
Abstract
The diet of entomophagous coccinellids is mainly based on aphids and other food sources such as pollen, nectar, or fungal spores. Knowledge of their foraging behavior on plants infected by powdery mildew and their survival on fungal spores is currently limited. In this study, we investigated the olfactory response of Coccinella septempunctata to odor emission of barley plants infected by powdery mildew and their survival on fungal spores in the presence or absence of aphids. Odors released by powdery-mildew infected plants were more attractive for ladybirds compared to those of uninfected controls. After 3 days, the survival rate of ladybirds feeding only on powdery-mildew spores was less than 50%, while for ladybirds feeding exclusively on Rhopalosiphum padi aphids, the survival rate was close to 90%. After 15 days, the highest survival rate (almost 80%) was observed for ladybirds feeding on plants with both aphids and powdery mildew. Molecular analyses confirmed the presence of fungal spores in ladybird guts when feeding either on powdery mildew or on a mixed diet. Our results provide new insights into foraging behavior of entomophagous coccinellids revealing the potential of powdery mildew to be utilized as important non-essential food in a mixed diet, but also its lethal effect if consumed alone.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Coccinellids are a heterogeneous group of insects divided into three major categories according to their food preferences: zoophagous (predating), phytophagous (plant eating), and mycophagous (fungus eating) (Giorgi et al. 2009). Predatory ladybirds have evolved from mycophagous ladybirds that first were adapted for feeding on sooty molds, but then accepted the insects that produce honeydew such as aphids (Leschen 2000). Most predatory ladybirds feed on honeydew-producing insects from the hemipteran suborder Sternorrhyncha, which they need to complete development. So far, there is no Coccinellid species for which the complete dietary breadth is entirely known (Weber and Lundgren 2009).
Entomophagous coccinellid species regularly consume non-prey food such as pollen and nectar (Togni et al. 2016), honeydew and plant parts (Giorgi et al. 2009), and fruits and fungi (Triltsch 1997). These types of food are an important component of diets for many ladybirds as an additional source of energy and often necessary for the development (Lundgren 2009a). Analyses of gut content have shown that fungal spores can be taken up by, e.g., Coccinella septempunctata, and even be more frequently present than prey body parts, especially at low aphid abundance (Triltsch 1999). Putman (1964) suggested that eating on the fungal spores by C. septempunctata happens fully accidentally, but according to Sutherland and Parrella (2009), fungal spores can be important seasonal food for ladybirds. The importance of the presence of this type of non-prey food in ladybird diet and impact on their foraging behavior are poorly understood.
Entomophagous coccinellids actively utilize olfactory cues associated with the prey odor (Francis et al. 2004), aphid-infested plants (Ninkovic et al. 2001; Togni et al. 2016), or the combination of both (Jamal and Brown 2001). Mycophagous ladybirds showed strong preference to characteristic odors released by squash plants infected by powdery mildew showing that such cues can play important roles in ladybird foraging behavior (Tabata et al. 2011). Non-prey food sources such as fungi are components of entomophagous coccinellid diets, but still we know almost nothing about how they locate infected plants. Detection and location of non-essential food such as fungi could be accidental or mediated by odors associated with fungus-infected plants.
To explore C. septempunctata foraging for powdery mildew, we tested their response to the odors from infected and uninfected barley plants using a two-arm olfactometer. We also investigated if powdery mildew would constitute an appropriate diet for ladybirds and if they eat it as a single food or in combination with Rhopalosiphum padi.
Materials and methods
Powdery mildew
An isolate of powdery mildew, Blumeria graminis f. sp. hordei (Bgh), was obtained from Lantmännen SW Seed AB, Svalöv, Sweden. For propagation of powdery mildew, the highly susceptible barley cultivar (cv.) Steffi was used. Plants were grown in a greenhouse and maintained at 18–22 °C with a light regime of L16:D8. Ten plants per pot (Ø 12 cm, height 9 cm) were grown in potting soil (Special Hasselfors garden, Hasselfors, Sweden). Barley plants infected 7 days earlier with B. graminis were used to inoculate 10-day-old plants (second leaf start to develop) with B. graminis by shaking infested plants above healthy plants. Infected plants were covered with Plexiglas cylinders (Ø 10 cm, height 32.8 cm, thickness 2 mm) closed on the top by a square of felt and a rubber band. Plants were incubated in a climate chamber at 18–22 °C with a light regime of L16:D8 and relative humidity 70% for 5–7 days, until new sporulating lesions of powdery mildew formed on the leaves.
Insects
Seven-spotted ladybird, Coccinella septempunctata, adults were collected from natural habitats close to Uppsala, Sweden (59°47′00.0″N, 17°39′00.0″E) and reared through at least five generations before they were used in experiments. Ladybirds were reared in cages (40 × 40 × 80 cm). They fed on mixed diet consisting of bird cherry-oat aphid (Rhopalosiphum padi), green peach aphid (Myzus persicae), pea aphid (Acyrthosiphon pisum), and pollen of flowering white mustard (Sinapis alba). Aphids were reared on specific host plants; R. padi on barley (Hordeum vulgare), M. persicae on rape seed (Brassica napus), and A. pisum on broad bean (Vicia faba). Ladybirds were reared under the same conditions as the test-plants. The adults in the experiments were 3–4 weeks old. Rhopalosiphum padi used in the experiments were reared on potted oat cv. Belinda under the same growing conditions as infected plants, but in a separate chamber.
Plants infested by aphids and infected by powdery mildew
Barley cv. Annabell was used in the experiments since it had been observed to have a high occurrence of ladybirds when infected with powdery mildew in the field. Ten plants per pot at the two-leaf stage were infested with R. padi (20 aphids per plant). Seven days after infestation, when the number of aphids per plant was around 100, the plants were used for bioassays.
Powdery-mildew infected plants were obtained by infecting barley plants at the two-leaf stage by shaking infected plants above healthy plants. Seven days after inoculation, when lesions were visible, the plants were exploited for experiments.
Plants treated with both aphids and powdery mildew were obtained by releasing 20 aphids per plant on plants that were infected by powdery mildew the day before. Seven days later, the plants were used in bioassays. Each pot with ten plants (treated or control) was placed in a Plexiglas cylinder (Ø 6.8 cm, height 32.8 cm, thickness 2 mm) and the top of the cylinder was sealed with a square of powdery-mildew proof cloth and tightened with a rubber band.
Olfactometer bioassay
Olfactory responses of ladybird adults were measured using a two-way airflow olfactometer consisting of two stimulus zones (arms) directly opposite of each other, with a central neutral zone separating them (Ninkovic et al. 2001). Air was drawn from the center of the olfactometer using a vacuum pump, establishing discrete air currents in the side arms. Airflow in the olfactometer was set to 180 ml/min, measured with a flow meter at the arm inlets. Each arm of the olfactometer was connected to a cage containing the plants. Three different arrangements were set up (1) barley infested by R. padi in one cage versus healthy barley plants in another cage, (2) barley infected by powdery mildew versus healthy barley plants, and (3) barley infected by powdery mildew versus barley infested by R. padi. A single ladybird, randomly chosen without sex determination, was introduced into the olfactometer and after an adaptation period of 10 min, its position was recorded every 2 min over a 20-min period. The number of tested individuals per combination varied from 21 to 23. Ladybirds remaining immobile for longer than 10 min were discarded. Each ladybird was tested only once. After each tested individual, the olfactometer was cleaned using 70% ethanol.
Test of ladybird survival
The effect of diets on ladybird survival was investigated in four different treatments: (1) plants infected with powdery mildew, (2) plants infested with R. padi, (3) plants both infected with powdery mildew and infested with R. padi, and (4) control plants without aphids or powdery mildew. After 7 days of treatment, one ladybird was introduced in each cylinder. Ladybirds had access to water through a cotton wool plug connected to the water in a small container placed near the plant. Every third day, we recorded alive or dead ladybirds during an experimental period of 15 days. Dead ladybirds were collected at each observation and put in − 20 °C. Ladybirds still alive at day 15 were also put in − 20 °C. All ladybirds were kept there until DNA analysis of gut content.
DNA extraction and PCR
Isolated guts were placed in Eppendorf tubes, flash frozen with liquid nitrogen, and homogenized with a sterile pestle. Total DNA was extracted by a Qiacube automated extraction robot (Qiagen) using the QIAamp DNA mini kit (Qiagen). PCR in 25 µl reactions was performed with Illustra PuReTaq Ready-To-Go PCR Beads (GE Healthcare, Uppsala, Sweden) according to Chen et al. (2015) using primers for B. graminis BF-F1 (5′-AAGCTATGCGGAACTTCGTTT-3′) and BF-R (5′-TTAGGAGTTTTGGCAAGTCCC-3′). The PCR program was as follows: 94 °C for 5 min, followed by 35 cycles of [94 °C for 50 s, 68 °C for 50 s, 72 °C for 50 s] followed by a final extension step at 72 °C for 10 min. Amplification products were sequenced at Macrogen (South Korea). Isolated guts of 17 ladybirds were individually DNA-extracted and analyzed with PCR for the presence of B. graminis.
Statistical analyses
The number of visits to one or the other arm of the olfactometer was compared using Wilcoxon matched pairs tests in the SAS statistical package Dell Statistica software (2015). The survival data were analyzed using the non-parametric survival analysis approach proposed by Kaplan and Meier (1958). The Life test procedure of the SAS (2014) package was used. The conclusions were based on the Log-rank test. Pair-wise comparisons were adjusted for multiplicity using the Bonferroni method. Tests comparing the survival at each time point were performed as exact Chi-square tests, i.e., Chi-square tests where the p value was calculated using a randomization test approach, similar to Fisher’s exact test.
Results
Ladybird olfactory responses
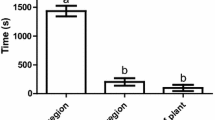
Odors released from stressed plants (powdery-mildew infected or aphid infested) influenced the olfactory responses of C. septempunctata (Fig. 1). The odors of powdery-mildew infected plants evoked a positive behavioral response by ladybirds compared to the odor of uninfected controls (Wilcoxon test: Z = 2.663, P = 0.0077, n = 21; Fig. 1a). The odor of R. padi infested plants also evoked a positive behavioral response by ladybirds compared to the odors released from non-infested plants (Wilcoxon test: Z = 2.84, P = 0.0045, n = 23; Fig. 1b). There was no difference in ladybird olfactory responses when they had a choice between the odor of powdery-mildew infected and aphid-infested barley plants (Wilcoxon test: Z = 0.243, P = 0.807, n = 22; Fig. 1c).

Olfactory responses of ladybirds, Coccinella septempunctata, to different odor sources: a powdery-mildew infected plants and undamaged plants as control, b aphid-infected plants and undamaged plants as control, and c powdery-mildew infected plants and aphid-infested plants. Box and whisker plots represent the cumulative number of visits in the arm zone after ten observations. Each box includes the middle 50% of the data; the dot symbol denotes the median value. The whiskers (vertical lines) indicate a minimum or maximum number of visits of a tested insect; n number of tested insects, P probability calculated by Wilcoxon matched pairs test
Ladybird survival
The overall test comparing all four treatments showed that the ladybird survival rate was strongly dependent on the type of diets (P = 0.0301) (Fig. 2). Pair-wise comparisons of effects of diet showed that the survival rate was significantly higher for ladybirds with mixed diet compared to control (P = 0.04) and compared to powdery mildew (P = 0.0069). After Bonferroni adjustment, only the comparison between mixed diet and powdery mildew remains significant (P = 0.0288). Already after 3 days, the survival rate of ladybirds kept on powdery mildew alone was reduced to 44% which was significant compared to ladybirds provided with a mixed diet (100%; P = 0.0085) or aphids (89%; P = 0.045), but not to the starvation control (78%; P = 0.1469). These results clearly demonstrate that feeding only on powdery mildew strongly reduced the survival rate of the ladybirds. The next considerable reduction of the survival rate was observed on day 9 when only 22% of the control ladybirds were still alive compared to 78% still alive on the mixed diet (P = 0.001). Until day 6, the survival rate for ladybirds kept on the starvation control was at the same level as those kept on the mixed diet or on the aphids alone, but from day 9 and until the end of the experiment the survival rate on the starvation control was at the same level as on the powdery-mildew diet. The mixed diet consisting of R. padi and powdery mildew resulted in the highest survival rate, 78% at day 15, while 56% of the ladybirds reared on R. padi alone were alive at day 15 (Fig. 2).

Survival rate of ladybirds reared on four different diets; only barley, powdery-mildew infected barley, aphid-infested barley, and barley with aphids and powdery mildew. The number of tested ladybirds per diet was 9. Different letters indicate significantly different values (Chi-Square test; P < 0.05)
Detection of powdery mildew in ladybird guts
PCR was used to detect the presence of powdery mildew in ladybird guts after different diets. All ladybirds feeding exclusively on powdery mildew had PCR products of the size corresponding to B. graminis. Also, one of the five starvation controls indicated the presence of powdery mildew. All of these PCR products were sent for sequencing confirming that they belong to powdery mildew. In contrast, no PCR products were obtained for ladybirds fed exclusively on aphids while only one weak PCR product was observed for a ladybird that had the access to both powdery mildew and aphids (Table 1).
Discussion
An observation of a high number of C. septempunctata ladybirds in barley fields infected by powdery mildew, but with extremely low aphid abundance, prompted us to investigate the mechanism of the high ladybird abundance implying the possibility of their survival on powdery mildew. In this study, we have shown that the odor of plants infected by powdery mildew, or the odor of aphid-infested plants, have strong attracting effects on walking behavior of ladybirds compared to the odor of unaffected plants (Fig. 1a). No differences in ladybird attraction between barley plants infected by powdery mildew and aphid-infested plants were observed (Fig. 1c). These results suggest that the odors of attacked/damaged plants can provide important signals for ladybirds in their search for essential and non-essential food. The highest survival rate of ladybirds was observed on plants with aphids and powdery mildew while the lowest survival rate was found for ladybirds living on plants infected with powdery mildew alone, indicating that a mixed diet consisting of non-essential and essential food is beneficial. Powdery-mildew infection can thus be an additional stimulus for predatory ladybirds in the search of more diversified food, but alone it is not an appropriate diet for them.
Finding appropriate food for predatory insects is partially guided by emission of plant volatiles induced by insect feeding (Ninkovic et al. 2001; de Vos and Jander 2010) making attacked plants attractive for natural enemies (Gencer et al. 2017), a phenomenon described as “cry for help” (Dicke and Baldwin 2010). Different plant stresses can induce changes in chemical composition of volatiles (Li et al. 2013; Ninkovic et al. 2011, 2013). For instance, the odor of barley plants infested by R. padi make them attractive for C. septempunctata (Ninkovic et al. 2001). In this study, we show that the odor of plants infected by powdery mildew can have significantly stronger attracting/arresting effects on C. septempunctata than the odor of undamaged plants, almost at the same level as the odor of aphid-infested plants. For mycophagous ladybirds it is known that they are attracted by “moldy” odorants reported as fungal volatiles, but not by elevated quantities of several compounds also present in the volatiles of healthy plants (Tabata et al. 2011). Interestingly, ladybirds did not prefer the odor of aphid-attacked plants over the odor of plants infected by powdery mildew, suggesting that either the different types of plant stresses induce similar compounds and/or that the ladybirds are attracted to both kinds of damages. For either of these options, the data suggest that the odors induced can be important signals for ladybirds in their food-searching behavior.
Powdery mildew developed on the leaf surface has been considered to be consumed accidentally by ladybirds (Sutherland and Parrella 2009). Instead, our molecular analysis of gut content of ladybirds that had only access to powdery mildew showed that they are able to consume it actively. However, their survival was significantly affected; after only 3 days almost 56% were dead and after 15 days the mortality of all individuals reached 78%. Structural polysaccharides found in fungi such as cellulose and lignin are not digestible for many insects and predatory insects lack digestive adaptations which would enable them to exploit the maximum nutrition from fungi (Lundgren 2009b). From our results, it thus seems that feeding on powdery mildew alone can have a lethal effect. This is also supported by the fact that these ladybirds experienced high mortality early on (56% dead at day 3), whereas ladybirds residing on control barley plants (and thus considered starving) started to die much later (80% still alive at day 6). High intake of less appropriate food may have a toxic effect and may potentially cause their death. How predatory ladybirds cope with toxin burden may depend on their energy status as well as their ability to withstand toxins. Well-fed predators may be able to invest more in detoxification (Sherratt et al. 2004). In order to improve its energy status, ladybirds may consume non-preferable food to get balanced range of important nutrients, resulting in a better survival rate than only on a non-preferable or preferable food source (Fig. 2), as is assumed by the model of self-selection of optimal diets (Waldbauer and Friedman 1991). The fungi in mixed diet, as shown in previous studies (Triltsch 1997, 1999; Sutherland and Parrella 2009), can thus be important seasonal food for ladybirds.
Taken together, data from this study and observations made in the field, show that odors released from stressed plants can attract ladybirds to habitats with the presence of both non-essential and essential food. Such foraging behavior suggests that ladybirds choose a feeding site with diversified food that may increase their survival rate via a balanced diet.
References
Chen S, Cao YY, Li TY, Wu XX (2015) Simultaneous detection of three wheat pathogenic fungal species by multiplex PCR. Phytoparasitica 43:449–460. https://doi.org/10.1007/s12600-014-0442-1
de Vos M, Jander G (2010) Volatile communication in plant–aphid interactions. Curr Opin Plant Biol 13:366–371. https://doi.org/10.1016/j.pbi.2010.05.001
Dell Inc (2015) Dell Statistica, version 13.dell.com
Dicke M, Baldwin IT (2010) The evolutionary context for herbivore-induced plant volatiles: beyond the ‘cry for help’. Trends Plant Sci 15:167–175. https://doi.org/10.1016/j.tplants.2009.12.002
Francis F, Lognay G, Haubruge E (2004) Olfactory responses to aphid and host plant volatile releases: (E)-β-Farnesene an effective kairomone for the predator Adalia bipunctata. J Chem Ecol 30:741–755. https://doi.org/10.1023/B:JOEC.0000028429.13413.a2
Gencer NS, Kumral NA, Seidi M, Pehlevan B (2017) Attraction responses of ladybird beetle Hippodamia variegata (Goeze, 1777) (Coleoptera: Coccinellidae) to single and binary mixture of synthetic herbivore-induced plant volatiles in laboratory tests. Turk J Entomol 41:17–26. https://doi.org/10.16970/ted.05956
Giorgi JA, Vanderberg NV, McHugh JV, Forrester JA, Slipinski SA, Miller KB, Shapiro LR, Whiting MF (2009) The evolution of food preferences in Coccinellidae. Biol Control 52:215–231. https://doi.org/10.1016/j.biocontrol.2009.05.019
Jamal E, Brown GC (2001) Orientation of Hippodamia convergens (Coleoptera: Coccinellidae) larvae to volatile chemicals associated with Myzus nicotianae (Homoptera: Aphidiidae). Environ Entomol 30:1012–1016. https://doi.org/10.1603/0046-225X-30.6.1012
Kaplan EL, Meier P (1958) Nonparametric estimation from incomplete observations. J Am Stat Assoc 53:457–481. https://doi.org/10.2307/2281868
Leschen RAB (2000) Beetles feeding on bugs (Coleoptera, Hemiptera). Repeated shifts from mycophagous ancestors. Invertebr Taxon 14:917–929. https://doi.org/10.1071/IT00025
Li YY, Zhou XR, Pang BP, Han HB, Yan F (2013) Behavioral responses of Hippodamia variegata (Coleoptera: Coccinellidae) to volatiles from plants infested by Aphis gossypii (Hemiptera: Aphidae) and analysis of volatile components. Acta Entomol Sin 56:153–160
Lundgren JG (2009a) Nutritional aspects of non-prey foods in the life histories of predaceous Coccinellidae. Biol Control 51:294–305. https://doi.org/10.1016/j.biocontrol.2009.05.016
Lundgren JG (2009b) Relationship of natural enemies and non-prey foods. Springer, Dordrecht. https://doi.org/10.1007/978-1-4020-9235-0
Ninkovic V, Al Abassi S, Pettersson J (2001) The influence of aphid-induced plant volatiles on ladybird beetle searching behavior. Biol Control 21:191–195. https://doi.org/10.1006/bcon.2001.0935
Ninkovic V, Al Abassi S, Ahmed E, Glinwood R, Pettersson J (2011) Effect of within species plant genotype mixing on habitat preference of a polyphagous insect predator. Oecologia 166:391–400. https://doi.org/10.1007/s00442-010-1839-2
Ninkovic V, Dahlin I, Vucetic A, Petrovic-Obradovic O, Glinwood R, Webster B (2013) Volatile exchange between undamaged plants—a new mechanism affecting insect orientation in intercropping. PLoS ONE 8:e69431. https://doi.org/10.1371/journal.pone.0069431
Putman WL (1964) Occurrence and food of some coccinellids (Coleoptera) in Ontario peach orchards. Can Entomol 96:1149–1155. https://doi.org/10.4039/Ent961149-9
SAS (2014) Package SAS Institute Inc. SAS/Stat User’s Guide. Version 9.4. SAS Institute Inc, Cary
Sherratt TN, Speed MP, Ruxton GD (2004) Natural selection on unpalatable species imposed by stat-dependent foraging behaviour. J Theor Biol 228:217–226. https://doi.org/10.1016/j.jtbi.2003.12.009
Sutherland AM, Parrella MP (2009) Mycophagy in Coccinellidae: review and synthesis. Biol Control 51:284–293. https://doi.org/10.1016/j.biocontrol.2009.05.012
Tabata J, De Moraes CM, Mescher MC (2011) Olfactory cues from plants infected by powdery mildew guide foraging by a mycophagous ladybird beetle. PLoS ONE 6:e23799. https://doi.org/10.1371/journal.pone.0023799
Togni PHB, Venzon M, Muniz CA, Martins EF, Pallini A, Sujii ER (2016) Mechanisms underlying the innate attraction of an aphidophagous coccinellid to coriander plants: implications for conservation biological control. Biol Control 92:77–84. https://doi.org/10.1016/j.biocontrol.2015.10.002
Triltsch H (1997) Gut contents in field sampled adults of Coccinella septempunctata (Col. Coccinellidae). Entomophaga 42:125–131. https://doi.org/10.1007/BF02769889
Triltsch H (1999) Food remains in the guts of Coccinella septempunctata (Coleoptera: Coccinellidae) adults and larvae. Eur J Entomol 96:355–364
Waldbauer GP, Friedman S (1991) Self-selection of optimal diets by insects. Annu Rev Entomol 36:43–63
Weber DC, Lundgren JG (2009) Assessing the trophic ecology of the Coccinellidae: their roles as predators and as prey. Biol Control 51:199–214. https://doi.org/10.1016/j.biocontrol.2009.05.013
Acknowledgements
This study is dedicated to the late Professor Jan Pettersson, for his kindness, endless scientific support, and his selflessness that always will be remembered. The study was financially supported by the Swedish Research Council for Environment (FORMAS) (Project Number 2014-225) and by the Ministry of Education, Science and Technological Development of the Republic of Serbia (Project Number III 46008) and by the Carl Tryggers Stiftelse för Vetenskaplig Forskning (Project Number 12:333). We gratefully acknowledge Professor Ulf Olsson for statistical support and Dr. Dimitrije Markovic for valuable comments.
Author information
Authors and Affiliations
Contributions
AR and VN conceived and designed the study; AR performed experiments; VN performed statistical analysis of the data; AR and VN drafted the manuscript; OT performed DNA analyses; VN and OT finalized the manuscript. All authors read and approved the paper.
Corresponding author
Additional information
Handling Editor: Heikki Hokkanen.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Radonjic, A., Terenius, O. & Ninkovic, V. The phytopathogen powdery mildew affects food-searching behavior and survival of Coccinella septempunctata. Arthropod-Plant Interactions 12, 685–690 (2018). https://doi.org/10.1007/s11829-018-9617-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11829-018-9617-x