
Abstract
Introduction
Due to its lack of conventional surface receptors, triple-negative breast cancer (TNBC) is inherently resistant to most targeted therapies. MAL2 overexpression prompts endocytosis, conferring resistance to novel therapeutics. This study explores the role of MAL2 and PD-L1 in TNBC patients’ prognosis.
Methods
We performed immunohistochemical analysis on 111 TNBC samples collected from 76 patients and evaluated the expression of MAL2 and PD-1. We expanded the study by including The Cancer Genome Atlas (TCGA) cohort.
Results
MAL2 expression did not correlate with stage, grade, tumor size, lymph node invasion, metastasis, and PD-1 expression. Patients with high MAL2 had significantly lower 5-year survival rates (71.33% vs. 89.59%, p = 0.0224). In the tissue microarray cohort (TMA), node invasions, size, recurrence, and low MAL2 (HR 0.29 [CI 95% 0.087–0.95]; p < 0.05) predicted longer patients’ survival. In the TCGA cohort, patients with low MAL2 had significantly longer overall survival and disease-specific survival than patients with high MAL2. Older age and high MAL2 expression were the only independent predictors of shorter patient survival in the BRCA TCGA cohort.
Conclusion
High MAL2 predicts unfavorable prognosis in triple-negative breast cancer, and its expression is independent of PD-1 levels and clinicopathological features of TNBC.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Breast cancer is the most common female malignancy worldwide. Although overall survival has significantly increased over the past few years, it remains the second most prevalent cause of cancer-related deaths in women, accounting for 2.3 million new cases and 685,000 deaths in 2020 [1]. Triple-negative breast cancer (TNBC) accounts for approximately 15–20% of all breast cancer cases and lacks the expression of estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2) [2]. This aggressive subtype of breast cancer affects predominantly young patients, especially those under 40 and African Americans, who rarely respond to standard targeted therapies and progress aggressively [2,3,4]. While early detection increases the chances of radical treatment and improves prognosis, current treatment modalities are limited to surgery, chemotherapy, radiation therapy, and only a few targeted therapies. Therefore, there exists a critical need to find new biomarkers and develop novel approaches to treat triple-negative breast cancer [5, 6].
Genetic and molecular factors play a key role in the development and progression of TNBC. Considering its inherent resistance to targeted therapies, developing novel prognostic markers became crucial to planning treatment, assessing disease progression, and stratifying patients’ risk [6]. Recently, T Cell Differentiation Protein 2 (MAL2) emerged as a potential driver of cancer immune escape and novel therapeutic target [7]. MAL2 was overexpressed in tumors and was associated with poor prognosis, and MAL2 knockdown improved major histocompatibility complex type 1 (MHC-1) recognition on tumor surfaces by CD8 + T cells, highlighting the role of MAL2 in immune regulation [8,9,10,11]. While the exact mechanism remains unknown, reducing MAL2 expression may be a novel therapeutic approach. More research is needed to fully understand the protein potential.
Targeting PD-1/PD-L1 has recently revolutionized patient care and started the era of immunotherapy in TNBC. PD-L1 is present on the surface of tumor cells and binds with Programmed Cell Death Protein 1 (PD-1) present on T cells, rendering them unable to induce antitumor response [12, 13]. Approximately 20% of TNBCs express Programmed Cell Death Ligand 1 (PD-L1), indicating their potential sensitivity to immune checkpoint inhibitors [14]. Blocking PD-1/PD-L1 interactions significantly improved the efficacy of cancer treatments in solid tumors, including breast cancer [15, 16]. While not all anti-PD-L1 regimens showed clinical benefits, blocking this interaction remains a promising treatment option [17]. Despite their distinct mechanisms of action, MAL2 and PD-1/PD-L1 expression suppress immune surveillance and facilitate cancer progression [9, 13]. Therefore, their concurrent targeting may hold the premise of improving therapeutic strategies to treat TNBC and prolong patient survival.
In this study, we explore the role of MAL2 in TNBC and investigate its interactions with PD-1 and PD-L1. Since both MAL2 and PD-L1 modulate tumor immune response, confirming their interactions may uncover the role of MAL2 as a novel prognostic marker and therapeutic target in TNBC.
Materials and methods
Sample acquisition
This study included 111 triple-negative breast cancer samples collected from 76 women in the Department of Pathomorphology between 2011 and 2015. Clinical data, including age, sex, overall survival, tumor differentiation (grade), stage T, lymph node invasion, metastasis, tumor size, and recurrence, were obtained and analyzed retrospectively (Table 1). The study followed the Declaration of Helsinki, and the protocol was approved by the Nicolaus Copernicus University Bioethics Committee (KB 746/2021). All 76 patients were treated by surgery, and the resection was radical in 75 cases. In 57 cases, it was breast-conserving surgery; in 19 cases, radical mastectomy was conducted. Three patients underwent neoadjuvant chemotherapy. Of 73 patients qualified for adjuvant therapy, 24 underwent chemotherapy, 47 chemotherapy and radiotherapy, while 2 were radiotherapy only. Three patients were disqualified from adjuvant therapy due to the high risk of adverse events relative to comorbidities. The high and low MAL2 expression groups (extracted from the tissue microarray (TMA), TCGA breast cancer, and TCGA TNBC cohorts) did not differ significantly in regard to administered treatment (p > 0.05).
Immunohistochemistry
A retrospective immunohistochemical analysis of MAL2 comprised 111 formalin-fixed, paraffin-embedded tissue blocks cut into 5 μm sections, attached to a glass slide, and incubated at 60 °C for 2 h. IHC staining was performed on the Ventana Benchmark Ultra platform according to NordiQC operating procedure. A primary MAL2 polyclonal antibody (ab75347, Abcam) was used for staining. The specimens were fixed in 10% buffered formalin for 24 h at room temperature.
Image acquisition and analysis
The pathologists evaluating the immunohistochemical expression of PD-L1 worked independently and were blinded to clinical and pathological data. Protein expression was evaluated using a light microscope at 20 × original objective magnification. All of the collected tissue samples were processed following the standard diagnostic protocol. For this study, representative material from diagnostic biopsies and matching resection specimens was selected for additional immunohistochemical studies. The immunohistochemical studies of PD-L1 expression were performed using an anti-PD-L1 antibody (clone SP142, Ventana Medical Systems) on material from diagnostic biopsies, and the original protocol provided by Ventana was followed. Finally, the brown color due to the histochemical reaction product was considered as observed in the site of the presence of the searched antigen. According to the guide provided by Ventana, tumors with PD-L1 expression ≥ 1% IC were considered PD-L1-positive [18].
Matching resection specimens were stained to assess MAL2 expression. For each sample, the expression of MAL2 was obtained by calculating the H-score in three representative areas of the tumor. The H-score was assigned using the formula [1 × (% cells low positive) + 2 × (% cells positive) + 3 × (%cells high positive)], obtaining a value from 0 to 300. The final result was the mean value of the three assessments [19] (Fig. 1).

Cross-section TNBC staining patterns: a H&E, b high MAL2 expression, c low MAL2 expression, d negative MAL2 expression, e negative PD-L1 expression, and f positive PD-L1 expression
In silico analysis
TCGA cohort clinical data were accessed through the cBioPortal [20]. USCS XENA was used to acquire normalized expression data of MAL2 and PD-1 [21]. FPKM gene expression was obtained through the Human Protein Atlas [22, 23]. Normalized gene expression was calculated as log2 (fpkm + 1). The cutoff was set at 126.57 FPKM for breast cancer and at 136.7 FPKM for TNBC.
Statistical analysis
All statistical analyses were performed using Statistica version 13.3 (Statsoft) and Microsoft Excel 2019. p value < 0.05 was considered statistically significant. Variables were tested for normality using the Kolmogorov–Smirnov test. The intergroup comparisons used the Mann–Whitney U test or the ANOVA Kruskal–Wallis test. The correlations between MAL2 expression and the clinicopathological features of triple-negative bladder cancer were evaluated using Spearman’s rank correlation coefficient. Univariate and multivariate analyses of potential predictors for overall survival implemented the Cox proportional hazard regression. The optimal cutoffs were calculated using Cutoff Finder [24] Results were expressed as hazard ratio (HR) and 95% confidence interval (CI). The estimation of survival differences between groups was based on the log-rank test.
Results
MAL2 expression is independent of clinical features of TNBC
First, we calculated the expression of MAL2 depending on the clinical features of TNBC (Table 2, Table S1). MAL2 did not correlate with stage, grade, tumor size, and lymph node invasion. Its expression was independent of age, PD-L1 status, Ki-67 status, progression, and recurrence. Although the PD-L1 status and lymph node invasions show a trend toward higher MAL2 expression, those correlations were not statistically significant. There was no correlation between MAL2 expression and received neoadjuvant or adjuvant therapy (p > 0.05).
MAL2 predicts shorter survival in TNBC
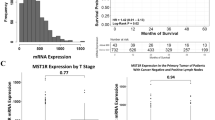
To examine the predictive value of MAL in patients with triple-negative breast cancer, we dichotomized the samples into low and high MAL2 expression groups, with the cutoff point set at 88 H-score. H-score ≤ 88 indicated low MAL2 expression, and H-score > 88 indicated high MAL2 expression. Patients with high MAL2 had significantly lower 5-year survival rates (71.33% vs. 89.59%, respectively; p = 0.0224) than patients with low MAL2 (Fig. 2). The Kaplan–Meier analysis of overall survival by quartiles showed significant differences in OS between patients in the first and second quartiles of MAL2 expression (p = 0.021). MAL2 expression did not predict disease-free survival in patients with TNBC (86.75% vs. 81.59% 5 years after diagnosis; p = 0.41).

Patients’ survival depends on MAL2 status (89.59% vs. 71.33% 5 years after diagnosis in low-MAL2 and high-MAL2 groups, respectively; p = 0.0244)
In univariate Cox regression analysis, lymph node invasions, size, recurrence, and low MAL2 expression (HR 0.29 [CI 95% 0.087–0.95]; p < 0.05) were associated with longer patients’ survival. In multivariate analysis, only disease recurrence and lymph node invasion retained statistical significance (Table 3). Given its significance in univariate analysis, we expand the analysis of MAL2 onto the TCGA cohort.

MAL2 and PD-1 are overexpressed in cancers compared to normal adjacent tissue (p = 0.0001 and p = 0.0007, respectively)
In silico analysis
We assessed the expression of MAL2 and PDCD1 in the TCGA BRCA cohort using the XENA browser (Table 4) [22, 23, 25]. The normalized gene expression for MAL (5.09 vs. 6.4; p < 0.000001) and PDCD1 (0.29 vs. 0.37; p = 0.00049) was higher in tumors than in adjacent normal tissues. MAL2 and PDCD1 are overexpressed in breast cancer (Fig. 3).

Overall survival in the TCGA cohort based on MAL2 expression (p = 0.0001)
We used the Ensembl gene ID from TCGA to map the TCGA RNA-seq data. The samples were dichotomized into low and high MAL2 expression groups based on the FPKMs value (number of fragments per kilobase of exon per Million reads).
Breast cancer patients with low MAL2 had significantly longer overall survival (p = 0.0001) and disease-specific survival (p = 0.0026) than patients with high MAL2 expression (Fig. 4). In multivariate COX regression analysis, older age and high MAL2 expression were the only independent predictors of shorter patient survival in the BRCA TCGA cohort (Supplementary materials).
Next, we analyzed the role of MAL2 in a subset of TNBC patients extracted from the TCGA database. In the TNBC TGCA cohort, patients with high MAL2 expression had significantly higher 5-year survival than patients with low MAL2 expression (87.25% vs. 55.45%, respectively) (Fig. 5). Furthermore, high MAL2 expression, older age, higher stage, and disease progression were independent prognostic factors of shorter TNBC patients survival (p < 0.05) (Table 5). The results obtained from the TCGA cohort are consistent with the findings from our tissue microarray (TMA) cohort.

Overall survival in the TCGA TNBC cohort based on MAL2 expression (p = 0.008)
Discussion
In our study, MAL2 overexpression was associated with shorter survival in triple-negative breast cancer. Although the results from the TCGA database analysis align with the results from our TMA cohort, MAL2 expression seems independent of PD-L1 status, tumor grade, stage, size, lymph node invasion, and metastasis. The lack of association with clinicopathological features of TNBC indicates that MAL2 independently contributes to cancer progression. However, the unclear relationship between MAL2, PD-1, and cancer immune escape needs to be addressed.
T Cell Differentiation Protein 2 (MAL2) is a 176 amino acid residue protein encoded by a gene located on chromosome 8 (8q24.12) [10, 11, 26]. It belongs to the MAL proteolipid family and encodes a transmembrane protein that participates in transcytosis [8, 27]. While the nature of the interactions between MAL2 signaling and breast cancer remains largely unexplored, recent studies have shed new light on its role in cancer immune escape [9]. Cancer cells interact with each other and modulate the adjacent stroma cells to form an immunosuppressive microenvironment (TME) where they dwell and proliferate [28]. In their niche, cells lose epithelial phenotype and acquire mesenchymal-like phenotype. This phenomenon, called epithelial–mesenchymal transition (EMT), is the key event that leads to cancer immune escape [29].
MAL2 is overexpressed in breast cancer, and its high expression correlates with poor prognosis [27]. The Bhandari et al. study showed that MAL2 knockdown decreased breast cancer cell proliferation, migration, and invasion. It was associated with increased E-cadherin and Vimentin levels, indicating that MAL2 drives disease progression by EMT. Those results seem to be confirmed by Yuan et al. study, in which MAL2 deletion in ovarian cancer cells reduced their proliferation, migration, invasion, and EMT [11].
The expression of PD-L1 on tumor cells increases along disease progression due to cancer immune escape, epithelial–mesenchymal transition, or the upregulation of the JAK-STAT, MAPK, and PI3K-AKT signaling pathways [30,31,32]. Contrary to MAL2, TNBC expressing PD-L1 seems to be associated with better outcomes and is more likely to respond to neoadjuvant chemotherapy [33]. While direct interactions between PD-1/PD-L1 and MAL2 are currently unknown, they seem to regulate T cell reactivity and anticancer immune response [9, 34]. By interacting with MHC-I and RAB, MAL2 caused the degradation of MHC-I, promoted tumor antigen endocytosis, and suppressed their presentation. MAL2 depletion in a mice xenograft suppressed breast tumor growth and enhanced tumor-infiltrating CD8 + T cells cytotoxicity, presumably by facilitating tumor antigen presentation [9].
While direct interaction between MAL2 and PD-1/PD-L1 remains unknown, their overexpression appears to reflect different mechanisms of therapeutic resistance. PD-1/PD-L1 affects the activation of T cells, while MAL2 renders them ineffective in recognizing malignant cells [13, 27]. Considering that we found no direct relationship between PD-1 and MAL2 expression, we hypothesize that both mechanisms occur separately and MAL2 overexpression may limit the efficacy of anti-PD-1/anti-PD-L1 therapy.
Conclusion
MAL2 is overexpressed in TNBC, and high MAL2 expression predicts shorter overall survival in triple-negative breast cancer in both the TMA and TCGA cohorts. MAL2 levels are independent of patients’ age, PD-1 status, Ki-67 status, hormonal therapy, progression, and recurrence status. We found no direct correlation between the expression of PD-L1 and MAL2, but both proteins partake in tumor immunoregulation and may be intertwined in mediating tumor immune escape. While targeting MAL2 appears promising, further trials are needed to examine its clinical applicability.
Data availability
The data presented in this study are available on request from the corresponding author. Due to ethical restrictions, they are not publicly available.
References
Breast cancer. [cited 17 May 2023]. Available: https://www.who.int/news-room/fact-sheets/detail/breast-cancer. Accessed 24 Apr 2024.
Triple-negative breast cancer. [cited 3 May 2023]. Available: https://www.cancer.org/cancer/breast-cancer/about/types-of-breast-cancer/triple-negative.html. Accessed 24 Apr 2024.
Irvin WJ Jr, Carey LA. What is triple-negative breast cancer? Eur J Cancer. 2008;44:2799–805. https://doi.org/10.1016/j.ejca.2008.09.034.
Smolarz B, Nowak AZ, Romanowicz H. Breast cancer-epidemiology, classification, pathogenesis and treatment (review of literature). Cancers. 2022. https://doi.org/10.3390/cancers14102569.
Wahba HA, El-Hadaad HA. Current approaches in treatment of triple-negative breast cancer. Cancer Biol Med. 2015;12:106–16. https://doi.org/10.7497/j.issn.2095-3941.2015.0030.
Li Y, Zhan Z, Yin X, Fu S, Deng X. Targeted therapeutic strategies for triple-negative breast cancer. Front Oncol. 2021;11: 731535. https://doi.org/10.3389/fonc.2021.731535.
Zhong Y, Zhuang Z, Mo P, Shang Q, Lin M, Gong J, et al. Overexpression of MAL2 correlates with immune infiltration and poor prognosis in breast cancer. Evid Based Complement Alternat Med. 2021;2021:5557873. https://doi.org/10.1155/2021/5557873.
Dersh D, Yewdell JW. Immune MAL2-practice: breast cancer immunoevasion via MHC class I degradation. J Clin Invest. 2021. https://doi.org/10.1172/jci144344.
Fang Y, Wang L, Wan C, Sun Y, Van der Jeught K, Zhou Z, et al. MAL2 drives immune evasion in breast cancer by suppressing tumor antigen presentation. J Clin Invest. 2021. https://doi.org/10.1172/jci140837.
Byrne JA, Maleki S, Hardy JR, Gloss BS, Murali R, Scurry JP, et al. MAL2 and tumor protein D52 (TPD52) are frequently overexpressed in ovarian carcinoma, but differentially associated with histological subtype and patient outcome. BMC Cancer. 2010;10:497. https://doi.org/10.1186/1471-2407-10-497.
Yuan J, Jiang X, Lan H, Zhang X, Ding T, Yang F, et al. Multi-omics analysis of the therapeutic value of MAL2 based on data mining in human cancers. Front Cell Dev Biol. 2021;9: 736649. https://doi.org/10.3389/fcell.2021.736649.
Schütz F, Stefanovic S, Mayer L, von Au A, Domschke C, Sohn C. PD-1/PD-L1 pathway in breast cancer. Oncol Res Treat. 2017;40:294–7. https://doi.org/10.1159/000464353.
Gou Q, Dong C, Xu H, Khan B, Jin J, Liu Q, et al. PD-L1 degradation pathway and immunotherapy for cancer. Cell Death Dis. 2020;11:955. https://doi.org/10.1038/s41419-020-03140-2.
Mittendorf EA, Philips AV, Meric-Bernstam F, Qiao N, Wu Y, Harrington S, et al. PD-L1 expression in triple-negative breast cancer. Cancer Immunol Res. 2014;2:361–70. https://doi.org/10.1158/2326-6066.cir-13-0127.
Lei Q, Wang D, Sun K, Wang L, Zhang Y. Resistance mechanisms of anti-PD1/PDL1 therapy in solid tumors. Front Cell Dev Biol. 2020;8:672. https://doi.org/10.3389/fcell.2020.00672.
Cortes J, Rugo HS, Cescon DW, Im S-A, Yusof MM, Gallardo C, et al. Pembrolizumab plus chemotherapy in advanced triple-negative breast cancer. N Engl J Med. 2022;387:217–26. https://doi.org/10.1056/nejmoa2202809.
Miles D, Gligorov J, André F, Cameron D, Schneeweiss A, Barrios C, et al. Primary results from IMpassion131, a double-blind, placebo-controlled, randomised phase III trial of first-line paclitaxel with or without atezolizumab for unresectable locally advanced/metastatic triple-negative breast cancer. Ann Oncol. 2021;32:994–1004. https://doi.org/10.1016/j.annonc.2021.05.801.
Ventana Medical Systems, Inc. and Roche Diagnostics International, Inc. Ventana PD-L1 (SP142) assay quick referene guide for ttiple-negative breast carcinoma (TNBC). 2022. Available: https://elabdoc-prod.roche.com/eLD/api/downloads/dba77a2d-da62-ea11-fa90-005056a772fd?countryIsoCode=be. Accessed 24 Apr 2024.
Nicholson RI, Bouzubar N, Walker KJ, McClelland R, Dixon AR, Robertson JF, et al. Hormone sensitivity in breast cancer: influence of heterogeneity of oestrogen receptor expression and cell proliferation. Eur J Cancer. 1991;27:908–13. https://doi.org/10.1016/0277-5379(91)90145-4.
Cerami E, Gao J, Dogrusoz U, Gross BE, Sumer SO, Aksoy BA, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012;2:401–4. https://doi.org/10.1158/2159-8290.cd-12-0095.
Goldman MJ, Craft B, Hastie M, Repečka K, McDade F, Kamath A, et al. Visualizing and interpreting cancer genomics data via the Xena platform. Nat Biotechnol. 2020;38:675–8. https://doi.org/10.1038/s41587-020-0546-8.
Expression of MAL2 in breast cancer - the human protein atlas. [cited 26 Jan 2023]. Available: https://www.proteinatlas.org/ENSG00000147676-MAL2/pathology/breast+cancer. Accessed 24 Apr 2024.
Expression of PDCD1 in breast cancer - the human protein atlas. [cited 26 Jan 2023]. Available: https://www.proteinatlas.org/ENSG00000188389-PDCD1/pathology/breast+cancer. Accessed 24 Apr 2024.
Budczies J, Klauschen F, Sinn BV, Győrffy B, Schmitt WD, Darb-Esfahani S, et al. Cutoff Finder: a comprehensive and straightforward Web application enabling rapid biomarker cutoff optimization. PLoS ONE. 2012;7: e51862. https://doi.org/10.1371/journal.pone.0051862.
UCSC Xena. [cited 30 Apr 2024]. Available: https://xenabrowser.net/datapages/?dataset=TCGA-BRCA.htseq_fpkm.tsv&host=https://gdc.xenahubs.net&removeHub=https://xena.treehouse.gi.ucsc.edu:443. Accessed 24 Apr 2024.
Zheng C, Wang J, Zhang J, Hou S, Zheng Y, Wang Q. Myelin and lymphocyte protein 2 regulates cell proliferation and metastasis through the Notch pathway in prostate adenocarcinoma. Transl Androl Urol. 2021;10:2067–77. https://doi.org/10.21037/tau-21-244.
Bhandari A, Shen Y, Sindan N, Xia E, Gautam B, Lv S, et al. MAL2 promotes proliferation, migration, and invasion through regulating epithelial-mesenchymal transition in breast cancer cell lines. Biochem Biophys Res Commun. 2018;504:434–9. https://doi.org/10.1016/j.bbrc.2018.08.187.
Fan Y, He S. The characteristics of tumor microenvironment in triple negative breast cancer. Cancer Manag Res. 2022;14:1–17. https://doi.org/10.2147/cmar.s316700.
Terry S, Savagner P, Ortiz-Cuaran S, Mahjoubi L, Saintigny P, Thiery J-P, et al. New insights into the role of EMT in tumor immune escape. Mol Oncol. 2017;11:824–46. https://doi.org/10.1002/1878-0261.12093.
Xu D, Li J, Li R, Lan T, Xiao C, Gong P. PD-L1 expression is regulated by NF-κB during EMT signaling in gastric carcinoma. Onco Targets Ther. 2019;12:10099–105. https://doi.org/10.2147/ott.s224053.
Zhang L, Xu L-J, Zhu J, Li J, Xue B-X, Gao J, et al. ATM-JAK-PD-L1 signaling pathway inhibition decreases EMT and metastasis of androgen-independent prostate cancer. Mol Med Rep. 2018;17:7045–54. https://doi.org/10.3892/mmr.2018.8781.
Quan Z, Yang Y, Zheng H, Zhan Y, Luo J, Ning Y, et al. Clinical implications of the interaction between PD-1/PD-L1 and PI3K/AKT/mTOR pathway in progression and treatment of non-small cell lung cancer. J Cancer. 2022;13:3434–43. https://doi.org/10.7150/jca.77619.
Oner G, Önder S, Karatay H, Ak N, Tükenmez M, Müslümanoğlu M, et al. Clinical impact of PD-L1 expression in triple-negative breast cancer patients with residual tumor burden after neoadjuvant chemotherapy. World J Surg Oncol. 2021;19:264. https://doi.org/10.1186/s12957-021-02361-9.
Shi L, Chen S, Yang L, Li Y. The role of PD-1 and PD-L1 in T-cell immune suppression in patients with hematological malignancies. J Hematol Oncol. 2013;6:74. https://doi.org/10.1186/1756-8722-6-74.
Funding
This study received no funding.
Author information
Authors and Affiliations
Contributions
Conceptualization: J.B. and K.S.; writing—original draft preparation: J.B., W.S., K.G., K.Sz.; formal analysis: J.Ł-M., H.A., M.M., J.B.; visualization: M.Z., Ł.S., M.B.; staining evaluation: M.Z., Ł.Sz.; investigation: J.B., M.M.; acquisition: J.Ł-M., H.A.; writing—review and editing: J.B. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest relevant to the article’s content.
Ethical approval
The study was conducted following the Declaration of Helsinki and approved by the Bioethics Committee (KB 746/2021).
Informed consent
The Bioethical Committee waived the requirement for patient consent due to the study’s retrospective nature.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Borowczak, J., Zdrenka, M., Socha, W. et al. High MAL2 expression predicts shorter survival in women with triple-negative breast cancer. Clin Transl Oncol (2024). https://doi.org/10.1007/s12094-024-03514-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12094-024-03514-4