
Abstract
ZCCHC9 is a human nuclear protein with sequence homology to yeast Air1p/Air2p proteins which are RNA-binding subunits of the Trf4/Air2/Mtr4 polyadenylation (TRAMP) complex involved in nuclear RNA quality control and degradation in yeast. The ZCCHC9 protein contains four retroviral-type zinc knuckle motifs. Here, we report the NMR spectral assignment of the zinc knuckle region of ZCCHC9. These data will allow performing NMR structural and RNA-binding studies of ZCCHC9 with the aim to investigate its role in the RNA quality control in human.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Biological context
RNA processing in eukaryotes has to undergo a strict RNA quality control. In yeast nucleus, aberrant or short-lived RNAs are degraded by the TRAMP-exosome pathway. TRAMP is a poly(A) polymerase complex consisting of the Trf4 or Trf5 non-canonical polymerases, the Air1 or Air2 retroviral-type zinc knuckle proteins as potential RNA-binding subunits and the Mtr4 RNA helicase (Vanacova and Stefl 2007). The complex acts on diverse classes of RNAs (San Paolo et al. 2009; Vanacova and Stefl 2007). It can add short poly(A) tails to 3′end of the RNA molecules to recruit the nuclear exosome nucleolytic activities (Vanacova and Stefl 2007). However, in vivo, poly(A) polymerase activity is dispensable for the degradation of most TRAMP4 RNA targets (San Paolo et al. 2009). The minimal TRAMP poly(A) polymerase consists of Trf4/Trf5 and Air1 or Air2 heteromeric complexes (Vanacova and Stefl 2007). It has been proposed that Air1p and Air2p, respectively support RNA recognition and binding while Trf4/5 proteins provide the polyadenylation activity (Vanacova and Stefl 2007). Air1/2 contain 4–5 CCHC zinc knuckle motifs. Zinc knuckle motifs of other proteins are known to contact RNA molecules (D’Souza and Summers 2004).
Human genome encodes for at least 22 zinc knuckle proteins of the CCHC-type (identified by HUGO Gene Nomenclature Committee), in most cases rather poorly characterized. Our BLAST search identified three closest homologues of Air1/2p; the ZCCHC7, ZCCHC3, ZCCHC9 proteins. No relevant functional information have been reported on ZCCHC7 nor ZCCHC3 to date and only little is known about ZCCHC9. ZCCHC9 (zinc finger, CCHC domain containing 9; alias DKFZp761J139) is a nucleolar protein (Zhou et al. 2008). Based on mRNA expression level profiling, it shows the highest expression levels in mouse brain, particularly cortex, and testicles. Genome-wide expression profiling study indicated ZCCHC9 up-regulation in the human neck cancers cells both HPV (human papillomaviruses) positive or negative and cervical cancer cells relative to normal cells (Pyeon et al. 2007). Overexpression of ZCCHC9 in HEK293 cells lead to the suppression of the transactivation by NF-kappa B and SRE, therefore ZCCHC9 has been implicated to be involved in the MAPK signaling cascade. However, the mechanism of ZCCHC9 function remains elusive.
ZCCHC9 contains 4 retroviral-type zinc knuckle motifs with consensus C–X2–C–X4–H–X4-C that is found mainly in the nucleocapsid protein of retroviruses and also in eukaryotic proteins involved in RNA or ssDNA binding (D’Souza and Summers 2005). To gain structural and functional insights into the mechanism of ZCCHC9 MAPK regulation and the involvement of ZCCHC9 in RNA metabolism, we have initiated an NMR study of ZCCHC9. Here, we report 1H, 13C, and 15N resonance assignments of ZCCHC9.
Methods and experiments
The coding sequence corresponding to the zinc knuckle region of the ZCCHC9 gene (accession number BC014841, Ref. Seq. ID NM_032280) from Homo sapiens (318 bp) was amplified by PCR and cloned into a pET30-G protein B1 fusion vector via BamHI and XhoI restriction sites (Zhou et al. 2001). A non-cleavable solubility-enhancement tag, GB1, was fused to the ZCCHC9 protein to enhance its solubility limit (Zhou et al. 2001), yielding a 175 amino acid construct (GB1 (58 aa) + ZCCHC9 (106 aa) + linker (5 aa) + hexahistidine tag). This tag is often used for NMR and X-ray studies of poorly behaving proteins (Zhou et al. 2001). The resulting N-terminal GB1 and C-terminal 6× histidine-tagged construct was verified by DNA sequencing. The protein was overexpressed in E.coli BL21-CodonPlus(DE3)-RIPL (Stratagene) at 37°C in M9 minimal medium, supplemented with 50 μM ZnSO4. For isotope labeling, the medium was supplemented with 15NH4Cl and U-13C6-glucose. Cells were grown at 37°C to OD600 ~0.5 and induced with 400 μM isopropyl-β-d-thiogalactoside (IPTG) for 6 h at 37°C. Cells were harvested by centrifugation (4,000 rpm, 10 min, 4°C), resuspended in lysis buffer (20 mM Tris pH 7.0; 400 mM NaCl; 100 μM ZnSO4; 10% glycerol; 25% sacharose; 2 mM MgCl2; 5 mM DTT; 500 μM PMSF; 2 μg/ml leupeptin; 1 μg/ml pepstatin) and disrupted by sonication. An equal volume of buffer A (20 mM Tris pH 7.0, 400 mM NaCl, 100 μM ZnSO4, 10% glycerol, 1% Nonidet P40, 40 mM imidazole, 5 mM DTT, 500 μM PMSF, 2 μg/ml leupeptin, 1 μg/ml pepstatin) was added to the lysate. The cell debris was cleared by centrifugation (14 000 rpm, 45 min, 4°C). The supernatant was next incubated for 45 min at 4°C with Ni-NTA agarose beads (QIAGEN), which were pre-equilibrated with a buffer B (20 mM Tris pH 7.0, 400 mM NaCl, 20 mM imidazole). The beads were washed with 10 volumes of wash buffer B and the protein was eluted in buffer EB (20 mM Tris pH 7.0, 400 mM NaCl, 100 μM ZnSO4, 10% glycerol, 5 mM DTT, 1 mM PMSF, 400 mM imidazole). The purified protein was dialyzed to buffer C (20 mM Tris pH 7.0, 200 mM NaCl, 100 μM ZnSO4, 14 mM 2-mercaptoethanol) and concentrated to ~1.5 mM in 550 μl of buffer C.
All NMR experiments were recorded at 293 K on a Bruker AVANCE 600 MHz spectrometer equipped with a cryogenic triple resonance probe (Bruker BioSpin). The NMR spectra were processed using the TopSpin 2.1 (Bruker BioSpin) and NMRPipe (Delaglio et al. 1995). The chemical shifts of backbone and side-chains were assigned manually using Sparky software (Goddard T.G. and Kellner D.G., University of California, San Francisco). For backbone resonance assignment, the following standard triple resonance experiments were carried out: HNCA, HNCACO, HNCACB (Sattler et al. 1999). For side-chain resonance assignment, 2D 1H-13C HMQC, 3D 1H-13C NOESY-HSQC, 1H-15N NOESY-HSQC (both NOESY with mixing time of 150 ms), 3D HNHA, 3D HCCH-TOCSY and 2D HB(CB)(CGCD)HD, were recorded (Sattler et al. 1999).
Extent of assignments and data deposition
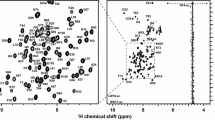
The 2D 1H-15N HSQC spectrum of GB1-ZCCHC9 is shown in Fig. 1 with indicated assignments. With standard sequential assignment procedure, near complete assignments of the backbone were obtained. For the entire protein construct, 98% of backbone amide protons and 15N nuclei, 98% of the Cα and Cβ were assigned; prolines and M1 not counted. The chemical shift deviations of Cα and Hα of the assigned backbone resonances of GB1-ZCCHC9 from the sequence-dependent random coil values (data not shown) agree well with the canonical secondary structure elements of the GB1 domain (β1β2α1β3β4; Gronenborn et al. 1991) and show no pattern for the ZCCHC9 region. The latter agrees well with the absence of secondary structure elements in the topology of zinc knuckles (D’Souza and Summers 2004). The histidines of CCHC motif of zinc knuckles (H71, H98, H127, and H154) are involved in coordination of zinc ions as the N-H correlations of these residues span the same region in the 2D 1H-15N HSQC (Fig. 1) that is characteristic for zinc fingers and knuckles. The histidine C2 chemical shifts that are sensitive probes to monitor the histidine protonation state, indicate that the histidines are not protonated under the conditions used in our study (pH 7). Further, the presence of folded zinc knuckles was confirmed by the addition of EDTA in excess of zinc ions that resulted in a loss of dispersion of many resonances in the 2D 1H-15N HSQC (data not shown), and consequently, in significant precipitation of the protein. All assignment for 1H, 15N and 13C backbone and side-chain chemical shifts of ZCCHC9 have been deposited at the BMRB (http://www.bmrb.wisc.edu) and can be accessed under the accession number 16958.

2D [1H-15N]-HSQC spectrum of 1.5 mM uniformly [15N,13C]-labeled ZCCHC9 in 20 mM Tris buffer (pH 7.0), 200 mM NaCl, 100 μM ZnSO4, 14 mM 2-mercaptoethanol, (90% H2O/10% D2O). The spectrum was acquired at 293 K on a Bruker Avance 600 MHz spectrometer. The assignments are labeled by the one-letter code of amino acids accompanied by a sequence number. The side-chain resonances of asparagine and glutamine are connected by horizontal lines
References
D’Souza V, Summers MF (2004) Structural basis for packaging the dimeric genome of Moloney murine leukaemia virus. Nature 431:586–590
D’Souza V, Summers MF (2005) How retroviruses select their genomes. Nat Rev Microbiol 3:643–655
Delaglio F, Grzesiek S, Vuister GW, Zhu G, Pfeifer J, Bax A (1995) NMRPipe: a multidimensional spectral processing system based on UNIX pipes. J Biomol NMR 6:277–293
Gronenborn AM, Filpula DR, Essig NZ, Achari A, Whitlow M, Wingfield PT, Clore GM (1991) A novel, highly stable fold of the immunoglobulin binding domain of streptococcal protein G. Science 253:657–661
Pyeon D, Newton MA, Lambert PF, den Boon JA, Sengupta S, Marsit CJ, Woodworth CD, Connor JP, Haugen TH, Smith EM et al (2007) Fundamental differences in cell cycle deregulation in human papillomavirus-positive and human papillomavirus-negative head/neck and cervical cancers. Cancer Res 67:4605–4619
San Paolo S, Vanacova S, Schenk L, Scherrer T, Blank D, Keller W, Gerber AP (2009) Distinct roles of non-canonical poly(A) polymerases in RNA metabolism. PLoS Genet 5:e1000555
Sattler M, Schleucher J, Griesinger C (1999) Heteronuclear multidimensional NMR experiments for the structure determination of proteins in solution employing pulsed field gradients. Prog Nucl Magn Reson Spectrosc 34:93–158
Vanacova S, Stefl R (2007) The exosome and RNA quality control in the nucleus. EMBO Rep 8:651–657
Zhou A, Zhou J, Yang L, Liu M, Li H, Xu S, Han M, Zhang J (2008) A nuclear localized protein ZCCHC9 is expressed in cerebral cortex and suppresses the MAPK signal pathway. J Genet Genomics 35:467–472
Zhou P, Lugovskoy AA, Wagner G (2001) A solubility-enhancement tag (SET) for NMR studies of poorly behaving proteins. J Biomol NMR 20:11–14
Acknowledgments
This work was supported by Wellcome Trust International Senior Fellowship 084316/Z/07/Z (to S.V.), EMBO Installation Grant Project No. 1642 (to S.V.), European Molecular Biology Organization/Howard Hughes Medical Institute Start-up Grant No. 1263 (to R.S.), and Human Frontier Science Program Career Development Award No. 0049/2006 (to R.S.), Ministry of Education of the Czech Republic (MSM0021622413, Ingo LA08008), GACR (204/08/1212 and 305/10/1490), GAAV (IAA401630903). MS is supported by the EC FP-7 (grant no. 205872).
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Maria Sanudo and Martin Jacko these authors contributed equally to this work.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Sanudo, M., Jacko, M., Rammelt, C. et al. 1H, 13C, and 15N chemical shift assignments of ZCCHC9. Biomol NMR Assign 5, 19–21 (2011). https://doi.org/10.1007/s12104-010-9257-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12104-010-9257-2