 944–951.
Surha, Y.J., K.S. Chuna, H.H. Cha, S.S. Han, Y.S. Keum, K.K. Park, and S.S. Lee. 2001. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: down-regulation of COX-2 and iNOS through suppression of NF-κB activation. Mutation Research 480–481: 243–268.
Thao, N.P., N.H. Nam, N.X. Cuong, T.H. Quang, P.T. Tung, B.H. Tai, B.T.T. Luyen, D. Chae, S. Kim, Y.S. Koh, P.V. Kiem, C.V. Minh, and Y.H. Kim. 2012. Diterpenoids from the soft coral Sinularia maxima and their inhibitory effects on lipopolysaccharide-stimulated production of proinflammatory cytokines in bone marrow-derived dendritic cells. Chemical and Pharmaceutical Bulletin 60: 1581–1589.
Thao, N.P., N.H. Nam, N.X. Cuong, T.H. Quang, P.T. Tung, L.D. Dat, D. Chae, S. Kim, Y.S. Koh, P.V. Kiem, C.V. Minh, and Y.H. Kim. 2013a. Anti-inflammatory norditerpenoids from the soft coral Sinularia maxima. Bioorganic and Medicinal Chemistry Letters 23: 228–231.
Thao, N.P., N.H. Nam, N.X. Cuong, B.H. Tai, T.H. Quang, N.T.T. Ngan, B.T.T. Luyen, S.Y. Yang, C.H. Choi, S. Kim, D. Chae, Y.S. Koh, P.V. Kiem, C.V. Minh, and Y.H. Kim. 2013b. Steroidal constituents from the soft coral Sinularia dissecta and their inhibitory effects on lipopolysaccharide-stimulated production of pro-inflammatory cytokines in bone marrow-derived dendritic cells. Bulletin of the Korean Chemical Society 34: 949–952.
Acknowledgments
This study was supported by a Grant from Vietnam Academy of Science and Technology (Project No: VAST.TĐ.ĐAB.02/13-15) and by a grant from the Priority Research Center Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2009-0093815), Republic of Korea.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Thao, N.P., Nam, N.H., Cuong, N.X. et al. Inhibition of NF-κB transcriptional activation in HepG2 cells by diterpenoids from the soft coral Sinularia maxima . Arch. Pharm. Res. 37, 706–712 (2014). https://doi.org/10.1007/s12272-013-0230-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12272-013-0230-3