
Abstract
Seleno-Yeasts (SY) used as feed additives are known to contain different Selenium (Se) species. Seleno-Yeasts has been shown, on previous analytical methods, to contain selenomethionine (SeMet), selenocysteine (SeCys), selenate (SeIV) and selenite (SeVI), and various other organic and inorganic Se forms identified but rarely quantified. A new advanced method has allowed elemental Se (Se0), an inorganic Se species, to be quantified, thereby obtaining better insight into the proportion of inorganic Se in SY products. The study aimed to quantify the Se0 in SY products and assess the proportion of inorganic Se in SY. The Se speciation of 13 fresh commercials SY from different suppliers and batches, was assayed for the total Se, inorganic Se species (SeIV, SeVI and Se0), and organic Se species (SeMet and SeCys). Results on total Se were in line with the expected Se concentrations for all evaluated samples. The proportion of Se present as Se0 ranged from 3.6% to 51.8%. The quantity of Se0 in the SY products, added to SeIV and SeVI, indicated an average proportion of inorganic Se of 14.2% for the 13 analyzed SY products. The proportion of Se as SeMet ranged from 19.0% to 71.8%, (average of 55.8%), and a large variability in the SeMet content was observed. The SeCys content was also variable, with an average of 3.8%, relative to the total Se. In conclusion, advances in the analytical characterization have revealed that SY products can have a significantly high proportion of inorganic Se, which could affect the bioavailability of Se from SY supplements and explain their variable and lower bio-efficacy than pure SeMet supplements, such as hydroxy-selenomethionine.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Selenium (Se) is an essential trace element for all animal species. A deficiency in Se is associated with poor health, increased susceptibility to various diseases and decreased productive and reproductive performances of animals [1]. To avoid the consequences of a Se deficiency and to meet the Se requirements, its supplementation in feeds has become a common practice worldwide. Se supplements already exist commercially in inorganic forms, such as sodium selenite (SeIV) and selenate (SeVI), and in organic forms, such as seleno-yeasts (SY) or pure forms of selenomethionine (SeMet or OH-SeMet). In general, the inorganic forms of Se are absorbed less effectively as they have a far lower bio-efficacy than organic forms [2, 3]. When comparing organic Se sources, their bio-efficacy has been attributed to their SeMet proportion, since it is the only Se species that can be stored in tissues and build up a Se reserve in the body [2, 3]. Seleno-yeasts often shows a lower and variable bio-efficacy than pure SeMet [3], due to the variable content of SeMet in SY products. However, there is still limited knowledge about the other Se species present in SY products, and their characterization still requires further investigation. Thus, Se speciation in SY is necessary to obtain a better understanding of its metabolism and biological significance in animal nutrition.
Seleno-yeasts are produced by the aerobic fermentation of yeasts, mainly the S. cerevisiae yeast strain, in a medium that contains sugar beet pulp or cane molasses, vitamins, nutritional salts and sodium selenite as the Se source [4, 5]. Briefly, the enzymatic systems in growing yeast are not able to distinguish between Se and Sulfur (S), due to their similar chemical and physical properties [6]. Therefore, during the fermentation process, Se is used for SeMet synthesis in a similar way to S during the synthesis of methionine. Commercial SY products contain between 1,000 to 3,000 mg of Se/kg, which is composed of SeMet and other Se species that have formed as secondary metabolites during the fermentation process [2, 7]. Seleno-yeasts are known to have a complex composition of Se species [7,8,9]. In fact, over 180 Se compounds have recently been identified in 5 Seleno-yeasts products, and the authors suggested that different SY could vary in efficacy due to differences in the compound composition of Se [10]. According to the European commission, SY should contain at least 97% of organic Se relative to the total Se, and a minimum of 63–70% of Se as SeMet [11, 12]. The proportion of inorganic Se has been indicated to be less than 2% [13,14,15,16]. Likewise, FDA [17] and AAFCO [18] have regulated SY related to animal feed additives; inorganic Se should be < 2% of the total Se. However, commercial SY products are facing several issues, including inconsistencies in SeMet concentrations [8, 19] and unreachable mass balances between quantified Se species and total Se concentration in SY [8, 19]. To date, the proportion of inorganic Se has been characterized by SeIV and SeVI, which often represent less than 2% of the total Se content of the SY products, and the remaining Se species are considered to be organic, including any unaccounted form of Se compounds. Nevertheless, recent advances in analytical techniques have allowed these Se forms to be further characterized. Vacchina et al. [20] developed a new method to quantify elemental Se (Se0) in SY products. Elemental Se is an inorganic form of Se that had previously been unquantified in SY, even though it was suspected of being generated through a fermentation process in ruminants fed selenite [21]. Therefore, the objective of this study has been to quantify Se0 in different commercial SY products used in animal nutrition by using an advanced analytical approach and to assess the contained proportion of organic/inorganic Se on the basis of these analytical advances.
Material and Methods
Seleno-Yeast Samples
Thirteen fresh SY samples (i.e. within expiration date) from different commercial products and batches were obtained from the market. The considered SY samples are from very common strains of Se enriched yeast, including CNCM I-3060, CNCM I-3399, NCYC R397, NCYC R645 and NCYC R646, which are authorized for animal feeds throughout the world. The labeled total Se content of these products ranged between 1,000 and 3,000 mg/kg.
Procedure Analysis
All the samples were sent to the UT2A lab (Ultra-Trace Analyses Aquitaine, Pau, 64,053, France) to establish the total Se content and conduct the speciation analysis. Se-enriched yeast certified reference material (SELM-1, NRCC, Canada) was used for a quality control to determine the total Se and SeMet. All the analysis were performed in duplicate.
Standards
For total Se determination, a multi-elemental stock standard containing 100 μg/mL of Se (MISA-4, CPAchem, Trappes, France) was used. For speciation analysis, 1000 μg Se/mL SeIV and SeVI were purchased from CPAchem (Trappes, France) and stock solutions of SeMet (1000 µg Se/mL) were prepared by dissolving the proper amount of the salt in ultrapure water. Elemental Se standard (1000 µg/mL) was prepared by dissolving Se0 in 1 M sodium sulfite (for details, see Vacchina et al. [20]).
Total Se Determination
The determination of the total Se in the SY samples was performed using the method described by Bierla et al. [22]. Briefly, the samples were mineralized with a mixture of HNO3/H2O2 on a hot plate and then analyzed by means of Inductively Coupled Plasma Atomic Emission Spectrometry (ICP-AES). Quantification of 10 mg/kg could be reached with an intermediate fidelity of 2%.
Determination of Selenomethionine, Selenite and Selenate
The quantification of SeMet, selenite (SeIV) and selenate (SeVI) in the SY samples was performed using high-performance liquid chromatography, coupled with inductively coupled plasma mass spectrometry (HPLC-ICP-MS) after double proteolytic digestion of the yeast, as described by Goenaga-Infante et al. [23] and Bierla et al. [22].
Selenocysteine Determination
Selenocysteine is an unstable molecule that is highly susceptible to oxidation [24]. Therefore, before digestion of the proteins by proteolysis, the selenocysteine was stabilized by means of carbamidomethylation, as described by Dernovics et al. [25] and Bierla et al. [22]. The proteolytic digestion and quantification of CAM-SeCys was then performed as for SeMet.
Elemental Se (Se0) Determination
The analysis of Se0 was conducted using the new advanced method developed by Vacchina et al. [20]. In short, the method consists of extracting Se0 from SY samples using an excess of sodium sulfite to transform insoluble Se0 into soluble selenosulfate. Selenosulfate, which is representative of the Se0 content in SY, was then quantified by means of HPLC-ICP-MS. The analytical figures of merit of the methods have shown recovery levels ranging from 93 to 101% and within and between-run precision below 8%. The quantification limit of Se0, when using this method, is 1 mg/kg.
Results and Discussion
Seleno-Yeast was the first approved source of organic Se to be used as a feed additive for farm animals. Although the speciation of SY has already been performed, its full composition still needs a more detailed elucidation. In this study, the Se speciation of different commercial SY products has been explored using state-of-the-art and new analytical methods.
Quantification of the Total Se in Seleno-Yeasts
The total Se content in the SY samples was found to be in accordance with the labeled values, that is, between 1,000 and 3,000 mg/kg (Table 1). High levels of Se induce toxicity in yeasts, which limits their capacity to ferment and convert selenite into different Se species. This explains why most commercial SY products have a maximum of 3,000 mg of Se/kg [2, 8, 24].
Quantification of Selenite (SeIV) and Selenate (SeVI)
The proportion of oxyanion Se species (SeIV and SeVI) in the 13 analyzed SY samples are illustrated in Fig. 1. The results show that all the tested samples contain only a residual concentration (< 1%) of inorganic Se as SeIV or SeVI. These results are in line with previous study [26] and with values reported by EFSA [13, 27] and FDA [17], thus confirming that the SeIV and SeVI used for the enrichment of a yeast growth medium is used by the yeast cells and transformed into other forms. Furthermore, the remaining SeIV and SeVI may be washed out during the production process prior to drying of the SY [28].

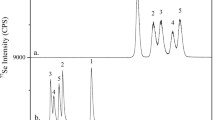
Se speciation compared with the total Se (%) of various fresh commercial Seleno-yeast (SY) products. The concentrations of Selenomethionine (SeMet), Selenocysteine (SeCys), Elemental Se (Se0), Selenite (SeIV), Selenate (SeVI) and other Se species are expressed as the % of the total Se. The analysis was conducted using the HPLC-ICP MS technique for all the Se species and ICP AES for the total Se. Limit of quantification (LOQ) are 10, 0.2, 1, 1, 0.2 and 0.5 mg/kg for total Se, SeMet, SeCys, Se0, SeIV and SeVI, respectively. The proportions of SeIV were: 0.1% for SY-A, 0.2% for SY-B, 0.1% for SY-C, 0.1% for SY-D, < LOQ for SY-E, < LOQ for SY-F, < LOQ for SY-G, 0.1% for SY-H, 0.4% for SY-I, < LOQ for SY-J, 0.1% for SY-K and < LOQ for SY-L and SY-M. The proportions of SeVI were: 0.1% for SY-G and < LOQ for the rest of SY products
Quantification of Elemental Se (Se0) in Seleno-Yeasts (SY)
The results of the quantification of Se0 in the 13 SY samples are presented in Table 1. The analysis revealed the presence of Se0 in all of the SY samples tested in this study, with an average of 14% (standard deviation; SD:13.2%). The proportion of Se0 to total Se ranged between 3.6% and 51.8%. These results corroborate those obtained by Vacchina et al. [20], who found the level of Se0 in 7 SY products ranged between 8 and 40%, with an average of 17.4% (SD:10.5%). These proportions can correspond to partially unknown Se species that have been mentioned in different EFSA reports [14,15,16]. In fact, the proportion of the unknown Se species reported by EFSA varies between 5% [15] and 29% [14], with an average of 17%. According to Jiménez-Lamana et al. [19], the Se mass balance before the quantification of Se0 rarely exceeded 90%, thus suggesting the presence of unaccounted forms of Se.
The Se0 in SY has been hypothesized to result from the reduction of selenite used in the fermentation process [29]. When yeast cells are exposed to high levels of SeIV, they convert it into Se0 in an attempt to reduce its toxicity [30, 31]. It is well known that many microorganisms can reduce highly toxic soluble oxyanion Se (SeIV and SeVI) to a much less toxic insoluble form, Se0 [32]. This transformation was reported to be induced by reductase enzymes [7]. It was observed that the addition of a protein synthesis inhibitor during the culture of yeast cells with SeIV led to the inhibition of the synthesis of Se0 [33]. The authors also studied the possibility of generating Se0 during the fermentation of yeast and found a significant reduction of the transformation of SeIV into Se0 at cold temperatures. They also observed that the process was not influenced by the presence/absence of oxygen and light. Another hypothesis is the transformation of hydrogen selenide, the major intermediate metabolite involved in the synthesis pathway of all forms of Se into Se0 via an oxidation reaction [29, 31, 34].
We also observed a negative correlation between the proportion of Se0 and SeMet in SY, with a coefficient of 0.78 (P < 0.001; Fig. 2). High levels of SeIV can reduce the growth of yeast cells and induce damage, as shown in the study of Rajashree and Muthukumar, [35]. The toxicity of the SeIV in SY was reported to be the result of such different physiological effects as: a genotoxicity effect, mitochondrial alteration and redox imbalance [34]. Thus, high levels of SeIV in the yeast growth media decrease their ability to synthesize SeMet and lead to a reduction of the transformation of SeIV into Se0. This could explain the negative correlation between SeMet and Se0. Hence, the proportion of Se0 in SY could be used as an additional indicator of the quality of SY products.

Correlation between the elemental Se content and selenomethionine content in Seleno-yeast (SY). Y = -0.732 + 0.549 (R2 = 0.781)
With respect to the bioavailability of Se0, it was shown that SeIV and Se0 have a similar bioavailability in different tissues (muscle, liver and plasma) of rats and can cause an equal regulation of plasma selenoprotein P, and it was thus concluded that both forms of Se were equally absorbed, distributed, metabolized and excreted [36]. Moreover, Galbraith et al. [21] reported that rumen microorganisms contributed to the conversion of dietary SeIV into Se0, a non-bioavailable Se form, thus leading to its eventual loss in feces.
Quantification of SeMet in Seleno-Yeasts
The proportions of SeMet in the 13 tested SY are summarized in Fig. 1. These results indicate a SeMet content ranging from 19 to 71.8%, with an average of 55.8% (SD:15.9%). The SeMet incorporation in SY is non-specific and involves the replacement of about 30% of methionine with selenomethionine [6, 37]. In a previous study, 12 SY samples of eight commercial products were collected and analyzed to establish the SeMet content [38], and the authors showed a high variation of SeMet, with an average of 60% total Se. The analysis of 11 samples from different batches of five different producers sourced in the EU and the USA revealed an SeMet content ranging from 24.8% to 69.7% of the total Se [39]. Bierla et al. [22] studied the composition of three SY products and found that the SeMet content varied between 60 and 81%. Rayman reviewed the composition of 7 commercial SY and found a content of 55–75% of Se as SeMet [4]. The variability of SeMet has been reported extensively and attributed to the yeast strain and the fermentation process [5, 9, 28]. Different factors can in fact affect the transformation of Se into SeMet in growing yeast, including the quantity of the sodium selenite used in the medium, the pH, yeast strain, etc. Most of the methodological developments concerning Se analysis have been focused on the quantitative determination of SeMet, because it is the driver of SY bio-efficacy [23]. A SeMet content above a certain value has been accepted as a crucial parameter of the quality control of SY products [6]. EFSA has reported that the SY authorized for animal feeds should contain at least 63%, and in some cases 70%, of Se as SeMet. In the current study, the SeMet proportion was below 63% in 8 of the analyzed SY samples and below 70% in 11 SY samples. Moreover, this proportion represented only one third, or even one-fifth, of the total Se in certain SY. The low proportion of SeMet could be explained by the transformation of oxyanion Se into Se0, rather than into SeMet, during the fermentation process, as mentioned above. In addition, the degradation of SeMet could also explain the low content of SeMet in SY [9]. In fact, the authors detected more SeMet after the addition of the reducing reagent, thus indicating the presence of oxidized forms of SeMet, as previously reported Gammelgaard et al. by [40]. Overall, the results of the SeMet determination in SY products confirm the difficulties of guaranteeing an exact and reliable content of SeMet in commercial SY products, which can explain the variable efficacy of SY when used as feed additives [3, 38].
Quantification of SeCys in Seleno-Yeasts
The results pertaining to the proportion of SeCys in the SY samples are reported in Fig. 1. The total Se in the form of SeCys, varied between 1.2 and 6.6%, with an average of 3.8% (SD: 1.7%). In 2013, 15–30% of total Se as SeCys was estimated in SY, which could represent 1/3 of the total Se [6]. Moreover, EFSA reported that the SeCys in SY was 13–26% of the total Se [15, 16, 27]. However, the findings of the current experiment reveal that the SeCys content in SY is low and thus corroborates the result obtained by Bierla et al. [22], who found an average of 4% of SeCys in 3 SY samples from different producers. The results of the present study are comparable with those of Bierla et al. [22] and those reported by EFSA [41] with 2 – 4% of the total Se being identified as SeCys. The SeCys in SY is found in the water-soluble fraction [42]. Seleno-yeast can incorporate SeCys in the proteins of the yeast proteome, despite the absence of the SeCys insertion sequence (SECIS) in the genome [14]. The SeCys proportion in SY is lower than the SeMet one because cysteine is less abundant than methionine [6, 43, 44], and for those sulfur homologues the degree of substitution of cysteine is lower than that of methionine [24]. Finally, the bio-efficacy of the SeCys form for animals is low and similar to selenite because, unlike SeMet, this Se form does not represent a storage form of Se and cannot be used as it is for selenoprotein synthesis, due to the complex and specific translation machinery, which uses a hydrogen selenide intermediate [3, 45, 46].
The proportion of Inorganic and Organic Se in SY
The proportion of inorganic Se (iSe) represents the sum of the analyzed Se0, SeIV and SeVI, while the proportion of organic (oSe) was computed as the difference between the total Se and iSe. The results showed that the proportion of iSe ranged between 3.8 and 51.8%, with an average of 14.2%, and the proportion of oSe ranged between 48 and 96%, with an average of 85.8% (Table 2). FDA and EFSA currently regulate the SY that has to be used in animal feeds so the iSe content is less than 2% of the total Se (FDA 2022). Organic Se is a generic term that covers different chemical Se species in which the Se atom is chemically bound an integral part of an organic molecule [24]. In fact, to date, only SeIV and SeVI have been considered as iSe species in SY, and the remaining Se metabolites have been presumed to be oSe [47]. This is the result of the assumption that the metabolism of the sodium selenite utilized in the process of fermentation leads to the formation of a bond between Se and carbon, thereby producing several organoselenium compounds [24]. Moreover, different studies performed to characterize the Se species of SY have only found organic Se compounds [10, 48]. However, it should be pointed out that those studies only analyzed the water-soluble fraction of SY, and Se0 was not detected because it is a water-insoluble species. Elemental Se is also an iSe that has already been detected [19, 33, 49, 50], but its accurate quantification in SY has not been possible until recently [20]. The presence of a significant quantity of Se0, and consequently of iSe, helps to explain the bio-efficacy results of SY when used as a feed additive [3]. The quantification of an important amount of iSe in SY has highlighted the need to review the proportion of organic and inorganic Se currently specified by SY suppliers. The increased bio-efficacy of oSe, in comparison to iSe, has been reported for different animal species, including broilers, laying hens, growing pigs, sows as well as dairy and beef cattle [2, 51,52,53,54,55].
Other Se Compounds
Other Se compounds are defined from the difference between the total analyzed Se and the sum of the analyzed Se species, as presented in Fig. 1. The proportion of these undefined Se species is between 12.9 and 48%, with an average of 26.3% (SD: 8.4%). SY has been reported to contain more than 100 unique Se species [56] or even 180 [10]. To date, only SeMet has been clearly described and has been proven to be the driver of the bio-efficacy of SY [3]. The possible roles and effects of these other Se compounds in animals/poultry still require further investigation, although, on the basis of the current knowledge, it can be concluded that, in most cases, their bio-efficacy is not so different from that of sodium selenite [2, 3]. Simon et al. [57], in a test on broiler chickens, compared sodium selenite and two SY with 56.7% and 63% of SeMet, respectively, and concluded that the two SY were not the same and their bioavailability depended on their SeMet content. Van Beirendonck et al. [58] also showed that the Se content in the muscles of broilers was significantly higher in broilers fed SY with 69% of SeMet than broilers fed SY with 26% of SeMet.
Indeed, SeIV, SeVI, and even SeCys are ineffective in increasing the Se concentration in muscle tissue and building Se reserves in monogastric animals, as shown by Deagen et al. [45] for rats and by De Marco et al. [3] for broilers, and only SeMet can non-specifically be incorporated into proteins in place of methionine [4, 8]. The higher bio-efficacy of SeMet has mainly been observed in farm animals’ studies, such us in the muscles of chickens [3, 52, 58], the eggs of laying hens [54, 59, 60], the muscles of growing pigs [53, 61, 62], in sow milk [63] and in milk and beef cattle [55, 64, 65].
The development of these new analytical methods is another step forward in achieving the mass balance of the Se speciation in SY. However, it is still not possible to conclude that the mass balance is complete. It is believed that the remaining Se species are water-soluble metabolites, with a high probability of being oSe, but the presence of other iSe species, in addition to Se0, SeIV and SeVI, cannot be excluded. The aforementioned analytical method may also lead to an underestimation of SeMet and/or the SeCys content, due to a low extraction or their oxidation after extraction during the analysis. The bio-efficacy of such an underestimated fraction should be considered, due to its low bio-accessibility for animals [66]. In addition, significant variation among different SY products was confirmed in this study. In fact, SY production is a complex process based on the growth and multiplication of the yeast and enrichment of the Se content by use of inorganic Sodium Selenite or Selenate. However, as inorganic Se is toxic for living organisms, the incorporation of Se into the yeasts is a delicate process to avoid killing the yeasts. Overall, the speciation of SY samples has revealed the presence of large amounts of Se0, which, consequently, increase the observed proportion of iSe in SY. The analysis has also shown a high variability of SeMet in the SY sources used in animal nutrition.
Conclusion
The recent developments in advanced analytical methodologies allow accurate, reliable and more comprehensive determinations of the Se composition of SY to be made. These analyses have revealed the presence of an inorganic Se species: Se0, which explains a great deal about the previously unknown Se species. These findings have revealed that the SY products used in animal nutrition contain, on average far less than 97% of total Se as organic Se, and consequently, the proportion of inorganic Se in SY products needs to be revised. According to these findings, the characterization of SY as a full organic form of Se is now questionable, and end users and the industry now have the opportunity to make more meaningful choices.
Data Availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Lyons MP, Papazyan TT, Surai P (2007) Selenium in Food Chain and Animal Nutrition: Lessons from Nature -Review-. Asian Australas J Anim Sci 20:1135–1155. https://doi.org/10.5713/ajas.2007.1135
Surai FP, Kochish II, Fisinin IV et al (2018) Selenium in Poultry Nutrition: From Sodium Selenite to Organic Selenium Sources. J Poult Sci 55:79–93. https://doi.org/10.2141/jpsa.0170132
De Marco M, Conjat AS, Briens M et al (2021) Bio-efficacy of organic selenium compounds in broiler chickens. Ital J Anim Sci 20:514–525. https://doi.org/10.1080/1828051X.2021.1894994
Rayman MP (2004) The use of high-selenium yeast to raise selenium status: How does it measure up? Br J Nutr 92:557–573. https://doi.org/10.1079/BJN20041251
Esmaeili S, Khosravi-Darami K (2014) Selenium-Enriched Yeast: As Selenium Source for Nutritional Purpose. Curr Opin Chem Biol 10:49–56. https://doi.org/10.2174/157340131001140328115753
Bierla K, Bianga J, Ouerdane L et al (2013) A comparative study of the Se/S substitution in methionine and cysteine in Se-enriched yeast using an inductively coupled plasma mass spectrometry (ICP MS)-assisted proteomics approach. J Proteomics 87:26–39. https://doi.org/10.1016/j.jprot.2013.05.010
LeBlanc KL, Mester Z (2021) Compilation of selenium metabolite data in selenized yeasts. Metallomics 13. https://doi.org/10.1093/mtomcs/mfab031
Schrauzer GN (2006) Selenium yeast: Composition, quality, analysis, and safety. Pure Appl Chem 78:105–109. https://doi.org/10.1351/pac200678010105
Rao Y, McCooeye M, Windust A et al (2010) Mapping of selenium metabolic pathway in yeast by liquid chromatography-Orbitrap mass spectrometry. Anal Chem 82:8121–8130. https://doi.org/10.1021/ac1011798
Ward P, Chadha M, Connolly C et al (2019) A comparative assessment of water-soluble selenium metabolites in commercial selenised yeast supplements by liquid chromatography-electrospray ionisation QTOF-MS. Int J Mass Spectrom 439:42–52. https://doi.org/10.1016/j.ijms.2018.10.040
Office P (2013) Commission Implementing Regulation (EU) No 427/2013 of 8 May 2013 concerning the authorisation of selenomethionine produced by Saccharomyces cerevisiae NCYC R646 as a feed additive for all animal species and amending Regulations (EC) No 1750/2006, (EC) No 634/2007 and (EC) No 900/2009 as regards the maximum supplementation with selenised yeastText with EEA relevance
Office P (2019) Commission Implementing Regulation (EU) 2019/ 804 - of 17 May 2019 - concerning the renewal of the authorisation of organic form of selenium produced by Saccharomyces cerevisiae CNCM I-3060 and of selenomethionine produced by Saccharomyces cerevisiae NCYC R397 as feed additives for all animal species and repealing Regulations (EC) No 1750 / 2006 and (EC) No 634 / 2007
EFSA (2009) Safety and efficacy of SELSAF (Selenium enriched yeast from Saccharomyces cerevisiae CNCM I-3399) as feed additive for all species. EFSA J 992:1–24. https://doi.org/10.2903/j.efsa.2009.992
EFSA (2012) Scientific Opinion on safety and efficacy of selenium in the form of organic compounds produced by the selenium-enriched yeast Saccharomyces cerevisiae NCYC R646 (Selemax 1000/2000) as feed additive for all species. EFSA J 10:2778. https://doi.org/10.2903/j.efsa.2012.2778
EFSA (2016) Safety and efficacy of selenium-enriched yeast (Saccharomyces cerevisiae NCYC R397) for all animal species. EFSA J 14:4624. https://doi.org/10.2903/j.efsa.2016.4624
EFSA (2017) Safety and efficacy of selenium-enriched yeast (Saccharomyces cerevisiae CNCM I-3399) for all animal species. EFSA J 15:04937. https://doi.org/10.2903/j.efsa.2017.4937
FDA (2022) 21 CFR 573.920 (up to date as of 6–30–2022)
AAFCO (2022) Official publication. Association of American Feed Control Officials
Jiménez-Lamana J, Abad-Álvaro I, Bierla K et al (2018) Detection and characterization of biogenic selenium nanoparticles in selenium-rich yeast by single particle ICPMS. J Anal At Spectrom 33:452–460. https://doi.org/10.1039/C7JA00378A
Vacchina V, Foix D, Menta M et al (2021) Optimization of elemental selenium (Se(0)) determination in yeasts by anion-exchange HPLC-ICP-MS. Anal Bioanal Chem 413:1809–1816. https://doi.org/10.1007/s00216-020-03129-y
Galbraith ML, Vorachek WR, Estill CT et al (2016) Rumen microorganisms decrease bioavailability of inorganic selenium supplements. Biol Trace Elem Res In press 171:338–343. https://doi.org/10.1007/s12011-015-0560-8
Bierla K, Lobinski R, Szpunar J (2018) Determination of proteinaceous selenocysteine in selenized Yeast. Int J Mol Sci 19:543. https://doi.org/10.3390/ijms19020543
Goenaga-Infante H, Sturgeon R, Turner J et al (2008) Total selenium and selenomethionine in pharmaceutical yeast tablets: assessment of the state of the art of measurement capabilities through international intercomparison CCQM-P86. Anal Bioanal Chem 390:629–642. https://doi.org/10.1007/s00216-007-1654-8
Bierla K, Szpunar J, Yiannikouris A et al (2012) Comprehensive speciation of selenium in selenium-rich yeast. TrAC Trends Anal Chem 41:122–132
Dernovics M, Lobinski R (2008) Speciation analysis of selenium metabolites in yeast-based food supplements by ICPMS-assisted hydrophilic interaction HPLC-hybrid linear ion trap/Orbitrap MS(n). Anal Chem 80:3975–3984. https://doi.org/10.1021/ac8002038
Larsen EH, Hansen M, Paulin H et al (2004) Speciation and Bioavailability of Selenium in Yeast-Based Intervention Agents Used in Cancer Chemoprevention Studies. J AOAC Int 87:225–232. https://doi.org/10.1093/jaoac/87.1.225
EFSA (2019) Assessment of the application for renewal of authorisation of selenomethionine produced by Saccharomyces cerevisiae NCYC R397 for all animal species. EFSA J 17:5539. https://doi.org/10.2903/j.efsa.2019.5539
Fagan S, Owens R, Ward P et al (2015) Biochemical Comparison of Commercial Selenium Yeast Preparations. Biol Trace Elem Res:Ahead. https://doi.org/10.1007/s12011-015-0242-6
Kieliszek M, Błażejak S, Gientka I et al (2015) Accumulation and metabolism of selenium by yeast cells. Appl Microbiol Biotechnol 99:5373–5382. https://doi.org/10.1007/s00253-015-6650-x
Nancharaiah YV, Lens PNL (2015) Ecology and biotechnology of selenium-respiring bacteria. Microbiol Mol Biol Rev 79:61–80. https://doi.org/10.1128/MMBR.00037-14
Tarze A, Dauplais M, Grigoras I et al (2007) Extracellular Production of Hydrogen Selenide Accounts for Thiol-assisted Toxicity of Selenite against Saccharomyces cerevisiae*. J Biol Chem 282:8759–8767. https://doi.org/10.1074/jbc.M610078200
Garbisu C, Ishii T, Leighton T et al (1996) Bacterial reduction of selenite to elemental selenium. Chem Geol 132:199–204. https://doi.org/10.1016/S0009-2541(96)00056-3
Pereira AG, Gerolis LGL, Gonçalves LS et al (2018) Selenized Saccharomyces cerevisiae cells are a green dispenser of nanoparticles. Biomed Phys Eng Express 4:35028. https://doi.org/10.1088/2057-1976/aab524
Herrero E, Wellinger RE (2015) Yeast as a model system to study metabolic impact of selenium compounds. Microb Cell 2:139–149. https://doi.org/10.15698/mic2015.05.200
Rajashree K, Muthukumar T (2013) Preparation of Organic Selenium Yeast by Fed-Batch Fermentation. Inte J Food Ferm Tech 3:135. https://doi.org/10.5958/2277-9396.2014.00341.9
Loeschner K, Hadrup N, Hansen M et al (2014) Absorption, distribution, metabolism and excretion of selenium following oral administration of elemental selenium nanoparticles or selenite in rats. Metabolomics 6:330–337. https://doi.org/10.1039/c3mt00309d
McSheehy S, Kelly J, Tessier L et al (2005) Identification of selenomethionine in selenized yeast using two-dimensional liquid chromatography-mass spectrometry based proteomic analysis. Analyst 130:35–37. https://doi.org/10.1039/b414246b
Liu YG, Geraert PA, Briens M (2017) How to compare organic selenium sources? Proc. 28th of Annual Australian Poultry Science Symposium, Sydney, New South Wales, pp 217-221
Perucchietti PLW (2012) Why check selenomethionine levels in selenium yeast? Allaboutfeednet 20(6):12–13
Gammelgaard B, Cornett C, Olsen J et al (2003) Combination of LC-ICP-MS, LC-MS and NMR for investigation of the oxidative degradation of selenomethionine. Talanta 59:1165–1171. https://doi.org/10.1016/S0039-9140(03)00026-2
EFSA (2008) Selenium-enriched yeast as source for selenium added for nutritional purposes in foods for particular nutritional uses and foods (including food supplements) for the general population - Scientific Opinion of the Panel on Food Additives, Flavourings, Processing Aids and Materials in Contact with Food. EFSA 766:1–42. https://doi.org/10.2903/j.efsa.2008.766
EFSA (2018) Assessment of the application for renewal of authorisation of selenomethionine produced by Saccharomyces cerevisiae CNCM I-3060 (selenised yeast inactivated) for all animal species. EFSA J 16:05386. https://doi.org/10.2903/j.efsa.2018.5386
Chiao JS, Peterson WH (1953) Yeasts, Methionine and Cystine Contents. J Agric Food Chem 1:1005–1008. https://doi.org/10.1021/jf60016a006
Maw GA (1963) Sulphur utilization by yeast. Pure Appl Chem 7:655–668. https://doi.org/10.1351/pac196307040655
Deagen JT, Butler JA, Beilstein MA et al (1987) Effects of dietary selenite, selenocystine and selenomethionine on selenocysteine lyase and glutathione peroxidase activities and on selenium levels in rat tissues. J Nutr 117:91–98. https://doi.org/10.1093/jn/117.1.91
Labunskyy VM, Hatfield DL, Gladyshev VN (2014) Selenoproteins: Molecular Pathways and Physiological Roles. Physiol Rev 94:739–777. https://doi.org/10.1152/physrev.00039.2013
Kubachka KM, Hanley T, Mantha M et al (2017) Evaluation of selenium in dietary supplements using elemental speciation. Food Chem 218:313–320. https://doi.org/10.1016/j.foodchem.2016.08.086
Arnaudguilhem C, Bierla K, Ouerdane L et al (2012) Selenium metabolomics in yeast using complementary reversed-phase/hydrophilic ion interaction (HILIC) liquid chromatography-electrospray hybrid quadrupole trap/Orbitrap mass spectrometry. Anal Chim Acta 757:26–38. https://doi.org/10.1016/j.aca.2012.10.029
Gharieb MM, Gadd GM (1998) Evidence for the involvement of vacuolar activity in metal(loid) tolerance: vacuolar-lacking and -defective mutants of Saccharomyces cerevisiae display higher sensitivity to chromate, tellurite and selenite. Biometals 11:101–106. https://doi.org/10.1023/A:1009221810760
Zhang L, Li D, Gao P (2012) Expulsion of selenium/protein nanoparticles through vesicle-like structures by Saccharomyces cerevisiae under microaerophilic environment. World J Microbiol Biotechnol 28:3381–3386. https://doi.org/10.1007/s11274-012-1150-y
Briens M, Mercier Y, Rouffineau F et al (2013) Comparative study of a new organic selenium source v. seleno-yeast and mineral selenium sources on muscle selenium enrichment and selenium digestibility in broiler chickens. Br J Nutr 110:617–624. https://doi.org/10.1017/S0007114512005545
Briens M, Mercier Y, Rouffineau F et al (2014) 2-Hydroxy-4-methylselenobutanoic acid induces additional tissue selenium enrichment in broiler chickens compared with other selenium sources. Poult Sci 93:85–93. https://doi.org/10.3382/ps.2013-03182
Jlali M, Briens M, Rouffineau F et al (2014) Evaluation of the efficacy of 2-hydroxy-4-methylselenobutanoic acid on growth performance and tissue selenium retention in growing pigs. J Anim Sci 92:182–188. https://doi.org/10.2527/jas.2013-6783
Jlali M, Briens M, Rouffineau F et al (2013) Effect of 2-hydroxy-4-methylselenobutanoic acid as a dietary selenium supplement to improve the selenium concentration of table eggs. J Anim Sci 91:1745–1752. https://doi.org/10.2527/jas.2012-5825
Grossi S, Rossi L, Marco M de et al (2021) The effect of different sources of selenium supplementation on the meat quality traits of young Charolaise Bulls during the finishing phase. Antioxidants 10. https://doi.org/10.3390/antiox10040596
Gilbert-López B, Dernovics M, Moreno-González D et al (2017) Detection of over 100 selenium metabolites in selenized yeast by liquid chromatography electrospray time-of-flight mass spectrometry. J Chromatogr B: Anal Technol Biomed Life Sci 1060:84–90. https://doi.org/10.1016/j.jchromb.2017.06.001
Simon E, Ballet N, Francesch M et al (2013) Comparison between the dietary supplementation of sodium selenite and selenium-yeast on meat Se accumulation in broiler. EggMear Symposia 2013
Van Beirendonck S, Driessen B, Rovers M et al (2016) Relation between selenomethionine content in dietary selenium sources and selenium deposition in broiler muscle tissue. The Proceedings of XXV World's Poultry Congress
Delezie E, Rovers M, Van der Aa A et al (2014) Comparing responses to different selenium sources and dosages in laying hens. Poult Sci 93:3083–3090. https://doi.org/10.3382/ps.2014-04301
Słupczyńska M, Jamroz D, Orda J et al (2018) Long-Term Supplementation of Laying Hen Diets with Various Selenium Sources as a Method for the Fortification of Eggs with Selenium. J Chem 2018:1–7. https://doi.org/10.1155/2018/7986591
Falk M, Bernhoft A, Framstad T et al (2018) Effects of dietary sodium selenite and organic selenium sources on immune and inflammatory responses and selenium deposition in growing pigs. J Trace Elem Med Biol 50:527–536. https://doi.org/10.1016/j.jtemb.2018.03.003
Rao Z-X, Tokach MD, Woodworth JC et al (2021) Evaluation of selenium source on nursery pig growth performance, serum and tissue selenium concentrations, and serum antioxidant status. Kansas Agric Exp Station Res Rep 7(11). https://doi.org/10.4148/2378-5977.8188
Li N, Sun Z, Ansari AR et al (2020) Impact of Maternal Selenium Supplementation from Late Gestation and Lactation on Piglet Immune Function. Biol Trace Elem Res 194:159–167. https://doi.org/10.1007/s12011-019-01754-y
Dumont T, Briens M, Cutard T, De MM, Leboeuf L, Pascard J (2018) L'administration alimentaire d'hydroxysélénométhionine peut améliorer le statut en sélénium chez les bovins. Renc Rech Ruminants 24:185
Hachemi MA, Sexton JR, Briens M, Whitehouse NL (2023) Efficacy of feeding hydroxy-selenomethionine on plasma and milk selenium in mid-lactation dairy cows. J Dairy Sci 106:S0022-0302(23)00101-7. https://doi.org/10.3168/jds.2022-22323
Reyes LH, Encinar JR, Marchante-Gayón JM et al (2006) Selenium bioaccessibility assessment in selenized yeast after “in vitro” gastrointestinal digestion using two-dimensional chromatography and mass spectrometry. J Chromatogr A 1110:108–116. https://doi.org/10.1016/j.chroma.2006.01.088
Acknowledgements
We are grateful to the researchers of the UT2A lab, and especially to Dr. Vacchina, for their involvement in conducting these analyses.
Author information
Authors and Affiliations
Contributions
Mohammed A. Hachemi: Methodology, formal analysis, visualization, investigation and writing of the original draft; Denise Cardoso: Investigation and writing of the original draft; Michele De Marco: Conceptualization, methodology, review and editing; Pierre-André Geraert: review and editing; Mickael Briens: Conceptualization, methodology, writing, review, editing and supervision.
Corresponding author
Ethics declarations
Conflict of Interest
All authors are employee of Adisseo France S.A.S. and this work has been financed by the company but conducted to respect a scientific and objective demonstration.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hachemi, M.A., Cardoso, D., De Marco, M. et al. Inorganic and Organic Selenium Speciation of Seleno-Yeasts Used as Feed Additives: New Insights from Elemental Selenium Determination. Biol Trace Elem Res 201, 5839–5847 (2023). https://doi.org/10.1007/s12011-023-03633-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12011-023-03633-z