
Abstract
Invasive clonal species may exhibit different growth strategies than their native clonal competitors. In this study, we examined the spatial distribution of tiller outgrowth and the bud bank by comparing the investment in phalanx versus guerilla growth of a native and invasive perennial grass in North America. We also examined the effect of altered precipitation frequency, clipping, and competition on their clonal growth strategies. Investment in phalanx and guerilla growth was assessed by examining live propagule and tiller production from the plant crown versus its rhizomes. Although invasive Bromus inermis and native Pascopyrum smithii exhibited similar clonal growth strategies as young seedlings, their clonal growth strategies significantly differed by the end of their first growing season. Pascopyrum smithii invested in dual phalanx and guerilla tiller outgrowth and bud placement, and B. inermis primarily invested in phalanx tiller outgrowth and bud placement. Competition rather than intra-annual precipitation variability and clipping altered the clonal growth strategy of these species. Intra- and inter- specific competition did not alter tiller outgrowth for either species. However, inter-specific competition caused both species to alter their bud placement. Bromus inermis shifted more buds from phalanx to guerilla positions while P. smithii shifted in the opposite direction. This may enable invasive B. inermis to expand while confining native P. smithii to more localized areas in the future. Clonal growth strategies appear to be species specific and responsive to inter-specific competition. Investigating the belowground bud aspect of clonal growth can reveal the mechanism driving the future aboveground clonal growth strategy of native and invasive rhizomatous grasses and help inform the patterns of invasion within a plant community.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Clonal plant growth and expansion, especially rhizomatous species, can be described along a “phalanx-guerilla” clonal growth form continuum (Lovett-Doust 1981a). At the two extremes, new ramets can be considered phalanx or guerrilla ramets (Lovett-Doust 1981a; Oborny and Cain 1997). Clonal plants with the phalanx growth strategy produce a compact structure of closely spaced ramets that slowly advance out from the parent plant, termed clumping ramets or tillers, whereas those with a guerilla growth strategy produce a loosely arranged group of widely spaced ramets spread far from the parent plant, termed as spreading ramets or tillers (Lovett-Doust 1981b; Bernard 1990). Guerrilla growth forms are commonly observed in early successional stages (Saiz et al. 2016), as well as in resource-heterogeneous, resource-poor, and disturbed habitats where new ramets are targeted to search out limited resources and favorable microsites (Lovett-Doust 1981b; Sutherland and Stillman 1988; Humphrey and Pyke 1998; Yu et al. 2020). Phalanx growth is more favored in late successional stages and in relatively homogeneous, resource-rich and less disturbed habitats (Schmid and Harper 1985; Yu et al. 2020). The phalanx growth strategy may enable clonal plants to tolerate more stressful conditions, make better use of locally abundant resources (monopolization strategy) and out-compete other species in a favorable microsite (Doust 1981b; Schmid and Harper 1985; Humphrey and Pyke 1998). Some species, such as Pascopyrum smithii, invest in both phalanx and guerilla tiller production in their native habitat (Ott and Hartnett 2015).
Perennial grasses can demonstrate plasticity in their growth forms under different environmental conditions (Lovett-Doust 1981b; Garnier and Roy 1988; Hutchings and Slade 1988). Perennial plants Ranunculus repens and Glechoma hederacea showed a guerrilla growth strategy (extensive foraging) under low-light and low nutrient, and a phalanx growth strategy (intensive foraging) under high-light and high nutrient conditions (Lovett-Doust 1981b; Slade and Hutchings 1987; Hutchings and Slade 1988). When released from competition by removing neighboring plants, the perennial grass Panicum virgatum experienced a >95% increase in radial clone expansion, intraconal ramet densities, and ramet population growth rates (Hartnett 1993). In Leymus secalinus, plants are more phalanx-like under high nutrient supply and more guerilla-like in low nutrient conditions (Ye et al. 2006; Zuo et al. 2023). In tidal wetlands, Elymus repens transitioned from a guerilla growth strategy pattern to a phalanx growth pattern upon its release from grazing pressure and increased competition from neighboring plants (Amiaud et al. 2008).
In the semi-arid mixed-grass prairie in the northern Great Plains of North America, the dominant perennial grasses are clonal and their local ramet population regeneration is primarily via vegetative reproduction from belowground buds (Benson and Hartnett 2006; Ott et al. 2017). These grasslands are dependent on a complex disturbance regime, including frequent fire, grazing by livestock and native herbivores, and strong inter-annual climatic variability (Schrag 2011). Because of climate change, growing season precipitation regimes are predicted to become more variable, with larger events and longer dry periods resulting in more temporally dynamic soil moisture regimes (Koerner et al. 2014). Increased climate variability is likely to interact with other disturbances, such as grazing, which may profoundly impact grassland community structure and function by affecting competitive dynamics between native and non-native invasive species, and potentially undermining the effectiveness of restoration activities (Koerner et al. 2015; Finch et al. 2021).
The northern Great Plains face the additional threat of invasion by introduced C3 perennial grasses, such as Bromus inermis Leyss. (smooth brome) (Christian and Wilson 1999; Grant et al. 2020; Larson et al. 2001; DeKeyser et al. 2013). Large areas of native grasslands have transformed into B. inermis monocultures with reduced native plant diversity (Frank and McNaughton 1992; Dillemuth et al. 2009) and habitat use by native ungulates (Trammell and Butler 1995). Bromus inermis expansion and success largely depends on its vegetative proliferation via belowground buds and rhizomes and its resulting integration among ramets (Otfinowski et al. 2007; Otfinowski and Kenkel 2008). Clonal integration can favor the successful invasion of alien plants into native plant communities (Wang et al. 2022). The spatial distribution of belowground buds and their varying levels of dormancy can drive where future tiller recruitment occurs (Ott et al. 2019). Therefore, it would be beneficial to examine how this invasive species alters its growth strategy in comparison to the dominant C3 native species, which has a similar rhizomatous growth form, and their responses to shifts in competition, predicted climatic changes, and grazing.
Comparing the resource investment of native P. smithii, and non-native B. inermis to tiller production and propagule supply from the plant-crown (phalanx) versus rhizome (guerilla) may reveal how each species prioritizes current and future guerilla versus phalanx growth in response to a change in environmental conditions. Tiller recruitment and bud placement in the crown places current and future growth near parent tillers allowing continued occupancy with a slow movement away from the parent tiller while rhizomes place current and future growth away from the parent tiller allowing new resource pools to be accessed. Tiller, rhizome, and propagule production declined for B. inermis as precipitation became more intermittent, while tiller and propagule production remained steady across multiple levels of precipitation variability, and rhizome production was highest at medium levels of precipitation variability for P. smithii (Bam et al. 2022). A solitary clipping event had no effect on tiller, rhizome, and propagule production (Bam et al. 2022). Competition reduced tiller and rhizome production in both species but only reduced propagule production in P. smithii (Bam 2018). Precipitation variability, clipping, and competition could also alter the location of tillers and propagules within the plant showing how these factors could influence the growth strategies of these plants. Using data from these same two greenhouse studies that examined tiller, rhizome, and propagule production of both species, our main objectives were to compare clonal growth strategies between native P. smithii, and non-native B. inermis under 1) altered precipitation frequency and simulated grazing, and 2) competition. Because of its invasiveness, we hypothesize that non-native B. inermis will invest more into current and future guerilla growth strategies under all precipitation frequencies, clipping, and competition conditions than the native P. smithii. We also hypothesize that both species will demonstrate plasticity in their growth strategies when encountering different environmental conditions and reduce their guerilla growth, allocating more to phalanx growth when experiencing lower precipitation frequencies, clipping, and intra- and inter- specific competition.
Materials and Methods
Species and Seed Source
Pascopyrum smithii (western wheatgrass), and B. inermis (smooth bromegrass) are both strongly rhizomatous, perennial C3 grasses that begin flowering in late May (the PLANTS database, USDA-NRCS 2021). Pascopyrum smithii is native to North America and is most abundant in northern mixed grass prairie, which usually receives between 254 to 508 mm of annual precipitation. In contrast, B. inermis was introduced to North America from Eurasia in the late 1880s for forage activity and has made an extensive impact on the grasslands of North America (Otfinowski et al. 2007; Palit and DeKeyser 2022). Bromus inermis establishes by invading disturbed prairies and through repeated introductions for soil retention and livestock grazing (Cully et al. 2003; DeKeyser et al. 2013; Stotz et al. 2019; Grant et al. 2020). In its native range in China, B. inermis is known to occur at 400 mm of annual precipitation (Gong et al. 2019).
Bromus inermis and P. smithii seeds were respectively obtained from commercial seed suppliers (Dakota’s Best Seed Limited Liability Company, Platte, South Dakota, USA; and Golden Willow Seeds, Inc., Midland, South Dakota, USA). Before each experiment, seeds were germinated in a commercial potting mix soil (Miracle-Gro®) with a temperature regime of 16 °C night/ 22 °C day in a greenhouse. Because of their slower germination, P. smithii seeds were sown five days earlier than B. inermis to obtain the same growth stage at time of transplant.
Experiment 1: Climate Change and Grazing
Treatments
Predicted climate change effects on precipitation frequency and their resulting effect on grazed and ungrazed seedlings of P. smithii and B. inermis were examined in a greenhouse at the South Dakota State University Seed Testing Laboratory in 2016. Precipitation frequency treatments began 19 days after seedling transplant and were subsequently applied for 20 weeks. Pots were watered every 2, 8, or 16 d to simulate high, medium, and low precipitation frequencies, respectively. Each treatment developed predictable volumetric water content (VWC) patterns during the study (Bam et al. 2022). Total water received was the same across treatments and simulated average growing season precipitation of 51 mm mo-1 (based on 1981–2010 March–August precipitation of Rapid City Regional Airport, South Dakota; https://climate.sdstate.edu). Therefore, 2-, 8-, or 16-d treatments received 72 mL, 288 mL, and 576 mL of water during each watering event, respectively.
A one-time clipping treatment was randomly assigned to half of the pots for each precipitation frequency treatment and species. This treatment was applied when seedlings of each species reached the three-collar leaf-stage, according to Moore et al. (1991), which occurred 20 days after seedling transplant. Seedlings were clipped to a 4-cm stubble height to simulate early-season grazing by ungulates.
Experimental Design
One single-leaf seedling was transplanted into each pot (16.5 cm dia. X 16.5 cm depth) filled with 600 g of non-fertilizer potting-soil (PRO-MIX® BX) and watered. A total of 480 pots were established (240 per species). Pots were watered to container capacity (44 – 45% VWC; Decagon Devices, Soil Moisture Sensor: Model EC-5 calibrated to the potting soil), and seedlings acclimated to their pots for 18 days. One week after seedling transplant, each pot received 100 mL solution (0.343g L-1 of Miracle-Gro® NPK (15-30-15); 5.8% ammoniacal nitrogen, 9.2% urea nitrogen; 30% P2O5; and 15% K2O).
Each treatment combination (species x precipitation frequency x clipping) was applied to 40 pots. Due to limited space, pots were randomly divided between two adjacent rooms of the greenhouse so that each treatment combination (species x clipping x precipitation frequency) was evenly represented in each room (n = 20). After seedlings acclimated to their pots for 20 days, precipitation frequency and clipping treatments were applied, and plants were grown for 20 additional weeks. The photoperiod and temperature regime mimicked mixed-grass prairie field conditions during the growing season (June–October; Bam 2018). As the 20 weeks treatment period of the experiment began on July 1, supplemental light (six 400-watt high-pressure sodium lamps) extended the ambient daylength between 0.5 and 1.5 hours depending on the month (Bam 2018). Due to a greenhouse roof leak altering water availability in 42 pots in one room, each treatment combination in that room had between 10 and 20 replicates when these pots were removed from the experiment.
Experiment 2: Competition
Treatments
A temperature-controlled competition greenhouse experiment was conducted at South Dakota State University in the Forestry and Horticulture Greenhouse in 2017. Competitive effects of B. inermis and P. smithii were tested using three competition treatments, including no competition (single plant without neighbor), intraspecific competition (two conspecific plants), and interspecific competition (one plant of each species). Therefore, the experiment consisted of five treatment combinations (Figure 1).

Experimental design for the competition study. BI = Bromus inermis, PS = Pascopyrum smithii
Experimental Design
Two-leaf seedlings were transplanted into pots (16.5 cm dia. X 16.5 cm depth) filled with 600 g of non-fertilizer potting-soil (PRO-MIX® BX) according to their treatment combination. Each treatment combination was applied to 30 pots (total of 150 pots). Pots were watered and fertilized in the same manner as Experiment 1. After being transplanted into the pots, volumetric water content (VWC) was maintained at 44–45% for the first week to assist with seedling establishment (Decagon Devices; Soil Moisture Sensor: Model 10HS). Soil moistures levels then dropped down to 25–28% VWC two weeks post-transplant and were maintained throughout the experiment by watering each pot every other day with 72 mL of water. Plants were grown for 12 weeks (June–August) under the same photoperiod and temperature regime as Experiment 1.
Data Collection
Plants were destructively harvested and washed free of soil at the end of each study (20 weeks for Exp. 1, 12 weeks for Exp. 2). For each individual plant, all rhizomes and adult tillers were counted. Rhizomes were belowground stems at least 0.5 cm in length with elongated internodes. Buds transition to adult tillers when they have vertically elongated aboveground past their prophyll and had reached 3.6 cm and 4.5 cm in height for B. inermis and P. smithii, respectively (following Ott et al. 2017). Three generations of tillers occurred during the experiment (Bam et al. 2022). Therefore, tillers and rhizomes were classified by generation following Ott and Hartnett (2015).
A random sub-sample of 3 tillers from each generation of tillers (primary, secondary, and tertiary), and 3 rhizomes from each generation of rhizome per individual plant was selected to assess bud production and bud development stages. Each tiller/rhizome was examined using a dissecting scope (Olympus® Stereo Microscope) with magnification between 6.7x and 45x. Rhizomes, basal/belowground buds, and new tillers borne on photosynthesizing tillers were counted and assessed to be living or dead and classified by their size (Ott et al. 2017). Collectively, all the live buds, small juvenile tillers, and large juvenile tillers were called live propagules. For each plant, live propagules per plant was calculated by summing the number of propagules belonging to each generation of adult tillers or rhizomes. These quantities for each stem type within a plant were obtained using the propagule count per stem type (e.g., primary rhizome, secondary tiller, etc.) multiplied by the total stems of that type. As these species have long rhizomes, buds and tillers located on rhizomes were categorized as guerilla growth while those located on parent tillers (i.e., the crown of the plant) were considered phalanx growth (Ott and Hartnett 2015). For a conceptual diagram, see Figure 6 in Bam et al. 2022 which shows three first generation tillers (T1) where two are close to the parent tiller (phalanx tillers) and one is placed far away from the parent tiller (guerilla). For each plant, the proportion of propagules that occur in guerilla versus phalanx positions were calculated as the number of propagules in the location divided by the total propagules. For each plant, total tillers was the sum of all adult tillers growing from the crown, rhizome node, and rhizome tip. The proportion of tillers from each of these types was calculated. Current guerilla growth would include tiller outgrowth from rhizome nodes and rhizome tips while current phalanx growth is from crown tillers. For each plant, tillers are considered evidence of current guerilla or phalanx growth, while propagules represent future guerilla or phalanx growth.
Statistical Analysis
Linear mixed models were used to analyze the effect of precipitation frequency, clipping, competition, and species on the proportion of total live propagules and the proportion of total tillers (PROC GLIMMIX; SAS Institute 2017). All treatments were applied to the experimental unit of plant, except location, which was applied to the experimental unit of tiller. Kenward-Roger’s (KR) method was used to approximate the denominator degrees of freedom.
Experiment 1: Climate Change and Grazing
The proportion of total live propagules from all tillers per plant was analyzed using a beta distribution in a three-way factorial treatment structure with the fixed factors of precipitation frequency (three levels), clipping (two levels), and species (two levels) in a randomized complete block design, with the greenhouse chamber as the block effect. The proportion of total tillers per plant belonging to each location (crown, rhizome nodes, and rhizome tips) was analyzed using a negative binomial distribution in a four-way factorial treatment structure with the fixed factors of precipitation frequency (three levels), clipping (two levels), species (two levels), and location (three levels) in a split-plot block design, with the greenhouse chamber as the block effect.
Experiment 2: Competition
The proportion of total live propagule from tiller per plant analyzed using a beta distribution in a two-way factorial treatment structure with the factor of competition (three levels), and species (two levels) in a randomized complete design. The proportion of total tillers per plant belonging to each location (crown, rhizome nodes, and rhizome tips) was analyzed using a negative binomial distribution in a three-way factorial treatment structure with the factors of competition (three levels), species (two levels), and location (three levels) in a split-plot design.
Results
Experiment 1: Climate Change and Grazing
Current Growth Strategy
Invasive B. inermis predominantly invested in phalanx growth, as 65 to 80% of tillers were borne from the crown and 20 to 31% were recruited from the rhizome tip, and less than 3% recruited from rhizome nodes. Precipitation frequency significantly affected the location of B. inermis tiller recruitment investment. At a medium precipitation frequency (8 d,) tiller recruitment from rhizomes (apical and axillary buds) increased by ~10% as compared to high and low precipitation frequency treatments (Figure 2A). In contrast, native P. smithii primarily invested in guerilla growth as 36% of tillers recruited from the crown, 27% of tillers recruited from rhizome nodes, and 37% recruited from rhizome tips (Figure 2A). This dual allocation of current tillers to phalanx and guerilla growth was consistent across the three precipitation frequencies (Figure 2A). Clipping significantly increased tiller recruitment from nodes of rhizomes by ~5% of P. smithii, but had no effect on B. inermis (Figure 2B).

Effect of (a) precipitation frequency and (b) clipping on proportion of tillers per plant of Bromus inermis and Pascopyrum smithii. 2d = watering every 2 days. 8d = watering every 8 days. 16d = watering every 16 days. C = clipping. NC = no clipping. G = Guerilla. P = Phalanx. Standard errors are not shown. Tillers recruited from crown contributed to phalanx growth and tillers recruited from nodes and tips of rhizomes contributed to guerilla growth. Values were mean proportion of tillers per plant. Full statistical results available in Appendix Table 1.
Future Growth Strategy Investment
As the location of propagules represent the investment in future plant growth strategies, invasive B. inermis predominantly invested in future phalanx growth as 68 to 83% live propagules were located on parent tillers in the crown and 17% to 32% live propagules were produced from nodes and tips of rhizomes (Figure 3A). This is consistent with its current growth strategy (i.e., location of tillers). Precipitation frequency significantly affected the investment of B. inermis on live propagule placement. At a medium precipitation frequency (8 d), B. inermis invested in more future phalanx growth with more propagules from the crown than at the high precipitation frequency (Figure 3A). Pascopyrum smithii maintained dual phalanx and guerilla growth as ~50% of live propagules were produced from tillers and ~50% from rhizomes. This investment did not shift with alteration in precipitation frequency (Figure 3A). Clipping did not alter either species’ investment in phalanx and guerilla growth (Figure 3B).

Effect of (a) precipitation frequency and (b) clipping on proportion of live propagules per plant of Bromus inermis and Pascopyrum smithii. 2d = watering every 2 days. 8d = watering every 8 days. 16d = watering every 16 days. C = clipping. NC = no clipping. Values are mean proportion of tillers per plant ± SE based upon the statistical model. Significant differences at p-value < 0.05 indicated by different letters. Interaction results are shown when p-value < 0.05. Full statistical results available in Appendix Table 2. The effect of clipping was similar for both species. Live propagules included live buds, small juvenile, large juvenile tillers, and excluded adult tillers. Tillers recruited from crown contributed to phalanx growth and tillers recruited from nodes and tips of rhizomes contributed to guerilla growth.
Experiment 2: Competition
Current Growth Strategy
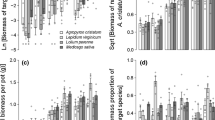
Competition did not significantly alter the tiller location, and therefore current growth strategy, of either species. Invasive B. inermis investment in dual phalanx and guerilla growth was consistent across types of competition with intraspecific competition producing 48% of new tillers from nodes and tips of rhizomes, and interspecific competition producing 38% of new tillers from nodes and tips of rhizomes (Figure 4). Native P. smithii prioritized phalanx growth (~73%) across all competition treatments (Figure 4). Bromus inermis had less, and P. smithii had more, guerilla growth in experiment 1 than experiment 2 when comparing similar treatments across experiments (2 d, no clip and no competition). Experiment 1 was 8 weeks longer than experiment 2. Therefore, B. inermis may allocate resources to guerilla growth and P. smithii may allocate resources to phalanx growth earlier in the growing season that may shift as the plants continue to develop.

Effect of competition treatment on proportion of total tillers based on location per plant of Bromus inermis and Pascopyrum smithii. None = no competition. Intra = Intra-specific competition. Inter = Inter-specific competition. G = Guerilla. P = Phalanx. Standard error is not shown. Tillers recruited from crown contributed to phalanx growth and tillers recruited from nodes and tips of rhizomes contributed to guerilla growth. Values were mean proportion of tillers per plant. Full statistical results available in Appendix Table 3.
Future Growth Strategy Investment
For invasive B. inermis, the intraspecific competition did not affect its live propagule growth investment strategy compared to no competition by maintaining a consistent dominant phalanx growth pattern with 80% of live propagules produced from tillers and 20% from rhizomes. However, interspecific competition shifted B. inermis from a phalanx growth strategy to dual phalanx and guerilla co-dominated growth strategy with 51% of live propagule produced from tillers and 49% from rhizomes (Figure 5). By contrast, native P. smithii responded to interspecific competition in an opposite manner than B. inermis. Pascopyrum smithii shifted from dual phalanx and guerilla with 53% of live propagules produced from tillers and 47% from rhizomes to a morephalanx strategy with 78% of live propagules produced from tillers and 22% from rhizomes when changing from intraspecific competition to interspecific competition (Figure 5). Furthermore, the intraspecific competition of P. smithii significantly increased investment in guerilla live propagules growth, whereas the interspecific competition decreased investment in guerilla live propagule growth when compared to no competition (Figure 5). When comparing comparable treatments between experiment 1 and 2, both species had lower guerilla investment in the shorter-term study indicating that investment in guerilla growth increases with plant development.

Effect of competition treatment on proportion of live propagules per plant of Bromus inermis and Pascopyrum smithii. None = no competition. Intra = Intra-specific competition. Inter = Inter-specific competition. Values are mean ± SE based on the statistical model. Significant differences at p-value < 0.05 indicated by different letters. Interaction results are shown when p-value < 0.05. Live propagules included live buds, small juvenile, large juvenile tillers, and excluded adult tillers. Tillers recruited from crown contributed to phalanx growth and tillers recruited from nodes and tips of rhizomes contributed to guerilla growth. Values were mean proportion of tillers per plant.
Discussion
Both the native and invasive species maintained phalanx and guerilla growth and placed buds in positions for future growth from phalanx and guerilla positions across precipitation variability, clipping, and competition treatments. Irrespective of change in precipitation frequency, clipping, and competition, P. smithii maintained a dual phalanx and guerilla growth form and had notable tiller recruitment from along its rhizomes as well as at its rhizome tips and tiller bases. Tiller outgrowth from along rhizomes enables space-filling between the most guerilla tillers (i.e., rhizome tips) and the phalanx tillers. In contrast, B. inermis primarily maintained a phalanx growth form but always had some guerilla tiller recruitment from rhizome tips, especially for the shorter-term experiment (i.e., competition experiment). Prioritization of the phalanx growth form by non-native B. inermis, irrespective of altered precipitation frequency, clipping, and competition conditions, may be affiliated with overall less rhizome production in B. inermis (Bam 2018; Bam et al. 2022). Non-native B. inermis tends to produce less rhizomes in first season’s growth regardless of the soil moisture variability (Dong et al. 2014). This predominantly phalanx growth form by B. inermis might have enabled it to establish from the seedling stage and make better use of locally abundant resources (Lovett-Doust 1981b; Schmid and Harper 1985; Humphrey and Pyke 1998).
Investment in a dual phalanx and guerilla clonal growth form by P. smithii may enable it to employ both conservative (phalanx) and foraging (guerilla) growth strategies which may facilitate its persistence under fluctuating resource availability associated with environmental change (Ott and Hartnett 2015). This also provides an opportunity to show architectural plasticity in P. smithii by the combination of both guerilla and phalanx traits in response to habitat and nutrient conditions (Lovett-Doust 1981b; Ye et al. 2006; Chen et al. 2011). Pascopyrum smithii disproportionately recruited more tillers from both rhizome tips and crown buds than from buds along the rhizome (Ott and Hartnett 2015). The higher density of rhizomes, greater rhizome length, greater bud supply and outgrowth also from rhizomes (guerilla growth form) in native P. smithii in comparison to non-native B. inermis as well as greater requirements to break bud dormancy as seen in wheatgrass species such as P. smithii and Pseudoroegneria spicata may indicate a disturbance-avoidance mechanism (Caldwell et al. 1981; Bam 2018; Bam et al. 2022). By keeping a large reserve of buds in between its growth front and parent plant, P. smithii may regrow in interstitial areas if guerilla tillers are separated by disturbance from their parent plant.
Examination of the placement of live propagules provides an indicator of where future growth will be prioritized. Clipping and precipitation variability had little impact on the placement of live propagules in guerilla or phalanx positions. Both of these disturbances are likely causing the plant to momentarily pause its growth during unfavorable periods but resume it when favorable conditions return and thus not putting undue pressure to shift the future growth strategy. Inter-specific competition caused the non-native B. inermis to shift from primarily prioritizing phalanx growth to a combination of the phalanx and guerilla growth. However, native P. smithii shifted its investment in future growth in the opposite direction under inter-specific competition and shifted from a combination of phalanx and guerrilla growth to more phalanx growth. This shift in bud placement may help us understand the mechanism behind the higher competitive ability of non-native B. inermis. Non-native B. inermis allocated extra effort to produce guerilla propagules when in competition with P. smithii. At the same time, P. smithii reduced its investment in guerilla tillers. As the competition continues, this may enable B. inermis to expand while confining P. smithii to more localized areas in the future. The prairies of the North American Great Plains are dominated by clonal perennial grasses that vary in architecture along the "phalanx - guerilla" clonal growth form continuum (Lovett-Doust 1981b; Harper 1985). In a study of the clonal perennial grass Panicum virgatum, neighborhood competition greatly influenced clonal architecture and expansion rates, where removal of neighbors resulted in a >95% increase in radial clone expansion, intraconal tiller densities, and tiller population growth rates (Hartnett 1993). The phalanx growth form by native P. smithii under the influence of non-native B. inermis as neighbor may be the mechanism to tolerate competition and make better use of locally abundant resources (monopolization strategy) but may enable the invasive B. inermis to expand (Lovett-Doust 1981b; Humphrey and Pyke 1998; Schmid and Harper 1985). Tiller position and clipping can also influence tiller ontogeny (Martínková and Klimešová 2017). Therefore, shifts in the spatial allocation of buds and disturbances may also lead to shifts in flowering which could demonstrate that clonal growth strategy may influence both vegetative and generative processes.
Growth strategies may shift with plant age. The competition experiment was conducted for a 12-week period, while the precipitation variability and clipping experiments were conducted for 20 weeks. When comparing these two studies, B. inermis started off with more tiller recruitment from the rhizome tip that lessened with age whereas P. smithii apparently increased its investment in tiller recruitment from rhizome nodes and rhizome tips with age. These shifts could indicate the initial priority of the invasive species to expand its occupying space while the native species initially aims to establish locally before investing in expansion but would require further direct experimental testing for verification.
Conclusion
Clonal growth strategies significantly differed between species. Pascopyrum smithii invested in dual phalanx and guerilla growth, and B. inermis primarily in phalanx growth. Pascopyrum smithii showed a lowered apical dominance by rhizome tip apical meristems than B. inermis as more tillers were recruited from rhizome buds in P. smithii. This would enable tiller recruitment between guerilla and phalanx tillers that would not occur as readily by B. inermis. Precipitation variability and clipping only had minor effects on clonal growth strategies, whereas inter-specific competition shifted the future placement of buds for tiller outgrowth. Pascopyrum smithii shifted from a dual to a more phalanx growth strategy, while B. inermis transitioned from a more phalanx to a dual growth strategy under inter-specific competition. This shift in bud location allocation by the native and invasive species when they are in competition with one another may explain why B. inermis is able to successfully invade stands of P. smithii.
Data Availability
Data from this study is available from the authors upon request.
References
Amiaud B, Touzard B, Bonis A, Bouzillé JB (2008) After grazing exclusion, is there any modification of strategy for two guerrilla species: Elymus repens (L.) Gould and Agrostis stolonifera (L.)? Plant Ecol 197:107-117. https://doi.org/10.1007/s11258-007-9364-z
Bam S (2018) Impacts of altered precipitation frequency, clipping, and competition on perennial grasses mediated through belowground bud bank in northern mixed-grass prairie. Master’s Thesis, South Dakota State University, Brookings, South Dakota
Bam S, Ott JP, Butler JL, Xu L (2022) Belowground mechanism reveals climate change impacts on invasive clonal plant establishment. Scientific Reports 12: 2860 https://doi.org/10.1038/s41598-022-06918-w
Benson EJ, Hartnett DC (2006) The role of seed and vegetative reproduction in plant recruitment and demography in tallgrass prairie. Plant Ecol 187:163-177. https://doi.org/10.1007/s11258-005-0975-y
Bernard JM (1990) Life-history and vegetative reproduction in Carex. Can J Bot 68:1441-1448. https://doi.org/10.1139/b90-182
Caldwell MM, Richards J, Johnson D, Nowak R, Dzurec R (1981) Coping with herbivory: photosynthetic capacity and resource-allocation in 2 semiarid Agropyron bunchgrasses. Oecologia 50:14-24. https://doi.org/10.1007/BF00378790
Chen XS, Xie Y-H, Deng Z-M, Li F, Hou Z-Y (2011) A change from phalanx to guerrilla growth form is an effective strategy to acclimate to sedimentation in a wetland sedge species Carex brevicuspis (Cyperaceae). Flora 206:347-350. https://doi.org/10.1016/j.flora.2010.07006
Christian JM, Wilson SD (1999) Long-term ecosystem impacts of an introduced grass in the northern Great Plains. Ecol 80:2397-2407. https://doi.org/10.2307/176919
Cully AC, Cully JF, Hiebert RD (2003) Invasion of exotic plant species in tallgrass prairie fragments. Conserv. Biol 17:990-998. https://doi.org/10.1046/j.1523-1739.2003.02107.x
DeKeyser ES, Meehan M, Clambey G, Krabbenhoft K (2013) Cool season invasive grasses in northern Great Plains natural areas. Nat Areas J 3:81-90. https://doi.org/10.3375/043.033.0110
Dillemuth FP, Rietschier EA, Cronin JT (2009) Patch dynamics of a native grass in relation to the spread of invasive smooth brome (Bromus inermis). Biol Invasions 11:1381–1391. https://doi.org/10.1007/s10530-008-9346-7
Dong X, Patton J, Wang G, Nyren P, Peterson P (2014) Effect of drought on biomass allocation in two invasive and two native grass species dominating the mixed-grass prairie. Grass and Forage Sci 69:160-166. https://doi.org/10.1111/gfs.12020
Finch DM, Butler JL, Runyon JB, Fettig CJ, Kilkenny FF, Jose S, Frankel SJ, Cushman SA, Cobb RC, Dukes JS, Hicke JA, Amelon SK (2021) Effects of climate change on invasive species. In: Invasive species in Forests and Rangelands of the United States: A comprehensive science synthesis for the United States Forest Sector (Poland TM, Patel-Weynand T, Miniat C, Hayes DC, Lopez VM eds.) Springer International Publishing Heidelberg, Germany. p. 57-84
Frank DA, McNaughton SJ (1992) The ecology of plants, large mammalian herbivores, and drought in Yellowstone National Park. Ecol. 73:2043-2058. https://doi.org/10.2307/1941454
Garnier E, Roy J (1988) Modular and demographic-analysis of plant leaf-area in sward and woodland populations of Dactylis glomerata and Bromus erectus. J Ecol 76:729-743. https://doi.org/10.2307/2260570
Gong K, Jin G, Sui X, Wang Y, Zhang X, Yuw Y (2019) Analysis on the distribution, breeding and utilization of Bromus inermis germplasm resource in China. Heilongjiang Animal Sci. and Vet. Medicine 21:33–36.
Grant TA, Shaffer TL, Flanders B (2020) Resiliency of Native Prairies to Invasion by Kentucky Bluegrass, Smooth Brome, and Woody Vegetation. Rangel. Ecol. & Manag 73:321–328. https://doi.org/10.1016/j.rama.2019.10.013
Harper JL (1985) Modules, branches, and the capture of resources. In: Population biology and evolution of clonal organisms (Jackson JBC, Buss LW, Cook RE eds.) Yale University Press, New Haven and London. p. 1-33.
Hartnett DC (1993) Regulation of clonal growth and dynamics of panicum-virgatum (Poaceae) in tallgrass prairie- effects of neighbor removal and nutrient addition. Am J Bot 80:1114-1120. https://doi.org/10.1002/j.1537-2197.1993.tb15342.x
Humphrey LD, Pyke DA (1998) Demographic and growth responses of a guerrilla and a phalanx perennial grass in competitive mixtures. J Ecol 86:854-865. https://doi.org/10.1046/j.1365-2745.1998.8650854.x.
Hutchings MJ, Slade AJ (1988) Aspects of the structure of clonal perennial herbs. In: Plant Form and Vegetation Structure: Adaptation, Plasticity and Relation to Herbivory (Werger MJA, Marinus J, Werger H, During J, Verhoeven JT eds.) SPB Academic Publishing, pp. 121-134.
Koerner SE, Collins SL, Blair JM, Knapp AK, Smith MD (2014) Rainfall variability has minimal effects on grassland recovery from repeated grazing. J Veg Sci 25:36-44. https://doi.org/10.1111/jvs.12065
Koerner SE, Avolio ML, Chang CC, Gray J, Hoover DL, Smith MD (2015) Invasibility of a mesic grassland depends on the time-scale of fluctuating resources. J Ecol 103:1538-1546 https://doi.org/10.1111/1365-2745.12479
Larson DL, Anderson PJ, Newton W (2001) Alien plant invasion in mixed-grass prairie: Effects of vegetation type and anthropogenic disturbance. Ecol Appl 11:128-141. https://doi.org/10.1890/1051-0761(2001)011[0128:APIIMG]2.0CO;2
Lovett-Doust LL (1981a) Population-dynamics and local specialization in a clonal perennial (Ranunculus repens). 1. The dynamics of ramets in contrasting habitats. J Ecol 69: 743-755. https://doi.org/10.2307/2259633
Lovett-Doust LL (1981b) Population-dynamics and local specialization in a clonal perennial (Ranunculus repens). 2. The dynamics of leaves, and a reciprocal transplant-replant experiment. J Ecol 69:757-768. https://doi.org/10.2307/2259634
Martínková J, Klimešová J (2017) Position of tillers in a clone determines their ontogeny: example of the clonal grass Phalris arundinacea. Folia Geobotanica 52:317-325
Moore KJ, Moser LE, Vogel KP, Waller SS, Johnson BE (1991) Describing and quantifying growth stages of perennial forage grasses. Agr J 83:1073-1077
Oborny B, Cain ML (1997) Models of spatial spread and foraging in clonal plants. In: Ecology and Evolution of Clonal Plants (de Kroon H, van Groenendael J eds.) Backbuy Pubshers, Leiden, The Netherlands. p. 155-183.
Otfinowski R, Kenkel NC (2008) Clonal integration facilitates the proliferation of smooth brome clones invading northern fescue prairies. Plant Ecol 199:235-242. https://doi.org/10.1007/s11258-008-9428-8
Otfinowski R, Kenkel NC, Catling PM (2007) The biology of Canadian weeds. 134. Bromus inermis Leyss. Can J Plant Sci 87: 183-198. https://doi.org/10.4141/P06-07
Ott JP, Hartnett DC (2015) Bud bank dynamics and clonal growth strategy in the rhizomatous grass, Pascopyrum smithii. Plant Ecol 216: 395-405. https://doi.org/10.1007/sl1258-014-0444-6
Ott JP, Butler, JL, Rong Y, Xu L (2017) Greater bud outgrowth of Bromus inermis than Pascopyrum smithii under multiple environmental conditions. J Plant Ecol 10: 518-527. https://doi.org/10.1093/jpe/rtw045
Ott JP, Klimešová J, Hartnett DC (2019) The ecology and significance of below-ground bud banks in plants. Ann Bot 123:1099-1118
Palit R, DeKeyser E (2022) Impacts and driver of smooth bromw (Bromus inermis Leyss.) invasion in native ecosystems. Plants 11: 1340 https://doi.org/10.3390/plants11101340
Saiz H, Bittebiere A-K, Benot M-L, Jung V, Mony C (2016) Understanding clonal plant competition for space over time: a fine-scale spatial approach based on experimental communities. J Veg Sci 27:759-770. https://doi.org/10.1111/jvs.12392
SAS Institute Inc. (2017) Studio 3.6 University Edition. Gary, NC.
Schmid B, Harper JL (1985) Clonal growth in grassland perennials .1. Density and pattern-dependent competition between plants with different growth forms. J Ecol 73:793-808. https://doi.org/10.2307/2260147
Schrag AM (2011) Addendum: Climate change impacts and adaptation strategies. In: Ocean of grass: a conservation assessment for the Northern Great Plains (Forrest SC, Strand H, Haskins WH, Freese C, Proctor J, Dinerstein E, eds.) Northern Plains Conservation Network and Northern Great Plains Ecoregion, World Wildlife Fund-US, Bozeman, MT.
Slade AJ, Hutchings MJ (1987) Clonal integration and plasticity in foraging behavior in Glechoma hederacea. J Ecol 75: 1023-1036
Stotz GC, Gianoli E, Cahill JF Jr. (2019) Biotic homogenization within and across eight widely distributed grasslands following invasion by Bromus inermis. Ecol 100:e02717. https://doi.org/10.1002/ecy.2717
Sutherland WJ, Stillman RA (1988) The Foraging Tactics of Plants. Oikos 52:239-244. https://doi.org/10.2307/3565196
Trammell M.A, Butler JL (1995) Effects of exotic plants on native ungulate use of habitat. J Wildlife Manage 59: 808-816. https://doi.org/10.2307/3801961
USDA-NRCS (2021) The PLANTS Database. National Plant Data Team, Greensboro, NC, USA. 2021 [cited 2021 6/13]; Available from: http://plants.usda.gov.
Wang YJ, Liu YY, Chen D, Du DL, Müller-Schärer, Yu FH (2022) Clonal functional traits favor the invasive success of alien plants into native communities. Ecol App e2756 https://doi.org/10.1002/eap.2756
Ye XH, Yu FH, Dong M (2006) A trade-off between guerrilla and phalanx growth forms in Leymus secalinus under different nutrient supplies. Ann Bot 98:187-191. https://doi.org/10.1016/j.flora.2010.07.006
Yu H, Wang L, Liu C, Yu D, Qu J (2020) Effect of a spatially heterogeneous nutrient distribution on the growth of clonal wetland plants. BMC Ecol. 20:59 https://doi.org/10.1186/s128998-020-00327-1
Zuo S, Liu L, Zhou C, Wang X, Huang D (2023) effects of nitrogen limitation on Leymuschinesis clonal growth form and bud bank. Flora 298:152186
Acknowledgments
The authors thank Dr. Brent Turnipseed at the Young Brothers Seed Technology Laboratory of South Dakota State University for contributing greenhouse space, M. Malecek, J. Shepherd, H. Becker, and A. Schauer for plant transplant and harvest assistance.
Funding
This study was supported in part by the USDA Forest Service, Rocky Mountain Research Station and the South Dakota Agriculture Experiment Station Hatch Project. The authors have no competing interests to declare that are relevant to the content of this article. Any opinions, findings, conclusions, or recommendations expressed in the material are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Author information
Authors and Affiliations
Contributions
Conceptualization, JPO, JLB, and LX; methodology, JPO, JLB, and LX; investigation, SB and LX; formal analysis, JPO and SB; writing—original draft preparation, JPO, SB, and LX; final draft preparation, JPO and LX; funding acquisition, JPO, JLB, and LX. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix
Appendix
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bam, S., Ott, J.P., Butler, J.L. et al. Belowground growth strategies of native and invasive rhizomatous perennial grasses in response to precipitation variability, clipping, and competition. Folia Geobot 58, 245–257 (2024). https://doi.org/10.1007/s12224-023-09438-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12224-023-09438-1