
Abstract
Members of the aldo–keto reductase family including aldose reductases are involved in antioxidant defense by metabolizing a wide range of lipid peroxidation-derived cytotoxic compounds. Therefore, we produced transgenic wheat genotypes over-expressing the cDNA of alfalfa aldose reductase gene. These plants consequently exhibit 1.5–4.3 times higher detoxification activity for the aldehyde substrate. Permanent drought stress was generated in the greenhouse by growing wheat plants in soil with 20 % water capacity. The control and stressed plants were monitored by a semi automatic phenotyping platform providing computer-controlled watering, digital and thermal imaging. Calculation of biomass values was based on the correlation (R 2 = 0.7556) between fresh weight and green pixel-based shoot surface area. The green biomass production by plants of the three transgenic lines was 12–26–41 % higher than the non-transgenic plants’ grown under water limitation. Thermal imaging of stressed non-transgenic plants indicated an elevation in the leaf temperature. The thermal status of transformants was similar at both normal and suboptimal water regime. In drought, the transgenic plants used more water during the growing season. The described phenotyping platform provided a comprehensive data set demonstrating the improved physiological condition of the drought stressed transgenic wheat plants in the vegetative growth phase. In soil with reduced water capacity two transgenic genotypes showed higher seed weight per plant than the control non-transgenic one. Limitation of greenhouse-based phenotyping in analysis of yield potential is discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
As a consequence of global warming the increase in the frequency of drought episodes threatens the sustainability of the world’s crop production. Therefore, there is an urgent need to develop adaptive agricultural strategies to reduce the devastating economic and social impacts. In these attempts, plant breeding has a pivotal role by continuous supply genotypes with improved water use efficiency and other yield- or drought tolerance-related traits. Nowadays, breeding of crops for stress tolerance and yield stability is efficiently supported by identification of quantitative trait loci (QTLs), association mapping to correlate genetic markers with phenotypes, cloning of resistance genes used for production of transgenic plants (Tuberosa and Salvi 2006; Takeda and Matsuoka 2008; Zhao et al. 2008, Deikman et al. 2012).
Bhatnagar-Mathur et al. (2008) reviewed a large set of isolated genes that has been tested in transgenic plants to analyze drought responses. These genes encode proteins involved in the perception and transduction of stress signals, activation of stress genes (Hsieh et al. 2002; Pellegrineschi et al. 2004; Karaba et al. 2007; Xu et al. 2007). The enhanced synthesis of osmoprotectants such as amino acids (proline), quaternary amines (glycinebetaine, polyamines) sugars and sugar alcohols (trehalose, mannitol) has been achieved and resulted in higher stress tolerance of transgenic plants (Abebe et al. 2003; Capell et al. 2004; Valliyodan and Nguyen 2006). Among other approaches, the over-expression of late embryogenesis abundant (LEA) proteins could provide an increased tolerance to dehydration (Rohila et al. 2002). Publications report on the improvements in grain yield of maize under water-limited conditions in the field. Expression of maize CAAT box transcription factor (ZmNF-YB2) or bacterial RNA chaperones (CSPs) opens a way towards engineering crop plants for yield stability under drought stress (Nelson et al. 2007; Castiglioni et al. 2008).
The decrease in plant productivity under suboptimal growing conditions such as drought largely originates from structural or physiological damages caused by the formation of reactive oxygen species (ROS) and their products from reactions with cellular constituents including lipids (Smirnoff 1993; Moran et al. 1994). Detoxification of lipid peroxidation-derived cytotoxic aldehydes (e.g., 4-hydroxynon-2-enal, HNE) can be a basic mechanism in reduction of stress-generated damages and maintaining physiological functions such as cell division, growth or photosynthesis. As other organisms, plants have a family of aldo–keto reductases (AKRs) that catalyze the NADPH-dependent reduction of different carbonyl metabolites (Bartels 2001). Previously we have shown that ectopic synthesis of the alfalfa aldose reductase (MsALR) enzyme in tobacco plants can prevent the increase of drought-induced formation of lipid peroxidation products (TBARs), OH· radical and H2O2; furthermore, this function has improved the photosynthetic activity (Oberschall et al. 2000; Hideg et al. 2003).
The development of phenomics as an independent discipline opens new dimensions for plant sciences by the acquisition and the interpretation of high-dimensional phenomic data from different levels of organization such as morphology, physiological state and molecular scale (Houle et al. 2010; Fabre et al. 2011, Salekdeh et al. 2009). Growth retardation is a direct stress symptom that can be monitored by phenotyping platforms based on the non-invasive imaging methods without physical damaging of analyzed plants. Green biomass is an integrative phenotypic character depending on both morphological traits such as plant height, leaf and head size and physiological parameters such as transpiration and photosynthesis. The biomass prediction can be based on RGB images taken through red, green, and blue filters to determine the pixel-based shoot surface area. Image analysis is one of the basic components of plant phenotyping (Golzarian et al. 2011; Hartmann et al. 2011). Monitoring the leaf growth rate can provide quantitative data about the functionality of stressed plants (Chenu et al. 2008). Various information systems have been developed for image analysis of digital RGB pictures or storage of environmental and plant watering data (Hartmann et al. 2011; Fabre et al. 2011). Stomatal closure is a key mechanism in plant response to drought stress and without transpirational cooling the canopy temperature is expected to increase relative to air temperature. Therefore, phenotyping platforms are generally equipped with thermal imaging facilities (Merlot et al. 2002; Munns et al. 2010; Berger et al. 2010).
In the present paper, we have quantified the green biomass of wheat plants on the basis of the green pixel-based shoot surface area recorded by digital photography. Based on the thermal imaging data plants from the transgenic (AKR 284, AKR 302, AKR 322) genotypes could maintain the transpiration function. Subsequently in comparison to non-transgenic genotype these plants used more water and produced higher biomass at suboptimal soil water content. The described data extend the previous observations to cereal crops such as wheat that the increase in the protective aldose reductase enzyme activity could ensure improvement in physiological functions under the reduced water supply in vegetative growth phase. We have identified transgenic wheat lines with a potential for better adaptation capability that resulted in higher seed weight per plants. The presented data support the future use of the described transgenic genotypes in studies to uncover the molecular events in the aldo–keto reductase-based stress tolerance.
Materials and methods
Wheat transformation
Immature embryos of the ‘CY-45’ spring wheat (Triticum aestivum L.) cultivar previously selected by Felföldi and Purnhauser (1992) for high morphogenic potential in somatic tissue cultures were used in the present gene transfer experiments. Protocols of biolistic transformation, selection and regeneration of transformants were described by Áy et al. (2012). We bombarded the pAHALR construct (see Supplementary Fig. 1) that was based on the pAHC25 vector (Christensen and Quail 1996). The β-glucuronidase (GUS) reporter gene was removed by SmaI-SacI enzyme digestion and similarly digested fragment of the Medicago sativa MsALR cDNA (Acc. number: X97606; Oberschall et al. 2000) was inserted between these sites. This vector contains two copies of promoters as the 5′-untranslated exons and the introns of the maize ubiquitin (Ubi-I) gene and the terminator sequence from the nopaline synthase (NOS) gene. These regulators insure the expression of the M. sativa MsALR (aldose reductase) and the BAR or PAT gene encoding phosphinothricin acetyltransferase from Streptomyces hygroscopicus. The pUC8 part of the vector contains the TEM-1 β-lactamase gene (bla) of E. coli. The presence of the transformed resistance gene (BAR) was tested by 0.5 % Finale 14 SL® spraying and gene-specific PCR amplification. Wheat plants of the AKR transgenic lines were consequently self-pollinated and all of the generations (T2–T3) were treated with the herbicide (Finale 14 SL®). From the Hungarian Gene Technology Authority the laboratories of the Biological Research Centre and the Cereal Research Non-profit Ltd. have permission for the closed system experiments (35476/2009). In total, 1,797 immature embryos were bombarded, and regeneration of 127 plantlets was achieved after applying the selective concentration of the herbicide.
Plant cultivation and stress treatment
In the present study seeds of plants from the transgenic wheat genotypes (AKR 284; AKR 304, AKR 322) were sown and after germination at two-leaf-stage seedlings were treated with 0.5 % Finale 14 SL® (effective molecule: 14 % glufosinate ammonium) to ensure the use of transgenic plants. We exposed the survived plants to cold-treatment at 3–4 °C for 1–2 weeks to synchronize their development. During this growing period we also tested the presence of MsALR transgene by PCR method. From each transgenic lines 10 MsALR gene positive plantlets were transferred into pots containing the mixture of 50 % Terra peat soil and 50 % sandy soil (Maros). Equal quantities of chemical fertilizer (SUBSTRAL® Osmocote Plus®) were added to each pot (2.8 g/1,870 g soil in pot) at the time of potting. The fertilizer was a complex fertilizer prepared by GARDENA Hungary Ltd. Budapest, included macro elements (10 % N, 10 % P2O5, 12 % K2O, 2 % MgO) supplemented with microelements (0.02 % B, 0.05 % Cu, 0.4 % Fe, 0.06 % Mn, 0.02 % Mo, 0.015 Zn). Single application serves 6 month continuous nutrition for the plants.
In the drought test, five pots from each genotype were exposed to water deficit and five others were treated as controls. We planted one plantlet/plastic pot in phenotyping experiments with our Complex Stress Diagnostic System. In the greenhouse, after planting all pots were irrigated with 100 cm3 water and then the stress treatment was started. Plants were first weighted by a computer-controlled balance (GSE model 350, 6,000 ± 1 g), and the amount of water which was necessary to keep the relative water content of the soil at either 20 % (water limitation) or 60 % (control, optimal water supply) was provided. The water capacity of the soil was determined before the present experiment. First the water from the soil was basically eliminated and this value represented the zero water capacity. We calibrated the maximum water capacity by adding water until saturation level. The 20 and 60 % water capacity values were calculated from these data and used for watering regimes in the Complex Stress Diagnostic System (Majer et al. 2008). The required amounts of water were added using computer-controlled peristaltic pump (Gilson Minipuls 3), while the pot was still on the balance. Each pot had a radiofrequency identifier (RF proximity tag), therefore, watering data could be stored automatically on the computer.
Digital imaging of plants
Growth of wheat plants was monitored by digital photography. Individual plants, planted into radio-tagged pots, were photographed by an Olympus C-7070WZ (Olympus Ltd., UK) digital camera from eleven different sideway positions, produced by 32°–33° step rotation of the pot. The plant pixels were determined by subtracting the homogenous background from the image. Monitoring of plant growth was performed during the whole growth period in every 2 weeks and in a weekly interval after heading. In separate experiment, a calibration curve has been constructed to establish correlation between the green pixel-based shoot surface area and the actual fresh weight of wheat plants determined after cutting the photographed plants.
Thermal imaging of plants
The water evaporation was estimated by measuring leaf temperature using a sensitive thermo camera (VarioSCAN 3021ST, 0.03 °C temperature resolution). These measurements were performed in a closed chamber in the morning hours within the greenhouse. The temperature of leaves of the tillering stage plants was determined as difference relative to the surrounding air. Air temperature data were obtained from a reference surface, which is in thermal equilibrium with air in the background of the plant. The relative humidity was ca. 60–65 %, and the air temperature was in the 17.4–19.7 °C range during the measurements.
Data management and analysis
Data analysis was performed by a home constructed software package based on Matlab software tools (version 2008b) with the Image Processing Toolbox™ (The MathWorks Inc., Natick, MA, USA). The experiments were carried out with at least three plants as replications, but in analysis of grain weight per plants five replications were used. Two-way ANOVA was used to analyze the effect of genotypes and treatment combinations (optimal- and suboptimal water supply). The statistical analysis was carried out by MICROSOFT® EXCEL 2002 statistical software package developed by Microsoft (Redmond, WA, USA) and shows the average thermal values of ‘plant pixels’ and the background.
RNA isolation and quantitative real-time PCR
The cDNAs for the quantitative-PCR reactions were synthesized from the total RNA of control and MsALR expressing transgenic wheat lines isolated according to the AGPC (acid guanidinium thiocyanate-phenol chloroform) method (Chomczynski and Sacchi 1987). 1 μg of total RNA was used for cDNA synthesis, following the First Strand cDNA Synthesis Kit manual (Fermentas Life Sciences).
For the quantitative RT-PCR reactions, we used the following primers: MsALR Fwd: 5′-GGTGTTGTTGCCAAAGCTGTC-3; MsALR Rev: 5′-TGATCCATAACTCCTCGCGC-3′. These were designed by the Primer Express Software from Applied Biosystems. The Q-PCR reactions were carried out in the ABI PRISM 7000 Sequence Detection System using the SYBR Green PCR Master Mix and the reaction conditions were the same as described by Lendvai et al. (2007). The 2−∆∆CT method was used to analyze the real-time PCR data (Livak and Schmittgen 2001) and the expression of examined genes was normalized to the reference gene (β-actin).
Western blot analysis
For Western analysis, proteins were extracted by grinding samples with quartz sand in a homogenization buffer containing 25 mM Tris–HCl pH 7.6, 15 mM MgCl2, 15 mM EGTA, 75 mM NaCl, 1 mM phenylmethylsulfonyl fluoride (PMSF), and 2 mM DTT. Equal protein loading was determined by Bradford protein assay (Bio-Rad, USA). 20–20 μg of protein was separated on 12 % SDS–polyacrylamide gels. The separated proteins were transferred onto polyvinylidene difluoride (PVDF) membranes (Sigma, USA) using the TRANS-BLOT SD Semy-Dry Transfer Cell (Bio-RAD, USA). A polyclonal antibody previously produced in rabbits has been used for the detection of the Medicago sativa L. aldose reductase as described by Oberschall et al. (2000). The goat anti-mouse IgG-peroxidase conjugated secondary antibody was purchased from Sigma (USA). The antigen–antibody complex was detected using the Lumi-Light Western Blotting Substrate (Roche, Switzerland) and X-ray films (AGFA, Germany). Relative band intensities were determined using the Image J software (http://rsbweb.nih.gov/ij/). The used antibody recognized the putative wheat aldose reductase protein in the extracts and its band intensity was used as a reference (=1).
Assay of aldose reductase (ALR) enzyme activities
Wheat leaves were homogenized on ice in extraction buffer (0.1 M Na-phosphate buffer pH 7.0, containing 1 mM PMSF and 1 % polyvinylpyrrolidone). The homogenate was centrifuged for 20 min at 15,000×g at 4 °C and the supernatant (crude extract) was used for enzymatic measurements. Aldose reductase activity was estimated by monitoring the oxidation of NADPH in the reaction mixture at 340 nm according to Vander Jagt et al. (1995). 0.1 M sodium phosphate buffer pH 7.0 contained 0.1 μmol NADPH, 0.1 μmol 2-naphthalene carboxaldehyde (NA) substrate and 50 μl crude extract in a total volume of 1 cm3. The decrease of NADPH was measured between the 3rd and 4th min, the non-specific NADPH decrease was corrected using additional measurements without substrate. In calculations, the molar extinction coefficient of NADPH at 340 nm was 6.22 mM−1 cm−1, the specific activity was expressed as oxidized NADPH μmol min−1 g−1 fresh weight.
Results
Production and molecular characterization of transgenic wheat plants with constitutively increased aldose reductase activity
For detailed studies, we have selected three transgenic lines (AKR 284; AKR 304; AKR 322) with plants carrying both the BAR (PAT) and the MsALR genes in their genome. Expression of the introduced MsALR gene in the transformed wheat plants was shown by the quantification of its mRNA products (Supplementary Fig. 2). The relative level of MsALR transcripts was the highest in plants from the genotype AKR 284 followed by that of AKR 304 and AKR 322. Considerable differences in the expression level of the introduced MsALR gene could be detected between individual plants within the selected transgenic genotypes (data not shown).
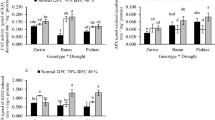
The Western blot analysis of leaf protein extracts detected an increased ALR protein accumulation in leaves of the T1 plants (Fig. 1a, b). This analysis revealed a low amount of cross-reacting protein in leaves from the control, non-transgenic (CY-45) plants. The highest ALR protein amount was present in samples from plants of the transgenic line AKR 284. The enzyme assay measuring aldehyde/aldose reductase activity in crude cell extracts provided further evidence that the transgenic plants have 1.5–4.3 times higher activities than the untransformed control plants (Fig. 1c).

Synthesis of the alfalfa (MsALR) aldose reductase enzyme in transgenic wheat lines. a and b Quantification of ALR protein in leaves of genotypes by Western blot analysis. c Specific aldose reductase activity in leaves of the wheat lines was estimated by monitoring the oxidation of NADPH in the presence of the naphthalene carboxaldehyde as substrate. The mean ± SD were calculated from the data of at least three measurements performed on three different plants. *, ***Significant differences between the control and transgenic plants at P ≤ 0.05 and 0.001 levels, respectively (Student’s t test)
Phenotyping of the vegetative growth of the control and the transgenic wheat plants
Our phenotyping platform uses a computer-controlled water supply system to keep the soil water capacity continuously at 60 % (reference condition with optimal watering) and at 20 % water capacity (water scarcity condition). As shown in the Supplementary Fig. 3, the overall responses of the studied wheat plants to the applied stress resulted in considerable growth retardation but it was not lethal.
Since biomass accumulation is one of the most comprehensive indicators of stress response, this parameter was monitored by recording green pixel-based shoot surface area of wheat plants. The biomass values were calculated with the calibration curve between the normalized plant pixel numbers and the fresh weights of areal plant organs (Fig. 2). In a separate experiment, CY-45 plants were photographed and weighed (see "Materials and methods"): comparison of calculated plant fresh weights from the different lines under non-stressed condition revealed that plants of the transgenic line AKR 322 produced higher biomass than plants of other genotypes (Fig. 3a). Under stress conditions all transgenic lines produced higher biomass than the control plants during the whole vegetative growth. At the last time point of the analysis, the average weight of plants from the control genotype (CY-45) was 2.7 + 0.04 g. Plants of the aldose reductase-overproducing transgenic lines exhibited moderate drought damage as shown by the average weight values as AKR 284 = 3.4 + 0.03; AKR 304 = 3.01 + 0.01 g; AKR 322 = 3.8 + 0.03 g. (Fig. 3b, c). As an example, the improved growth characteristics of transgenic plants can be seen by comparing them to recipient control plants (Fig. 4).

Correlation between fresh weight and green pixel-based shoot surface area in wheat plants. Plants were photographed and their above ground fresh weight was measured as described in “Materials and methods”

Monitoring the biomass production of the control and of the aldose reductase-overproducing transgenic plants by measuring the green pixel-based fresh weight of areal plant organs under optimal (a) and water deficit (b) conditions (means–SE = standard error. Empty square CY-45 (control), cross symbol AKR 284, empty circle AKR 322, empty triangle AKR 304. c Box-plots illustrate the maximum and the minimum values of fresh weight of areal plant organs as well the medians for the different genotypes grown for 9 weeks after planting under water limitation Pooled St Dev = 0.02932

Morphology of plants from the control (CY-45; a) and the transgenic genotype (AKR 284; b) after 8 weeks of drought stress treatment. The improved physiology is reflected by the plant height and advanced development stage in the case of transgenic plants
Over-expression of the aldose reductase detoxification enzyme ensures optimal water use
As suggested by Collins et al. (2008) the stress avoidance may originate from maintaining or even increasing the rate of transpiration and photosynthesis by the excessive use of water. Here we monitored the transpiration function of different genotypes by thermal imaging that allowed determining the temperature difference (ΔT) between the leaf and surrounding air. In the case of well-watered plants, the normal rate of water evaporation resulted in a larger air temperature minus leaf temperature values as shown by ΔT values (3.2–3.6 °C) in Fig. 5. The limitation in water supply increased the leaf temperature of control (CY-45) plants that was reflected by the lower ΔT value (2.7 °C). The thermal imaging data clearly showed the maintenance of transpiration in the case of transgenic plants under water limitation. The ΔT values of these plants remained higher than 3 °C indicating cooler leaf temperature. The described differences in thermal status of tested plants from different genotypes are demonstrated by thermal images shown by Fig. 6.

The temperature differences (ΔT) between leaves and surrounding air indicate a cooler thermal status of leaves of the transgenic plants with better leaf evaporation function. In the control (CY-45) plants, the water deficit resulted in an increase in leaf temperature as shown by the lower ΔT value (2.7 °C). Deviations are marked on the top of columns. Filled bar 60 % soil water capacity. Open bar 20 % soil water capacity

Cooler leaves of the transgenic (AKR 284, AKR 304) plants were detected by the thermal imaging. Under drought condition the leaf of the control (CY-45) wheat plant exhibits an increased temperature according to the artificial color scale representing a 5 °C range with the temperature decrease in the white–red–green–blue direction (color figure online)
The improved physiological status of wheat genotypes synthesizing the alfalfa aldose reductase under drought stress is expected to be consistent with an increase in the water utilization of transgenic plants. The watering system allowed the calculation of the total water amounts used during the life cycle of individual wheat plants. As shown by Fig. 7 in an optimal condition, plants from all genotypes have consumed 2,000 cm3 water during the cultivation period. Under water limitation plants from CY-45 genotype could utilize 430 cm3 water. Plants of all the transgenic lines showed higher water use. These plants absorbed 470–530 cm3 water from the soil with low water capacity.

Plants from the transgenic genotypes (AKR 284; AKR 322; AKR 304) present a better water usage capability than the non-transgenic (CY-45) plants under water stress during the whole life cycle. The standard deviations are marked on the top of the columns. Light grey bars 60 % soil water capacity. Dark grey bars 20 % soil water capacity
Grain weight data show an increased productivity of aldose reductase overproducer plants under permanent drought stress in the greenhouse
After maturation seeds were harvested from five plants per treatment representing plants grown in pots with either optimal or suboptimal water regime. The box-plot figure indicates a strong effect of the different water management and differential responses of the control and the transgenic genotypes to drought (Fig. 8). As shown by analysis of the average grain weight data there was no significant difference among the tested genotypes if the plants were grown in soil with constant 60 % water capacity. The applied water stress caused 59 % reduction in seed production of the control (CY-45) plants. Similar loss (57 %) could be recorded in seed yield in the case of plants from the transgenic AKR 322 genotype. Plants from the other two transgenic genotypes (AKR 284 and AKR 304) yielded significantly higher grain weight than the control or AKR 322 plants (at 0.05 probability level). The statistical analysis (ANOVA) revealed the significant effect of treatments (optimal- and suboptimal irrigation) and genotypes CY-45 and AKR overproducer lines at 0.001 probability level on the yield data (Supplementary table 1). The effect of treatment and genotype interaction was also significant at the 0.001 probability level showing that water scarcity had a significant effect on the control and tested AKR lines, respectively. This drought-induced damage was moderated by an improved physiology of two (AKR 284 and AKR 304) transgenic genotypes carrying the functional alfalfa aldose reductase cDNA.

Box-plot is from data of the grain weight (g) per plants grown under optimal or suboptimal water conditions. 1–4: plants were cultured in pots with normal irrigation; 5–8: plants were cultured in pot with reduced water supply 1 and 5: CY-45 control; 2 and 6: AKR 284; 3 and 7: AKR 304; 4 and 8: AKR 322. LSD5 % = 0.02812
Discussion
The aldo–keto reductase enzyme family attracts increasing attention as an antioxidant defense system also in plants (Bartels 2001). Overproduction of the Medicago sativa L. aldose reductase (MsALR) in tobacco plants could reduce the accumulation of thiobarbituric acid reactive species (TBARS) and H2O2; furthermore, this enzyme detoxified the 4-hydroxynon-2-enal and the methylglyoxal (Oberschall et al. 2000; Hideg et al. 2003). The vegetative organs of these transformants recovered from a severe drought after rewatering. The present study extends the test of this enzyme into a cereal crop such as wheat by analyzing both green biomass and seed yield parameters of transgenic lines exhibiting elevated aldose reductase activity. Recognizing the key role of wheat production in ensuring sustainable food supply worldwide, there is a great need for the functional analysis of genes with potential agronomic benefits also in wheat, despite limitations in the methodology of gene transfer and the regeneration of transformed plants (Zhao et al. 2008).
In the present study we used a morphogenic wheat cultivar (CY-45) for gene bombardment and sufficient number of the Finale 14 SL® resistant plants could be regenerated from immature embryo-derived callus tissues. This method is frequently used for cereal transformation, but the Agrobacterium-mediated transformation is gaining increasing significance (Tamás-Nyitrai et al. 2012). In our study, the selected transgenic wheat genotypes showed high variability in the synthesis of transgene mRNAs (Supplementary Fig. 2) and the accumulation of Medicago MsALR protein (Fig. 1a, b). Expression of this cDNA in wheat plants resulted in higher NADPH-dependent aldose reductase activities in leaf extract (Fig. 1c). In both parameters, we detected considerable variation between the transgenic lines and individual plants but the presented data support a general trend that GM plants exhibit an enhanced detoxification capability for removing stress-generated toxic metabolites. These opened a unique way to evaluate multiple changes caused by the transgene at phenotypic level and test its potential protective function.
The growth, photosynthetic and yield responses to drought stress are highly complex and flexible (Chaves et al. 2008; Passioura 2007). At phenotypic level, the stress exposure of higher plants can alter morphological traits, growth parameters and seed productivity. As reported the large-scale phenotyping of stressed plants could be based on digital image analysis to measure the leaf area and relative growth rate (Granier et al. 2006; Berger et al. 2010). In our studies, we have established a calibration curve to predict the weight of areal plant organs as a major indicator for water deficit tolerance during vegetative growth phase. As shown by Fig. 2, this approach can be safely used in the earlier phase of plant life cycle. During the maturation phase, the senescence of plant organs may influence the quality of pictures; therefore, the correlation between the pixel number and fresh weight becomes lower (Fig. 2). According to the pixel number-derived weight data, the wheat plants carrying the alfalfa detoxification gene were less affected by drought treatment than the control non-transgenic plants (Figs. 3b, 4). At this stage, we have no explanation for the considerably better green biomass production of the plants from the AKR 322 line under optimal growing conditions (Fig. 3a). This side effect may originate from changes caused by the integration of foreign DNA into the wheat genome. The moderate stress tolerance of transgenic plants was shown by variable increase in biomass relative to the control plants. The 12 and 26 % higher plant weight in AKR 284 and 304 lines is expected to be linked with the protective function of the MsALR enzyme. The 41 % improvement in biomass of plants from AKR 322 line cannot be explained by the enzyme data (Fig. 1a, b), therefore, the influence of additional factor may be involved in the case of this genotype.
Among the remote-sensing technologies, the thermal imaging has been successfully applied to monitor leaf temperature being indicative for changes in the leaf transpiration and stomatal closure (Jones 2004). In the control CY-45 wheat plants, the temperature difference between leaves and surrounding air was reduced by near to 0.8 °C after drought treatment. This shows that water limitation caused an increase in canopy temperature (Fig. 5). This leaf temperature status could originate from the stomatal closure and reduced water evaporative activity. In contrast the ΔT values of leaves from plants carrying the transgene did not change significantly during water withdrawal compared to leaf temperatures at optimal water supply (Figs. 5, 6). We can conclude that water metabolism of these plants was functionally less damaged by the drought stress. The computer-controlled water supply system allowed the calculation of the cumulative water use during the complete growing season (Fig. 7). In accordance with the thermal imaging data, this study shows that plants of the GM lines used extra water in comparison with the control CY-45 plants. The higher water usage could contribute to the increased biomass production of these transgenic plants (Fig. 3). The presented green biomass values in conjunction with the canopy temperature and the water use data indicate an improved physiology of the analyzed transgenic plants under non-lethal drought stress. All these observations strengthen the previous results with the transgenic tobacco plants as an example for dicot species (Oberschall et al. 2000).
The present characterization of wheat genotypes was carried out under stabilized soil water content as in other studies (Yu and Setter 2003; Granier et al. 2006). Based on the degree of growth retardation (46 % inhibition in the case of CY-45 plants) the reduction of soil water capacity to 20 % can be considered as an intermediate stress effect, since the wheat plants could survive the treatment (see Supplementary Fig. 3). The degree of stress exposure is a key factor in the analysis of stress tolerance of different genotypes. Skirycz et al. (2011) reported that transgenic Arabidopsis plants with stress tolerance genes showed improved survival under severe drought, whereas these plants failed to grow better under mild drought conditions. This observation emphasizes the need for the precise regulation of the soil’s water status that translates to different degrees of stress. The described phenotyping platform with the computer-controlled water supply ensured the precise regulation of the watering regime. Maintaining the stress exposure during the whole growing period can provide advantages for genotypes with the capability of long-term adaption and efficient use of limited water.
The majority of phenotyping facilities are established in greenhouse and crop plants are cultivated with small amounts of soil in pots. Under these artificial circumstances the cultivation parameters can be standardized but the agronomic value of the tested genotypes cannot be predicted. Acknowledging this limitation we present the seed weight per plant data as physiological parameters indicating the level of stress tolerance. As shown by Fig. 8, the seed production was significantly reduced in the stressed wheat plants. In the case of wild-type plants, seed yield was lowered by the drought stress to 41 % of well-watered plants. Only two (AKR 284, AKR 304) out of three transgenic genotypes could produce significantly more seeds than the control plants. Plants from the AKR 322 genotype showed similar degree of yield loss (57 %) as the CY-45 plants (59 %). Interestingly, this genotype yielded the highest biomass both under optimal and suboptimal conditions (Fig. 3). In phenotyping of drought response of barley genotypes, we failed to detect correlation between shoot surface parameters and seed yield (r 2 = 0.249) (Cseri et al. 2013). The seed productivity may require the functionality of green organs, but another level of control is represented by the reproductive organs.
The described wheat genotypes overproducing the aldose reductase enzyme extend the list of examples for transgenic approaches successfully used to improve the drought tolerance in wheat (Zhao et al. 2008). The mannitol accumulating transgenic line produced 92 % dry weight under mild water stress (88 % soil water capacity) if they were compared to the non-stressed plants (Abebe et al. 2003). The biomass production has been increased by constitutive expression of the ABA-responsive barley HVA1 gene in transgenic wheat lines (Sivamani et al. 2000). Under moderate water deficit conditions, both growth characteristics and water use efficiency were improved as the result of transgenic function. The expression of the Arabidopsis DREB1A transcription factor gene in wheat resulted in the delayed development of water stress symptoms and improved root system (Pellegrineschi et al. 2004). The listed publications are primarily focused on the improved drought response of vegetative organs. In the case of AKR genotypes field studies will be required for the analysis of potential breeding values of these wheat plants with elevated aldose reductase activities.
Author contribution
Erzsébet Fehér-Juhász: wheat transformation and molecular characterization, Petra Majer: phenotyping data collection, László Sass: computer program development and data analysis, Csaba Lantos: statistical analysis, Jolán Csiszár: aldose reductase enzyme activity measurements, Zoltán Turóczy: transcript analysis, Róbert Mihály: wheat transformation, Antal Mai: vector construction, Gábor V. Horváth: isolation and characterization of aldose reductase gene, Imre Vass: establishment of imaging technologies, Dénes Dudits: designing and coordinating the study, editing the manuscript, János Pauk: establishment of the wheat transformation technology and test of plant material in closed system.
References
Abebe T, Guenzi AC, Martin B, Cushman JC (2003) Tolerance of mannitol-accumulating transgenic wheat to water stress and salinity. Plant Physiol 131:1748–1755
Áy Z, Mihály R, Cserháti M, Kótai É, Pauk J (2012) The effect of high concentrations of glufosinate ammonium on the yield components of transgenic spring wheat (Triticum aestivum L.) constitutively expressing the bar gene. ScientificWorldJournal. doi:10.1100/2012/657945
Bartels D (2001) Targeting detoxification pathways: an efficient approach to obtain plants with multiple stress tolerance? Trends Plant Sci 6:284–286
Berger B, Parent B, Tester M (2010) High-throughput shoot imaging to study drought responses. J Exp Bot 61:3519–3528
Bhatnagar-Mathur P, Vadez V, Sharma KK (2008) Transgenic approaches for abiotic stress tolerance in plants: retrospect and prospects. Plant Cell Rep 27:411–424
Capell T, Bassie L, Christou P (2004) Modulation of the polyamine biosynthetic pathway in transgenic rice confers tolerance to drought stress. P Natl Acad Sci USA 101:9909–9914
Castiglioni P, Warner D, Bensen RJ, Anstrom DC, Harrison J, Stoecker M, Abad M, Kumar G, Salvador S, D’Ordine R, Navarro S, Back S, Fernandes MH, Targolli J, Dasgupta S, Bonin C, Luethy MH, Heard JE (2008) Bacterial RNA chaperones confer abiotic stress tolerance in plants and improved grain yield in maize under water-limited conditions. Plant Physiol 147:446–455
Chaves MM, Flexas J, Pinheiro C (2008) Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Ann Bot 103:551–560
Chenu K, Chapman SC, Hammer GL, McLean G, Salah HBH, Tardieu F (2008) Short-term responses of leaf growth rate to water deficit scale up to whole-plant and crop levels: an integrated modelling approach in maize. Plant Cell Environ 31:378–391
Chomczynski P, Sacchi N (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162:156–159
Christensen AH, Quail PH (1996) Ubiquitin promoter-based vectors for high-level of expression of selectable and/or screenable marker gene in monocotyledone plants. Transgenic Res 5:213–218
Collins NC, Tardieu F, Tuberosa R (2008) Quantitative trait loci and crop performance under abiotic stress: where do we stand? Plant Physiol 147:469–486
Cseri A, Sass L, Törjék O, Pauk J, Vass I, Dudits D (2013) Monitoring drought responses of barley genotypes with semi-robotic phenotyping platform and association analysis between recorded traits and allelic variants of some stress genes. Aust J Crop Sci 7:1560–1570
Deikman J, Petracek M, Heard JE (2012) Drought tolerance through biotechnology: improving translation from the laboratory to farmers’ fields. Curr Opin Biotechnol 2:243–250
Fabre J, Dauzat M, Nègre V, Wuyts N, Tireau A, Gennari E, Neveu P, Tisné S, Massonnet C, Hummel I, Granier C (2011) PHENOPSIS DB: an Information System for Arabidopsis thaliana phenotypic data in an environmental context. BMC Plant Biol 11:77–83
Felföldi K, Purnhauser L (1992) Induction of regenerating callus from immature embryos of 44 wheat and 3 Triticale cultivars. Cereal Res Commun 20:273–277
Golzarian MR, Frick RA, Rajendran K, Berger B, Roy S, Tester M, Lun DS (2011) Accurate inference of shoot biomass from high-throughput images of cereal plants. Plant Methods 7:2–12
Granier C, Aguirrezabal L, Chenu K, Cookson SJ, Dauzat M, Hamard P, Thioux JJ, Roland G, Bouchier-Combaud S, Lebaudy A, Muller B, Simonneau T, Tardieu F (2006) FENOPSIS, an automated platform for reproducible phenotyping of plant responses to soils water deficit in Arabidopsis thaliana permitted the identification of an accession with low sensitivity to soil water deficit. New Phytol 169:623–635
Hartmann A, Czauderna T, Hoffmann R, Stein N, Schreiber F (2011) HTPheno: an image analysis pipeline for high-throughput plant phenotyping. BMC Bioinform 12:148–156
Hideg É, Nagy T, Oberschall A, Dudits D, Vass I (2003) Detoxification function of aldose/aldehyde reductase during drought and ultraviolet-B (280–320 nm) stresses. Plant Cell Environ 26:513–522
Houle D, Govindaraju DR, Omholt S (2010) Phenomics: the next challenge. Nat Rev Genet 11:855–866
Hsieh TH, Lee JT, Chang YY, Chan MT (2002) Tomato plants ectopically expressing Arabidopsis CBF1 show enhanced resistance to water deficit stress. Plant Physiol 130:618–626
Jones HG (2004) Application of thermal imaging and infrared sensing in pant physiology and ecophysiology. Adv Bot Res 41:107–163
Karaba A, Dixit S, Greco R, Aharoni A, Trijatmiko KR, Marsch-Martinez N, Krishnan A, Nataraja KN, Udayakumar M, Pereira A (2007) Improvement of water use efficiency in rice by expression of HARDY, on Arabidopsis drought and salt tolerance gene. P Natl Acad Sci USA 104:15270–15275
Lendvai A, Pettkó-Szandtner A, Csordás-Tóth É, Miskolczi P, Horváth GV, Györgyey J, Dudits D (2007) Dicot and monocot plants differ in retinoblastoma-related protein subfamilies. J Exp Bot 58:1663–1675
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2 −Δ ΔCT T method. Methods 25:402–408
Majer P, Sass L, Lelley T, Cseuz L, Vass I, Dudits D, Pauk J (2008) Testing drought tolerance of wheat by complex stress diagnostic system installed in greenhouse. Acta Biol Szeged 52:97–100
Merlot S, Mustilli AC, Genty B, North H, Lefebvre V, Sotta B, Vavasseur A, Giraudat J (2002) Use of infrared thermal imaging to isolate Arabidopsis mutants defective in stomatal regulation. Plant J 30:601–609
Moran JF, Becana M, Iturbe-Ormaetxe I, Frechilla S, Klucas RV, Aparicio-Tejo P (1994) Drought induces oxidative stress in pea plants. Planta 194:346–352
Munns R, James RA, Sirault XR, Furbank RT, Jones HG (2010) New phenotyping methods for screening wheat and barley for beneficial responses to water deficit. J Exp Bot 61:3499–3507
Nelson DE, Repetti PP, Adams TR, Creelman RA, Wu J, Warner DC, Anstrom DC, Bensen RJ, Castiglioni PP, Donnarummo MG et al (2007) Plant nuclear factor Y (NF-Y) B subunits confer drought tolerance and lead to improved corn yields on water-limited acres. P Natl Acad Sci USA 104:16450–16455
Oberschall A, Deák M, Török K, Sass L, Vass I, Kovács I, Fehér A, Dudits D, Horváth VG (2000) A novel aldose/aldehyde reductase protects transgenic plants against lipid peroxidation under chemical and drought stresses. Plant J 24(4):434–446
Passioura J (2007) The drought environment: physical, biological and agricultural perspectives. J Exp Bot 58:113–117
Pellegrineschi A, Reynolds M, Pacheco M, Brito RM, Almeraya R, Yamaguchi-Shinozaki K, Hoisington D (2004) Stress-induced expression in wheat of the Arabidopsis thaliana DREB1A gene delays water stress symptoms under greenhouse conditions. Genome 47:493–500
Rohila JS, Jain RK, Wu R (2002) Genetic improvement of Basmati rice for salt and drought tolerance by regulated expression of a barley Hva1 cDNA. Plant Sci 163:525–532
Salekdeh GH, Reynolds M, Bennett J, Boyer J (2009) Conceptual framework for drought phenotyping during molecular breeding. Trends in Plant Sci 14:488–496
Sivamani E, Bahieldin A, Wraith JM, Al-Niemi T, Dyer WE, Ho T-HD, Qu R (2000) Improved biomass productivity and water use efficiency under water deficit conditions in transgenic wheat constitutively expressing the barley HVA1 gene. Plant Sci 155:1–9
Skirycz A, Vandenbroucke K, Clauw P, Maleux K, De Meyer B, Dhondt S, Pucci A, Gonzalez N, Hoeberichts F, Tognetti VB, Galbiati M, Tonelli C, van Breusegem F, Vuylsteke M, Inzé D (2011) Survival and growth of Arabidopsis plants given limited water are not equal. Nature Biotechnol 29:212–214
Smirnoff N (1993) The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol 125:27–58
Takeda S, Matsuoka M (2008) Genetic approaches to crop improvement: responding to environmental and population changes. Nat Rev Genet 9:444–457
Tamás-Nyitrai C, Jones HD, Tamás L (2012) Biolistic- and Agrobacterium-mediated transformation protocols for wheat. Methods Mol Biol 877:357–384
Tuberosa R, Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops. Trends Plant Sci 11:405–412
Valliyodan B, Nguyen HT (2006) Understanding regulatory networks and engineering for enhanced drought tolerance in plants. Curr Opin Plant Biol 9:189–195
Vander Jagt DL, Kolb NS, Vander Jagt TJ, Chino J, Martinez FJ, Hunsaker LA, Royer RE (1995) Substrate specificity of human aldose reductase. Identification of 4-hydroxynonenal as an endogenous substrate. Biochem Biophys Acta 1249:117–126
Xu C, Jing R, Mao X, Jia X, Chang X (2007) A wheat (Triticum aestivum) protein phosphatase 2A catalytic subunit gene provides enhanced drought tolerance in tobacco. Ann Bot-Lond 99:434–450
Yu L, Setter TL (2003) Comparative transcriptional filing of placenta and endosperm in developing maize kernels in response to water deficit. Plant Physiol 131:568–582
Zhao C-X, Guo L-Y, Cheruth JA, Shao H-B, Yang H-B (2008) Prospectives for applying molecular and genetic methodology to improve wheat cultivars in drought environments. C R Biol 331:579–586
Acknowledgments
This work was supported by a grant from the National Research and Technology Office (Budapest) in the frame of the “Wheat Spike Consortium” and the joint German-Hungarian “NAP_BIO 2006 ALAP3-01435/2006” projects. The improvement of the Complex Plant Stress Diagnostic System was supported by the European Plant Phenotyping Network (EPPN) INFRA-2011-1.1.8 grant (grant agreement number: 284443). The authors express their thanks to Zsuzsa Keczán, Judit Szabad and Klára Godó for their help in the preparation and to Dr Erzsébet Fejes and Dr Attila Fehér for the critical reading of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. V. Huylenbroeck.
E. Fehér-Juhász and P. Majer contributed equally.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Fehér-Juhász, E., Majer, P., Sass, L. et al. Phenotyping shows improved physiological traits and seed yield of transgenic wheat plants expressing the alfalfa aldose reductase under permanent drought stress. Acta Physiol Plant 36, 663–673 (2014). https://doi.org/10.1007/s11738-013-1445-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11738-013-1445-0