
Abstract
Mycophenolic acid (MPA) is an occurring antibiotic produced through Penicillium brevicompactum. Its production was achieved by systematic process optimization under submerged fermentation. In shake flask, single-factor test experiments, Box–Behnken design (BBD) experiments, and fermentation strategy were determined, and the MPA yield was reached at 3002 ± 47, 3610 ± 51, and 4748 ± 59 µg/mL, respectively. For fermentation strategy, MPA production was 58.1% higher than that initial fermentation condition without optimization. Then, the optimized medium was further carried out in 5-L stirred fermenter for 180 h; MPA titer was increased from 3712 ± 65 µg/mL to 5786 ± 76 µg/mL, 55.9% higher than that of single-factor optimized medium. The results of this investigation will provide a vital step toward industrial-scale production of MPA.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
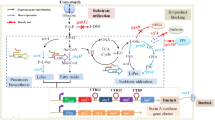
Mycophenolic acid (MPA) is a secondary metabolite of many Penicillium species, including Penicillium brevicompactum, P. paxillin, P. rugulosum, P. canescens, P. roqueforti, P. viridicatum, P. glabrum, and other fungi from genus [1,2,3,4]. Subsequent studies have proved that MPA is not only anti-fungal, anti-tumor, anti-bacterial and anti-psoriasis, but also has an immunosuppression effect and anti-severe acute respiratory syndrome coronavirus 2 activity [5,6,7,8,9]. MPA has a reversible inhibitory impact on hypoxanthine mononucleotide deaminase and selective inhibition of lymphocyte activity [10,11,12]. In addition, mycophenolate mofetil, a 2-malinyl ethyl ester derivative of MPA, is used to prevent rejection after organ and tissue transplantation [13, 14]. At present, a variety of MPA-based immunosuppressants have been commercialized, such as Cellcept and Myfortic [15, 16]. Given such known functions, the production of MPA with high efficiency tends to be important to improve the production level.
The biosynthesis method using Penicillium sp. microbial fermentation to produce MPA was usually divided into two routes: submerged fermentations and solid-state fermentations. With solid fermentation, optimization of fermentation medium for Penicillium brevicompactum increased MPA yield from 3.4 to 4.5 g/kg [1]. Nevertheless, solid-state fermentations are generally not applicable to the commercial production of medicinal products. For submerged fermentation, the yield of MPA was increased from 1.2 to 1.7 g/L using the wild strain of P. brevicompactum (MTCC 8010) with central composite design (CCD) and one-at-a-time variation factors method at the shake flask level [17]. In the submerged fermentation process, the maximum MPA production was reached 3.1 g/L using P. brevicompactum with ultrasonicated batch fermentation at the shake flask level, which is 1.64-fold higher than without ultrasonicated batch fermentation [18]. It was reported that a titer of 6.7 g/L MPA was reached using strain of NRRL864 by increasing the intracellular Ca2+ concentration of MPA high-producing strain NRRL864 and medium optimization at the shake flask level [19]. Various bioreactor and different feeding strategies may play a significant role in the growth of mycelia and biofermentation of MPA. A combined feeding strategy for increasing MPA production in a 7 L fermenter using P. brevicompactum ATCC 16,024, the maximum MPA increased from initial of 1.72 g/L to 2.68 g/L [20]. While using P. brevicompactum MTCC 8010 to raise the production of MPA, it was increased from 1.26 g/L to 3.26 g/L with fed-batch strategy and controlled pH in a 14 L bioreactor [21]. Up to now, research on production MPA mainly deals with screening of high-yield strains producing MPA. High-yield strains can be improved by mutation strategy, such as ultraviolet mutation, atmospheric and room temperature plasma (ARTP), auxotroph screening using random mutagenic method, and insertion mutation [22,23,24]. It was reported that used various antibiotic-resistant mutants for screening strain, mutant strain reached 4.7 g/L of MPA [22]. Agrobacterium tumefaciens-mediated transformation was used to insert a mutated β-Hydroxy-β-methylglutaryl-CoA lyase gene into P. brevicompactum. The maximum MPA concentration by transformants was 2.94 g/L [23]. Knowledge-based approaches in genetic modification and metabolic regulation requires somewhat experience because multiple genes and pathways are involved in overproduction of metabolites. Nevertheless, gene-modified strain or chemical-defined medium is commonly difficult to achieve desirable MPA production. There are limited reports on enhancing production of MPA by systematic process optimization under submerged fermentation. Medium optimization is a vital part of process optimization for MPA production because most of highly MPA production was attributed to the optimal medium.
In this study, attempts were carried out to improve MPA production using strain Penicillium brevicompactum, with combined use of traditional single-factor test method, Box–Behnken design method, and fed-batch fermentation strategy. The fed-batch fermentation was compared with a batch fermentation that used the single-factor method optimized medium. The fed-batch fermentation was further carried out in 5-L stirred fermenter to produce MPA. Hence, the aim of this study was to improve MPA yield through fermentation process optimization in hope to provide a solid basis for the industrialization of fermentation.
Materials and Methods
Organism and Culture
Penicillium brevicompactum was used in this study was obtained from the partner company and kept in our lab. The strains were cultured on the sterile slope of potato glucose agar slants (potatoes 200 g/L, glucose 20 g/L, agar power 18 g/L) at 22 °C for 9 days. The spores were collected from the potato glucose agar slants petri dish and adjusted to (107–108) CFU/mL with distilled water [1]. This spore suspension was used as an inoculum in the shake flask and fermenter experiments.
Preparation of Seed Culture and Fermentation Media
Seed medium was as follows (g/L): sucrose 20, raw soybean flour 5.0, corn steep powder 5.0, KH2PO4 1.0, and MgSO4 1.0. The medium pH was adjusted to 6.5 with 1.5 M NaOH solution prior to sterilization at 121 °C for 20 min. 2.0 mL spore suspension was inoculated into a 500-mL shaking flask with 100-mL seed culture medium and cultivated at 24 °C for 26–27 h in shake incubator at 250 rpm.
Original medium was consisted of (g/L): glucose 150, yeast extract 20, glycine 12.0, methionine 0.4, potassium dihydrogen phosphate (KH2PO4) 3.0, and trace elements solution 1 mL. The trace element solution consisted of (g/L): FeSO4·7H2O 0.22, CuSO4·5H2O 0.3, MnSO4 4H2O 0.12, ZnSO4·7H2O 2.0. The media components expect carbon source and trace element solution, were autoclaved separately at 121 °C for 20 min after the pH had been adjusted to 6.0 with 1.5 M HCl or 1.5 M NaOH. Carbon source and trace element were sterilized at 115 °C, 15 min. The medium was inoculated with an inoculum of 10.0% (v/v).
Optimization of Fermentation Conditions for MPA
The effects of carbon sources (glucose, sucrose, fructose, soluble starch, and glycerol), nitrogen sources (peptone, raw soybean meal, yeast extract, tryptone, and urea), phosphate sources (KH2PO4, K2HPO4, NaH2PO4, and Na2HPO4), and precursor sources (methionine, glycine), temperatures (20 to 30 °C), and initial pH (4.0–7.0) on the production of MPA in 250-mL shake flask each containing 50 mL of the culture medium were studied by single-factor test experiments. 1 mL/L trace elements solution were added to all the medium, and fermentation was conducted at 250 rpm for 180 h. All fermentation experiments were carried out for triplicate.
According to the results of the single-factor test experiments, each factor was evaluated to scrutinize their impacts on the response pattern of MPA yield. Other medium components and fermentation conditions remained same as the single-factor test above. A quadratic polynomial equation was used to predict the response. The experiments were designed and analyzed by the software Design Expert, version 11.0 (State-Ease Inc., USA) [25]. A Box–Behnken experimental design was used to optimize the key factors impacting the production of MPA.
where y is the predicted response, \({x}_{i}\) and \({x}_{j}\) are independent variables, \(k\) is the number of variables, \({\beta }_{o}\) is a constant term, \({\beta }_{i}\), \({\beta }_{ii}\), and \({\beta }_{ij}\) are coefficients for the linear, the coefficients of the quadratic regression, and the coefficients of the interactive regression, respectively, and \(\delta\) is the random error.
Bioreactor Fermentation
According to the shaking flask experiment results, the optimized fermentation strategy medium was further verified in 5-L stirred fermenter (Winpact FS-05 fermenter), and compared with performance of conventionally optimized medium. In a 3.3 L capacity fermenter, the fermentation medium was incubated at 24 °C with a stirring speed of 250 rpm for 180 h, and aeration rate of 1.5 vvm. The fermentation process was tested every 24 h to detect the concentration of amino acid, glucose, and biomass, pH, oxygen capacity, and MPA yield in the fermentation broth.
Analytical Methods
MPA was determined by high-performance liquid chromatography with a C18 (4.6 mm × 250 mm, 5 µm) column at 40 °C. The mobile phase consisted of 0.14% (v/v) triethylamine solution (the pH was adjusted to 5.0 by acetic acid) and acetonitrile at a ratio of 3: 2, and the flow rate was used at 1.0 mL/min. The UV detection was at 249 nm wavelength, and the injection volume was 20 µl. The glucose concentration of fermentation broth was determined by a colorimetric method using the dinitrosalicylic acid (DNS) method [26] on a full-wavelength microplate reader (SpectraMax Plus384, USA) at 540 nm. The experimental results were compared with the standard curve of glucose solution, and the glucose concentration in the fermentation broth was calculated. The determination of the fungal biomass in the fermentation broth was used by the dry cell weight (DCM) method [27]. The amino nitrogen concentration of fermentation broth was determined by formaldehyde titration [28].
Results and Discussion
Screening Experiments by the Single-Factor Test Method
To stepwise increase MPA production, the optimized medium obtained by traditional one-factor-at-a-time method was used to form the control medium for the next screening. As shown in Fig. 1b, 200 g/L of glucose provided with maximum MPA production, reaching at 1966 ± 40 µg/mL. When peptone, tryptone, raw soy flour, and yeast extract been used as nitrogen sources (20.0 g/L for each) for fermentation (Fig. 1c), yeast extract producing MPA was slightly higher than any other one, and the biomass production and MPA concentration were 41.54 ± 0.67 g/L and 2138 ± 52 µg/mL, respectively. As shown in Fig. 1d, 10.0 g/L yeast extract could produce MPA as high as 2682 ± 40 µg/mL. Previous studies reported that the phosphorous source for mycophenolic acid production was KH2PO4 superior to other phosphorous sources [6, 17, 27]. KH2PO4 with a concentration of 3.0 g/L was the preferred phosphorus source (Fig. 1f).

Effects of the following on the production of MPA and biomass. (a) Different carbon sources (150 g/L). (b) Different concentrations of glucose (100–300 g/L). (c) Different nitrogen sources (20 g/L). (d) Different concentrations of yeast exact (1–20 g/L). (e) Different phosphorous sources (3.0 g/L). (f) Different concentrations of KH2PO4 (1–5 g/L)
In the experiment of a single-factor method, the effects of different initial concentrations of methionine and glycine on biomass and MPA yield were studied (Fig. 2). The optimum concentration of glycine was 12.0 g/L (Fig. 2a). When the methionine concentration was 0.4 g/L, the MPA reached at 2987 ± 49 µg/mL (Fig. 2b). The effect of initial pH and temperature on the production of MPA and biomass is shown in Fig. 3. An initial pH of 4.5 a little significantly influences the MPA concentration (Fig. 3a). The maximum MPA production has been reported to vary with respect to the incubation pH. Ultrasound-treated cells of P. brevicompactum were stimulated for MPA production at pH 4.5 [29]. The maximum yield of MPA was produced by P. brevicompactum cultured in a fermentation medium adjusted to pH 5.0 [17]. The maximum MPA production from P. brevicompactum by sorbitol-fed fermentation with the pH controlled at 6.0 [21]. The maximum MPA production by P. brevicompactum with combined feeding strategy and the controlled pH of 6.5 was from 120 h to the end [20]. The yield of MPA had an obvious peak at 24 °C (Fig. 3b).

Effects of different concentrations of glycine and methionine on MPA and biomass production. (a) Glycine (8.0–18.0 g/L) and (b) methionine (0.2–1.0 g/L) on the MPA and biomass

Effects of incubation temperatures and pH on the production of MPA and biomass. (a) Incubation temperatures (20–30 ℃) and pH (4.0–7.0) on the MPA and biomass
According to the single-factor experiment, the highest concentration was 3002 ± 47 µg/mL after continuous fermentation for 180 h. The optimal composition of fermentation medium (g/L): glucose 200, yeast extract 10, methionine 0.4, KH2PO4 3.0, glycine 12.0; trace element solution of 1 mL; the initial pH of 4.5 and an incubation temperature of 24 °C.
Fermentation Medium Optimization Using Box–Behnken Design Method
Based on the above single-factor test experiments, glucose, yeast extract, methionine and glycine were chosen as variables. And the optimal levels of four factors affecting MPA yield were determined by the Box–Behnken design (BBD) method. The glucose (150, 200, 250 g/L), yeast extract (5, 10, 15 g/L), glycine (10, 12.5, 15 g/L), and methionine (0.2, 0.5, 0.8 g/L) were determined for 29 experiments, all of which were performed in triplicate (Table 1). The measured values of MPA produced by BBD fermentation are presented in Table 2. The difference between the experimental results and the simulation results was insignificant. The influence of experimental factors on the response value can be shown by regression fitting and regression equation.
In the formula, A represents glucose, B represents glycine, C represents yeast extract, D represents methionine.
The variance analysis of the regression model is shown in Table 2. The F value of the model was greater than 14 (Table 2), indicating importance of this model. The coefficient (R2) of the model was 0.9442 and adjusted R2 (0.8883), as well as the not significantly lack of fit (P = 0.0702), so a good correlation between actual and predicted MPA yield (Fig. 4). As shown in Table 2, A (glucose) was small than 0.0001, indicating this of the most significant variable to influence MPA yield. It is well known that glucose is vital to MPA fermentation [21]. The production of MPA was significantly affected by the alone, glycine, and yeast extract (P < 0.005). Nonetheless, the linear effect of methionine on MPA yield was not significant (P = 0.975). However, the interaction between yeast extract and methionine meaningfully affected the reaction (Table 2, P < 0.05).

Response surface plot representing interactive effects. (A) Glucose and glycine, (B) glucose and yeast extract, (C) glucose and methionine, (D) glycine and yeast extract, (E) glycine and yeast extract, and (F) yeast extract and methionine on the production of MPA
The smallest ellipse in the contour plot indicates the optimum values of variables at the maximum predicted response [6, 20, 30]. The response surface diagram illustrated the effect of the composition between these four factors on the MPA response. The three-dimensional (3D) response surfaces plots showed the optimal value of MPA yield, and interactions between any two nutrition variables among the four variables are shown in Fig. 4. It was displayed a significant interaction between methionine and yeast extract on MPA production (Fig. 4F). On the contrary, poor ellipticity of the contour lines represented in the plots of glucose vs glycine (Fig. 4A), glucose vs yeast extract (Fig. 4B), glucose vs methionine (Fig. 4C), glycine vs yeast extract (Fig. 4D), and glycine vs methionine (Fig. 4E) suggested a lack of interaction between these factors. The MPA yield of 3606 µg/mL by the model predicted existed as the concentration of glucose, yeast extract, glycine, and methionine were 190.217 g/L, 8.747 g/L, 11.616 g/L, 0.502 g/L, respectively. The MPA yield achieved an experimentally of 3610 ± 51 µg/mL using the optimized conditions, close to the predicted value. The difference between the predicted value of the model and the experimental value was ± 2%. The concentration of MPA in the ordinary optimized medium was 3002 ± 41 µg/mL, which was 20% higher than that of the medium (glucose 200 g/L, glycine 12.0 g/L, methionine 0.4 g/L, yeast extract 10.0 g/L). The final composition of medium optimized was (g/L): glucose 190.217, yeast extract 8.747, glycine 11.616, methionine 0.502, KH2PO4 3.0 and trace element solution 1 mL.
Fermentation Strategy in the Shake Flask
Feeding methionine, an important precursor of MPA, has been reported to enhance MPA production [31]. It was reported that addition of precursors to improve the fermentation yield of antibiotics significantly [32, 33]. Studies have shown that glycine was a potential precursor and can be used as nitrogen source [6, 20]. There have been few reports about the effect of glycine on the production of MPA in the batch fermentation in the shake flask or stirred tank bioreactor. A single-factor test experiment should be carried out in a shaking flask to investigate the effect of different concentrations of glycine on the yield of MPA to obtain better results in the fermenter. The components of the fermentation medium were maintained at the following initial concentrations (g/L): glucose 190.217, KH2PO4 3.0, yeast extraction 8.747, methionine 0.502. The use of trace elements solution was still 1 mL/L. The fermentation conditions were as follows: 180 h at 24 °C and 250 rpm. At 32 h, to study the effect of adding glycine on MPA synthesis, different concentrations of glycine (8.0 g/L, 10.0 g/L, 12.0 g/L, 14.0 g/L, 16.0 g/L) were added at one time. The results in Fig. 5 that the yield of MPA was 4748 ± 59 µg/mL when the feed concentration of glycine was 12.0 g/L and fermented for 180 h, increased by 58% compared with the original fermentation, and effectively improved the fermentation level of MPA.

Effects of different concentrations of glycine on the production of the biomass and MPA. Glycine was added at 32 h fermentation
MPA Fermentation in 5-L Stirred Fermenters
Using the single-factor method, the medium was composed of (g/L) glucose 200, yeast extract 10.0, methionine 0.4, KH2PO4 3.0, glycine 12.0. The composition of a fed-batch fermentation medium was (g/L): glucose 190.217, yeast extract 8.747, methionine 0.502, KH2PO4 3.0. Fed-batch fermentation, at 32 h, 12.0 g/L glycine were added at one time. The above two media were cultured at 24 °C and pH 4.5 for 180 h.
MPA is a typical secondary metabolite during the stationary phase in microorganism sand biosynthesis of MPA occurs during the stationary phase of microorganisms [20]. As shown in Fig. 6, from 0 to 24 h, the glucose and amino nitrogen consumption concentration decreased by 28 g/L and 513 µg/mL in the batch fermentation. In contrast, the consumption of glucose and amino nitrogen was faster than in batch fermentation, and decreased by 33 g/L and 578 µg/mL, respectively. Besides, pH was decreased to 3.81 due to the rapid consumption of glucose. At 32 h fermentation, glycine was added of 12.0 g/L, the pH value was increased from 3.81 to 4.12, and the amino acid nitrogen concentration was increased to 2442 ± 98 µg/mL. After 180 h, the maximum MPA production reached 5786 ± 76 µg/mL, and increased by 55.9% compared to the batch fermentation. It was reported that the yield of MPA was 2.68 g/L (fermentation 312 h in 7 L fermenter), 3.26 g/L (fermentation 312 h in a 14 L stirred bioreactor), and 3.63 g/L (fermentation 280 h in 7 L fermenter), respectively [20, 21, 27]. Through the analysis of reported, MPA production in this study showed the apparently maximum yield, indicating a great potential in industrial process.

Batch fermentation and fed-batch fermentation profile in the stirred fermenter. (a) Batch fermentation profile in stirred fermenter. (b) Fed-batch fermentation profile with feeding of 12.0 g/L glycine at 32 h. [■ pH, ▲ Glucose concentration, ▼ Amino nitrogen concentration, ○ Wet biomass concentration, ● MPA concentration, ♦ Dissolved oxygen]
Conclusion
In this study, fed-batch fermentation was concluded to be superior to batch culture for MPA production using Penicillium brevicompactum. The optimized medium was further verified carried out in a 5-L stirred fermenter for 180 h, MPA titer was increased from 3712 ± 65 µg/mL to 5786 ± 76 µg/mL, a 55.9% increase was achieved through glycine was added at the concentration of 12.0 g/L at 32 h. The fed-batch fermentation method may solve the chemical synthesis of mycophenolic acid with many steps and low yield. We proposed that this combination of response surface methodology and fed-batch fermentation could be efficient for enhancing the production of the submerged fermentation. Therefore, MPA production was greatly improved by optimizing the fermentation strategy, which showed a great potential in application of MPA production.
Data Availability
Not applicable.
References
Patel, G., Patil, M. D., Soni, S., Chisti, Y., & Banerjee, U. C. (2017). Production of Mycophenolic Acid by Penicillium brevicompactum Using Solid State Fermentation. Applied Biochemistry and Biotechnology, 182, 97–109.
Song, X., Tu, R., Mei, X., Wu, S., Lan, B., Zhang, L., Luo, X., Liu, J., & Luo, M. (2020). A mycophenolic acid derivative from the fungus Penicillium sp. SCSIO sof101. Natural Product Research, 34, 1206–1212.
Mahmoudian, F., Sharifirad, A., Yakhchali, B., Ansari, S., & Fatemi, S. S. (2021). Production of Mycophenolic Acid by a Newly Isolated Indigenous Penicillium glabrum. Current Microbiology, 78, 2420–2428.
Mouhamadou, B., Sage, L., Perigon, S., Seguin, V., Bouchart, V., Legendre, P., Caillat, M., Yamouni, H., & Garon, D. (2017). Molecular screening of xerophilic Aspergillus strains producing mycophenolic acid. Fungal Biology, 121, 103–111.
Wang, Y., Zhou, X., Debing, Y., Chen, K., Van Der Laan, L. J., Neyts, J., Janssen, H. L., Metselaar, H. J., Peppelenbosch, M. P., & Pan, Q. (2014). Calcineurin inhibitors stimulate and mycophenolic acid inhibits replication of hepatitis E virus. Gastroenterol, 146, 1775–1783.
Xu, Z., & Yang, S. (2007). Production of mycophenolic acid by Penicillium brevicompactum immobilized in a rotating fibrous-bed bioreactor. Enzyme and Microbial Technology, 40, 623–628.
Kobayashi, A., Yamamoto, I., Nakada, Y., Kidoguchi, S., Matsuo, N., Tanno, Y., Ohkido, I., Tsuboi, N., Yamamoto, H., Yokoyama, K., & Yokoo, T. (2014). Successful treatment of BK virus nephropathy using therapeutic drug monitoring of mycophenolic acid. Nephrology (Carlton, Vic.), 19(Suppl 3), 37–41.
Patel, G., Thakur, N. S., Kushwah, V., Patil, M. D., Nile, S. H., Jain, S., Kai, G., & Banerjee, U. C. (2020). Mycophenolate co-administration with quercetin via lipid-polymer hybrid nanoparticles for enhanced breast cancer management. Nanomedicine, 24, 102147.
Thakur, N. S., Patel, G., Kushwah, V., Jain, S., & Banerjee, U. C. (2019). Self-Assembled Gold Nanoparticle-Lipid Nanocomposites for On-Demand Delivery, Tumor Accumulation, and Combined Photothermal-Photodynamic Therapy. ACS Applied Bio Materials, 2, 349–361.
Ferreira, P. C. L., Thiesen, F. V., Pereira, A. G., Zimmer, A. R., & Fröehlich, P. E. (2020). A short overview on mycophenolic acid pharmacology and pharmacokinetics. Clinical Transplants, 34, e13997.
Anand, S., & Srivastava, P. (2020). Optimization Strategies for Purification of Mycophenolic Acid Produced by Penicillium brevicompactum. Applied Biochemistry and Biotechnology, 191, 867–880.
Neuberger, M., Sommerer, C., Bohnisch, S., Metzendorf, N., Mehrabi, A., Stremmel, W., Gotthardt, D., Zeier, M., Weiss, K. H., & Rupp, C. (2020). Effect of mycophenolic acid on inosine monophosphate dehydrogenase (IMPDH) activity in liver transplant patients. Clinics and Research in Hepatology and Gastroenterology, 44, 543–550.
Resendiz Galvan, J. E., Romano Aguilar, M., Medellin Garibay, S. E., Milan Segovia, R. D. C., Nino Moreno, P. D. C., Jung Cook, H., Chevaile Ramos, J. A., & Romano Moreno, S. (2020). Population pharmacokinetics of mycophenolic acid in adult kidney transplant patients under prednisone and tacrolimus regimen. European Journal of Pharmaceutical Sciences, 150, 105370.
Klangjorhor, J., Chaiyawat, P., Teeyakasem, P., Sirikaew, N., Phanphaisarn, A., Settakorn, J., Lirdprapamongkol, K., Yama, S., Svasti, J., & Pruksakorn, D. (2020). Mycophenolic acid is a drug with the potential to be repurposed for suppressing tumor growth and metastasis in osteosarcoma treatment. International Journal of Cancer, 146, 3397–3409.
Holt, C. D. (2017). Overview of Immunosuppressive Therapy in Solid Organ Transplantation. Anesthesiology Clinics, 35, 365–380.
Takahashi, K., Go, P., Stone, C. H., Safwan, M., Putchakayala, K. G., Kane, W. J., Malinzak, L. E., Kim, D. Y., & Denny, J. E. (2017). Mycophenolate Mofetil and Pulmonary Fibrosis After Kidney Transplantation: A Case Report. American Journal of Case Reports, 18, 399–404.
Patel, G., Patil, M. D., Soni, S., Khobragade, T. P., Chisti, Y., & Banerjee, U. C. (2016). Production of mycophenolic acid by Penicillium brevicompactum-A comparison of two methods of optimization. Biotechnol Rep, 11, 77–85.
Patel, G., Patil, M. D., Tangadpalliwar, S., Nile, S. H., Garg, P., Kai, G., & Banerjee, U. C. (2021). Machine Learning Modeling for Ultrasonication-Mediated Fermentation of Penicillium brevicompactum to Enhance the Release of Mycophenolic Acid. Ultrasound in Medicine and Biology, 47, 777–786.
Chen, M., Wang, J., Lin, L., Xu, X., Wei, W., Shen, Y., & Wei, D. (2022). Synergistic Regulation of Metabolism by Ca(2+)/Reactive Oxygen Species in Penicillium brevicompactum Improves Production of Mycophenolic Acid and Investigation of the Ca(2+) Channel. ACS Synthetic Biology, 11, 273–285.
Dong, Y., Xu, R., Wang, L., Zhang, J., Bai, C., Sun, A., & Wei, D. (2015). A combined feeding strategy for enhancing mycophenolic acid production by fed-batch fermentation in Penicillium brevicompactum. Process Biochemistry, 50, 336–340.
Patel, G., Biswas, K., Patil, M. D., Chisti, Y., & Banerjee, U. C. (2018). Bioreactor studies of production of mycophenolic acid by Penicillium brevicompactum. Biochemical Engineering Journal, 140, 77–84.
Hachiro, O., Masaru, I., Takao, K., Shigeru, Y., & Hiroshiro, S. (1987). Mycophenolic Acid Production by Drug-resistant and Methionine or Glutamic-Acid Requiring Mutants of Penicillium brevicompactum. Agricultural and Biological Chemistry, 51, 2509–2514.
Dong, Y., Zhang, J., Xu, R., Lv, X., Wang, L., Sun, A., & Wei, D. (2016). Insertion Mutation in HMG-CoA Lyase Increases the Production Yield of MPA through Agrobacterium tumefaciens-Mediated Transformation. Journal of Industrial Microbiology and Biotechnology, 26, 1924–1932.
Gillot, G., Jany, J. L., Dominguez-Santos, R., Poirier, E., Debaets, S., Hidalgo, P. I., Ullan, R. V., Coton, E., & Coton, M. (2017). Genetic basis for mycophenolic acid production and strain-dependent production variability in Penicillium roqueforti. Food Microbiology, 62, 239–250.
Adesina, O. A., Taiwo, A. E., Akindele, O., & Igbafe, A. (2021). Process parametric studies for decolouration of dye from local ‘tie and dye’ industrial effluent using Moringa oleifera seed. South African Journal of Chemical Engineering, 37, 23–30.
Miller and G., L. (1959). Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Analytical Biochemistry, 31, 426–428.
Ardestani, F., Fatemi, S. A., Yakhchali, B., Hosseyni, S. M., & Najafpour, G. (2010). Evaluation of mycophenolic acid production by Penicillium brevicompactum MUCL 19011 in batch and continuous submerged cultures. Biochemical Engineering Journal, 50, 99–103.
Adler-Nissen and Jens. (1979). Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. Journal of Agriculture and Food Chemistry, 27, 1256.
Zhao, Y., Ang, W. T., Xing, J., Zhang, J., & Chen, J. (2012). Applications of ultrasound to enhance mycophenolic acid production. Ultrasound in Medicine and Biology, 38, 1582–1588.
Qing, Q., Guo, Q., Wang, P., Qian, H., Gao, X., & Zhang, Y. (2018). Kinetics study of levulinic acid production from corncobs by tin tetrachloride as catalyst. Bioresource Technology, 260, 150–156.
Muth, W. L., & Nash, C. H., 3rd. (1975). Biosynthesis of mycophenolic acid: Purification and characterization of S-adenosyl-L-methionine: Demethylmycophenolic acid O-methyltransferase. Antimicrobial Agents and Chemotherapy, 8, 321–327.
Shiomi, N., Fukuda, H., Murata, K., & Kimura, A. (1995). Improvement of S-adenosylmethionine production by integration of the ethionine-resistance gene into chromosomes of the yeast Saccharomyces cerevisiae. Applied Microbiology and Biotechnology, 42, 730–733.
Vaněk, Z., & Mikulík, K. (1978). Microbial growth and production of antibiotics. Folia Microbiologica, 23, 309.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
Qiaowei Wu contributed to experiment design, execution, and paper writing. Xiangqian Li is the corresponding author and guides the research direction of the project. Mingzhen Li executed the experiment for fermentation by 5-L fermenter. Muhammad Bilal1 contributed to writing—original draft. Yuxiang Yang executed the experiments. Jinfeng Zhang supports experiment design and data analysis. All the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics Approval
Not applicable.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Wu, Q., Li, M., Bilal, M. et al. Enhanced Production of Mycophenolic Acid from Penicillium brevicompactum via Optimized Fermentation Strategy. Appl Biochem Biotechnol 194, 3001–3015 (2022). https://doi.org/10.1007/s12010-022-03886-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-022-03886-9