
Abstract
The release of phosphorus (P) stored in the sediment may cause long-term delay in the recovery of lakes, ponds, and lagoons from eutrophication. In this paper, we tested on a laboratory scale the efficacy of the flocculant polyaluminium chloride (PAC) and a strong P-binding agent (lanthanum-modified bentonite, LMB) on their ability to flocculate a cyanobacterial bloom and hamper P release from a hypertrophic, brackish lagoon sediment. In addition, critical P loading was estimated through PCLake. We showed that cyanobacteria could be effectively settled using a PAC dose of 2 mg Al L−1 combined with 400-mg L−1 LMB; PAC 8 mg Al L−1 alone could also remove cyanobacteria, although its performance was improved adding low concentrations of LMB. The efficacy of LMB to bind P released from the sediment was tested based on potentially available sediment P. A dose of 400 g LMB m−2 significantly reduced the P release from sediment to over-standing water (either deionized water or water from the lagoon with and without cyanobacteria). In sediment cores, LMB + PAC reduced sediment P flux from 9.9 (± 3.3) to − 4.6 (± 0.3) mg P m−2 day−1 for the experimental period of 3 months. The internal P load was 14 times higher than the estimated P critical load (0.7 mg P m−2 day−1), thus even if all the external P sources would be ceased, the water quality will not improve promptly. Hence, the combined LMB + PAC treatment seems a promising in-lake intervention to diminish internal P load bellow the critical load. Such intervention is able to speed up recovery in the brackish lagoon once external loading has been tackled and at a cost of less than 5% of the estimated dredging costs.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Eutrophication is one of the main anthropogenic stressors leading to major degradation of coastal waters worldwide (Kennish 2002). Water quality problems caused by eutrophication include fish deaths due to anoxia, loss of biodiversity, bad smells, and massive plant growth (Paerl and Huisman 2008; Conley et al. 2009). Key symptom of eutrophication is a blooming of harmful algae and cyanobacteria, which pose an additional risk to wildlife and humans because of the toxins they may produce (Correl 1998; Huszar et al. 2000; Paerl and Paul 2012). Hence, there is a great need to control these nuisance blooms.
Since blooms are fueled by nutrients, the first step in mitigation would be reducing the nutrient discharge into the receiving waters (Cooke et al. 2005; Paerl et al. 2014). Although some waters will clear up and recover rapidly from such lowered external nutrient loading, many will show a considerable delay in recovery due to internal nutrient cycling (Jeppesen et al. 1991; Søndergaard et al. 1999; Søndergaard et al. 2001; Cooke et al. 2005). This legacy of nutrients created from decades of uncontrolled excessive external nutrient loading will periodically be recycled between sediment and water column and is viewed as one of the main reasons why many restoration attempts have failed (Gulati and Van Donk 2002). Removal of polluted sediments is then a straightforward restoration approach, but it may come with relatively high costs compared to other techniques such as in situ fixation or capping (Cooke et al. 2005; Perelo 2010).
In recent years, the use of solid phase phosphorus (P)-adsorbing compounds has gained interest to tackle the widespread internal loading issue (Spears et al. 2013). The rationale to target stored P lays in the fact that it is the only essential element that can be easily be made to limit algal growth through the formation of insoluble precipitates (Golterman 1975). Internal P loading not only is a major issue in inland freshwater systems (Søndergaard et al. 2001) but also occurs in brackish coastal lagoons (Markou et al. 2007). While there is a growing number of studies demonstrating efficacy and applicability of solid-phase P sorbents in freshwater systems, studies on brackish coastal waters are virtually lacking. Nonetheless, eutrophication is considered the most common problem affecting coastal lagoons (Esteves et al. 2008; Kennish et al. 2014). For instance, the coastal system Jacarepaguá lagoon in the western part of Rio de Janeiro city (Brazil) suffers heavily from eutrophication and perennial presence of cyanobacterial blooms (Gomes et al. 2009; De-Magalhães et al. 2017).
Recently, we have explored the possibility of removing cyanobacteria from Jacarepaguá lagoon water using a coagulant (poly-aluminium chloride, PAC, or chitosan) and red soil or local sediment as ballast (De-Magalhães et al. 2017). While PAC was effective, chitosan appeared ineffective to flock cells even when combined with ballast compounds (De-Magalhães et al. 2017). Elevated pH and high alkalinity were identified as factors that may hamper the coagulation of chitosan and impair its ability to effectively remove cyanobacteria from the water column (Lürling et al. 2017).
In the present study, we elaborated on these findings and first tested the combination of PAC and the solid phase P adsorbent Phoslock®, which is a lanthanum-modified bentonite (LMB) with strong P binding capacity and widely used in freshwater systems (Copetti et al. 2016), on the ability to remove cyanobacteria from the brackish water of Jacarepaguá lagoon. In addition, we were particularly interested in the performance of LMB as Copetti et al. (2016) reported that even moderately saline environments of > 0.5 ppt will render LMB ineffective. A thorough scientific underpinning of this statement is, however, lacking in that review paper (Copetti et al. 2016) and also finds no support in the few studies that included more saline environments (Haghseresht 2006; Reitzel et al. 2013). Given the current uncertainty on applicability of LMB in brackish environments, we tested the hypotheses that (1) LMB will block P release from the sediment of the eutrophic coastal lagoon Jacarepaguá and (2) that a combination of PAC with LMB will clear the water and block the P release effectively.
Material and Methods
Study Ecosystem
The Jacarepaguá lagoon (43° 17′–43° 30′ W, 22° 55′–23° 00′ S) is part of a brackish water lagoon complex located in the western part of Rio de Janeiro City (Fig. 1). The Jacarepaguá lagoon is 3.7 km2 in area; it has an average depth of 3.3 m; drainage area of 103 km2; and the freshwater inflow from the six tributaries is about 0.8 m3 s−1 (Gomes et al. 2009). This system has a direct communication with the sea water by the Joatinga channel, giving an average salinity of 5.35 ppt (De-Magalhães et al. 2017). The lagoon usually presents high pH and alkalinity with perennial relatively high chlorophyll-a concentrations (mostly exceeding 100 μg L−1) and long periods of cyanobacteria dominance promoted by the constant sewage input (Gomes et al. 2009; De-Magalhães et al. 2017).

a Location of the Jacarepaguá lagoon near to the Olympic 2016 venues and the sediment sampling station (JAC 20). b The green water of the lagoon (January 19th 2015) and c the main phytoplankton species (M. aeruginosa colonies and P. agardhii filaments)
Sediment and Water Sampling
On November 2014, 10 L of surface water was collected for experiments with coagulants and ballast. The most important species in this moment was Microcystis aeruginosa, and the chlorophyll-a concentration was 225 μg L−1. Jacarepaguá sediment was collected with a Kajak sediment core sampler on January 19, 2015 at station JAC20 (Fig. 1). At this moment, the chlorophyll-a concentration, collected by an integration tube, was 226 μg L−1 composed mainly by M. aeruginosa with some undergrowth of Planktothrix agardhii (Fig. 1). The pH of the water was 9.04 (± 0.24); salinity was 5.49 ppt and the alkalinity 4.35 mEq L−1. On September 2015, more sediment was collected using the gravity Uwitec Corer sampler at station JAC20, and at this moment, the pH was 9.88, salinity 5.17 ppt, and the alkalinity was 3.74 mEq L−1. No cyanobacteria bloom was observed, and the phytoplankton community was composed mainly by Cryptophyceae and green algae.
Chemicals and Materials
The lanthanum-modified bentonite Phoslock® (LMB) was obtained from HydroScience (Porto Alegre, Brazil). This LMB was developed by the Australian CSIRO, as dephosphatization technique aiming at removing soluble reactive phosphorus (SRP) from the water and blocking the release of SRP from the sediment (Douglas 2002). The coagulant PAC-AP (polyalumnium chloride; Aln(OH)mCl3n-m, ρ ≈ 1.37 kg L−1, 8.9% Al, 21.0% Cl) was obtained from Pan-Americana (Rio de Janeiro, Brazil).
Effect of Different Concentrations of LMB and PAC on Cyanobacteria Removal
The first experiment tested the efficacy of a combination of LMB with PAC to settle the cyanobacteria from Jacarepaguá water. Different concentrations of LMB (0 to 400 mg L−1) in the presence of two fixed doses of PAC were used. The low PAC dose (2 mg Al L−1) was based on the results from previous experiments in freshwaters (Lürling and van Oosterhout 2013; Noyma et al. 2016); the higher PAC dose (8 mg Al L−1) was based on the effective removal of cyanobacteria from Jacarepaguá water with red soil as ballast (De-Magalhães et al. 2017). The experiment was run in 75-mL glass tubes that were filled with 60 mL of unfiltered water from Jacarepaguá. The water collected from Jacarepaguá contained cyanobacteria at a chlorophyll-a concentration of 222 (± 2) μg L−1; the cells were healthy as indicated by a photosystem II (PSII) efficiency of 0.53 (± 0.03), both determined using a PHYTO-PAM phytoplankton analyzer (Heinz Walz GmbH, Effeltrich, Germany). The experiment included a control without any compound added and was performed with three replicates per treatment. Immediately after adding the designated amount of LMB, the PAC coagulant was added and the content in the test tube mixed briefly using a glass rod. Tubes were placed in the laboratory at 25 °C under stagnant conditions. After 1 h, 5-mL samples were taken from both the top and the bottom of the tubes in which chlorophyll-a concentrations and PSII efficiencies were measured. The 5 mL from the top and the bottom of the tubes was sampled, since an accumulation at the top would indicate a scum formation in the field, which is an unwanted effect, whereas the accumulation at the bottom is the intended effect from the combined coagulant and ballast (De-Magalhães et al. 2017; Miranda et al. 2017). After the top and bottom samples were taken, the pH was measured in the middle of the tubes. The chlorophyll-a concentrations in the top of the tubes and those measured at the bottom of the test tubes, as well as PSII-efficiencies and pH values were statistically evaluated running a one-way ANOVA in the program SigmaPlot version 13. Homogeneity of variance was tested by the equal variance test (Brown-Forsythe) and normality, by the Shapiro-Wilk normality test. In cases where normality failed data were log-transformed to fulfill this prerequisite, an all pairwise multiple comparison was performed to distinguish means that were significantly different at the 0.05 level (Holm-Sidak method; p = 0.05).
LMB Dose
The manufacturers advice to dose the LMB in an LMB:P ratio 100:1, with P the “labile” P-pool in the sediment. The ratio LMB:P 100:1 is based on the 1:1 molar La:P from the precipitation reaction equaling a 4.485:1 La:P weight ratio and a 4.5% La in LMB. Based on the 0.05 g P/kg (wet sediment), we estimated a dose of 400 and 507.5 g LMB m−2 assuming a communicating sediment depth of 8 and 10 cm, respectively, which are consistent with LMB doses applied in the field (Dithmer et al. 2016).
Sediment P Extraction
To determine the dose of LMB needed in the experiments, an estimate of the potentially releasable P in the sediment was required. Hereto, a sequential extraction protocol modified from Paludan and Jensen (1995) and used by Cavalcante et al. (2018) to measure different P forms in the sediment was adopted. One gram of wet sediment was brought into each of four 50-mL Falcon tubes to which, as a first step, 25-mL anoxic demineralized water was added to extract the immediately available P. The tubes were shaken for 30 min (oxygen at the start was 0.21 and at the end 0.44 mg L−1). The tubes were centrifuged and the supernatant collected. A second aliquot of 25-mL anoxic demineralized water was added to the pellets and shaken for 5 min, where after the tubes were centrifuged and the supernatants joined, filtered through 0.6-μm glass fiber filters (GF-3, Macherey-Nagel), acidified with 0.5-mL 2-M H2SO4 and stored in the refrigerator until P analysis. In the second step, to the pellets 25 mL of anoxic bicarbonate/dithionite (BD: 0.11-M NaHCO3 and 0.11-M Na2S2O4) was added to extract P bound to Fe-hydroxides and Mn-compounds from the sediment pellets. The tubes were shaken for 30 min, subsequently centrifuged, and the supernatant collected. To the pellets, another 22-mL anoxic BD was added and tubes were shaken for 5 min, centrifuged, and supernatants joined. The joined 47-mL supernatants were aerated for half an hour, filtered through 0.6-μm glass fiber filters, acidified with 3-mL 2-M H2SO4, and stored in the refrigerator for P analysis. In the third and last step, to the pellets, 25-mL 0.1-M NaOH was added aiming to extract P bound to metal oxides of AI. The tubes were shaken for 30 min, centrifuged, and supernatants collected, followed by a second extraction with 25-mL 0.1-M NaOH for 5 min and a washing step for 5 min with 23.5-mL demineralized water. The three joined supernatants (73.5 mL) were filtered as before, acidified with 1.5-mL 2-M H2SO4, and stored in the refrigerator. The filtrates were analyzed on their SRP and total phosphorus (TP) concentrations using a flow injection analysis system (model 2500, FIAlab, USA). The dry weight of the sediment was determined by weighing triplicate samples of 10-mL sediment before and after drying at 105 °C.
Effect of Different Over-standing Water on Sediment Phosphate Release
Fifty grams wet sediment from Jacarepaguá, corresponding to a 2.43 mg of releasable P in the sediment, considering the P content determined as described above, was transferred into 250-mL Schott glass bottles. To six bottles 100-mL demineralized water was added, to nine bottles 100-mL filtered Jacarepaguá water (0.6-μm glass fiber filters; GF-3, Macherey-Nagel) was added, while to nine other bottles 100-mL unfiltered Jacarepaguá water was added, which was collected on January 19, 2015. Three bottles of each series were left untreated (control) and three were treated with 400-g m−2 LMB, while the two series with filtered and unfiltered Jacarepaguá water also included a treatment with PAC (8 mg Al L−1) and LMB (400 g m−2) in triplicates. This dose of PAC was found effective in flocculating the cyanobacteria out of the water column without strong effects on the pH of Jacarepaguá water (De-Magalhães et al. 2017). PAC was not included in the demineralized water series, because of strong effects on pH (Gebbie 2001). We calculated a dose of 400 g LMB m−2 assuming a communicating sediment depth of 8 cm, which is consistent to the La profile in 10 LMB treated lakes where La was mixed in the sediment from ~ 5 cm to more than 10 cm (Dithmer et al. 2016). The experimental bottles were placed at 25 °C at low light (≅ 1 μmol photon m−2 s−1) in day-night regime (13-h light:11-h dark). Initially and after 7, 14, and 21 days, samples were taken, filtered through 0.6-μm glass fiber filters (GF-3, Macherey-Nagel), and analyzed on their SRP concentrations using a flow injection analysis system (model 2500, FIAlab, USA). Differences in SRP concentrations between start and 1 week incubations were used to derive an estimate of SRP fluxes using the known water volume (100 mL) and the surface area of the sediment at the Schott glass bottles (28.27 cm2).
Treating Sediment Cores with PAC or LMB + PAC—Short-Term Experiment
On January 19, 2015, seven sediment cores were drilled from Jacarepaguá using a Kajak core sampler. The cores contained between 18- and 30-cm length of black sediment and 9- to 21-cm over-standing, cyanobacteria dominated water. Hereto, considering a communicating sediment depth of 8 cm and the results of the extraction described above, the cores contain an estimated amount of 10.15 mg of P releasable to the water column. Two cores were treated with sole PAC (8 mg Al L−1), two cores with LMB (400 g m−2) plus PAC (8 mg Al L−1), while three cores remained untreated (controls). The cores were incubated in the laboratory at 25 °C at low light (≅ 1 μmol photon m−2 s−1) in day-night regime (13-h light:11-h dark). Initially and after 1.5, 3.5, 18, 42, 90, 138, 186, and 306 h, water samples were taken and analyzed on their chlorophyll-a concentrations. Additional samples taken before, just after application and after 138 and 306 h incubation, were filtered through 0.6-μm glass fiber filters (GF-3, Macherey-Nagel) and analyzed on their SRP concentration as previously described. The differences between SRP concentrations from the start and after incubation of 306 h were used to estimate the SRP fluxes using the formula: {(Pfinal − Pstart) × water height} / Δt, with P in mg m−3, water height in m, and Δt in days (day).
Treating Sediment Cores with LMB + PAC—Long-Term Experiment
On September 29, 2015, additional sediment cores were taken from Jacarepaguá using a gravity Uwitec Corer sampler. The tubes contained between 18- and 28-cm length of black sediment and 32 to 43 cm of over-standing water. The potential available P was determined as outlined above and the SRP concentration in the water was determined. Both were used to estimate the dose of LMB required assuming a 10-cm communicating sediment depth. The 10-cm communicating sediment depth contain a calculated amount of 12.68 mg of P releasable yielding a dose of 507.5 g m−2 to be added together with PAC (8 mg Al L−1) to each of four replicate cores (treatment), while four other cores remained untreated (controls). The chlorophyll-a concentration of the over-standing water was 86 (± 1) μg L−1. The cores were closed with a rubber stopper and placed in the laboratory at 25 °C in the dark. The experiment lasted 96 days to give insight in the durability and efficacy of the treatment. The experiment was conducted under anoxia 0.20 (± 0.45) mg L−1 at a circumneutral pH 7.01 (± 0.54). Ten milliliters of water from the middle of the core tubes were sampled initially and after 1, 3, 15, 22, 29, 35, 64, and 96 days and filtrated before been analyzed using a flow injection analysis system (model 2500, FIAlab, USA) for SRP measurements. The treatment took place on October 1, i.e., 2 days after collection, because the sediment P had to be determined prior to application. Consequently, the course of SRP concentrations was statistically evaluated running a rmANOVA in the toolpack SPSS (version 22) using the whole period as well as using only the data obtained after application (days 3, …, 96). The differences between SRP concentrations from start and after 96 days of incubation were used to estimate the SRP fluxes using the formula: {(Pfinal − Pstart) × water height} / Δt, with P in mg m−3, water height in m, and Δt in days (day).
Comparison of SRP Fluxes with Estimated Critical P Loadings
The PCLake Metamodel is used to estimate the critical P load sufficient to cause a shift between a clear water state (P load below its critical value) and a turbid water state (P load above its critical value) (Mooij et al. 2010, PBL 2015), available at http://themasites.pbl.nl/modellen/pclake/index.php. In the clear water state, blooms of cyanobacteria are not expected as they are P-limited. PCLake simulates the influence of phosphate on lakes based on water and sediment P, transparency, amount of water plants, phytoplankton concentration, fish stock, and swamp and bank vegetation, while taking into consideration soil type, size, and depth of a lake. PCLake simulations have been run by the model builders using a whole range of P loads for a number of lake types and both starting conditions. In these 100,000 simulations, depth, lake surface, retention time, soil type, and swamp area defined the lake types. All results are stored in a database and the critical transitions determined for each combination. The critical transition is the P-load yielding a transparency of half the water column depth. In case of new combinations, the critical P loads are estimated using a neural network (http://www.pbl.nl/dossiers/water/modellen/WerkingModelPCLake). PCLake was run with the following parameters based on previous Jacarepaguá lagoon studies (Barbosa and Almeida 2001; Ferrão-Filho et al. 2002; Gomes et al. 2009). The input parameters were as follows: average depth of 3.3 m, swamp area 0.1, fetch 4000 m, discharge (19 mm day−1 = residence time of 176 days), average depth = 3.3 m, background extinction = 0.5 m−1, and sand as soil type. We compare the SRP fluxes derived from our current experiments to the PCLake critical P loadings. An additional critical P load estimate was made, targeting a TP concentration of 30 μg L−1, which correspond to a decrease of 98% of TP in Jacarepaguá lagoon (De-Magalhães et al. 2017), using the Vollenweider (1976) model: Pcritical = Ptarget × (1 + √τ) × zm × τ−1, where Pcritical is the critical P load (g m−2 year−1), Ptarget is the target in-lagoon P concentration (g m−3), τ is the water retention time (year), and zm is the mean water depth (m). The same model was used to derive an estimate of the actual load based on the current in-lagoon P concentration.
Results
Effect of Different Concentrations of LMB and PAC on Cyanobacteria Removal
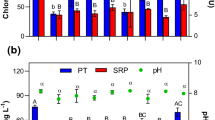
With the PAC dose fixed at 2 mg Al L−1, the chlorophyll-a concentrations in the top of the test tubes declined with increasing LMB dose. The F test revealed a significant difference among the treatments (F5,12 = 135.0; p < 0.001). The pairwise multiple comparison revealed no difference between the control and the sole PAC treatment (0 mg LMB L−1), while with higher LMB dose, all chlorophyll-a concentrations in the top were significantly different and decreased with higher concentrations of LMB as ballast (Fig. 2). Also, in the bottom of the test tubes, significantly different (F5,12 = 495.4; p < 0.001) chlorophyll-a concentrations were found. The post hoc comparison, considering the top of the bottles revealed four homogenous groups that were significantly different from each other: (1) the lowest chlorophyll-a concentrations were in the control and the 0-mg LMB L−1 treatment; (2) significantly higher chlorophyll-a concentrations were measured in the bottom of the tubes treated with concentrations 50-mg LMB L−1 treatment; (3) even higher chlorophyll-a concentrations were measured in the bottom of the tubes treated with concentrations 100-mg LMB L−1 treatment; and (4) the highest chlorophyll-a concentrations were measured in the bottom of the tubes treated with 200 and 400 mg LMB L−1 (Fig. 2). PSII-efficiencies in the top of the tubes were also statistically different (F5,12 = 6.19; p = 0.005), the post hoc comparison revealed that the PSII in the 200-mg LMB L−1 treatment was lower than in the 100 and 400 mg LMB L−1. Nonetheless, values varied on average between 0.49 and 0.55. PSII efficiency was also statistically different in the bottom of the tubes (H5 = 13.8; p = 0.017). The pairwise multiple comparison test revealed that PSII in the control (0.59) was significantly higher than in the 400-mg LMB L−1 treatment (0.42) (Fig. 2). Although pH in the control was significantly higher than the treatments (F5,12 = 94.1; p < 0.001), the differences were relatively small varying from 9.1 (control) to 8.7 (400 mg LMB L−1; Fig. 2).

Chlorophyll-a concentrations (μg L−1) in the top 5 mL (top light gray bars) and bottom 5 mL (lower dark gray bars) of 60-mL cyanobacteria suspensions from Jacarepaguá lagoon incubated for 1 h in the absence (control) or presence of the coagulant PAC (2 mg Al L−1) and different concentrations of lanthanum-modified bentonite, LMB (0–400 mg L−1). Also included the photosystem II efficiency (PSII) of the cyanobacteria collected at the water surface (filled circles) and at the bottom (open circles). Error bars indicate one standard deviation (n = 3). Similar symbols (a, …, δ) above/below the bars indicate homogeneous groups that are not different at the 95% level (Holm-Sidak test)
In the series with PAC dosed at 8 mg Al L−1 and concentration range of LMB, chlorophyll-a concentrations in the top of the test tubes were significantly different among treatments (F5,12 = 66.7; p < 0.001). The post hoc comparison revealed four homogeneous groups: (1) the control; (2) the 0-mg LMB L−1 treatment, i.e., the sole PAC treatment; (3) the combined PAC and 50, 100, and 400-mg LMB L−1 treatments; and (4) the 100, 200, and 400-mg LMB L−1 treatments combined with PAC (Fig. 3). The bottom chlorophyll-a concentrations were also significantly different (F5,12 = 217.7; p < 0.001). Three significantly different groups were detected: (1) the control; (2) the 0, 50, 100, and 200-mg LMB L−1 treatments combined with PAC; and (3) the 400-mg LMB L−1 treatment also combined with PAC (Fig. 3). PSII-efficiencies in the top of the test tubes were similar (F5,12 = 2.78; p = 0.068) and on average 0.55 (± 0.05). In the bottom, the ANOVA indicated significant differences (F5,12 = 6.50; p = 0.004), where the post hoc comparison indicated PSII in the 400-mg LMB L−1 treatment was significantly lower than those in the 0-, 50-, and 100-mg LMB L−1 treatments. However, differences were very small, as were within group variations. The mean PSII-efficiency at the bottom was 0.53 (± 0.02) (Fig. 3). The pH was significantly different (F5,12 = 288.3; p < 0.001) and three different groups were found: (1) the control; (2) the 0-mg LMB L−1 treatment, i.e., the sole PAC treatment; and (3) all PAC + LMB treatments (Fig. 3).

Chlorophyll-a concentrations (μg L−1) in the top 5 mL (top light gray bars) and bottom 5 mL (lower dark gray bars) of 60-mL cyanobacteria suspension from Jacarepaguá lagoon incubated for 1 h in the absence (control) or presence of the coagulant PAC (8 mg Al L−1) and different concentrations of lanthanum-modified bentonite, LMB (0–400 mg L−1). Also included are the photosystem II efficiencies (PSII) of the cyanobacteria collected at the water surface (filled circles) and at the bottom (open circles) as well as the pH of the water (open triangles). Error bars indicate one standard deviation (n = 3). Similar symbols (a, …, γ) above/below the bars indicate homogeneous groups that are not different at the 95% level (Holm-Sidak test)
Effect of Different Over-standing Water on Sediment Phosphate Release
The initial SRP concentration in the Jacarepaguá lagoon was 786 μg L−1 in the filtered water and 783 μg L−1 in the unfiltered water, while it was below the detection limit (3 μg L−1) in the demineralized water. The SRP concentrations in the filtered and unfiltered lagoon water standing over Jacarepaguá sediment were reduced by treatments with LMB or PAC + LMB, while it was the same in the controls (Fig. 4). Contrary, in over-standing demineralized water, SRP concentrations seem to increase in treatments with LMB and control (Fig. 4); however, the rmANOVA indicated no time effect (F1.5,6.0 = 2.12; p = 0.200), no treatment effect (F1,4 = 1.60; p = 0.274), and no time × treatment interaction (F1.5,6.0 = 0.36; p = 0.655) (Fig. 4a). The SRP fluxes, from demineralized water, determined after 1-week incubation were not significantly different (t4 = 2.25; p = 0.065), despite they were on average 4.0 (± 2.5) mg P m−2 day−1 in the control and 0.4 (± 0.2) mg P m−2 day−1 in the LMB treatment (Fig. 5).

Phosphate (SRP) concentrations (μg L−1) after 7, 14, and 21 days in 100-mL demineralized water (a), filtered (b), and unfiltered (c) Jacarepaguá water standing above 50-g Jacarepaguá sediment that was untreated (controls) or treated with either LMB (400 g m−2) or PAC + LMB (PAC at 8 mg Al L−1). Error bars indicate one standard deviation (n = 3). The gray line represents the initial SRP values at day “0” in each type of water used

Estimated SRP fluxes in the different experiments and treatments performed in this study. Negative values indicate a net SRP removal from over-standing water and thus an accumulation in the sediment, whereas positive values indicate a net release from the sediment (internal loading). The green line represents the estimated critical transition from clear to turbid water, while the gray line represents the critical transition from turbid to clear based on the PCLake Metamodel
In the series where Jacarepaguá sediment was incubated with filtered lagoon water, the rmANOVA indicated no time effect (F2,12 = 1.15; p = 0.351), a significant treatment effect (F2,6 = 107.5; p < 0.001), and no time × treatment interaction (F4,12 = 0.79; p = 0.552). Tukey’s post hoc comparison revealed that SRP concentrations in the LMB and LMB + PAC treatments were significantly lower than in the controls (Fig. 4b). Likewise, the SRP fluxes were significantly different (F2,8 = 22.8; p = 0.002). Tukey’s test showed that controls differed from treatments with values of 2.3 (± 1.2) mg P m−2 day−1 in the control, − 2.7 (± 0.9) mg P m−2 day−1 in LMB treatment, and − 2.4 (± 0.9) mg P m−2 day−1 in LMB + PAC treatment. The negative values indicate a net removal of SRP from the over-standing water and thus a flux towards the sediment (Fig. 5).
The treatment effects in the series with sediment and unfiltered water were comparable to those obtained with filtered water (Fig. 4b, c). The rmANOVA indicated no time effect (F2,12 = 1.90; p = 0.192), a significant treatment effect (F2,6 = 13.1; p = 0.006), and no time × treatment interaction (F4,12 = 1.37; p = 0.301). Tukey’s post hoc comparison revealed that SRP in the LMB and LMB + PAC treatments was significantly lower than in the controls (Fig. 4). The SRP fluxes were significantly different (F2,8 = 8.87; p = 0.016), and Tukey’s test showed that the control differed from treatments with values of 6.1 (± 4.8) mg P m−2 day−1 in the control, − 2.5 (± 1.0) mg P m−2 day−1 in LMB treatment, and − 2.3 (± 0.9) mg P m−2 day−1 in LMB + PAC treatments. Again, the negative values indicate a net removal of SRP from the over-standing water and thus a flux towards the sediment (Fig. 5).
“Labile” P-Pool in Jacarepaguá Lagoon Sediment
The average value from the phosphate concentration sum, for all three extraction steps, was 362.5-μg P/g DW. The major part of the phosphorus was extracted in step 2, with BD (167.1 ± 27.1-μg P/g DW). The P sorbed by clay minerals and oxides of AI extracted using NaO contributed with 131.8 (± 20.5) μg P/g DW. Lower contribution of loosely adsorbed P (extracted with anoxic demineralized water) was observed in a concentration of 63.5 (± 12.1) μg P/g DW (Fig. 6). Considering 14% of dry weight in each ml of sediment, it yields a concentration of 51.5 μg P/ml.

Average concentrations of P-fractions (μg P/g DW) in Jacarepaguá Lagoon sediment core. Error bars indicate one standard deviation (n = 3). BD = strongly reducing reagent (anoxic bicarbonate/dithionite)
Treating Sediment Cores with PAC or LMB + PAC—Short-Term Experiment
The sediment cores treated with PAC + LMB or only PAC caused a rapid decline in both the chlorophyll-a and the SRP concentrations in the water column (Fig. 7a, b). Already after 1.5 h, chlorophyll-a concentrations in the sole PAC treatment were 59% lower than in the control, while in the LMB + PAC treatment, it was more than 90% lower. The rmANOVA indicated a significant time effect (F3.5,14.3 = 34.3; p < 0.001), a significant treatment effect (F2,4 = 11.1; p = 0.023), and a significant time × treatment interaction (F7.1, 14.3 = 3.84; p = 0.015). The chlorophyll-a concentrations in the control also gradually decreased to values similar as in the treatments (Fig. 7a). Tukey’s test revealed that only the control and the LMB + PAC treatments were significantly different from each other. The SRP concentrations were strongly influenced by the treatments where PAC reduced the SRP concentrations by 72% within 1.5 h, while LMB + PAC caused a 92% reduction (Fig. 7b). However, SRP concentrations in PAC treatments started to increase again and after 306 h, they were similar to the control (t3 = 1.25; p = 0.299), while SRP in the PAC + LMB treatments was still significantly lower (Tukey’s test following one-way ANOVA; F2,6 = 49.2; p = 0.002). The SRP fluxes were on average 9.2 (± 6.7) mg P m−2 day−1 in the control, 3.6 (± 1.7) mg P m−2 day−1 in PAC treatment, and − 10.4 (± 5.5) mg P m−2 day−1 in LMB + PAC treatment. The ANOVA indicated that SRP fluxes were significantly different (F2,6 = 7.70; p = 0.043) (Fig. 5), but this was not confirmed by the Tukey post hoc comparison yielding a marginal difference between the control and LMB + PAC treatment (p = 0.052).

Course of the chlorophyll-a concentrations (upper panel a) and of SRP concentrations (lower panel b) in a short-term experiment in which sediment cores from Jacarepaguá lagoon (collected January 19th 2015) were left untreated (control; n = 3) or were treated with either PAC (8 mg L−1; n = 2) or with PAC (8 mg L−1) and LMB (400 g m−2; n = 2)
Treating Sediment Cores with LMB + PAC—Long-Term Experiment
When sediment cores from Jacarepaguá were treated with PAC + LMB a strong reduction in SRP concentrations could be observed (Fig. 8). The rmANOVA on SRP data over the whole period indicated a significant time effect (F3.4,16.8 = 7.29; p = 0.002), a significant treatment effect (F1,5 = 136.0; p < 0.001), and a significant time × treatment interaction (F3.4,16.8 = 16.4; p < 0.001). To check whether the interaction effect was caused by the initial data obtained prior to the treatment (days 0 and 1), an additional rmANOVA was run on data after the application (days 3, …, 96). This rmANOVA yielded similar results; a significant time effect (F3.3,16.7 = 7.22; p = 0.002), a significant treatment effect (F1,5 = 146.7; p < 0.001), and a significant time × treatment interaction (F3.3,16.7 = 7.62; p = 0.002). The time × treatment interaction effect was caused by the gradual increase in SRP in the controls, while SRP in the treatments remained equally low over the course of the experiment (Fig. 8). SRP in the treatments was on average only 2% of the values in the control. The SRP fluxes were 9.9 (± 3.3) mg P m−2 day−1 for the control and − 4.6 (± 0.3) mg P m−2 day−1 for the PAC + LMB treatments (Fig. 5).

Course of the SRP concentrations in sediment cores collected on September 29th 2015 in Jacarepaguá lagoon that were left untreated (control; n = 4) or were treated with PAC (8 mg L−1) and LMB (507.5 g m−2; n = 4)
Comparison of SRP Fluxes with Estimated Critical P Loadings
Based on the output from the PCLake Metamodel, a critical SRP flux of 0.7 mg P m−2 day−1 indicates the shift from a clear to turbid stable state in Jacarepaguá lagoon. While after the turbid stable state is established, only decreasing the SRP flux to values lower than 0.2 mg P m−2 day−1 would shift the water from the turbid to clear. The Vollenweider (1976) model yielded -for a target in-lagoon P concentration of 30 μg L−1—a critical P load of 0.95 mg P m−2 day−1 (Table 1). These values are considerably lower than the fluxes that have been estimated in the controls of the different experiments conducted and higher than the LMB treatments in this study (Fig. 5).
Discussion
The results of this study are in agreement with our hypotheses that LMB will block P release from the sediment of the eutrophic coastal lagoon Jacarepaguá and that a combination of PAC with LMB also clears the water effectively from cyanobacteria. The LMB strongly reduced the internal loading from the nutrient rich sediment from Jacarepaguá lagoon. These results add to the growing body of evidence that LMB is an effective eutrophication management agent. Meanwhile, LMB has been used in dozens of freshwater systems where it in general led to an improved water quality (e.g., Copetti et al. 2016; Spears et al. 2013, 2016; Epe et al. 2017). In freshwater lakes with cyanobacterial blooms, the combination of LMB with a coagulant clearly improved water quality through effective control of the bloom and internal SRP loading (Lürling and Van Oosterhout 2013; Waajen et al. 2016a). A growing number of studies show that LMB is effective in reducing the SRP efflux from freshwater sediments (e.g., Waajen et al. 2016a, b), but reports on its efficacy in eutrophic coastal lagoons is still limited. Few studies have reported that LMB could adsorb SRP effectively in saline water (Haghseresht 2006; Zamparas et al. 2012) and brackish water (Reitzel et al. 2013), while reports on effective SRP efflux reduction from brackish and saline sediments are even more rare (Douglas et al. 1999). Hence, our study is one of the few that demonstrates the effectiveness of LMB in hampering the SRP efflux from a brackish sediment and the first that shows that combined with PAC and LMB can control the sediment SRP release for at least 3 months. In our short-term experiment, we could see that PAC alone (at 8 mg Al L−1) was ineffective in hampering sediment P efflux as within 2 weeks SRP was as high as in the control, but when combined with LMB, the SRP remained at 2% of the control for the entire experimental period of 3 months.
Our results also refute the claim that even moderately saline environments of > 0.5 ppt will render LMB ineffective, because La would be freed from the clay matrix and therewith prohibit formation of a reactive layer for the absorption of labile P species at the sediment-water interface (Copetti et al. 2016). Liberation of some of the La from the clay matrix is likely in more ion rich environments, but formation of “soluble La species” (Copetti et al. 2016) is less likely, since any La will immediately react with oxyanions in the water to form complexes (Byrne and Kim 1993), including precipitates with phosphate (Firsching and Kell 1993). High pH and elevated alkalinity as in Jacarepaguá lagoon imply a higher proportion of hydroxyl- and carbonate ions in the water that could interfere with La-phosphate precipitation (Byrne and Kim 1993; Johannesson and Lyons 1994). Nonetheless, our results show that such interference in the water of Jacarepaguá is insufficient to render LMB ineffective and that sufficient La remained to effectively reduce the SRP efflux from the brackish sediment of Jacarepaguá lagoon. Effective blocking of sediment P release has also been found in a short-term (4 days) experiment, where 0.1 g of LMB and 1.0 g of bottom sediment from Swan River were incubated with 30-mL autoclaved water of 0.5- and 30-ppt salinity (Haghseresht 2006). It would be advisable to conduct additional research with LMB under more saline conditions to test the claim that “soluble La species” are released into the saline water and to evaluate the efficacy of LMB in more saline conditions.
The main reason for including PAC is the year round presence of a relatively high biomass of phytoplankton in Jacarepaguá lagoon (Gomes et al. 2009) and the incapacity of solely ballast to precipitate cyanobacteria, while their combination is highly effective (Noyma et al. 2017). The effectiveness of ballast compounds and a low-dose coagulant is, however, inversely related to cyanobacterial biomass—more biomass requires more ballast to effective settling (Noyma et al. 2017). Our results showed that 2-mg Al L−1 PAC in itself was insufficient to settle the cyanobacteria, but with a dose of 400 mg LMB L−1, effective removal could be achieved. It should be noted, however, that such LMB dose comes down to 4884 tons for the entire lagoon. Increasing the PAC dose to 8 mg L−1, which proved to be the best concentration in our previous studies with red soil as ballast (De-Magalhães et al. 2017), showed that comparable results could be achieved with 100 mg LMB L−1, which is 1221 tons for the entire lagoon.
Of course, the main reason for applying LMB is not the needed ballast weight, but the effectiveness in reducing the sediment SRP efflux. In the first trials, we used 400 g m−2, but this was increased to 507.5 g m−2 in the last sediment core experiment as we increased the communicating sediment depth to 10 cm and included the water column P in the calculations. This dose is in close vicinity with the average dose of 388 g LMB m−2 (range 159–530) given in Copetti et al. (2016) and the 348 g LMB m−2 (range 6–667) listed in Spears et al. (2013). Considering the total sediment area of the lagoon (3.7 km2), a 507.5-g m−2 dose implies 1878 tons of LMB would be needed, which with an average water depth of 3.3 m yields a dose of 154 mg LMB L−1 in the lagoon. Common application of LMB is as a slurry from the water surface—LMB granules are mixed with surface water just before spraying on the water (Copetti et al. 2016). Since this LMB dose is between suboptimal and optimal when combined with 2 mg Al L−1 of PAC (see Fig. 2), a dose of 8 mg Al L−1 of PAC seems better suited (see Fig. 3). Although 8 mg Al L−1 of PAC by itself was already sufficient to precipitate the cyanobacteria, the short-term sediment core experiment clearly evidenced that it was ineffective in hampering the sediment P efflux (see Fig. 7b). Using a higher dose of PAC to counteract the sediment, P release is not recommended for several reasons despite the cost of PAC is only about 15–20% that of LMB. First, the minimum dose needed, based on an Al:P ratio of minimally 10:1 (cf. De Vicente et al. 2008; Egemose et al. 2010), would boil down to at least 16 mg Al L−1. At such PAC dose, the pH in Jacarepaguá water will drop and depending on the pH at application could drop to pH values below 6 (De-Magalhães et al. 2017). Second, the Al polymerization seems to be the most important factor for acute hypoxic death in fish (Poléo 1995), and thus, negative effects of a relatively high Al dose are likely on the abundant fish, such as Tilapia rendalli, which is an important feed and source of income to fishermen living adjacent to the lagoon. Finally, Al-flocks are easily resuspended and hence distributed, while LMB is not (Egemose et al. 2010). In fact, LMB strongly increased sediment stability/consolidation and the resuspension data obtained by Egemose et al. (2010) drove them to conclude that wind driven events will most probably not cause any resuspension of LMB in contrast to Al flocks. Nonetheless, the influence of wind driven resuspension on flocks and P-binding capacity in a large shallow system like Jacarepaguá lagoon needs to be determined.
The combination of a higher dose of PAC (8 mg L−1) with dose of LMB targeting both water column and sediment P, not only ensures a stripping of the water column of cyanobacteria and phosphate, which is around 0.8 mg SRP L−1 despite cyanobacteria flourishing (De-Magalhães et al. 2017), but also sufficient SRP binding capacity to counteract any P that might diffuse from a bit deeper than 10-cm sediment as the PAC brings an additional SRP binding capacity equal to about 5 cm of sediment.
The negative P flux and the low SRP concentration in the long-term experiment (− 4.6 ± 0.3 mg P m−2 day−1) showed the strong P binding capacity of LMB + PAC. This P binding occurred in cores that had very low oxygen concentrations, which is common for Jacarepaguá lagoon and is in line with other studies that also found good P adsorption by LMB under anoxia (Robb et al. 2003; Akhurst et al. 2004; Ross et al. 2008). The combination of a low-dose PAC with a sediment P target dose of LMB has been applied in a whole lake application in 2008 (Lürling and Van Oosterhout 2013). The hypertrophic water with dominance of cyanobacteria in Lake Rauwbraken (The Netherlands) was changed to a mesotrophic clear water state with total P concentrations for more than 4 years being less than 10% pre-application concentrations (Lürling and Van Oosterhout 2013). It should be noted, however, that Lake Rauwbraken, unlike Jacarepaguá lagoon, has no major inflows of nutrient rich water, but rather diffuse sources via ground water, litter fall, and birds (Lürling and Van Oosterhout 2013). In Jacarepaguá, the external load is overwhelming with an average discharge of 0.8 m3 s−1 (Gomes et al. 2009) from the tributaries and an average total P concentration of 1477 mg m−3; the external load comes to 27.6 mg P m−2 day−1 (~ 10 g P m−2 year−1). Given such ongoing external P loading in Jacarepaguá lagoon, it is beyond doubt that first these external sources should be tackled before massive in-lake rehabilitation actions are undertaken.
External load reductions could reduce the eutrophication symptoms within several years depending on efficacy of P load reduction and retention time (Fastner et al. 2016). Although the retention time in Jacarepaguá is around 0.5 years, suboptimal mixing and particular the high internal P load could hamper delay in recovery for many years (Fastner et al. 2016). In Jacarepaguá lagoon, the internal P flux of ~ 10 mg P m−2 day−1 (or ~ 3.6 g P m−2 year−1) is ~ 14 times higher than the critical load, here calculated. Although the model has not been developed for brackish water and estimated critical loadings come with some uncertainties (Janse et al. 2010), the internal P loading was substantially larger than the critical loading. Reducing the P loading to below the critical P load is the only option for rehabilitation (Vollenweider 1976; Janse et al. 2008). Thus, even when external loadings are completely stopped, the water quality in Jacarepaguá lagoon will not improve rapidly unless internal loading is tackled adequately. Consequently, additional in-lake actions seem inevitable in this system. Treatments with only LMB or LMB + PAC could bring the internal P load below the critical load, but as emphasized the external load should also be strongly controlled by implementing efficient waste water treatment in the catchment. Furthermore, those catchment treatments should keep the P load below the critical load to ensure enduring improved water quality.
The effective control of the sediment P release from Jacarepaguá sediment becomes important in view of the planned but not yet executed dredging plans for the lagoon. According to media reports, the dredging project will cost $250 million (http://www.dailymail.co.uk/wires/ap/article-2947878/Rio-official-visits-filthy-lagoon-near-Olympic-Park.html). Of course, as mentioned before, such actions should only be undertaken once external loading has been reduced drastically, but then LMB or LMB + PAC might prove a much cheaper option or could be considered as addition to dredging. The effective dose used here (507.5 g m−2) will boil down to around 1878 tons of LMB for the entire lagoon. With a pricing between $2500 and $3000 per ton of LMB, the total material costs to tackle the internal load would be between $4.7 and $5.6 million. Assuming that also the entire water column needs to be stripped of 1 mg P L−1, which requires an additional 1000 tons ($2.5–3.0 million) and including operational costs a total budget of around $10 million would suffice. This is only 4–5% of the estimated dredging costs. Moreover, it remains to be seen if the planned dredging and storage of sediment in geotextiles in a newly to create island in the lagoon will sufficiently reduce in-lake nutrients to improve water quality. The in-lagoon P concentrations need to be pushed below the threshold concentration needed to minimize the risk on cyanobacterial blooms (Fastner et al. 2016). The here tested combination of PAC and LMB proved an efficient method to settle cyanobacteria out of the water column and to block the sediment P release. Hence, the combination seems promising to test at a larger scale in-situ using enclosures.
Conclusions
-
Positively buoyant cyanobacteria could be precipitated using low dosage PAC (2 mg Al L−1) combined with higher LMB dose and solely with a higher dose of PAC (8 mg Al L−1) or also combined with lower LMB dose.
-
The determined internal P loading from the sediment exceeded estimated critical P loading for rehabilitation meaning that only external load reduction will not improve water quality in the lagoon and that both internal and external P load should be tackled.
-
The PAC dose used (8 mg Al L−1) was not capable to block P release from the sediment, but the LMB proved highly efficient in a brackish system.
-
In all treatments with LMB and LMB + PAC, negative SRP fluxes were determined meaning a net removal of P from the water column towards the sediment.
-
In a 3-month sediment core experiment, combined LMB + PAC treatment kept SRP as low as 2% of the controls underpinning the strong and robust interception of P released from the heavily P enriched sediment of Jacarepaguá lagoon.
References
Akhurst, D., G.B. Jones, and D.M. McConchie. 2004. The application of sediment capping agents on phosphorus speciation and mobility in a sub-tropical dunal lake. Marine and Freshwater Research 55 (7): 715–725.
Barbosa, M.C., and M.S.S. Almeida. 2001. Dredging and disposal of fine sediments in the state of Rio de Janeiro, Brazil. Journal of Hazardous Materials 85 (1-2): 15–38.
Byrne, R.H., and I.H. Kim. 1993. Rare earth precipitation and coprecipitation behavior: The limiting role of PO4 3− on dissolved rare earth concentrations in seawater. Geochimica et Cosmochimica Acta 57 (3): 519–526.
Cavalcante, H., F. Araújo, N.P. Noyma, and V. Becker. 2018. Phosphorus fractionation in sediments of tropical semiarid reservoirs. Science of the Total Environment. 619-620: 1022–1029. https://doi.org/10.1016/j.scitotenv.2017.11.204.
Conley, D.J., H.W. Paerl, R.W. Howarth, D.F. Boesch, S.P. Seitzinger, K.E. Havens, C. Lancelot, and G.E. Likens. 2009. Controlling eutrophication: Nitrogen and phosphorus. Science 323 (5917): 1014–1015.
Cooke, G.D., E.B. Welch, S. Peterson, and S.A. Nichols. 2005. Restoration and management of lakes and reservoirs. Boca Raton: CRC press.
Copetti, D., K. Finsterle, L. Marziali, F. Stefani, G. Tartari, G. Douglas, K. Reitzel, B.M. Spears, I.J. Winfield, G. Crosa, P. D'Haese, and M. Lürling. 2016. Eutrophication management in surface waters using a lanthanum-modified bentonite: A review. Water Research 97: 162–174.
Correl, D.L. 1998. The role of phosphorus in the eutrophication of receiving waters: A review. Journal of Environmental Quality 27 (2): 261–266.
De Vicente, I., P. Huang, F.Ø. Andersen, and H.S. Jensen. 2008. Phosphate adsorption by fresh and aged aluminum hydroxide. Consequences for lake restoration. Environmental Science and Technology 42 (17): 6650–6655.
De-Magalhães, L., N. Noyma, L.L. Furtado, M. Mucci, F. van Oosterhout, V.L.M. Huszar, M.M. Marinho, and M. Lürling. 2017. Efficacy of coagulants and ballast compounds in removal of cyanobacteria (Microcystis) from water of the tropical lagoon Jacarepaguá (Rio de Janeiro, Brazil). Estuaries and Coasts. 40 (1): 121–133. https://doi.org/10.1007/s12237-016-0125-x.
Dithmer, L., U.G. Nielsen, M. Lürling, B.M. Spears, S. Yasseri, D. Lundberg Moore, N.D. Jensen, and K. Reitzel. 2016. Responses in sediment phosphorus and lanthanum concentrations and composition across 10 lakes following applications of lanthanum modified bentonite. Water Research 97: 101–110. https://doi.org/10.1016/j.watres,2106.02.011.
Douglas, G. B. 2002. Remediation material and remediation process for sediment. USA Patent 6350383.
Douglas, G. B., J. A. Adeney, and M. S. Robb. 1999. A novel technique for reducing bioavailable phosphorus in water and sediments. International Association Water Quality Conference on Diffuse Pollution: 517–523.
Egemose, S., K. Reitzel, F.Ø. Andersen, and M.R. Flindt. 2010. Chemical lake restoration products: Sediment stability and phosphorus dynamics. Environmental Science and Technology 44 (3): 985–991.
Epe, T.S., K. Finsterle, and Y. Nine. 2017. Years of phosphorus management with lanthanum modified bentonite (Phoslock) in a eutrophic, shallow swimming lake in Germany. Harmful Algae 61: 32–45.
Esteves, F.A., A. Caliman, J.M. Santangelo, R.D. Guariento, V.F. Farjalla, and R.L. Bozelli. 2008. Neotropical coastal lagoons: An appraisal of their biodiversity, functioning, threats and conservation management. Brazilian Journal of Biology 68 ((4) supp l.0): 967–981. https://doi.org/10.1590/S1519-69842008000500006.
Fastner, J., S. Abella, A. Litt, G. Morabito, L. Vörös, K. Pálffy, D. Straile, R. Kümmerlin, D. Matthews, G. Phillips, and I. Chorus. 2016. Combating cyanobacterial proliferation by avoiding or treating inflows with high P load—Experiences from eight case studies. Aquatic Ecology 50 (3): 367–383.
Ferrão-Filho, A.S., P. Domingos, and S.M.F.O. Azevedo. 2002. Population dynamics during a Microcystis aeruginosa bloom in Jacarepaguá lagoon (RJ, Brazil). Limnologica 32 (4): 295–308.
Firsching, F.H., and J.C. Kell. 1993. The solubility of the rare-earth-metal phosphates in sea water. Journal of chemical and Engeneering Data 38 (1): 132–133.
Gebbie, P. 2001. Using polyaluminium coagulants in water treatment. 64th Annual Water Industry Engineers and Operators Conference: 39–47.
Golterman, H.L. 1975. Physiological limnlogy: an approach to the physiology of lake ecosystems. Amsterdam: Elsevier.
Gomes, A.M.A., P.L. Sampaio, A.S. Ferrão-Filho, V.F. Magalhães, M.M. marinho, A.C.P. Oliveira, V.B. Santos, P. Domingos, and S.M.F.O. Azevedo. 2009. Toxic cyanobacterial blooms in an eutrophicated coastal lagoon in Rio de Janeiro, Brazil: Effects on human health. Oecologia brasiliensis 13 (2): 329–345.
Gulati, R.D., and E. Van Donk. 2002. Lakes in the Netherlands, their origin, eutrophication and restoration: State-of-the-art review. Hydrobiologia 478 (1/3): 73–106.
Haghseresht, F. 2006. A revolution in phosphorus removal. PS-06, Phoslock Water Solutions Limited, Rosebery, Australia.
Huszar, V.L.M., L.H.S. Silva, M.M. Marinho, P. Domingos, and C.L. Sant’anna. 2000. Cyanoprokaryote assemblages in eight productive tropical Brazilian waters. Hydrobiologia 424 (1): 67–77.
Janse, J.H., L.N. De Senerpont Domis, M. Scheffer, L. Lijklema, L. Van Liere, M. Klinge, and W.M. Mooij. 2008. Critical phosphorus loading of different types of shallow lakes and the consequences for management estimated with the ecosystem model PCLake. Limnologica 38 (3-4): 203–219.
Janse, J.H., M. Scheffer, L. Lijklema, L. Van Liere, J.S. Sloot, and W.M. Mooij. 2010. Estimating the critical phosphorus loading of shallow lakes with the ecosystem model PCLake: Sensitivity, calibration and uncertainty. Ecological Modelling 221 (4): 654–665.
Jeppesen, E., P. Kristensen, J.P. Jensen, M. Søndergaard, E. Mortensen, and T. Lauridsen. 1991. Recovery resilience following a reduction in external phosphorus loading of shallow, eutrophic Danish lakes: Duration, regulating factors and methods for overcoming resilience. Memorie dell, stituto Italiano di Idrobiologia 48: 127–148.
Johannesson, K.H., and B.W. Lyons. 1994. The rare earth element geochemistry of Mono Lake water and the importance of carbonate complexing. Limnology and Oceanography 39n (5): 1141–1154.
Kennish, M.J. 2002. Environmental threats and environmental future of estuaries. Environmental Conservation 29 (1): 78–107.
Kennish, M.J., M.J. Brush, and K.A. Moore. 2014. Drivers of change in shallow coastal photic systems: An introduction to a special issue. Estuaries and Coasts 37 (Suppl 1): 3–19.
Lürling, M., and F. van Oosterhout. 2013. Controlling eutrophication by combined bloom precipitation and sediment phosphorus inactivation. Water Research 47 (17): 6527–6537.
Lürling, M., N. Noyma, L. de Magalhães, M. Miranda, M. Mucci, F. van Oosterhout, V.L. Huszar, and M.M. Marinho. 2017. Critical assessment of chitosan as coagulant to remove cyanobacteria. Harmful Algae 66: 1–12.
Markou, D.A., G.K. Sylaios, V.A. Tsihrintzis, G.D. Gikas, and K. Haralambidou. 2007. Water quality of Vistonis Lagoon, Northern Greece: Seasonal variation and impact of bottom sediments. Desalination 210 (1-3): 83–97.
Miranda, M., N. Noyma, F.S. Pacheco, L. de Magalhães, E. Pinto, S. Santos, M.F.A. Soares, V.L. Huszar, M. Lürling, and M.M. Marinho. 2017. The efficiency of combined coagulant and ballast to remove harmful cyanobacterial blooms in a tropical shallow system. Harmful Algae 65: 27–39. https://doi.org/10.1016/j.hal.2017.04.007.
Mooij, W.M., D. Trolle, E. Jeppesen, G. Arhonditsis, P.V. Belolipetsky, D.B.R. Chitamwebwa, A.G. Degermendzhy, D.L. DeAngelis, L.N. De Senerpont Domis, A.S. Downing, J.A. Elliott, C.R. Fragoso, U. Gaedke, S.N. Genova, R.D. Gulati, L. Hákanson, D.P. Hamilton, M.R. Hipsey, J. t’Hoen, S. Hulsmann, F.H. Los, V. Makler-Pick, T. Petzoldt, I.G. Prokopkin, K. Rinke, S.A. Schep, K. Tominaga, A.A. Van Dam, E.H. Van Nes, S.A. Wells, and J.H. Janse. 2010. Challenges and opportunities for integrating lake ecosystem modelling approaches. Aquatic Ecology 44 (3): 633–667. https://doi.org/10.1007/s10452-010-9339-3.
Noyma, N., L. de Magalhães, L. Lima Furtado, M. Mucci, F. van Oosterhout, V.L.M. Huszar, M.M. Marinho, and M. Lürling. 2016. Controlling cyanobacterial blooms through effective flocculation and sedimentation with combined use of flocculents and phosphorus adsorbing natural soil and modified clay. Water Research 97: 26–38. https://doi.org/10.1016/j.watres.2015.11.057.
Noyma, N.P., L. de Magalhães, M. Miranda, M. Mucci, F. van Oosterhout, V.L.M. Huszar, M.M. Marinho, E.R.A. Lima, and M. Lürling. 2017. Coagulant plus ballast technique provides a rapid mitigation of cyanobacterial nuisance. PLoS One 12 (6): e0178976. https://doi.org/10.1371/journal.pone.0178976.
Paerl, H.W., and J. Huisman. 2008. Blooms like it hot. Science 320 (5872): 57–58.
Paerl, H.W., and V.J. Paul. 2012. Climate change: Links to global expansion of harmful cyanobacteria. Water Research 46 (5): 1349–1363.
Paerl, H.W., N.S. Hall, B.L. Peierls, and K.L. Rossignol. 2014. Evolving paradigms and challenges in estuarine and coastal eutrophication dynamics in a culturally and climatically stressed world. Estuaries and Coasts 37 (2): 243–258.
Paludan, C., and H.S. Jensen. 1995. Sequential extraction of phosphorus in freshwater wetland and lake sediment: Significance of humic acids. Wetlands 15 (4): 365–373.
PBL 2015. Metamodel PCLake. Planbureau voor de Leefomgeving. http://themasites.pbl.nl/modellen/pclake/index.php. Accessed 20 Apr 2017.
Perelo, L.W. 2010. Review: In situ and bioremediation of organic pollutants in aquatic sediments. Journal of Hazardous Materials 177 (1–3): 81–89.
Poléo, A.B.S. 1995. Aluminium polymerization—a mechanism of acute toxicity of aqueous aluminium to fish. Aquatic Toxicology 31 (4): 347–356.
Reitzel, K., S. Lotter, M. Dubke, S. Egemose, H.S. Jensen, and F.Ø. Andersen. 2013. Effects of Phoslock_ treatment and chironomids on the exchange of nutrients between sediment and water. Hydrobiologia 703 (1): 189–202.
Robb, M., B. Greenop, Z. Goss, G. Douglas, and J. Adeney. 2003. Application of Phoslock_, an innovative phosphorus binding clay, to two Western Australian waterways: Preliminary findings. Hydrobiologia 494 (1-3): 237–243.
Ross, G., F. Haghseresht, and T.E. Cloete. 2008. The effect of pH and anoxia on the performance of Phoslock®, a phosphorus binding clay. Harmful Algae 7 (4): 545–550.
Søndergaard, M., J.P. Jensen, and E. Jeppesen. 2001. Retention and internal loading of phosphorus in shallow, eutrophic lakes. Scientific World Journal 1: 427–442.
Søndergaard, M., J.P. Jensen, and E. Jeppesen. 1999. Internal phosphorus loading in shallow Danish lakes. Hydrobiologia 408: 145–152.
Spears, B.M., S. Meis, A. Anderson, and M. Kellou. 2013. Comparison of phosphorus (P) removal properties of materials proposed for the control of sediment p release in UK lakes. Science of the Total Environment 442: 103–110.
Spears, B.M., E.B. Mackay, S. Yasseri, I.D.M. Gunn, K.E. Waters, C. Andrews, S. Cole, M. De Ville, A. Kelly, S. Meis, A.L. Moore, G.K. Nürnberg, F. van Oosterhout, J. Pitt, G. Madgwick, H.J. Woods, and M. Lürling. 2016. A meta-analysis of water quality and aquatic macrophyte responses in 18 lakes treated with lanthanum modified bentonite (Phoslock®). Water Research 97: 111–121.
Vollenweider, R.A. 1976. Advances in defining critical loading levels for phosphorus in lake eutrophication. Memorie dell, stituto Italiano di Idrobiologia 33: 53–83.
Waajen, G., F. Van Oosterhout, G. Douglas, and M. Lürling. 2016a. Geo-engineering experiments in two urban ponds to control eutrophication. Water Research 97: 69–82. https://doi.org/10.1016/j.watres.2015.11.070.
Waajen, G., F. Van Oosterhout, G. Douglas, and M. Lürling. 2016b. Management of eutrophication in Lake De Kuil (The Netherlands) using combined flocculant—Lanthanum modified bentonite treatment. Water Research 97: 83–95. https://doi.org/10.1016/j.watres.2015.11.034.
Zamparas, M., A. Gianni, P. Stathi, Y. Deligiannakis, and I. Zacharias. 2012. Removal of phosphate from natural waters using innovative modified bentonites. Applied Clay Science 62: 101–106.
Funding
This study was sponsored by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Brasil, through a Sciencewithout Borders Grant, SwB (400408/2014-7) and by Fundação de Apoio à Pesquisa do Estado do Rio de Janeiro, FAPERJ, Brasil(111.267/2014). L. De Magalhães PhD scholarship was funded by Federal Government of Brazil, Ministry of Education, through CAPES(Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Ministério da Educação). V. Huszar was partially supported by CNPq(309700/2013-2). M. Mucci PhD scholarship was funded by SwB/CNPq (201328/2014-3). N. Noyma Postdoctoral fellowship was fundedby SwB/CNPq (159537/2014-2). This study was conducted under the flag of the CAPES (Brazil)/NUFFIC (The Netherlands) project 045/12.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Margaret R. Mulholland
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
de Magalhães, L., Noyma, N.P., Furtado, L.L. et al. Managing Eutrophication in a Tropical Brackish Water Lagoon: Testing Lanthanum-Modified Clay and Coagulant for Internal Load Reduction and Cyanobacteria Bloom Removal. Estuaries and Coasts 42, 390–402 (2019). https://doi.org/10.1007/s12237-018-0474-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-018-0474-8