
Abstract
In order to correctly plan conservation and management of animal populations, it is fundamental to obtain reliable information on population abundance using cost-effective monitoring methods. In the case of small terrestrial vertebrates, capture–mark–recapture, removal sampling and counts of unmarked individuals have proven to be reliable techniques to estimate population abundance. In the present study, we applied a multinomial N-mixture modelling approach, performed through a double-observer sampling protocol, to estimate population size of the endemic terrestrial salamander Speleomantes ambrosii. We aimed at comparing cost-effectiveness of this protocol with those of other estimation methods (i.e., capture–mark–recapture and removal sampling), with particular attention to management and monitoring guidelines. The double-observer multinomial N-mixture protocol was the most cost-effective among the different methods: given its ease of application and cost-effectiveness, we encourage its employment for long-term monitoring and conservation plans of the near-threatened plethodontid salamander Speleomantes ambrosii.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
1 Introduction
In a context of “biodiversity crisis”, where amphibian populations are systematically declining worldwide (Stuart et al. 2004), the identification of less expensive monitoring methods for robust population estimates may promote investments into conservation actions. For this reason, it is essential to obtain reliable ecological and demographic information by correctly coordinating efforts and available resources to obtain accurate predictions, adequate monitoring plans, and effective conservation strategies (Williams et al. 2002; Griffiths et al. 2015). However, since it is unlikely that all individuals present within a population are detected during a sampling session (i.e., detection error is always present), accounting for detection probability is fundamental to obtain reliable estimates of population abundance (Schmidt 2003; Costa et al. 2020).
Capture–mark–recapture (CMR) or temporary removal sampling and related statistical methods have been widely used to estimate demographic parameters of terrestrial salamanders in their habitats, while accounting for imperfect detection (Salvidio and Pastorino 2002; Lunghi et al. 2019, 2020; Renet et al. 2019). The former consists in capturing a number of animals, marking them appropriately, releasing them back into the population, repeating this for subsequent sessions and then determining the ratio of marked to unmarked animals in the population, while in the latter, all animals captured are physically removed from the population, kept in a holding area, and released at the end of the study. However, these methods may be expensive in terms of time, materials, and sampling effort (Dodd 2016). The recent implementation of N-mixture models allows analysis of count data of unmarked animals without the need for temporal (i.e., space-for-time substitution; Kéry and Royle 2016) or spatial replication (i.e., time-for-space substitution; Costa et al. 2019, 2021). These models, despite some criticism regarding parameter identifiability, assumption violation, and overdispersion (e.g., Barker et al. 2018; Link et al. 2018; but see Kéry 2018 for a comprehensive and favorable analysis of N-mixture models), have proven to be reliable within a disparate range of applications, since they provide reliable estimates of abundance and other demographic parameters with reduced time and field effort (Ariefiandy et al. 2014; Ficetola et al. 2018; Broker et al. 2019; Costa et al. 2019).
In recent years, some studies applied multinomial N-mixture techniques, obtained within a multiple-observer protocol, for the estimation of population abundance of small cryptic terrestrial animals, such as amphibians and reptiles (Costa et al. 2020; Romano et al. 2021; Rosa et al. 2022). These studies, by comparing abundance estimates with other methods, such as removal sampling (Costa et al. 2020; Romano et al. 2021) or distance sampling (Rosa et al. 2022), showed that multinomial N-mixture models give reliable and robust estimates of population abundance, even when detection and density are relatively low. The reliability of this method, in underground habitats, has been validated by Costa et al. (2020), which compared this approach with a removal sampling protocol of three sessions on the congener Speleomantes strinatii, during the main activity period of the species (i.e., summer), demonstrating that during the population activity peak, a single visit is sufficient to approximate total population size and density. Therefore, we applied this multinomial N-mixture framework on the near-threatened cave salamander Speleomantes ambrosii in three underground environments, using a double-observer sampling protocol. The European plethodontid Speleomantes ambrosii, a fully terrestrial salamander endemic to a limited area in NW Italy, is listed as Near Threatened (NT) by IUCN (2009), given that its range is less than 5000 km2 and suitable habitat may be declining locally. The Ambrosi’s cave salamander is found in humid underground habitats such as natural and artificial caves, but also in humid forest environments (e.g., leaf litter near streams and on wet rocky outcrops) along small streams (Lanza 2007). Given the reliability and time-effectiveness of the double-observer sampling protocol, we compared the cost-effectiveness of this protocol with those of removal, binomial N-mixture models and CMR methods. The evaluation of this methodology–cost relationship could help to define conservation strategies and prioritization of a poorly known and small-range species, by promoting the selection of accurate and precise estimators and using financial resources more efficiently.
2 Materials and methods
2.1 Survey design
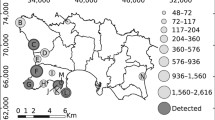
The studied sites are three natural caves located in the Province of La Spezia (Liguria region, NW Italy; Fig. 1) that extend underground 15, 27, and 39 m, respectively. Each cave was divided using a metal tape ruler into 1-m linear sections and extending on both side walls and on the ceiling. These 1-m sections were used as spatial replicates (i.e., plots) for the application of multinomial N-mixture models. Therefore, we sampled 15, 27, and 39 plots in Cave 1, 2, and 3, respectively. Since the multinomial protocol employed does not require multiple visits and salamanders did not move from one plot to another during sampling, we are not concerned about the demographic closure assumption and independence of sites (Royle 2004). We are not even concerned about the spreading of fungal infection, since, before entering the caves, we followed the procedures provided by Garner et al. (2016) for mitigating amphibian disease.

Map showing species distribution range (orange area), provided by IUCN (2009), and location of the three sampling sites, across Northern Italy
In September 2021, two experienced observers searched for salamanders with the aid of head lamps in all the 1-m plots, by applying a dependent-double-observer (DDO) method (Nichols et al. 2000). In the DDO method, a primary observer reports all the individuals counted to a secondary observer who records the data. In addition, the secondary observer takes records of any salamander that the primary observer failed to detect (Nichols et al. 2000). In the field, the two researchers alternated as primary and secondary observer. This protocol yields to three possible multinomial outcomes. An individual could be detected: (i) by the primary observer; (ii) by the secondary observer but not by the primary observer; (iii) by neither of the two observers (i.e., the quantity to estimate).
2.2 N-mixture modelling
Data were analyzed with a Poisson error distribution for the mean abundance at each sampling plot (λ) and considering constant detection probability across observers (Royle and Dorazio 2006). We modelled λ accounting also for the distance from the entrance of each cave, to test if animal distribution changed from the twilight zone to the inner parts of the underground environment. We selected the best between these two models (i.e., null model and model with the effect of distance from cave entrance) according to the Akaike’s Information Criterion and considered that a ΔAICc > 4 prove substantial differences (Burnham and Anderson 2002). We used a goodness-of-fit (GOF) test for model assessment. Total population estimates (N-hat) for each cave, along with 95% confidence intervals (CI), were obtained by posterior distribution of the latent abundance from the best model (Kery and Royle 2016). We conducted all statistical analyses in R with the package “unmarked”, through the function “gmultmix” (Fiske and Chandler 2011), and ‘‘AICcmodavg’’ (Mazerolle 2017) and built graphs using the package “forestplot” (Gordon and Lumley 2019).
2.3 Cost-effectiveness analysis
Finally, we provided a detailed estimate of costs for the application of this sampling protocol, compared with temporary removal and the CMR methods. In the case of removals, we considered three successive sampling occasions, this being also the usual minimum requirement for the application of repeated counts on unmarked individuals in the framework of binomial N-mixture modelling to obtain reliable estimates (Ficetola et al. 2018; Costa et al. 2021). To attain the final costs, we calculated the expenses required for monitoring all the sites where the presence of the species is confirmed. We used the available data on Global Biodiversity Information Facility (GBIF) and the Ligurian Regional Cartographic Service. We considered both real costs such as researchers’ remuneration, field trip, and equipment and survey costs, calculated as hours and days of working effort (Ariefiandy et al. 2014; Romano et al. 2017). The estimates of costs regarding the equipment (laser meter for measuring caves’ plots during DDO monitoring, fauna boxes for storing animals during removal sampling and camera to photograph animals in a CMR study) were obtained by the mean of mid-range equipment costs from international online sellers, while the expenditure for researchers’ remuneration and accommodation has been calculated on the base of the average international values (euros and dollars). Concerning data analyses, we did not consider the remuneration cost for researchers, but only estimated the level of complexity of each analysis, since time for data computerization and computation could be highly variable. Finally, we calculated the cost per kilometer during field trips on the basis of the Automobile Club Italia (ACI) parameters.
3 Results
We counted a total of 202, 7 and 34 salamanders, in Cave 1, 2, and 3, respectively. The best model for site abundance was that accounting for the distance from the entrance (AICc = − 338.92 vs AICcdist = − 363.54), which had a significant negative effect on λ = 0.602; SE = 0.120; p value < 0.001). The GOF showed a good fit of the model with the data (p value = 0.153; c-hat = 1.2). Detection estimates were very similar among sites and all > 0.80 (Fig. 2). Population estimates from the best model were N-hat1 = 207 (95% CI 185–231), N-hat2 = 7 (95% CI 2–13), and N-hat3 = 35 (95% CI 24–46). Plot abundance was also very different among sites (Table 1) with λ1 = 14 (95% CI 12–16), λ2 = 0.27 (95% CI 0.12–0.56), and λ3 = 0.89 (95% CI 0.63–1.24).

Forest plot with total population estimates (left) and estimated site detections (right), along with 95% confidence intervals (horizontal bars) for the three sites
The estimated total cost for the DDO method was 4624 €, while for removal sampling and CMR method was 6930 € and 8506 €, respectively (Table 2). The highest gap was related to the remuneration for research officers conducting field surveys, given the greater number of sampling days for removal sampling (20) and CMR (24) compared to DDO (7), while there was no difference related to sampling material.
4 Discussion
Detection probabilities were high and very similar in the three study sites, despite the different estimated abundances (Table 1). These results resemble those obtained with other congeneric Speleomantes species, whose individual mean capture probabilities in subterranean environments are usually > 0.50 (Lindstrom et al. 2010; Lunghi et al. 2018; Costa et al. 2020).
The negative effect of cave depth on λ seems to be in good agreement with the results of previous studies on Speleomantes (Cimmaruta et al. 1999; Lunghi et al. 2015; Salvidio et al. 2020). In fact, the spatial distribution of salamanders along the cave is affected by the interaction between multiple environmental and social factors (e.g., light, humidity, temperature, and competition). Specifically, there is a clear age-class segregation, with juveniles living in more external cave sectors than adults which, in turn, are dispersed along the inner parts of the cave, with high abundances in the intermediate sectors (Salvidio and Pastorino 2002; Ficetola et al. 2013; Salvidio et al. 2020). To better understand what influences population abundance and spatial distribution of the Ambrosi’s cave salamander, it will be necessary to carry out further investigations, taking into account age classes and environmental factors.
The estimated field activity to evaluate the abundances of Speleomantes ambrosii populations within its distributional range, through the DDO approach, required only 7 working days for a two-person team, about a third less than removal sampling, binomial N-mixture models, and CMR. This difference is due to the characteristics of the different sampling protocols. In fact, to obtain reliable estimates through removal sampling, binomial N-mixture, or CMR, it is usually necessary to carry out at least three sampling sessions for each site, in contrast to the single session of the dependent double-observer multinomial N-mixture protocol used in this study (Ficetola et al. 2018; Costa et al. 2020). In any case, the correct application of the DDO sampling protocol should require a very good knowledge of the species seasonal activity pattern (e.g., with a pilot survey over the entire population activity period), because DDO has to be performed during the period of highest activity of the focal population. Since in the present study, we performed field sampling during the activity peak of the species, we are confident that our abundance estimates of the active population are representative of the entire population, even if not accounting directly for availability, and our results are comparable with those obtained with removal sampling or CMR protocol, as reported by Costa et al. (2020) after comparing DO with a multi-day removal sampling on the congener S. strinatii on similar conditions. Furthermore, the manipulation of animals, required by methods that need individual marking or physical capture, implies the request for official capture and handling permits to be issued by the Italian Ministry of Environment or local authorities, whose processing may take a long time, and a greater risk of spreading amphibians’ pathogens, such as chytridiomycosis.
5 Conclusions
The benefits related to the application of the DDO sampling method in a hierarchical framework are several, since it is a reliable, quick, and economic protocol to estimate population abundance of amphibians without the need of handling individuals, preventing any possible negative effects on animals. Given the reliability, the easy application and the cost-effectiveness of this sampling protocol, we suggest that the employment of this sampling framework should be considered when planning conservation strategies or long-term monitoring of Speleomantes ambrosii and other terrestrial salamander species.
References
Ariefiandy A, Purwandana D, Seno A, Chrismiawati M, Ciofi C, Jessop TS (2014) Evaluation of three field monitoring-density estimation protocols and their relevance to Komodo dragon conservation. Biodivers Conserv 23:2473–2490. https://doi.org/10.1007/s10531-014-0733-3
Barker RJ, Schofield MR, Link WA, Sauer JR (2018) On the reliability of N-mixture models for count data. Biometrics 74:369–377. https://doi.org/10.1111/biom.12734
Broker KCA, Hansen RG, Leonard KE, Koski WR, Heide-Jørgensen MP (2019) A comparison of image and observer based aerial surveys of narwhal. Mar Mam Sci 35:1253–1279. https://doi.org/10.1111/mms.12586
Burnham KP, Anderson DR (2002) Model selection multimodel inference a practical information-theoretic approach. Springer, New York
Cimmaruta R, Forti G, Nascetti G, Bullini L (1999) Spatial distribution and competition in two parapatric sibling species of European plethodontid salamanders. Ethol Ecol Evol 4:383–398. https://doi.org/10.1080/08927014.1999.9522821
Costa A, Oneto F, Salvidio S (2019) Time-for-space substitution in N-mixture modeling and population monitoring. J Wildlife Manage 83:737–741. https://doi.org/10.1002/jwmg.21621
Costa A, Romano A, Salvidio S (2020) Reliability of multinomial N-mixture models for estimating abundance of small terrestrial vertebrates. Biodivers Conserv 29:2951–2965. https://doi.org/10.1007/s10531-020-02006-5
Costa A, Salvidio S, Penner J, Basile M (2021) Time-for-space substitution in N-mixture models for estimating population trends: a simulation-based evaluation. Sci Rep 11:1–10. https://doi.org/10.1038/s41598-021-84010-5
Dodd CK (2016) Reptile ecology and conservation. Oxford Univ. Press, Oxford
Ficetola GF, Pennati R, Manenti R (2013) Spatial segregation among age classes in cave salamanders: Habitat selection or social interactions? Popul Ecol 55:217–226. https://doi.org/10.1007/s10144-012-0350-5
Ficetola GF, Barzaghi B, Melotto A, Muraro M, Lunghi E, Canedoli C, Carretero MA (2018) N-mixture models reliably estimate the abundance of small vertebrates. Sci Rep 8:10357. https://doi.org/10.1038/s41598-018-28432-8
Fiske I, Chandler R (2011) Unmarked: an R package for fitting hierarchical models of wildlife occurrence and abundance. J Stat Softw 43:1–23. https://doi.org/10.18637/jss.v043.i10
Garner TWJ, Schmidt BR, Martel A, Pasmans F, Muths E, Cunningham AA, Weldon C, Fisher MC, Bosch J (2016) Mitigating amphibian chytridiomycoses in nature. Phil Trans R Soc 371(1709):20160207. https://doi.org/10.1098/rstb.2016.0207
GBIF.org. GBIF Occurrence Download. Available at https://doi.org/10.15468/dl.fe5dtd.
Gordon M, Lumley T (2019) Package ‘forestplot’. Advanced forest plot using ‘grid’ graphics. The Comprehensive R Archive Network, Vienna
Griffiths RA, Foster J, Wilkinson JW, Sewell D (2015) Science, statistics and surveys: a herpetological perspective. J Appl Ecol 52:1413–1417. https://doi.org/10.1111/1365-2664.12463
IUCN (2009) European Species on the IUCN Red List. Available at: http://www.iucnredlist.org/europe.
Kéry M (2018) Identifiability in N-mixture models: a large-scale screening test with bird data. Ecology 99:281–288. https://doi.org/10.1002/ecy.2093
Kéry M, Royle JA (2016) Applied hierarchical modelling in ecology. Academic Press, Cambridge
Lanza B (2007) Speleomantes strinatii (Aellen). In: Lanza B, Andreone F, Bologna MA, Corti C, Razzetti E (eds) Fauna d’Italia-amphibia. Edizioni Calderini, Bologna, pp 152–156
Lindstrom L, Reeve R, Salvidio S (2010) Bayesian salamanders: analysing the demography of an underground population of the European plethodontid Speleomantes strinatii with state-space modelling. BMC Ecol 10:4. https://doi.org/10.1186/1472-6785-10-4
Link WA, Schofield MR, Barker RJ, Sauer JR (2018) On the robustness of N-mixture models. Ecology 99:1547–1551
Lunghi E, Manenti R, Ficetola GF (2015) Seasonal variation in microhabitat of salamanders: environmental variation or shift of habitat selection? PeerJ 3:e1122. https://doi.org/10.7717/peerj.1122
Lunghi E, Manenti R, Mulargia M, Veith M, Corti C, Ficetola GF (2018) Environmental suitability models predict population density, performance and body condition for microendemic salamanders. Sci Rep 8:7527. https://doi.org/10.1038/s41598-018-25704-1
Lunghi E, Giachello S, Zhao Y, Corti C, Ficetola GF, Manenti R (2020) Photographic database of the European cave salamanders, genus Hydromantes. Sci Data 7:171. https://doi.org/10.1038/s41597-020-0513-8
Lunghi E, Romeo D, Mulargia M, Cogoni R, Manenti R, Corti C, Ficetola GF, Veith M (2019) On the stability of the dorsal pattern of European cave salamanders (genus Hydromantes). Herpetozoa 32: 249–253. http://hdl.handle.net/2434/733296
Mazerolle MJ (2017) Package ‘AICcmodavg’. R package 281
Nichols JD, Hines JE, Sauer JR, Fallon F, Fallon J, Heglund PJ (2000) A double-observer approach for estimating detection probability and abundance from avian point counts. Auk 117:393–408. https://doi.org/10.1093/auk/117.2.393
Renet J, Leprêtre L, Champagnon J, Lambret P (2019) Monitoring amphibian species with complex chromatophore patterns: a non-invasive approach with an evaluation of software effectiveness and reliability. Herpetol J 29:13–22. https://doi.org/10.33256/hj29.1.1322
Romano A, Costa A, Basile M, Raimondi R, Posillico M, Scinti D, Crisci A, Piraccini R, Raia P, Matteucci G, De Cinti B (2017) Conservation of salamanders in managed forests: methods and costs of monitoring abundance and habitat selection. For Ecol Manage 400:12–18. https://doi.org/10.1016/j.foreco.2017.05.048
Romano A, Roner L, Costa A, Salvidio S, Trenti M, Pedrini P (2021) When no color pattern is available: application of double observer methods to estimate population size of the Alpine salamander. Arct Antart Alp Res 53:300–308. https://doi.org/10.1080/15230430.2021.1994103
Rosa G, Salvidio S, Trombini E, Costa A (2022) Estimating density of terrestrial reptiles in forest habitats: the importance of considering availability in distance sampling protocols. Trees for People 7:100184. https://doi.org/10.1016/j.tfp.2021.100184
Royle JA (2004) N-mixture models for estimating population size from spatially replicated counts. Biometrics 60:108–115. https://doi.org/10.1111/j.0006-341X.2004.00142.x
Royle JA, Dorazio RM (2006) Hierarchical models of animal abundance and occurrence. J Agric Biol Environ Stat 11:249–263. https://doi.org/10.1198/108571106X129153
Salvidio S, Pastorino MV (2002) Spatial segregation in the European plethodontid salamander Speleomantes strinatii in relation to age and sex. AMphib Reptil 23:505–510
Salvidio S, Costa A, Oneto F, Pastorino MV (2020) Variability of a subterranean prey-predator community in space and time. Diversity 12:17. https://doi.org/10.3390/d12010017
Schmidt BR (2003) Count data, detection probabilities, and the demography, dynamics, distribution, and decline of amphibians. CR Biol 326:119–124. https://doi.org/10.1016/S1631-0691(03)00048-9
Stuart SN, Chanson JS, Cox NA, Young BE, Rodrigues ASL, Fischman DL, Waller RW (2004) Status and trends of amphibian declines and extinctions worldwide. Science 306:1783–1786. https://doi.org/10.1126/science.1103538
Williams BK, Nichols JD, Conroy MJ (2002) Analysis and management of animal populations. Academic Press, Cambridge
Acknowledgements
I warmly thank Sebastiano Salvidio for his assistance during field sampling and Andrea Costa for his suggestions concerning data analysis. Research permits were issued by the Italian Ministry of Environment (no 0039130 of 15/04/2021).
Funding
Open access funding provided by Università degli Studi di Genova within the CRUI-CARE Agreement.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rosa, G. Optimizing monitoring of an endemic terrestrial salamander (Speleomantes ambrosii): comparing cost-effectiveness of different methods for abundance estimation. Rend. Fis. Acc. Lincei 33, 807–813 (2022). https://doi.org/10.1007/s12210-022-01099-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12210-022-01099-0