
Abstract
The physiological reasons for the differences in sensitivity of C3 and C4 plant species to environmental stresses have not been thoroughly explained. In this study the effects of drought stress on the growth and selected physiological traits were examined in the seedlings of 13 single cross maize (C4 plant) hybrids and 11 spring triticale (C3 plant) breeding lines and varieties differing in drought sensitivity. For plants in the seedling stage the results demonstrated a genetic variation in dry matter accumulation of shoots and roots (DWS, DWR), number (N) and length (L) of particular components (seminal, seminal adventitious, nodal) of the root system, membrane injury by soil drought (LID), osmotic and high temperature stress (LIOS, LIHT), water potential (ψ), water loss (WL), grain germination in osmotic stress (FG, PI), and seedling survival (SS). Seedlings grown under moderate soil drought showed a decrease in dry matter of the top parts and roots and a decrease in the length of seminal, seminal adventitious and nodal roots in comparison to seedlings grown in control conditions. The observed harmful effects of drought stress were more distinct in drought sensitive genotypes. Used in this paper drought susceptibility indexes (DSIGY) were calculated in other experiment by determining the changes in grain yield (GY) under two soil moisture levels (irrigated and drought). The variation of DSIGY for maize ranges from 0.381 to 0.650 and for triticale from 0.354 to 0.578. The correlations between DSIGY and laboratory tests (LI, FG, SS) confirmed that they are good indicators of drought tolerance in plants. The highest values of genetic variation were observed in LI, DWS, SS and WL and the lowest in the measurements of ψ FG, PI, LS, LSA and LN. The correlation coefficients between LIOS and LIHT tests were, in most of the considered cases, statistically significant, which indicates that in maize and triticale the mechanisms of membrane injury caused by simulated drought or high temperature are physiologically similar. It can be concluded that an approach to the breeding of maize and triticale for drought tolerance using these tests can be implemented on the basis of separate selection for each trait or for all of them simultaneously. In that case, it would be necessary to determine the importance of the trait in relation to growth phase, drought timing and level, as well as its associations with morphological traits contributing to drought tolerance. The obtained values of the correlation coefficient between laboratory tests suggest that the same physiological traits may be applied as selection criteria in drought tolerance of maize and triticale genotypes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
As drought is the most important environmental phenomenon affecting a plant’s growth, development and crop yield, considerable progress has been made in understanding the changes in physiological processes caused by drought stress. It has been shown that the physiological responses of plants to drought stress are extremely complex and vary with plant species as well as with the degree and time of the exposure to drought (Levitt 1980; Bennett 1990; Evans et al. 1990, 1991; Jones 1993; Reynolds 2002; King 2011). Plants develop different morphological, physiological and biochemical mechanisms which inhibit or remove the harmful effects of drought stresses (Sullivan and Ross 1979; Boyer 1982; Larsson and Górny 1988; Chaves et al. 2002; Reynolds et al. 1998; Asharaf 2010). Drought tolerance of a plant species is usually determined by the plant’s genes and also by morphological, phonological, physiological, and biochemical traits. The responses of plants to drought stress depend on the species, genotype, plant age, level and duration of drought, and physical parameters of the soil. Differences in tolerance to drought are known to exist within genotypes of plant species and were found in many studies, e.g. in maize (Martinielio and Lorenzoni 1985; Lorens et al. 1987; Grzesiak 1990; Grzesiak et al. 2012), wheat (Winter et al. 1988; Reynolds et al. 1998; Paknejad et al. 2007), rape seed (Richards and Thurling 1978), oat (Larsson and Górny 1988), coconut (Gomez et al. 2008) and triticale (Royo et al. 2000; Grzesiak et al. 2012).
Methods of evaluating the degree of drought tolerance allow for a direct or indirect estimation of the various physiological, biochemical or morphological traits of the examined genotypes. Measurements of different physiological processes of plant response to drought provide important information about the reactions of the plant intended to remove or to reduce the harmful effects of water deficit in the soil or plant tissues. Techniques of screening for drought tolerance were devised by selecting genotypes in a field or greenhouse study. Conducting field experiments is necessary for the verification of the drought tolerance estimated on the basis of physiological laboratory tests (Grzesiak 1990; Richards 1991; Kpoghomou et al. 1990). For proper field testing a number of methodological problems must be solved to enable water content in the soil to be controlled by irrigation or by limiting the inflow of water from rainfall. The relations between the plant yield obtained under conditions of drought and that obtained under conditions of optimal soil moistening were preferred among the field indices of drought tolerance. Such tests, however, are not accurate enough or too simplified to show important relations between the crop forming processes and soil–water-plant relationship. A more precise quantitative formulation of this relationship can be found in the studies by Fischer and Maurer (1978), Hanson and Nelson (1985), Winter et al. (1988), Stanley (1990) and in FAO reports by Doorenbos and Pruit (1977), Doorenbos and Kassam (1986).
Methods of screening for drought tolerance within a large number of genotypes should be easy, rapid and inexpensive (Hanson and Nelson 1985; Palta 1990; Zagdańska 1992). It is also necessary for the laboratory testing method to be characterized by a significant correlation with drought resistance observed under field conditions (Sullivan and Ross 1979; Blum et al. 1980; Bouslama and Schapauch 1984; Hanson and Nelson 1985; Kpoghomou et al. 1990; Chaves et al. 2002; Grzesiak et al. 2012). The most important laboratory methods suggested for screening for drought tolerance in crop plants were: germination in osmotic substances (mannitol, PEG), growth or survival of young seedlings subjected to soil or simulated water stress and high temperature stress (Sullivan and Ross 1979; Blum and Ebercon 1981; Martinielio and Lorenzoni 1985), leaf injury, leaf water content, leaf temperature and parameters of leaf gaseous exchange (Passioura et al. 1993; Farquhar et al. 1993; Dubey 1997). Other traits that may be promising as screening traits are different parameters of chlorophyll fluorescence (Reynolds 2002; Hura et al. 2007) and associations between dark respiration under drought conditions and heat tolerance of sorghum lines (Gerik and Eastin 1985) and of wheat (Reynolds et al. 1998).
The physiological reasons for the differences in sensitivity of C3 and C4 plant species to environmental stresses have not been thoroughly explained and understood (Edwards and Ku 1987; Medrano et al. 2002; Nayyar and Gupta 2006; Lopes et al. 2011). Maize and triticale have different types of photosynthesis (C3—in triticale, C4—in maize), different metabolic pathways and structure of bundle sheath chloroplasts (Kranz syndrome). According to Iijima and Kono (1991), cereal species develop two types of root system, depending on the angle of the growth of branches (lateral roots) and their distribution in the soil profile. For triticale, the root system structure is “concentrated” whereas it is “scattered” for maize. Maize and spring triticale appear to be sensitive to drought stress during grain germination, seedling emergence, early vegetative growth and pollination. Moreover, both maize and triticale are important crops widely cultivated throughout the world (Fageria et al. 2006).
The objectives of this study were to evaluate the variation of selected physiological characteristics of maize and triticale seedlings grown in drought conditions and to compare them to variations in drought susceptibility index based on grain yield (DSIGY) of plants grown in a stressed environment under field conditions and then determine which tests are most useful for screening drought resistance genotypes.
Materials and methods
Plant materials
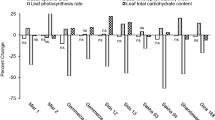
Experiments were conducted on 13 single-cross maize hybrids and on 4 strains and 7 cultivars of spring triticale. Maize grains were obtained from Pioneer Overseas GmbH (Austria), Pioneer Saaten (Poland), Garst Seed Company (USA), Agriculture Canada and SAMPLO Holding (Slovakia). Triticale grains of breeding lines were obtained from DANKO in Choryn and cultivars from IHAR, Małyszyn (Poland). The choice of the maize hybrids and triticale lines and cultivars to be examined was done on the basis of the information on the effect of drought on plant yield received from breeders. Plant materials and drought susceptibility indexes (DSIGY) according to Grzesiak et al. (2012) are listed in Fig. 1.

Maize and triticale genotypes ordered according to their Drought Susceptibility Index (DSIGY)
Experimental conditions
The experiment was carried out in a greenhouse under the conditions of 25/20 °C day/night temperature and 65 % relative humidity. Plants were grown in Mitscherlich pots and root-boxes filled with a mixture of garden soil, peat and sand (1:1:1). Air-dried soil substrate was sieved in a 0.25 cm mesh. Soil substrate pH was 7.1 and the percent of organic material was 0.7 %. For the determination of root length and number of root system components, seedlings were grown in root-boxes, which enabled non-destructive isolation of all compartments of the root system (Kono et al. 1987). A set for the “root-box and pin board method” consists of: a Plexiglas box (width—0.25 m, depth—0.40 m, thickness—0.02 m), a pin board for sampling the root system, and a polyethylene sheet (envelope) for handling and preserving the root system. In root-boxes soil compaction level was 1.30 g cm−3. Mechanical impedance in soil substrate was measured with penetrometer DIK-5520 (Daiki Rika Kogyo Co. Ltd, Japan).
Field soil water capacity (FWC) for soil mixture was determined according to Kopecky methods. Air-dried soil samples were placed inside metal cylinders, with a 1 mm hole at the bottom. The volume for the samples was 100 cm3. Cylinders with the samples were placed inside a container with water for 30 min. After 8 h, maximal soil water content in the samples was 0.43 (g cm−3) and after 48 h it decreased to 0.21 (g cm−3). Following Hillel and van Bavel (1976), those last values were assumed to be 100 % of soil field water capacity (FWC). The pots and root-boxes were weighed every day, and the amount of water loss through transpiration was added to maintain the original weight in each treatment. For control (C) treatment soil water content was maintained from sowing for 42 days at the level of 65–70 % FWC. Soil water content in the root-box experiment for drought (D) treatment was kept at the level of 30–35 % FWC from the 21st to 42nd day. Similarly, in the pot experiment from the 21st to 42nd day it was kept at the level of 30–35 % FWC for drought treatment D35 and at the level 15–20 % FWC for treatment D20. In order to limit water evaporation from pots and root-boxes, soil surface was covered with 1 cm layer of ground Styrofoam. A single pregerminated grain was planted at the depth of 3–4 cm. After 42 days of plants’ growth under C treatment Zadoks scale was about 17 or 18 for maize and about 16 or 17 for triticale.
On the 42th day seedlings grown in pots were used for measurements of leaf water potential, membrane injury and water loss in the excised leaf. For each species (2), genotypes (13 or 11) and treatments (3) 5 pots with 3 plants were used (n = 5). Similarly, seedlings grown in root-boxes were cut into top parts and roots for the determination of dry weigh (DWS, DWR) and number and length of particular components of the root system. The roots were sampled after the soil from the pot had been washed away by a gentle stream of water. After the measurements root samples were preserved in a FAA (formalin, acetic acid, and ethanol) solution. For each species (2), genotypes (13 or 11) and treatments (2) 4 root-boxes with 1 plants were used (n = 4).
Measurements
Germination and seedling survival (FG, PI, SS). Twenty grains of each genotype were germinated under 0.00, −0.47, −0.85 and −1.30 MPa of osmotic stress. Grains of the tested genotypes were surface sterilized in 70 % ethanol for 5 min and placed on petri dishes and incubated in an air-conditioned growth cabinet at the temperature of 25 °C. Osmotic stress was simulated using mannitol solutions (C6H1406, M.W. 182.17), Lobe Chemia. The concentration of solutions at the desired chemical water potential (ψ) was calculated according to Michel et al. (1983), the control was distilled water (ψ = 0.0 MPa). Germination was recorded when the radicle reached 5 mm in length. There were 4 replications for each treatments. Counts of germinated grains were made each day to compute the final germination percentage (FG) and promptness index (PI) and calculated as follows:
where ndx = number of germinated grains by the xth day, Seedling survival (SS), 25 germinated grains were planted in 5 pots and after 14 days the number of growing seedling was recorded, and seedling survival index (SS) was calculated as follows:
Leaf water potential (ψ) was measured with thermocouple psychrometer HR 33T (Wescor Inc., Logan, USA) in “dew point” mode, equipped with sample chamber C-52 SF and digital multimeter Metex M-3640 D. Measurements were taken on leaf discs—diameter of 0.3 cm for triticale and 0.5 cm for maize—and immediately placed inside the psychrometer chamber and left to balance temperature and water vapor equilibrium for 30 min before measurements. For each treatments there were 5 replications. Psychrometric readout and ψ determination were made as described by Johnson and Brown (1977).
Excised-leaves water loss (WL) was measured only for control treatment (C) of each maize and triticale genotype. Leaves were cut from the upper half of the plant, weighed (FW0) and left to desiccate at 25 °C in the dark. After 12 and 24 h samples were reweighed (FW12, FW24) and next oven dried at 70 °C and weighed again for the determination of dry weight of samples (DW). There were 5 replications. Water loss of excised leaves (WL) was calculated by the following formula:
Dry weight of shoots (DWS) and roots (DWR) was sampled in each root-box and was determined on the 42nd day after sowing through drying at 65 °C for 72 h.
Number (N) and length (L) of seedling root components (seminal-LS, seminal adventitious-LSA, nodal-NN·LN) was measured with DELTA-T SCAN (England) analyzer.
Relative loss of intracellular electrolytes from leaf tissues (LI) was measured with the conductivity method using conductivity meter OK-102/1 (Radelkis, Hungary), according to the procedure and formula described by Sullivan and Ross (1979) and Blum et al. (1980).
where C and T refer to the conductivity of control and treatment solutions, respectively, and subscript 1 and 2 refer to initial and final conductance, respectively.
Nine leaf discs (0.5 cm diameter for maize and 0.3 cm for triticale) were cut from leaves and immersed in test tubes containing 30 cm3 redistilled water. After 24 h initial conductance measurements were taken. Final conductance measurements were taken after autoclaving all tubes at 110 °C for 15 min and cooling them to room temperature.
In the experiment additional conductivity tests of leaf injury by simulated drought (LIOS) and by high temperature (LIHT) were made. For these tests leaf discs were cut only from control (C) plants. For LIOS leaf discs were immersed for 12 h in test tubes with 30 cm3 of redistilled water (control 0.0 MPa) or 30 cm3 of mannitol solution (treatment −0.47, −0.85 and −1.30 MPa). High temperature stress (LIHT) was imposed by immersing leaf discs for 1 h in test tube with 30 cm3 of redistilled water by placing them in well-stirred water bath at three temperatures: 25 °C (control), 35 and 45 °C (treatments). There were 5 replications for each treatments.
All measurements except germination tests (FG, PI, SS) were made after 42 days of seedlings’ growth. Samples for measurements of leaf water potential (ψ), relative loss of intracellular electrolytes (LI), and water loss of excised leaves (WL) were taken between 11:00 and 13:00 on most recent fully expanded leaf. Results of all measurements are presented as stress index (SI) which was calculated as indicated below:
Statistical analysis
All data were analyzed by one-way ANOVA and LSD tests (p < 0.05) using Statistica 10.0 (StatSoft, Inc., USA). Linear correlation analyses were used to determine the relationship between drought susceptibility index (DSIGY) and stress indices (SI) obtained in laboratory tests. Also correlation coefficients were calculated for the determination of the relationship between and within laboratory tests.
Results and discussion
Drought susceptibility indexes (DSIGY) of maize and triticale genotypes
According to our earlier research (Grzesiak et al. 2012), drought susceptibility indexes (DSIGY) for maize and triticale genotypes were calculated by determining the changes in grain yield (GY) under two soil moisture levels (irrigated and drought). Variation of DSIGY for maize ranges from 0.381 to 0.650 and for triticale from 0.354 to 0.578. The values of DSIGY made it possible to rank the examined maize and triticale genotypes according to their drought tolerance. In the maize hybrids the drought resistant group (0.381 < DSIGY > 0.439) comprises the hybrids Tina, Garst 8344, Pioneer 3925 and Pioneer 3957, while the drought sensitive group (0.607 < DSIGY > 0.650) comprises the hybrids Ankora, Garst 8702, Garst 8388 and Garst R5515. For triticale the drought resistant group (0.354 < DSIGY > 0.419) comprises the lines CHD 247 and CHD 220, and cultivars Migo and Wanad, while the drought sensitive group (0.544 < DSIGY > 0.578) comprises the strains CHD 147 and CHD 12, and cultivars Mieszko and Maja (Fig. 1).
Germination under simulated drought conditions (FG, PI) and seedling survival (SS)
In the control treatment (0.0 MPa), maize and triticale genotypes did not differ in the final germination index (FG) and the correlation coefficients between FG and DSIGY were insignificant. The imposed drought from −0.43 to 1.30 MPa caused a decrease in the values of stress indexes of FG and the correlation coefficients between FG and DSIGY were statistically significant for maize in −0.43 MPa treatment and for triticale in −0.85 and −1.30 MPa treatments (Table 1). Differences between maize and triticale genotypes in promptness index (PI) values in the control were statistically significant only for maize, whilst the correlation coefficient with DSIGY was statistically significant for both species. As with drought FG, osmotic stress caused a decrease in the values of PI and the correlation coefficients between PI and DSIGY were statistically significant for maize in −0.43 and −0.85 MPa treatments, but for triticale only in −1.30 MPa treatment (Table 2). The results of seedling survival (SS) after grain germination in different water potential of mannitol solutions are presented in Table 3. Germination under osmotic stress conditions (−0.43, −0.85. −1.30 MPa) caused a decrease in the number of live seedlings in maize to 86, 79 and 76 %, respectively, and for triticale to 97, 86 and 84 %, respectively. Correlation coefficients between DSIGY and the values of stress index of seedling survival were statistically significant in −0.43, −0.85 and −1.30 MPa treatments of maize genotypes, but for triticale only −0.85 and −1.30 MPa treatments were affected.
Germination is strongly influenced by plant species, grain age and storage conditions and is also highly sensitive to soil water quality and temperature (Ashraf and Mehmood 1990; Ahmad et al. 2009). Grain germination indexes were used with different results in selecting procedures for identifying drought resistant genotypes (Jajarmini 2009). Opinions on the efficiency of plant drought tolerance assessments on the basis of different parameters of germination are divergent and indicate their limited usefulness. It is believed that germination traits are affected by drought stress less than other physiological and biochemical traits (Winter et al.1988; Morgan 1992). Genotype tolerance to drought stress is process-specific with regard to grain water imbibition, endosperm utilization, activation of the dormant enzyme system and seedling emergence, growth and survival after stress. The tolerance to drought in any of those stages will affect the results and thus screening for drought tolerance reaction should also be process-specific, and the prediction of genotype performance from one process to another would not necessarily be possible (Blum et al. 1980; Ahmad et al. 2009). In this study the tolerance to simulated drought stress proved a relatively good indicator in the screening of maize and triticale genotypes. Our results showed that the efficiency of FG, PI and SS is related to how strongly a trait is expressed and the measurement must be performed at the right moment in order to maximize the expression of genetic variations of the particular trait.
Seedling dry matter (DWS, DWR), root length (LS, LSA, LN) and the number of nodal roots (NN)
Soil drought decreased the dry matter of the above ground parts (DWS) in 13 genotypes of maize from 12.0 to 67.0 % and in 11 genotypes of triticale from 5.0 to 17.0 %. Similarly, the dry matter of roots (DWR) decreased in maize genotypes from 3.0 to 30.0 % and in triticale genotypes from 1.0 to 25.0 %. Drought also strongly influenced the ratio of DWS to DWR within maize genotypes from 0.0 to 60.0 % and in triticale genotypes from 0.0 to 25.0 % (Table 4). The variation coefficients (CV) in control (C) seedlings of DWS, DWR and the ratio of DWS to DWR in maize were about 29, 18 and 37 %, respectively, and in triticale 23, 29 and 22 %, respectively. CV calculated for treatment D as a stress index (SI) for those traits in maize were about 34, 13 and 31 %, respectively, and in triticale 15, 9 and 11 %, respectively (Table 4). In the control treatment (C), the statistically significant correlation between DSIGY and DWS, DWR and the ratio of DWS to DWR were found only in triticale for the ratio of DWS to DWR. In the drought treatment, the statistically significant correlation between DSIGY and DWS, DWR and the ratio of DWS to DWR were found in maize for DWS and the ratio of DWS to DWR, and in triticale, only for DWS (Table 4).
The results presented in Table 5 show that in the control (C) treatment, differences within drought resistant and drought sensitive genotypes of maize and triticale in the length of seminal (LS), seminal adventitious (LSA), nodal roots (LN), and number of nodal roots (NN) were in most cases statistically insignificant. Also under control conditions the correlation coefficient between DSIGY and the measured traits was statistically insignificant. LS in drought treatment decreased in the drought resistant genotypes of maize (Tina, Garst 8344) about 10 % and in drought sensitive genotypes (Ancora, Garst 8702) about 23 %. In drought resistant genotypes of triticale (CHD-220, CHD-247) the decrease was about 6 % and in drought sensitive genotypes (CHD-12, CHD-147) about 13 %. In maize the decrease in the total length of two seminal adventitious roots (LSA) in comparison to the control was about 7 % in drought resistant hybrids and in drought sensitive about 25 %. For triticale the decrease in the total length of three seminal adventitious (LSA) roots in comparison to the control was statistically insignificant in drought resistant and sensitive lines. In seedlings exposed to drought, significant differences between drought resistant and sensitive genotypes were observed in the measurements of the total length of nodal roots (LN). In maize the decrease in LN in drought sensitive genotypes was about 22 % and in the drought resistant ones about 10 %, and in triticale about 30 and 13 %, respectively. In C treatments no statistically significant correlation between DSIGY and LS, LSA, NN and LN was found in either maize or triticale genotypes. In seedlings exposed to drought, a statistically significant correlation between DSIGY and SI calculated for the measurements of root traits was found in maize, but in triticale only for LN (Table 5).
As one might expect, a root system characteristic, such as the number and length of particular components of root system structure, their depth and abundance, are known to be associated with performance under drought conditions in many studies of cereal species (Richards 1996; Reynolds 2002). Nevertheless, decreased allocation in roots in the top layer of the soil has been shown to be an effective drought stress adaptive mechanism (Richards 1991). In the studies by Kono et al. (1987) the specific response of cereal species to drought stress was clearly noticeable in root distribution, nodal root number, leaf number and grain yield. Those authors reported that the responses in root growth for maize and rice were different in the downward penetration of the main axis and in the higher order laterals. Species with a “concentrated” type of root system showed less restriction of root and shoot growth compared to species with a “scattered” type. A decrease in size of the root system and an increasing irregularity of root distribution resulted in water and nutrients being transported greater distances to the nearest roots (Tardieu 1991; Lipiec et al. 1996). Drought also modifies root system components, such as the main root axis and lateral roots of different orders in rice and maize (Iijima and Kono 1991). Changes in the morphological structure of the root system in triticale and maize were also observed in our earlier studies in response to waterlogging and soil compaction (Grzesiak et al. 2012).
Leaf water potential (ψ) and water loss of excised leaves (WL)
Moderate (D35) and severe (D20) soil drought decreased ψ in maize and triticale (Table 6). Differences between resistant genotypes of maize (Garst 8344, Tina) and triticale (CHD-247, CHD-220) in terms of a decrease of ψ were statistically significant in comparison with sensitive genotypes of maize (Ankora, Garst 8702, Garst 8388) and triticale (Maja, CHD-247).
In seedlings exposed to drought, a statistically significant correlation between DSIGY and SI calculated for the measurement of ψ in maize was found for both treatments but in triticale only for treatment D20. The differences in WL between drought resistant and sensitive genotypes of maize and triticale genotypes were statistically significant. A statistically significant correlation between DSIGY and WL in maize was found only in the measurements taken after 12 h of leaf desiccation but in triticale in the measurements taken after 12 and 24 h (Table 6).
Decreases in leaf water potential initially induced stomatal closure, resulting in a decrease in the supply of CO2 to the mesophyll cells and subsequently in a decrease in the rate of leaf photosynthesis (Williams et al. 1999; Lawlor and Cornic 2002). According to Hura et al. (2007), a statistically significant correlation between water potential and photosynthetic rate and stomatal conductance was found in maize and triticale during various stages of plant development. Dehydration in C3 and C4 plants impairs various physiological processes, especially the changes in leaf water content, water potential and photosynthesis. There is a controversy as to whether drought limits photosynthesis due to leaf water status, stomatal closure, metabolic impairment or injuries to photosynthetic apparatus (Flexas et al. 2006). The first response to leaf water deficit is stomata closure, which limits CO2 diffusion to chloroplasts (Berkowitz et al. 1983; Cornic and Masacci 1996; Muller and Whitsitt 1996). Non-stomatal mechanisms under prolonged or severe soil drought include changes in chlorophyll synthesis, functional and structural changes in chloroplasts and also disturbances in accumulation and distribution of assimilation products (Medrano et al. 2002). However, it is known that during drought stress, plants with C4 photosynthesis increase water use efficiency and suppress photorespiration. Thus, C4 plants are often more competitive than C3 plants in drought-prone areas (Edwards and Ku 1987; King 2011).
Relative loss of intracellular electrolytes from leaf tissues (LI)
In drought sensitive genotypes of maize and triticale, the values of indexes of leaf injury by soil drought (D35, D20), osmotic stress (−0.47, −0.85, −1.30 MPa), and heat temperature (25, 35, 45 °C) were in general higher than in drought resistant genotypes and in most cases the differences between resistant and sensitive genotypes were statistically significant (Table 7). In treatment D35, the values of coefficients of variation (CV) in maize and triticale were higher than in D20 treatment. Under osmotic stress, higher values of CV were found in −0.85 MPa treatment and under high temperature stress in 35 °C treatment.
The ability to maintain the structure and function of cytoplasmatic membranes under water deficit is one of the most important physiological traits. Conductometric measurements of LI are applied as a screening test for the estimation of tolerance to various stresses (Vietor et al. 1977; Richards 1978; Blum and Ebercon 1981; Poljakoff-Mayber 1981; Martinielio and Lorenzoni 1985; Palta 1990). Differences between sensitive and resistant genotypes might stem from the fact that drought resistant genotypes possess more efficient mechanisms protecting membrane functions and structure. Drought stress causes a loosening of lamellar membranes in chloroplasts, loss of a certain amount of grana, and increase in the level of coarse-grain matrix (Haupt-Harting and Fock 2002; Lawlor and Cornic 2002; Tang et al. 2002). Some authors suggest that drought resistant plant species show stronger binding of chlorophyll molecules to the lipid-protein complex of chloroplast membranes (Smirnoff and Colombe 1988; Bukhov et al. 1990). Our earlier results indicate that leaf age is very important because differences in LI between drought resistant and sensitive genotypes were smallest in the oldest and youngest leaves, though the greatest differences were observed in the leaves where cellular divisions had taken place and which had reached the maximal area (Grzesiak et al. 2006).
Correlations among stress parameters (DWS, DWR, DWS/DWR) were significant, except for the relationship between DWS and DWR in maize. In this experiment the correlations between DWR and other traits were not significant except for seedling survival (SS). For both species, high and significant correlations were found between measurements of membrane injures due to drought (LID), osmotic stress (LIOS), and high temperature (LIHT). Also for both species, high and significant correlations were found between leaf water potential (ψ) and water loss (WL) and between FG and PI, except for maize (Table 8).
Most of physiological processes are affected by the stresses of soil drought, osmotic and high temperatures (Levitt 1980). The usefulness of methods for studying plant drought tolerance has been discussed in many papers and reviews (Blum et al. 1980; Kpoghomou et al. 1990; Zagdańska 1992; Reynolds 2002). According to some authors, besides the evaluation criteria used in the present research, positive results were also obtained by utilizing the measurements of canopy infra-red temperature, changes of leaf color and responses of root architecture (Clarke and McCaig 1982; Stanley 1990). Partly positive results were obtained in tests of the plant’s ability to reduce leaf area, the development of wax bloom on leaves, leaf rolling and the ability of the leaf to hold water (Passioura et al. 1993). On the other hand, the effectiveness of the evaluation of tolerance based on the measurement of different parameters of leaf gaseous exchange and the content of various metabolites, including proline, has not been definitely confirmed. (Hanson and Nelson 1985; Farquhar et al. 1993; Dubey 1997; Bandurska and Stroiński 2003).
The maize and triticale genotypes used in these experiments show a relatively wide range of drought tolerance. This study contributes to the understanding of responses of different genotypes to drought, though in this research, the method of estimating drought susceptibility index was relatively simple and did not take into account other important factors of soil–water-plant relationship. The correlations between DSIGY and laboratory tests (LI, FG, SS) showed that they are good indicators of plant drought tolerance. The correlation coefficients between LIOS and LIHT tests were, in most of the considered cases, statistically significant, thus indicating that in maize and triticale the mechanisms of membrane injury caused by simulated drought or high temperature were physiologically similar. It can be concluded that an approach to the breeding of maize and triticale for drought tolerance using these tests can be implemented on the basis of separate selection for each trait or for all of them simultaneously. In that case, it would be necessary to determine the importance of the trait in relation to the growth phase, drought timing and level as well as associations with morphological traits contributing to drought tolerance (Kono et al. 1987; Kpoghomou et al. 1990).
The results presented in this paper, our earlier research (Grzesiak 1990; Grzesiak et al. 2012) and the results of other authors (Lorens et al. 1987; Martinielio and Lorenzoni 1985; Kono et al. 1987) confirm the existence of a wide range of genotypic variability of response to drought in cereal plants. In maize and triticale, similar to other crop plants, the physiological reasons for this variability have not as yet been fully recognized and explained. In the literature one can find many contradictory conclusions, as the reduction of crop yield of the tested genotypes depends not only on the drought level and its duration, but also on the phase of plant growth and development and interaction with other environmental factors (Hanson and Nelson 1985; Blum 1988; Naylor and Su 1998).
The grounds for genotypic variation of drought tolerance in cereals has not yet been entirely elucidated and future research is necessary (Zagdańska 1992; Royo et al. 2000; Reynolds 2002). The frequent occurrence of drought stress in many regions of the world and the deteriorating water conditions for plant growth and productivity have raised interest in research into the responses of crops to periodic water deficiency. Progress in the breeding of drought resistant cereal plants requires future study of the physiological mechanisms underlying the responses of plants to water stress. It is a widely held opinion that the breeding of drought resistant crop plants will not be an easy task (Hanson and Nelson 1985; Zagdańska 1992; Reynolds 2002). The complexity of the property of drought tolerance will make it necessary to take into consideration various tolerance evaluation criteria in the breeding programs (Levitt 1980; Turner 1986; Blum 1988; Richards 1991; Jones 1993; King 2011).
Author contribution
MTG, FJ, PW, KH designed the research; MTG, PS, IM and TG conducted the research; MTG, FJ, PS and KH analyzed the data; MTG, FJ, IM and KH wrote the paper; MTG had primary responsibility for the final content. All authors have read and approved the final manuscript.
Abbreviations
- DSIGY :
-
Drought susceptibility index
- C:
-
Control
- D:
-
Drought
- SI:
-
Stress index
- FWC:
-
Field water capacity
- FG:
-
Final germination
- PI:
-
Promptness index
- SS:
-
Seedling survival
- LI:
-
Membrane injury index
- Ψ:
-
Leaf water potential
- WL:
-
Water loss
- DWR :
-
Root dry matter
- DWS :
-
Shoot dry matter
- FW:
-
Leaf fresh weight
- DW:
-
Leaf dry matter
- LS, LSA, LN :
-
Length of seminal, seminal adventitious and nodal roots, respectively
- NN :
-
Number of nodal roots
References
Ahmad S, Ahmad R, Ashraf MY, Ashraf M, Waraich EA (2009) Sunflower (Helianthus annuus L.) response to drought stress at germination and seedling growth stages. Pak J Bot 41:647–654
Asharaf M (2010) Inducing drought tolerance in plants: recent advances. Biotech Adv 28:199–238
Ashraf M, Mehmood S (1990) Response of four Brassica species to drought stress. Environ Exp Bot 30:93–100
Bandurska H, Stroiński A (2003) ABA and proline accumulation in leaves and roots of wild (Hordeum spontaneum) and cultivated (Hordeum vulgare Maresi) barley genotypes under water deficyt conditions. Acta Physiol Plant 25:55–61
Bennett JM (1990) Problems associated with the measuring plant water status. HortScience 25:1551–1554
Berkowitz GA, Chen C, Gibbs M (1983) Stromal acidification mediates in vivo water stress inhibition of nonstomatal controlled photosynthesis. Plant Physiol 72:1123–1126
Blum A (1988) Plant breeding for stress environments. CRC Press, Boca Raton
Blum A, Ebercon A (1981) Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop Sci 21:43–47
Blum A, Sinmena B, Ziv O (1980) An evaluation of seed and seedling drought tolerance screening tests in wheat. Euphytica 29:727–736
Bouslama M, Schapauch WT (1984) Stress tolerance in soybean. I. Evaluation of three screening techniques for heat and drought tolerance. Crop Sci 24:933–937
Boyer JS (1982) Plant productivity and environment. Science 218:443–448
Bukhov NG, Sabat SC, Mohanty P (1990) Analysis of chorophyll a fluorescence changes in weak light in heat-treated Amaranthus chloroplasts. Photosyn Res 23:81–87
Chaves MM, Pereira JS, Maroco J, Rodrigues ML, Ricardo CPP, Osorio LM, Carvalho I, Faria T, Pinheiro C (2002) How plants cope with water stress in the field. Photosynthesis and growth. Ann Bot 89:907–916
Clarke JM, McCaig TN (1982) Evaluation of techniques for screening for drought resistance in wheat. Crop Sci 22:503–506
Cornic G, Masacci A (1996) Leaf photosynthesis under drought stress. In: Baker NR (ed) Photosynthesis and the environment. Kluwer Academic Publication, Dordrecht, pp 347–366
Doorenbos J, Kassam AH (1986) Yield response to water. FAO Irrigation and drainage paper. Food and Agriculture Organization of the United Nations. Rome
Doorenbos J, Pruit WO (1977) Guidelines for predicting crop water requirements. FAO Irrigation and drainage paper. Food and Agriculture Organization of the United Nations. Rome
Dubey RS (1997) Photosynthesis in plants under stressful conditions. In: Pessarakli M (ed) Handbook of Photosynthesis, University of Arizona, Tuscon. Arizona. Marcel Dekker, Inc., New York, pp 859–875
Edwards GE, Ku MSB (1987) Biochemistry of C3–C4 intermediates. In: Hatch MD, Boardman NK (eds) The Biochemistry of plants, a comprehensive treatise, vol 10. Photosynthesis. Academic Press, New York, pp 275–324
Evans RO, Skagss RW, Sneed RE (1990) Normalized crop susceptibility factors for corn and soybean to excess water stress. Trans ASAE 33:1153–1161
Evans RO, Skagss RW, Sneed RE (1991) Stress day index models to predict corn and soybean relative yield under high water table conditions. Trans ASAE 5:1997–2005
Fageria NK, Balingar VC, Clark RB (2006) Physiology of crop production. The Haworth Press Inc., New York, pp 23–60
Farquhar GD, Wong SC, Evans JR, Hubiek KT (1993) Photosynthesis and gas exchange. In: Jones HG, Flowers TJ, Jones MB (eds) Plants under stress. Cambridge University Press, New York, pp 47–70
Fischer RA, Maurer R (1978) Drought resistance in spring wheat cultivars. I Grain yield responses. Aus J Agric Res 29:897–912
Flexas J, Bota J, Galme′s J, Medrano H, Ribas-Carbo′ M (2006) Keeping a positive carbon balance under adverse conditions: responses of photosynthesis and respiration to water stress. Physiol Plant 127:343–352
Gerik TJ, Eastin JD (1985) Temperature effects on dark respiration among diverse sorghum genotypes. Crop Sci 25:957–961
Gomez F, Oliva MA, Mielke MS, de Almeida AAF, Leita HG, Aquino LA (2008) Photosynthetic limitations in leaves of young Brazilian Green Dwarf coconut (Cocos macifera L.) ‘‘nana’’ palm under well-watered conditions or recovering from drought stress. Environ Exp Bot 62:195–204
Grzesiak S (1990) Reaction to drought of inbreds and hybrids of maize (Zea mays L.) as evaluated in field and greenhouse experiments. Maydica 35:303–331
Grzesiak MT, Grzesiak S, Skoczowski A (2006) Changes of leaf water potential and gas exchange during and after drought in triticale and maize genotypes differing in drought tolerance. Photosyntetica 44:561–568
Grzesiak MT, Marcińska I, Janowiak F, Rzepka A, Hura T (2012) The relationship between seedling growth and grain yield under drought conditions in maize and triticale genotypes. Acta Physiol Plantarum. doi:10.1007/s11738-012-0973-3
Hanson AD, Nelson ChE (1985) Water adaptation of crop to drought. In: Carlson PS (ed) The biology of crop productivity. Academic Press, New York, pp 79–149
Haupt-Herting S, Fock HP (2002) Oxygen exchange in relation to carbon assimilation in water-stressed leaves during photosynthesis. Ann Bot 89:851–859
Hillel D, Van Bavel CHM (1976) Simulation of profile water storage as related to soil hydraulic properties. Soil Sci Soc Am J 40:s807–s815
Hura T, Hura K, Grzesiak MT, Rzepka A (2007) Effect of long-term drought stress on leaf gas exchange and fluorescence parameters in C3 and C4 plants. Acta Physiol Plantarum 29:103–113
Iijima M, Kono Y (1991) Interspecific differences of the root system structures of four cereal species as affected by soil compaction. Jpn J Crop Sci 60:130–138
Jajarmini V (2009) Effects of water stresson germination indices in seven wheat cultivar. World Acad Sci Eng Technol 45:105–106
Johnson DA, Brown RW (1977) Psychrometric analysis of turgor pressure response; a possible technique for evaluating plant water stress resistance. Crop Sci 17:507–510
Jones HG (1993) Drought tolerance and water-use efficiency. In: Smith JAC, Griffiths H (eds) Water deficits plant responses from cell to community. Bios Scientific Publishers Limited, Oxford, pp 193–204
King J (2011) Reaching for the sun. How plant work. Part IV Stress, defense, and decline, 2nd edn. Cambridge University Press, Cambridge, p 185–244
Kono Y, Yamauchi A, Nonoyama T, Tatsumi T, Kawamura N (1987) A revised system of root-soil interaction for laboratory work. Environ Conltrol Biol 25:141–151
Kpoghomou BK, Sapra VT, Beyl CA (1990) Screening for drought tolerance: soybean germination and its relationship to seedling response. J Agron Crop Sci 164:153–159
Larsson S, Górny AG (1988) Grain yield and drought resistance indices of oat cultivars in field rain shelter and laboratory experiments. J Agron Crop Sci 161:277–286
Lawlor DW, Cornic G (2002) Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ 25:275–294
Levitt J (1980) Response of plants to environmental stresses. Academic Press, New York
Lipiec J, Ishioka T, Szustak A, Pietrusiewicz J, Stępniewski W (1996) Effects of soil compaction and transient oxygen deficiency on growth, water use and stomatal resistance in maize. Acta Agric Scand Sect Soil Plant Sci 46:186–191
Lopes MS, Araus JL, van Heerden PDR, Foyer CH (2011) Enhancing drought tolerance in C4 crops. J Exp Bot 62:3135–3153
Lorens GF, Bennett JM, Loggale LB (1987) Differences in drought resistance between two corn hybrids. I. Water relations and root length density. Agron J 79:802–807
Martinielio P, Lorenzoni C (1985) Response of maize genotypes to drought tolerance tests. Maydica 30:361–370
Medrano H, Escalona JM, Bota J, Gulias J, Flexas J (2002) Regulation of photosynthesis of C3 plants in response to progressive drought: stomatal conductance as a reference parameter. Ann Bot 89:895–905
Michel BE, Wiggins KO, Outlow WHJ (1983) A guide to establishing water potential for aqueous two-phase solutions (Polyethylene glycol plus dextran) by amendment with mannitol. Plant Physiol 72:60–65
Morgan JM (1992) Osmotic components and properties associated with genotypic differences in osmoregulation in wheat. Aust J Plant Physiol 19:67–76
Muller JE, Whitsitt MS (1996) Plant cellular response to water deficit. Plant Growth Regul 20:41–46
Naylor RES, Su J (1998) Plant development of triticale cv. Lasko at different sowing date. J Agric Sci 130:297–306
Nayyar H, Gupta D (2006) Differential sensitivity of C3 and C4 plants to water deficit stress: association with oxidative stress and antioxidants. Environ Exp Bot 58:106–113
Paknejad F, Nasri M, Moghadam HRT, Zahedi H, Alahmadi MF (2007) Effects of drought stress on chlorophyll fluorescence parameters, chlorophyll content and grain yield of wheat cultivars. J Biol Sci 7:841–847
Palta JP (1990) Stress interactions at the cellular and membrane level. Hort. Sci. 25:1337–1381
Passioura JB, Condon AG, Richards RA (1993) Water deficits, the development of leaf area and crop productivity. In: Smith JAC, Griffiths H (eds) Water deficits plant responses from cell to community. BIOS Scientific Publishers Ltd, Oxford, pp 253–264
Poljakoff-Mayber A (1981) Ultrastructural consequences of drought. In: Paleg LG, Aspinall D (eds) The physiology and biochemistry of drought resistance in plants. Academic Press, New York, pp 389–403
Reynolds MP (2002) Physiological approaches to wheat breeding. In: Curtis BC, Rajaram S, Gomez Macpherson H (Eds) Bread wheat: improvement and production. Food and Agriculture Organization, Rome. pp118–140
Reynolds MP, Singh RP, Ibrahim A, Agech OAA, Larque-Saavedra A, Quick JS (1998) Evaluation physiological traits to complement empirical selection for wheat in warm environments. Euphytica 100:84–95
Richards RA (1978) Variation between and within species of rapeseed (Brassica campestris and B. napus) in response to drought stress. III. Physiological and physicochemical characters. Aust J Agric Res 29:495–501
Richards RA (1991) Crop improvement for temperate Australia: future opportunities. Field Crop Res 39:141–169
Richards RA (1996) Defining selection criteria to improve yield under drought. Plant Grow Regul 20:157–166
Richards RA, Thurling N (1978) Variation between and within species of rapeseed (Brasica campestris and B. napus) in response to drought stress. I. Sensitivity at different stages of development. Aust J Agric Res 29:469–477
Royo C, Abaza M, Bianco R, del Moral LFG (2000) Triticale grain growth and morphometry as affected by drought stress, late sowing and simulated drought stress. Aust J Physiol 27:1051–1059
Smirnoff N, Colombe SV (1988) Drought influences the activity of enzymes of the chloroplast hydrogen peroxide scavenging system. J Exp Bot 39:1097–1108
Stanley CD (1990) Proper use and data interpretation for plant- and soil water status measuring instrumentation: introductory remarks. HortScience 25:1534
Sullivan ChY, Ross WN (1979) Selecting for drought and heat resistance in grain sorghum. In: Mussel H, Staples R (eds) Stress physiology in crop plant. Wiley, New York, pp 263–281
Tang AC, Kawamitsu Y, Kanechi M, Boyer JS (2002) Photosynthetic oxygen evolution at low water potential in leaf discs lacking an epidermis. Ann Bot 89:861–870
Tardieu F (1991) Spatial arrangement of maize roots in the field. In: McMichael BL, Person H (eds) Plant roots and their environment. Elsevier, Amsterdam, pp 506–514
Turner NC (1986) Adaptation to water deficits: a changing perspective. Aust J Plant Physiol 13:175–190
Vietor DM, Ariyanayagam RP, Musgrave RB (1977) Photosynthetic selection of Zea mays L. I. Plant age and leaf position effects and relationship between leaf and canopy rates. Crop Sci 17:567–573
Williams MH, Rosenqvist E, Buchhave M (1999) Response of potted miniature roses (Rosaxhybrida) to reduced water availability during production. J Hortic Sci Biotechnol 74:301–308
Winter SR, Musick JT, Porter KB (1988) Evaluation of screening techniques for breeding drought resistant winter wheat. Crop Sci 28:512–516
Zagdańska B (1992) Physiological criteria for estimation of plant resistance to drought. Biul Inst Hod i Aklimatyzacji Roślin 183:11–19 (In Polish)
Acknowledgments
The authors are grateful to the National Science Centre (NCN) for financial support (Project No. N N310 782540). We are thankful to SEMPOL–Holding, Trnava, Slovakia and Choryn Breeding Station for providing maize and triticale genotypes.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by W. Filek.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Grzesiak, M.T., Waligórski, P., Janowiak, F. et al. The relations between drought susceptibility index based on grain yield (DSIGY) and key physiological seedling traits in maize and triticale genotypes. Acta Physiol Plant 35, 549–565 (2013). https://doi.org/10.1007/s11738-012-1097-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11738-012-1097-5