
Abstract
Three varieties of oil palm seedlings (Deli Yangambi, Deli Urt, Deli AVROS) were exposed to three levels of CO2 (400, 800, 1,200 μmol/mol) in split plot design to determine growth (net assimilation rate, NAR; relative growth rate, RGR) and photosynthetic patterns of the seedlings under short-term CO2 exposure of 15 weeks. Increasing CO2 from 400 to 800 and 1,200 μmol/mol significantly enhanced total biomass and leaf area, net photosynthesis (A) and water use efficiency (WUE) especially from weeks 9 to 15. By the end of week 15, total biomass increased by 113%, and A and WUE by one- and fivefold, respectively, while specific leaf area decreased by 37%. Both enhanced biomass and A under elevated CO2 were effective in modifying NAR and RGR as shown by high correlation coefficient values (r 2 = 0.68 and 0.72; r 2 = 0.63 and 0.67, respectively), although WUE seemed to have more influence over the NAR (r 2 = 0.97) and RGR (r 2 = 0.93). Neither interspecific preference nor its interaction with CO2 imposed any significant effect on parameters observed. Growth improvement with CO2 seemed able to produce healthy, bigger and vigorous oil palm seedlings, and the technique may have potential to be developed for use to reduce nursery period.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Nursery stage is very crucial phase for oil palm planting material. Due to replanting of old and unproductive palms that are more than 25 years old, the demand for oil palm seedling in Malaysia keeps expanding by the years (MPOB 2007). Mortality rate in the field is usually high due to weak seedlings and possible infestation by pests (beetle, rats and wild boars) which can be a serious problem in some plantations. Vigorous and healthy oil palm planting materials should be used in the transplants to the filed to ensure low mortality rate. Subsequently, this would ensure high accumulated yields and harvest for that transplanted to the field would ensure good yield and harvest for next 20 years. So, good establishment during this stage is vital.
Generally, standard nursery period takes about 13–15 months before seedlings are ready for field planting. As the nursery period prolongs, mortality rate in the field tends to decrease. Carlos et al. (2006) have observed this phenomenon where seedlings extended in the field for more than 18 months (also known as advanced planting material; APM) produced higher accumulated early yields of their first 10 years compared to palms planted from conventionally raised seedlings. These palms also had high standing biomass due to increased leaf area and frond numbers, which are able to intercept more sunlight, hence enhancing net assimilation per plant growth. However, the practice of APM nursery requires wide spaces and high capital in maintaining the palms. The supplies of APM are also inconsistent and scarce. The introduction of a new technique that is able to reduce the nursery time and enhance standing biomass of planting material is important for the supply of good, uniform and vigorous palms for successful field planting.
Lengthening the time in nursery might produce vigorous palms that could ensure successful field establishment, and that have the potential for high initial accumulated yield than plants coming from traditional nurseries (Carlos et al. 2006). The use of CO2 enrichment may substitute for the length of time in the nurseries. Basically, increasing CO2 level is able to enhance photosynthesis, reduce photorespiration rate and produce extra photoassimilates that are partitioned to plant organs for stimulation of growth (Eric et al. 2000; Jaafar 2004). The latter is caused by enhanced maximum photosynthesis (A max) and quantum efficiency (α) that is simultaneously followed by the reduction in dark respiration rate (Jaafar et al. 2005; Loats and Rebbeck 1999). Under elevated CO2, carboxylation process also increases producing glycolate and inhibiting photorespiration. These effects are important and they increase the efficiency of net carbon gain by decreasing photorespiratory CO2 loss and diverting ATP and NADPH away from photorespiratory metabolism to photosynthetic assimilation (Elizabeth and Alistair 2007). The extra assimilates produced are then allocated to biomass and leaf area resulted in increases in total dry weight, shoot to root ratio, leaf thickness, stem diameter, relative growth rate (RGR) and net assimilation rate (NAR). However, growth enhancement under elevated CO2 is dependent on the plant species, duration of CO2 exposure and the sink strength of the plant (Kowlozsky et al. 1991). The seedlings that are enhanced with elevated CO2 also usually have higher water use efficiency (WUE) as decrease in stomatal aperture and stomatal density reduce stomatal conductance (g s) and subsequently reducing transpiration rate (Ghannoum et al. 2000).
This technique may have high potential to revolutionize oil palm nursery management where early growth and biomass of oil palm seedling could be enhanced by CO2 enrichment, thus reducing nursery period. Information on growth enhancement of oil palm seedlings using CO2 enrichment technique, however, are still insufficient due to lack of studies being carried out. The present work aims to investigate the growth responses of three commercial tenera progenies of oil palm seedlings exposed to three levels of elevated CO2 in order to determine the best CO2 concentration and the most responsive progenies to duration of CO2 enrichment. Total plant biomass, leaf area per plant, specific leaf area (SLA), shoot to root ratio were also correlated with NAR and RGR to determine the factors that are more influential on the growth patterns of oil palm seedlings under CO2 enrichment.
Materials and methods
The 3-month-old tenera (Dura × Pesifera) oil palm progenies of Deli Urt, Deli Yangambi and Deli AVROS were left for 2 months to acclimatize in a nursery until ready for the treatments. Carbon dioxide enrichment treatments started when seedlings reached 5 months old by exposing them to three levels of CO2, viz., ambient CO2 (400 μmol/mol), twice ambient CO2 (800 μmol/mol) and thrice ambient CO2 (1,200 μmol/mol). The split plot 3 × 3 factorial experiment was designed using randomized complete block design with CO2 levels being the main plot and progenies as the sub-plot replicated three times. Each treatment consisted of ten seedlings. Carbon dioxide at 99.8% purity was supplied from a high-pressure CO2 cylinder and injected through a pressure regulator into fully sealed 2 m × 3 m growth compartment at 2 h/day from 08:00 to 10:00 (Jaafar 2006) continuously. The CO2 concentration at different enrichment treatments was measured using Vaisala™ (Ouijart, Finland) CO2 portable meter during CO2 exposition period.
Total plant biomass was taken by calculating the dry weight of root, boles and leaves per seedling. Destructive plant analysis was carried out every 3 weeks for 15 weeks. The plant parts were placed in paper bags and oven dried at 80°C until constant weight was reached using electronic weighing scale (CDS 125, Mitutoyo Inc, Japan). Leaf area per plant was measured using a leaf area meter (LI-3100, Lincoln Inc, USA). The leaves were arranged within the field of view, and overlapping of adjacent leaves was avoided. Growth analysis was calculated on an individual plant basis through the measurement of total plant leaf area and dry weight. NAR, RGR and SLA were calculated based on biomass and leaf area parameters according to the formula reported by Gardner et al. (1986). The mean value of 18 representative seedlings was used for each plot.
Leaf gas exchange measurements were carried out on weeks 9 and 15 after exposition with CO2 enrichment. Measurements were taken using closed system, infrared gas analyzer LICOR 6400 Portable Photosynthesis System (IRGA: LICOR Inc., Lincoln, NE, USA) by placing the lamina of fully expanded leaflets of the second frond on the terminal bifid lobes in a leaf cuvette supported by a tripod stand and set with optimal growth conditions. The measurements used were standard optimal cuvette conditions for oil palm at 1,000 μmol/m2/s photosynthetic photon flux density, 400 μmol/mol CO2, 30°C leaf temperature and 60% relative humidity. Photosynthesis (A) and instantaneous WUE were calculated using the equation by Tanner and Sinclair (1983). Data were analyzed using analysis of variance by SPSS version 13. Mean separation tests between treatments were performed using Duncan multiple range test and standard error of differences with the assumption that data were normally distributed and equally replicated.
Results
Plant total biomass
Generally, most of treatment effects were contributed by CO2 enrichment; no progeny and interaction effects between two factors were observed on all parameters. As weeks progressed, plant total biomass showed an increasing trend with those enhanced under CO2 enrichment being enhanced significantly in all weeks measured (Fig. 1). However, total biomass of oil palm seedling under 800 and 1,200 μmol/mol was significantly more than the control from week 3 onwards till the end of the experiment although differences between elevated 800 and 1,200 μmol/mol were not observed. The highest biomass increment was shown from weeks 12 to 15 with elevated treatments (800 and 1,200 μmol/mol) where total plant biomass had increased by 1.6-fold from the previous week 12 (62.88 g/plant) to week 15 (225.5 g/plant) compared to total plant biomass from the ambient treatment (400 μmol/mol) which had increased only by onefold (34.2 vs 105.81 g/plant for the 12th and 15th week, respectively).

Total biomass of oil palm seedling (g) as affected by different levels of CO2 during 15 weeks of exposure. Data are mean ± SEM of 18 replicates. Mean values followed by *P ≤ 0.05 differ significantly by Duncan multiple range test procedure. There were no progenies and interaction effect (CO2 × progeny) observed during the experimental period
Total leaf area
Similar to plant total biomass, total leaf area of oil palm seedlings was also influenced by CO2 levels. Neither progeny nor interaction effects between factors were observed during all weeks of measurement (Fig. 2). Although total leaf area in all treatments increased with age due to ontogenical phase, seedlings enriched with elevated CO2 significantly (P ≤ 0.05) recorded higher total leaf area compared to the control starting from week 6 after exposure onwards. There was, however, no significant difference on total leaf area obtained between the elevated CO2 treatments (800 and 1,200 μmol/mol). The lowest enhancement of total leaf area in elevated treatments was shown in week 3 where total leaf area was increased by 32% in elevated treatments (800 and 1,200 μmol/mol) compared to control. Acute increase in total leaf area under elevated CO2 was observed between week 9 (1,085 vs 2,299) and week 12 (2,981 vs 4,561) with respective 2.0- and 1.5-fold enhancement compared to the control.

Total leaf area (cm2) of oil palm seedling as affected by different levels of CO2 during 15 weeks of exposure. Data are mean ± SEM of 18 replicates. Mean values followed by **P ≤ 0.01 differ significantly by Duncan multiple range test procedure. There were no progenies and interaction (CO2 × progeny) effects observed during the experimental period
Specific leaf area
As both plant biomass and total leaf area increased with time, especially under CO2 enrichment, SLA showed a decreasing trend (Fig. 3). However, SLA was only influenced (P ≥ 0.05) by CO2 levels, on week 15, which recorded lower SLA in plants exposed to 800 and 1,200 μmol/mol compared to control by 37% (61 vs 98 cm2/g).

Specific leaf area (cm2/g) of oil palm seedling as affected by different levels of CO2 during 15 weeks of exposure. Data are mean ± SEM of 18 replicates. Mean values followed by **P ≤ 0.01 differ significantly by Duncan multiple range test procedure. There were no progenies and interaction (CO2 × progeny) effects observed during the experimental period
Relative growth rate
Contrastingly, the reduction in SLA was followed by increasing trends of RGR from week 6 until 15 after start of treatments (Fig. 4). The RGR of the seedlings was significantly enhanced under elevated CO2 compared to the control at weeks 9 and 15. Throughout the experiment, palms exposed to 15 weeks of elevated treatments displayed higher RGR than control palms (400 μmol/mol). However, there was no enhancement of the RGR observed between palms in 800 and 1,200 μmol/mol enrichment treatments.

Relative growth rate (g/g/week) of oil palm seedling as affected by different levels of CO2 during 15 weeks of exposure. Data are mean ± SEM of 18 replicates. Mean values followed by *P ≤ 0.05 and **P ≤ 0.01 differ significantly by Duncan multiple range test (DNMRT) procedure. There were no progenies and interaction effect (CO2 × progeny) observed during the experimental period
Net assimilation rate
Similar to RGR, NAR also increased as the weeks increased. Oil palm seedlings enriched with CO2 gave the higher NAR than control seedlings (Fig. 5) from weeks 6 to 15 although differences in NAR between treatments were only observed significant on week 15. NAR on the latter week was found to increase by 124 and 113% for 1,200 and 800 μmol/mol treatments, respectively, compared to the control (400 μmol/mol).

Net assimilation rate (g/cm2/week) of oil palm seedling as affected by different levels of CO2 during 15 weeks of exposure. Data are mean ± SEM of 18 replicates. Mean values followed by **P ≤ 0.01 differ significantly by Duncan multiple range test procedure. There were no progenies and interaction (CO2 × progeny) effects observed during the experimental period
Shoot to root ratio
The balance between shoot and root growth of oil palm seedlings under elevated CO2 is presented in Fig. 6. Oil palm seedlings mostly partitioned their biomass to the shoot from week 3 to 15. No significant difference was observed throughout the weeks. Shoot to root ratio of oil palm seedlings was not affected by the progeny, CO2 treatments or their interaction. However, there was an increasing trend in shoot:root ratio for plants under elevated CO2 (800 and 1,200 μmol/mol) throughout the experiment period compared to the control, implying that there was higher investment of seedling to the shoot under elevated CO2.

Shoot to root ratio of oil palm seedling as affected by different levels of CO2 during 15 weeks of exposure. Data are mean ± SEM of 18 replicates
Net photosynthesis
The A pattern of seedlings enriched with CO2 was shown in Fig. 7. Carbon dioxide enrichment significantly (P ≤ 0.01) influenced A of oil palm seedlings on the 9 and 15 weeks after treatment. After 9 weeks of CO2 exposure, A of oil palm seedlings increased tremendously by 60 and 41% under 800 and 1,200 μmol/mol treatments, respectively, compared to the control. Extending the CO2 exposure into the 15th week developed a general decrease in A of seedlings in elevated treatments compared to the previous reading at week 9. However, seedlings under CO2 elevation depicted a very significant enhancement of A at both 800 and 1,200 μmol/mol CO2 by respective 140 and 112% compared to the control. There was no clear difference observed between the two elevated CO2 treatments.

Net photosynthesis (A; μmol/m2/s) of oil palm seedling as affected by different levels of CO2 during 15 weeks of exposure. Data are mean ± SEM of 18 replicates. Mean values followed by **P ≤ 0.01 differ significantly by Duncan multiple range test procedure. There were no progenies and interaction (CO2 × progeny) effects observed during the experimental period
Water use efficiency
Similar to A, CO2 significantly enhanced WUE of oil palm seedlings (Fig. 8). In week 9 after exposure, 800 and 1,200 μmol/mol CO2 enrichment treatments significantly (P ≤ 0.01) recorded higher WUE compared to control with former treatment registering a onefold (8.36 vs 4.14) higher value. However, there was no clear difference existed between the two elevated CO2 treatments. By the end of the experiment (week 15), WUE had slightly decreased in all treatments although WUE of seedlings exposed to 800 and 1,200 μmol/mol was still higher than the control by four- and sevenfold, respectively. A very obvious difference was also noted for WUE between the two CO2 treatments.

Water use efficiency (WUE; μmol CO2 assimilated/mmol H2O loss) of oil palm seedling as affected by different levels of CO2 during 15 weeks of exposure. Data are mean ± SEM of 18 replicates. Mean values followed by **P ≤ 0.01 differ significantly by Duncan multiple range test procedure. There were no progenies and interaction (CO2 × progeny) effects observed during the experimental period
Correlation analysis
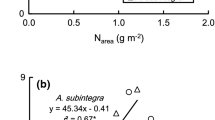
Correlation analysis was carried out to establish relationship among parameters. There was a significant positive linear relationship between RGR and NAR recording a correlation coefficient value of 0.6586 (Fig. 9a). RGR and NAR displayed a very significant positive linear relationship with plant total biomass (Fig. 9b) and a quadratic relationship with total leaf area (Fig. 9c). In both relationships, RGR depicted higher correlation coefficient values than NAR with respective values of 0.7207 and 0.678 for total plant biomass and 0.5016 and 0.3829 for total leaf area. The latter had weaker relationship with RGR and NAR compared to the former as indicated by the lower correlation coefficient. The increase in SLA reduced RGR and NAR as indicated by the negative correlation coefficients of −0.5167 and −0.4811, respectively (Fig. 9d). Meanwhile, shoot to root ratio showed positive linear relationship with RGR and NAR depicting values of 0.1668 and 0.1781, respectively (Fig. 10a). Leaf gas exchange parameters of A (Fig. 10b) and WUE (Fig. 10c) also recorded significant positive linear relationships with RGR and NAR. Stronger linear relationship was observed between net photosynthesis and RGR than the former with NAR as translated by the respective correlation coefficients of 0.6699 vs 0.6345. While A depicted a positive linear relationship with WUE recording a high correlation value (0.7124), its relationship with SLA was a quadratic (0.3273; Fig. 10d). A stronger relationship between growth (RGR and NAR) and WUE was observed in a significant positive linear relationship (Fig. 10c) as indicated by the high values of correlation coefficients at respective 0.9718 and 0.9343.

Relationships between relative growth rate (RGR) and net assimilation rate (NAR) (a), total biomass, NAR and RGR (b), total leaf area, NAR and RGR (c) and specific leaf area (SLA), NAR and RGR (d). *P ≤ 0.05, **P ≤ 0.01. n = 54

Relationships between shoot: root, NAR and RGR (a), net photosynthesis (A), NAR and RGR (b), water use efficiency (WUE), NAR and RGR (c) and net photosynthesis (A), WUE and SLA (d). *P ≤ 0.05, **P ≤ 0.01. n = 54
Total biomass (Fig. 11) and total leaf area (Fig. 12) of seedlings under both the ambient and CO2-enriched conditions were regressed against duration of exposure period to treatment for 15 weeks, and positive linear relationships were established between these parameters and the exposure period. The calculated data from regression formula were plotted to obtain an extended period of time to 65 weeks, which enabled the estimation of total biomass and total leaf area of seedlings under both ambient and CO2-enriched conditions. Under normal ambient condition, a 52-week seedling ready for transplanting recorded total biomass of about 285 g, and total leaf area of 8,450 cm2, while it took only 25 and 30 weeks, respectively, for seedlings under 4-month CO2 enrichment to achieve similar values, exhibiting a reduction in time by 52% for total biomass (Fig. 11) and 42% for total leaf area (Fig. 12).

Comparison of total plant biomass of oil palm seedlings between elevated and ambient treatments. Under elevated CO2 enrichment the 25-week-old oil palm seedlings had attained similar value of total plant biomass at 285 g as the 52-week-old seedlings grown under an ambient CO2 condition

Comparison of total leaf area of oil palm seedlings between elevated and ambient treatments. The 30-week-old seedlings exposed to elevated CO2 enrichment had exhibited similar value of total leaf area at 8,450 cm2 as the 52-week-old seedlings under ambient CO2 condition
Discussion
Carbon dioxide significantly (P ≤ 0.05) improved total biomass accumulation in double and triple than ambient CO2 treatments in all weeks measured. Result showed that oil palm seedlings exposed to increased CO2 concentration had developed sufficient sink strength to accommodate the high level of CO2 enrichment, thus enhancing the total biomass accumulation (Jeffrey and Richard 1999). Increasing level of CO2 to 1,200 μmol/mol did not further enhance growth. The enhancement of total dry weight might have been attributed to starch accumulation in plant parts, which comprises of 20–50% of total plant weight (Idso and Kimball 1988). The positive response of total biomass under elevated CO2 seemed to be the main factor that had contributed to growth enhancement as being signified by the higher correlation coefficient values with RGR (0.7207) and NAR (0.6781) compared to the other measured parameters (total leaf area, SLA, shoot to root ratio). In the present study, elevated CO2 had shown to enhance both the RGR and NAR of oil palm seedlings throughout the weeks of measurement. By the end of week 15, 113% increase in biomass in elevated treatments had increased RGR and NAR by 92 and 118%, respectively, than the control.
As plant total biomass increased, total leaf area also increased and this was solely influenced by CO2 enrichment, particularly in weeks 6, 9, 12 and 15 after the start of treatments. Throughout these weeks, elevated CO2 (800 and 1,200 μmol/mol) resulted in higher total leaf area compared to the control palms. Elevated CO2 improved light harvesting component of oil palm seedlings as increased biomass produced under elevated CO2 is being partitioned to the leaves as manifested by the positive linear relationships between total leaf area and RGR and NAR. Although total leaf area was enhanced by elevated CO2, significant reduction in SLA occurred more with increasing CO2 than with the ambient. SLA had shown to be negatively correlated with RGR and NAR. This implies that the leaves of oil palm seedlings enhanced with 2 × and 3 × than ambient CO2 were thicker than control, probably due to thicker mesophyll layer under elevated CO2, which results in more efficient photosynthesis per area basis (Beadle 1998).
Net photosynthesis (A) under CO2 enrichment was found to be highest in week 9, prolonging the enrichment to 15 weeks recorded comparatively lower than A. The latter might be due to starch accumulation in the leaves under prolonged elevated CO2, which could impair net photosynthesis by disrupting the chlorophyll protein that reduced net photosynthesis (Bindi et al. 2002). Although A decreased under prolonged exposure to CO2, it did not affect growth parameters as plant total biomass and total leaf area continuously increased and the significant differences in growth between ambient and elevated CO2 were maintained during the experimental period. Similar result was observed with sugarcane grown under closed top chamber where peak A was recorded at the middle of the experiment and slightly reduced with extended CO2 imposition (Amanda et al. 2008). However, plant height, leaf biomass and stem biomass of enriched sugarcane were found to be continuously improved growth as indicated by the significant positive linear relationship between RGR and NAR. It was observed that the enhancement of A in oil palm seedlings under elevated CO2 might be attributed to increase in leaf thickness or reduced SLA. The latter was also found to be negatively correlated with A suggesting that thicker mesophyll layer might contain high photosynthesis protein especially Rubisco that might up-regulate several enzymes related to carbon metabolism (Vu et al. 2006).
Increase in A simultaneously enhanced WUE of oil palm seedling grown under elevated CO2. As WUE increased, growth was acutely stimulated as shown in Fig. 10c where increases in WUE enhanced NAR and RGR very significantly recording respective correlations of 0.9718 and 0.9343. As much as 60% increase in A in enriched seedlings (800 and 1,200 μmol/mol) improves WUE by 100% compared to the control recorded at week 9, while in week 15, 140% increase in A enhanced WUE by 610%. The improvement in WUE is due to an enhanced A and reduced transpiration rate under elevated CO2 (Ainsworth and Rogers 2007). According to Taylor et al. (1994), improvement of WUE under elevated CO2 correlated with greater plant turgor pressure, as shown by the higher total leaf area values recorded in the present work. Subsequently, this could explain the higher rate of leaf expansion, which also occurred under elevated CO2.
The enrichment of oil palm seedlings by CO2 can reduce the nursery time. The CO2-enriched seedlings demonstrated higher growth (total biomass and leaf area) than the ambient-treated palms. From calculation and the observed regression data, oil palm seedlings enriched with CO2 for 25 weeks would have similar total plant biomass as the 52-week-old seedlings raised in conventional nursery under ambient CO2 condition, exhibiting a reduction in nursery time by 27 weeks (6.8 months; Fig. 11). In the case of total leaf area, the reduction in nursery time is about 22 weeks to achieve similar total leaf area as in enriched seedlings (5.5 months; Fig. 12). Results showed the possibility of enhancing growth and reducing nursery period through enrichment of oil palm seedlings with CO2.
The positive response observed in growth for oil palm in elevated CO2 could also be attributed to an increase cell production or cell elongation (Pritchard et al. 1999). This hypothesis was supported by the findings of Amanda et al. (2008) that cell wall-related genes, viz., alpha-l-arabinofuranosidase (EC 3.2.1.55), xyloglucan endotransglycosylase/hydoxase (EC 2.4.1.207), caffeoyl-CoA-3-O-methyltransferase (EC 2.1.1.68) and cell wall invertase (EC 3.2.1.26) were induced under elevated CO2. Higher sucrose content of seedlings under elevated CO2 has also been suggested by Ranasinghe and Taylor (1996) to contribute in the increased growth due to improved activities of growth enzyme known as cyclin-dependent protein kinases (EC 2.7.11.22) that enhances cell division.
Conclusion
The manipulation of CO2 enrichment on oil palm seedling seemed able to reduce the nursery phase through growth enhancement, which is solely contributed by CO2 enrichment. Neither progenical differences in tenera oil palm nor the interaction effects between factors were observed. Growth under CO2 enrichment was stimulated through enhancement of plant total biomass than increase in total leaf area. Although net photosynthesis and WUE decreased towards the end of experiment, they did not affect growth parameters, as plant total biomass and leaf area continued to increase, and the significant differences in growth between ambient (400 μmol/mol) and elevated CO2 (800 and 1,200 μmol/mol CO2) were observed during all the experimental weeks. However, there were no significant differences between 800 and 1,200 μmol/mol CO2. Hence, CO2 enrichment has a vast potential to be developed to substitute the need for APM for vigorous and healthy oil palm planting materials.
References
Ainsworth EA, Rogers A (2007) The response of photosynthetic and stomatal conductance to rising CO2: mechanism and environmental interaction. Plant Cell Environ 30:258–270
Amanda PDS, Marilla G, Emerson A, Euginio CU, Allesandro JW, Milton Y, Renato V, Marcelo MT, Glaucia MS, Marcos SB (2008) Elevated CO2 increases photosynthesis, biomass and productivity, and modifies gene expression in sugarcane. Plant Cell Environ 31:1116–1127
Beadle CL (1998) Growth analysis. In: Hall DO, Scurlock JMO, Bolhar N, Leegood RC, Long SP (eds) Photosynthesis and production in a changing environment. Chapman & Hall, New York
Bindi M, Fibbi L, Fra BA, Chiessi M, Selvaggi G, Magliudo V (2002) Free air carbon dioxide enrichment of potato (Solanum tuberosum). Eur J Agric 17:319–335
Carlos MIC, Juan B, Geovany C, Alfredo S (2006) Advanced oil palm planting material: vegetative growth and yield. http://www.asd-cr.com/ASD-Pub/Bo117/B17c11NG.htm. Accessed 27 May 2006
Elizabeth AA, Alistair R (2007) The response of photosynthesis and stomatal conductance to rising CO2: mechanism and environment interaction. Plant Cell Environ 30:258–270
Eric LS, Donald RO, Evan HD (2000) Diurnal regulation of photosynthesis in understory saplings. New Phytol 145:39–49
Gardner FPI, Pearce RB, Mitchell RL (1986) Physiology of crop plants. Iowa States University Press, Amsterdam
Ghannoum O, Von CS, Ziska LH, Conroy JP (2000) The growth response of C4 plants to rising CO2 partial pressure: a reassessment. Plant Cell Environ 23:931–942
Jaafar HZE (2004) Response of Brassica chinensis var. chinensis under CO2-enriched controlled environment production system in the tropics. Trans Malays Soc Plant Physiol 13:59–62
Jaafar HZE, Mohd Haniff H, Ramlan MF, Mohd Hafiz I (2005) Carbon dioxide fertilization for seedling management in the tropics. Trans Malays Soc Plant Physiol 14:75–77
Idso SB, Kimball BA (1988) Atmospheric CO2 enrichment and plant dry matter content. Agric For Meteorol 43:171–181
Jaafar HZE (2006) Carbon dioxide enrichment technology for improved productivity under controlled environment system in the tropics. Acta Hortic 742:353–363
Jeffrey DH, Richard BT (1999) Effects of carbon dioxide enrichment on the photosynthetic light response of sun and shade leaves of canopy sweetgum trees in a forest ecosystem. Tree Physiol 19:779–786
Kowlozsky TT, Kramer PJ, Pallardy SG (1991) The physiological ecology of woody plants: carbon dioxide (chapter 10). Academic, New York
Loats KV, Rebbeck J (1999) Interactive effects of ozone and elevated CO2 on the growth and physiology of black cherry, green ash and yellow poplar seedlings. Environ Pollut 106:237–248
Malaysian Palm Oil Board (MPOB) (2007) Oil palm statistics-2007. MPOB Publication, Kuala Lumpur
Pritchard SG, Hugo HR, Stephen A, Curt MP (1999) Elevated CO2 and a plant structure: a review. Glob Change Biol 5:807–837
Ranasinghe S, Taylor G (1996) Mechanism for increased leaf growth in elevated carbon dioxide. J Exp Bot 47:349–358
Tanner CB, Sinclair TR (1983) Efficient water use in crop production research. In: Taylor HM, Jordan WR, Sinclair TR (eds) Limitation to efficient water use in crop production. ASA/CSSA/SSSA, Madison, pp 1–28
Taylor G, Ranasinghe S, Basac C, Gardener SDL, Ferris R (1994) Elevated CO2 and plant growth: cellular mechanism and responses of whole plants. J Exp Bot 45:1761–1774
Vu JCV, Allen LH, Gesch RW (2006) Up-regulation of photosynthesis and sucrose metabolism enzymes in young expanding leaves of sugarcane under elevated CO2. Plant Sci 171:123–131
Acknowledgments
The authors wish to acknowledge the financial support from Malaysia Palm Oil Board (MPOB) for sponsoring the research.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Z. Gombos.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Ibrahim, M.H., Jaafar, H.Z.E., Harun, M.H. et al. Changes in growth and photosynthetic patterns of oil palm (Elaeis guineensis Jacq.) seedlings exposed to short-term CO2 enrichment in a closed top chamber. Acta Physiol Plant 32, 305–313 (2010). https://doi.org/10.1007/s11738-009-0408-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11738-009-0408-y