
Abstract
Effective breeding requires multiplying desired genotypes, keeping them at a convenient location to perform crosses more efficiently, and building orchards to generate material for reforestation. While some of these aims can be achieved by conventional grafting involving only rootstock and scion, topgrafting is known to deliver all in a shorter time span. In this study, Scots pine scions were grafted onto the upper and lower tree crowns in two clonal archives with the aim of inducing early female and male strobili production, respectively. Their survival rates and strobili production were analyzed with generalized linear mixed models. Survival was low (14%) to moderate (41%), and mainly affected by the topgraft genotype, interstock genotype, crown position and weather conditions in connection with the grafting procedure. Survival was not affected by the cardinal position in the crown (south or north). Male flowering was ample three years after grafting and reached 56% in the first year among live scions, increasing to 62 and 59% in consecutive years. Female flowering was scarce and was 9% at first, later increasing to 26 and 20% of living scions but was strongly affected by the topgraft genotype. In one subset of scions, female flowering was observed 1 year after grafting. Overall, flowering success was mainly affected by the topgraft and interstock genotypes, and secondary growth of scions. This is one of few reports on topgrafting in functional Scots pine clonal archives.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Predictable quality of tree crops can be achieved by deployment of tested material in seed orchards (Kroon et al 2011). Information from the tested material also allows for the assessment of selection effectiveness and realization of long-achieved genetic gain (Danusevičius and Lindgren 2004). Testing of selected phenotypes occurs through assessment of field performance of progeny from controlled crosses, but to cross plants in a controlled and well-organized way, all genotypes should be available within one location (Kang and Bilir 2021). Scions also need to come from larger, ontogenetically mature trees to be able to produce flowers (Ahuja and Libby 1993). However, the first flowering years can be highly variable in the number of flowers (Nikkanen and Ruotsalainen 2000). Thus, phenotypic selection for new accessions and their testing requires convenient access to genotypes and methods that reduce the flowering time of mature material.
Grafting is an ancient technique of vegetative plant propagation that originated in horticulture and was adapted to conifers (Jinks 1999; Mudge et al 2009). By assembling different genotypes together, grafting reduces the maintenance costs and logistics, and modifies production (Miller and DeBell 2013; Loewe-Muñoz et al 2022). Topgrafting, (also known as top-working), is a variation of a grafting technique where scions are grafted onto a combination of interstock and rootstock that have different genotypes, as opposed to normal grafting, consisting of two genotypes—rootstock and a scion. The resulting union consists of three parts—rootstock, interstock, and scion, each representing a separate genotype (Almqvist and Ekberg 2001).
The advantage of topgrafting is the shortened time to first flowering—ranging from 1 to 4 years after the grafting procedure (Greenwood and Gladstone 1978; Bramlett and Burris 1995). Reproductive competence of mature interstock is transferred via flowering signals (genes related to flowering initiation that are already expressed in the tissues of the interstock) to scion genotypes that carry desirable traits (Ma et al 2022) which promote early flowering, even in juvenile scions (Almqvist 2001). At the same time, more traditional grafting with only two genotypes does not guarantee a such shortening in generation time (Simak 1978), mainly due to the much smaller size of rootstocks (Philipson 1987).
Thus, topgrafting not only combines, multiplies and archives valuable genotypes but also overcomes the long juvenile phase so that sufficient yearly flowering of desired genotypes is ensured within a reduced time (Bramlett 1997; Miller and DeBell 2013). Currently, topgrafting is a productive option in advanced breeding strategies that contributes to the progress of tree breeding programs worldwide (Harfouche et al 2012). Examples include the radiata pine (Pinus radiata D. Don) breeding program in New Zealand and the loblolly pine (Pinus taeda L.) breeding program in the southeastern US (Li and Dungey 2018; Isik and McKeand 2019). It can be used both at the planning stage to create new clonal archives and seed orchards, and to preserve tree genotypes economically, thus allowing great flexibility of breeding (Zobel and Talbert 1984).
However, in breeding programs at the northern parts of a species’ distribution range, topgrafting is also used for flowering induction on ontogenetically older scion material. In the Scots pine (Pinus sylvestris L.) breeding program in Sweden, normal grafts reduce the breeding cycle but still require 10–15 years until first flowering (Kroon et al 2009). Topgrafting have produced the first female flowers within a much shorter time than normal (Almqvist 2013b). However, experience in topgrafting has shown that vital grafts are only produced with a limited number of genotype combinations. This indicates the possible influence of interstock genotype on grafting success (Melchior 1984; Schmidtling 1991), for example, of loblolly pine (McKeand and Raley 2000), slash pine (Pinus elliottii Engelm.) (Medina Perez et al 2007) and Scots pine (Almqvist and Ekberg 2001; Almqvist 2013b). Topgrafting has generally been unsuccessful in inducing early flowering in species like Norway spruce, even when interstocks flowered regularly (Almqvist 2013a).
The importance of studying the interstock effect is due to its influence on the topgraft—scion genotypes for a particular breeding program which may change, depending on the breeding objectives, but the interstock part forms the crown and remains as part of the tree despite these changes (Kang and Bilir 2021). Therefore, interstock interactions may pose a serious bottleneck in the process of breeding and conclusive outcomes of experiments. The extent of these effects and their interactions determine the prerequisite for building an archive with a broad, combining capacity for many breeding programs. So far, topgrafting reports on Scots pine are scarce and best practice is still inconclusive as to what the main factors that determine topgrafting success.
The aim of this study was to determine the role of interstock genotypes in topgrafting success for Scots pine in current clonal archives in northern Sweden. The particular focus was to understand whether interstock genotype governs the outcome of topgrafting success solely or in combination, and potentially—interaction, with the topgraft genotype. Additionally, we aimed to assess whether there was a difference in survival rate and type of flowers produced between crown positions and geographic orientation within the crown. To answer these questions, topgraft vitality and strobili production was assessed in two clonal archives with relation to topgrafting, interstock genotypes, geographic position, and compared topgraft vitality and flowering between the two archives where scions were topgrafted in different parts of the crown.
Materials and methods
Topgraft archives
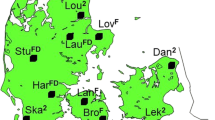
The trees for grafting were located in two functioning clonal archives that belong to the currently running breeding program, located in Sävar, northern Sweden (63°89' N 20°55'' E) and referred to as the male and female archives. In the female archive, topgrafts were made on upper crowns of small (≈ 1.5 m) grafted trees established in 2013 with the aim to promote female flowering. In the male archive, topgrafts were made in relatively large (≈5–6 m) grafted trees in a clonal breeding archive established in 1992 and made in the lower crown with a high presence of male flowers. Interstock donor trees in the female archive were more than 150 years old at the time of scion collection and 50–80 years old in the male archive. In both archives, interstock donor genotypes originated from northern parts of Sweden (Fig. 1; Table S1). One interstock genotype in the female archive belonged to a congener species–bog pine (Pinus mugo Turra). There were no records on the rootstock genotypes. Topgraft donor trees were either young (≈ 20 years; grafted in 2020 in the female archive) or mature (≈ 45 years; grafted in both archives in 2017) at the time of scion collection.

Map of interstock genotype origin
Topgrafting of scions in the female archive was carried out in 2017 and 2020. In 2017, 816 scions belonging to 47 unique topgraft genotypes were grafted on to 21 unique interstock genotypes. In 2020, 405 scions from 133 new unique topgraft genotypes were grafted into this archive on 11 interstock genotypes that had the highest survival in the previous grafting procedure. In this archive, the topgrafts with interstock genotypes were randomly distributed among 22 plots, each consisting of four rows with five trees per row within six blocks, with each interstock genotype replicated five times.
Experimental design in the grafting archives
Grafting in the male archive was performed in 2017 and 45 unique topgraft genotypes were grafted on to 24 unique interstock genotypes. In this archive, the interstock genotypes were planted in one row with 2–3 ramets of the interstock genotype and without any other allocated design effects.
The same 45 topgraft genotypes were used in both archives in 2017 (Fig. 2), and the new set of topgraft genotypes used for grafting in 2020. Topgraft genotypes were randomized in pairs on three different interstocks in the female archive and two different interstocks in the male archive. Each topgraft genotype was represented within a tree crown with three scions (except for four genotypes grafted in 2020 represented by one scion). Trees in both male and female archives were pruned.

Venn diagram describing the experiment setup
Data processing
The topgrafts from 2017 were evaluated yearly from 2019 to 2022, 2–5 years after grafting. In 2019 only survival was recorded. The topgrafts from 2020 were assessed in 2021 and 2022, 1–2 years after grafting.
The following variables were recorded: survival of scions (binary: 0–dead, 1–alive), the number of male or female flowers per scion, a binary variable—flowering success (binary: 0–no flowering, 1–flowering), the total number of flowers of any type per scion and grafting side within the crown in the male archive (orientation either south or north part of the crown). The frequency of successful flowering of the interstock was recorded in the female archive as a binary variable (“ + ” for successful flowering and “–” for unsuccessful) for all blocks and was also recorded as a number of cones in one block.
The data were divided into subsets according to the type of the archive, year of grafting and year of assessment (Table 1). Analysis was performed on subsets C, F, H and J (which corresponds to the subset “common topgrafts” mentioned in the Fig. 2), and data from the subsets A–I were summarized in Figs. 3, 4, 6, 7 and 8. To enable analyses of the binary data and prevent singularities, datasets were pruned to remove interstock and topgraft genotypes that only produced dead scions.

Survival and flowering results in the female archive. Survival is represented by the number of scions surviving in the female archive in 2019–2022 as bars (light grey for scions grafted in 2017, dark grey for scions grafted in 2020). Flowering is represented as the number of scions that produced female strobili in the archive in 2020–2022 and depicted as lines. Numbers on the bottom of the bars represent the percent of scions surviving in the female archive to the total number of grafted scions

Survival and flowering results in the male archive. Survival is represented by the number of scions surviving in the male archive in the north and south sides of the archive in 2019–2022 as bars. Flowering is the number of scions that produced male strobili in the male archive, north and south side in 2020–2022 and depicted as lines. Numbers on the bars represent the percent of surviving scions in the male archive to the total number of grafted scions, irrespective of the geographic side
Survival of scions, number of male/female flowers, total number of flowers, flowering success were the response variables and analyzed using a generalized linear mixed model (GLMMs). The following models were fitted with mixed (1) or random effects (2) to analyze survival of topgrafts in the subsets C, F, H, J:
where, yit is the dependent value (topgraft survival); µ is the overall mean; bi is the main effect of the interstock genotype (i = 1,.., 21); ct is the random effect of the topgraft genotype (t = 1, …, 47); eit is the error term. *For the subset J, model 1) was extended with the model term f, which is the main effect of side (north or south side in the male archive, or “none” in the female archive), and applied in models with topgraft survival and total number of flowers as response variables.
where, yit is the dependent value (topgraft survival, number of male/female flowers); bi is the random effect of the interstock genotype (I = 1,.., 21; I = 11); ct is the random effect of the topgraft genotype (t = 1, …, 47; t = 1, …, 133).
Models were specified by applying binomial models with binary variables as response variables, or Poisson distribution models with categorical variables as response variables, the logit or log-link function, increased number of iterations, and “bobyqa” optimizer within the model specification. Significance of random effects was verified via a likelihood ratio test between a model with the random effects and a model without. All modelling procedures were performed in R package “lme4” (Bates et al 2015) and performed in R studio v 4.2.3 (R Development Core Team 2024).
Results
Topgraft survival
The model with the interstock genotype as a main effect showed that in the subsets C and F several interstock genotypes had a significant negative effect on topgraft survival (Figs. S1 and S2). In the subset H, several interstock genotypes had a significant effect on survival of topgrafts and one genotype had a positive effect, compared to the subset F. Effect sizes are summarized for all subsets in Tables S4, S5, S6 and Figs. S1 and S2.
A model fitted with only random effects showed that survival of scions in 2022 depended on genotypes of interstock and topgraft almost equally in the subsets C and H, mostly on interstock in the subset F, and mostly on topgraft genotype in the subset J (Table 2). According to the model summary of a mixed model for the subset J, topgraft genotype explained three times more variance (Table 3). Fixed effects of this mixed model detected no significant improvement of survival of scions located in the south side of the crown. The opposite side of the crown, the north side, negatively affected the survival of scions (Table 3).
Overall, mortality was high but the dynamics of the two archives was different. Survival of scions in the female archive grafted in 2017 was low but barely decreased by the end of the experiment (Fig. 3). Among all 816 scions grafted that year, 14.6% (or 119) survived in 2019, 14.3% in 2020 (SD = 0.98), 14.0% in 2021 (SD = 1.03), 13.9% in 2022 (5 years after grafting; SD = 1.03). Survival among the 405 scions added in 2020 was higher (Fig. 3): 41.7% (169 scions; SD = 1.37) survived in 2021 and 41.2% in 2022 (2 years after grafting; SD = 1.47).
Survival of scions in the male archive was low at the beginning of the observations and continued to drop throughout the period of recording (Fig. 4). Of the 516 grafted scions, 86 scions (or 16.7%) survived in 2019 (2 years after grafting), 12.4% in 2020 (SD = 0.81), 9.7% in 2021 (SD = 0.67), and 9.1% in 2022 (5 years after grafting; SD = 0.83).
Low survival limited the possibilities to fit models with the main effects of both interstock and topgraft genotypes, and it was not possible to assess interactions between the genotypes directly. Survival was greater in the second grafting procedure, but the number of newly introduced topgraft genotypes was too high to assess possible interactions.
Topgraft strobili production
Low survival and scarce flowering in some subsets did not allow direct estimation of the interaction between topgrafted and interstock genotypes, nor the estimation of main effect models. Output from the random effect models assessing two different flowering-related variables exhibited similar partitioning of variance: for subsets F and H, variance explained by interstock genotype was almost 10 times greater than those explained by topgraft genotype in both binomial and Poisson-based models. A similar trend appeared for subset J where the interstock genotype variance explained the majority of the random effects part, although the difference between variances of random effects was smaller in the Poisson model. Differences emerged for variance explained by interstock genotype in the subset C in binomial and Poisson-based models, where the former showed greater variance explained by interstock genotype, but the latter had similar variances for both random effects. The mixed model fitted to subset J also showed similar variances for both random effects. The results of a likelihood-ratio test determined that random effects for the subset J were significant in models assessing flowering because their removal led to a model with less explanatory power.
Scions grafted in 2017 in both archives started producing flowers in 2020, three years after grafting, but the scale was considerably different between female and male archives (Figs. 5, 6 and 7). Scions grafted in the female archive in 2020 started producing flowers in 2021, one year after grafting.

Survival and strobili production of topgrafts on different interstocks in the male archive. The percentage of surviving topgraft genotypes per interstock genotype is on the y-axis and depicted with three bars in descending order—percentage of survived topgraft genotypes in 2020, 2021 and 2022. Strobili production is represented as three graph lines corresponding to percentage of flowered topgraft genotypes on interstock genotype in 2020, 2021 and 2022. Interstock genotypes that were associated with only dead topgrafts are omitted

Survival and strobili production of topgrafts on different interstocks in the female archive in 2017. The percentage of surviving topgraft genotypes per interstock genotype is represented on the y-axis and depicted with three bars in descending order—percentage of surviving topgraft genotypes in 2020–2022, respectively. Strobili production is represented as three graph lines corresponding to percentage of flowered topgraft genotypes on interstock genotypes in 2020–2022, respectively. Interstock genotypes associated with only dead topgrafts are omitted

Survival and strobili production of topgrafts grafted on different interstocks in the female archive in 2020. The percentage of surviving topgraft genotypes per interstock genotype is represented on the y-axis and depicted by two bars in descending order—percentage of survived topgraft genotypes in 2021 and 2022. Strobili production is represented as two graph lines corresponding to percentage of flowered topgraft genotypes on interstock genotypes in 2021 and 2022. Interstock genotypes associated with only dead topgrafts are omitted
In the subset of scions grafted into the female archive in 2017, 11 out of 117 living topgrafts (9.4%) flowered and produced 18 flowers three years after grafting in 2020 (Fig. 6). By 2021, four years after grafting, 30 scions out of 114 living topgrafts (26.3%) produced 80 flowers. In 2022, five years after grafting, 23 scions out of 113 living topgrafts (20.3%) produced 101 flowers. Interstock flowering was regular for most of the grafts, although the differences in the cone number suggest yearly dynamics, based on cone counts in block 1, and show that the least number of cones on the interstock was produced mainly in 2020. The average number of strobili per scion increased each year: 1.63 in 2020, 2.67 in 2021, and 4.39 in 2022. The maximum number of strobili per scion was 4.0 in 2020 and 16.0 in both 2021 and 2022. Male flowers produced by the scions in this subset were 3.0 in 2020, 18.0 in 2021, and 149.0 in 2022.
Among the 169 scions grafted in 2020 that survived, five (2.9%) flowered and produced seven female flowers in 2021, one year after grafting. In 2022, nine scions (5.4%) flowered and produced 14 female flowers. Flowering scions were associated with three interstock genotypes (Fig. 7). Both the average and the maximum number of strobili per scion increased slightly in this subset by 2022, two years after grafting. The average number of strobili per scion started from 1.4 in 2021 and was 1.55 in 2022; the maximum number per scion was two in 2021 and three in 2022. Male flowers produced by the scions in this subset were 30 in 2021 and 47 in 2022.
For the male archive in 2020, three years after grafting, 36 live topgrafts (56.2%) flowered out of 64 surviving and produced 77 strobili (Fig. 5). This number changed to 32 topgrafts (62.7%) that produced 77 strobili four years after grafting in 2021, and later to 28 flowering scions (59.6%) that produced 100 male strobili in 2022. Almost all topgraft and interstock genotypes that produced flowers in 2020 flowered in 2021 and 2022. The average number of pollen clusters per topgraft was 2.13 in 2020, three years after grafting, 2.41 pollen clusters in 2021 (four years after grafting), and 3.57 five years after grafting in 2022. The maximum number of pollen clusters per topgraft was seven, both in 2020 (three years after grafting) and 2021, and 14 pollen clusters in 2022 (five years after grafting). Female flowers produced by the scions in this subset were three in 2020, 18 in 2021, and 14 in 2022.
Discussion and conclusion
Survival
Grafting success in conifers is usually moderate to high (Pérez-Luna et al 2020; Świerczyński et al 2020), and low rates similar to our results, (especially in the subsets B, C, F), are unusual. These results could be attributed to both factors considered in the study (scion position and orientation within the tree crown) as well as those not considered. Success rates may depend on the grafting season (Pérez-Luna et al 2019; Lawson 2020), particular the grafting technique (Barrera-Ramírez et al. 2021), age of donor plants (Pérez-Luna et al 2019; Velisevich et al 2021), pre- and post-grafting status of plants and management of the new grafts (McKeand and Jett 2000; Larson 2006), and genetic affinity (Schmidtling 1983; Darikova et al. 2011).
Success rates are also expected to be affected by the interactions (Jayawickrama et al 1997; Almqvist and Ekberg 2001) or incompatibility (Darikova et al. 2011; Susilowati et al 2016) between the genotypes involved, which unfortunately, due to the low survival did not allow for testing. It is of considerable practical significance to select interstock genotypes with a broad ability to form stable union of tissues with various scion genotypes and to secure stability and flexibility of the breeding efforts. Using the topgrafts with some degree of genetic relation to interstocks might be helpful (Copes 1973).
Topgrafting, (as with any other grafting technique), relies on functioning biochemical communication between genotypes (Fernández de Simón et al 2021; Feng et al 2024), which infers the possibility of interactions not only between interstock and topgraft, but also with the rootstock (Jayawickrama et al 1997). A review of grafting techniques in loblolly pine suggests that resistance of the rootstock genotype to local pathogens is essentially the only genetic criterion of serious consideration (McKeand and Jett 2000; Larson 2006). This conclusion may reflect the importance of the ability of the rootstock genotype to withstand external stressors and thus contribute to the health of a grafted tree, as has been shown in Norway spruce and loblolly pine grafting studies (Schmidtling 1983; Melchior 1984). In our experiment, there were no records on rootstocks; further studies would benefit from keeping such records and taking potential effects into consideration.
The difference in survival rates between the subsets F (13.8%) and H (41.2%), C (9.1%) and F could be attributed to weather conditions and to repeated grafting with only the best performing interstocks. Harsh weather after grafting in early spring 2017 slowed down growth initiation and could have reduced the opportunity for quick appearance of the contact layer and successful tissue reconnection, important steps in callus formation and restoring the flow of metabolites (Weatherhead and Barnett 1986).
Second, interstock genotypes in the female archive that only produced dead topgrafts after grafting in 2017 (1176, 1195, 3028) were excluded in 2020, and the number of new unique topgraft genotypes was expanded. Testing more topgraft genotypes on interstocks to detect the most productive ones is the common recommendation (Almqvist and Ekberg 2001; Medina Perez et al 2007). The increase in survival from 14 to 41% (Fig. 3) verifies the importance of interstock genotype and the utility of this strategy, although we expect weather to have had the largest impact.
The discrepancy in survival between the subsets C and F can arise partly due to the placement within the crown. The top position is an important factor for survival, as established in previous grafting experiments, which suggests that the middle to high crown areas have a better chance for growth because interstock branches are thinner and do not differ substantially in size from scions (Medina Perez et al 2007). Simak (1978) found that crown position and the presence or absence of the top shoot created different outcomes on survival and growth of scions of Scots pine. Lower positions in the crown are also more vulnerable to disturbance from equipment (ladders, lifts), and there is a tendency for pines to lose lower branches with age due to lower hydraulic properties (Protz et al 2000), hence the lower scion survival in this part of the crown. While the scions in the female archive were mainly in the upper crown, the trees were not tall, and the crowns were not well developed in the beginning of the experiment in 2017, so the branches were of comparable size with the grafts and allowed the scions to form a stable mechanical union with their interstock.
Additionally, such properties of interstock genotypes as latitudinal origin may have played a role in survival (Karlsson and Woods 1992). Many of the previous grafting studies with Scots pine used provenances with stocks originating from milder climates (south-eastern and southern Sweden and Germany) (Dormling 1962; Simak 1978; Almqvist and Ekberg 2001; Almqvist 2013b). There are clear differences in the growth rhythms between southern and northern Scots pine populations in Sweden, where the latter grow slower and shorter than their southern counterparts (Oleksyn et al 1999; Andersson Gull et al 2018). When transferred south to milder locations, northern genotypes exhibit enhanced scion survival due to earlier dehardening and the onset of growth further exaggerated by the upper crown position. Anatomical studies established that the differences in such traits as total resin duct area, or density of resin ducts, may have a key significance for robust tissue connection (Darikova et al. 2011; Susilowati et al 2016; Castro-Garibay et al 2023), and that stock re-growth is usually rapid so branches selected for grafting should be smaller than the scion (Dormling 1962). Consequently, the top position within the crowns of early dehardening and slow growing northern clones may have created an advantage for survival.
From a practical perspective, it might be useful to test different tying and/or sealing materials in combination with various grafting techniques, particularly those that allow deeper insertion of the scion into the interstock, to find those that prevent desiccation but also provide more support to the fragile union between the scion and interstock (Humphrey 2019), and apply treatments such as heat (Ho 1991). Since both archives studied are included in current breeding activities, there were limited possibilities to manipulate the experimental design, and some of the noted factors are surprising (crown position, tree size, origin of interstock genotypes and type of archive, age of interstock donor and type of archive). For more conclusive results, further research is needed.
Finally, while the binary values of vitality were used to increase the number of useful observations, they did not distinguish the most vigorous scions from less healthy ones, and there is still a degree of oversimplification in assessments. By the end of the field observations in July 2022, the most vital topgrafts in the female archive developed into vigorous branches within the grafted tree crowns, with abundant, healthy foliage, and some were almost unproportionally large and easily seen from a distance; whereas many of scions in the male archive were still short, had scarce foliage and rough, poorly healed grafting scars.
Flowering
Despite the scarcity of data, some general patterns were observed, namely the discrepancy between the extent of flowering in the male and female archives, the induction of flowering one year after grafting in subset I, and more consistent contribution to flower production in some topgraft genotypes in subsets I and J. Ample male flowering aligns well with the results of earlier studies, but the stark discrepancy between male and female archive flowering is uncommon. In general, grafting experiments gain moderate to high results in terms of induction of both female and male flowers (Gooding et al 1999; Lott et al 2003; Almqvist 2013c) and reduce the time to reproduction (Loewe-Muñoz et al 2022).
In our experiment, the extent of variation among genotypes is greater than yearly variation, especially in the female archive: some interstock genotypes were responsible for most of the flowering within their respective archive every year, whereas the majority induced strobili on their scions only occasionally (Figs. 5, 6, 7). These observations mirror previous results that revealed substantial genetic variation of flowering behavior (in terms of number and sex ratio of produced flowers) in the genus Pinus (Schmidtling 1983; Ying et al 1985) and in Scots pine in particular (Bilir et al 2006, 2008). It should also be noted that the observation period was rather short (three to five years) and a long-term study may clarify which factor prevails.
Some topgraft genotypes in our experiment were the vigorous and prolific in producing both male and female flowers (Fig. 8). Similar variances in subset J may indicate that the role of the topgraft genotype is important in the number of flowers produced. Almqvist and Ekberg (2001) and Almqvist (2013b) encountered an effect of genotype interaction on flowering in Scots pine, where the interaction was significant for female flowering but not for male flowering Their conclusions might help to explain ample flowering in the subset C (male archive) and scant output of all subsets from the female archive. Due to generally low survival and scarce flowering in the female archive, we were unable to test for the main effect of the topgraft genotype and the interaction between genotypes, and statistical results to verify or disprove its presence were lacking. Indirect support for potential interactions may be the successful interstock-topgraft combinations that produced flowers in 2020 also flowered in 2021 and 2022 (Figs. 5 and 6).

Relationships between survival and flowering of 45 common topgraft genotypes grafted in 2017 in the male (a) and female (b) archives as a cumulative sum over three years of observation
The relationships summarized in Fig. 8 might also indicate that female flower production is more resource demanding, thus a greater cumulative sum of vitality was associated with prolific female flowering. Experimental excision of developing female flowers in grafted Aleppo pine (Pinus halepensis Mill.) caused an increase in female flowering the year after, suggesting a reproductive cost for scions (Santos‐del‐Blanco and Climent 2014). Flowering initiates when available resources are sufficient, and yearly flower production would keep them continuously at a low level while extreme weather such as drought would further reduce resource allocation to reproduction (Fober 1976; Mutke et al 2005a; He et al 2018). All of these, combined with the influence of crown position in topgrafting, could help explain the greater vigour and scarcer flowering in the female archive in our study. The poor weather conditions following grafting in 2017, together with an unusually hot and dry summer of 2018, likely affected bud initiation and limited female flowering in 2020. The small increase in female flowers produced in the male archive supports the conclusion of greater cost of female flowers, compared to the large increase in male flowers produced in the female archive, which rose to 149 by the end of the observation period.
An alternative explanation to the discrepancy between male and female flowering may be an age-specific (or species-specific) sex ratio of scions (Ahuja and Libby 1993) and tree size (Greenwood et al 2010). Size of the grafted tree, its crown width and number of branches were positively correlated with flowering in a Scots pine study in Finland (Nikkanen and Velling 1987). These results might support the role of tree size from the female archive and that taller trees in the male archive had on flowering (Philipson 1987)—the latter flowered abundantly even when scions exhibited signs of incompatibility and had lower vitality. If greater resources for female flower production are indeed required, the interplay of crown position and tree size, topgraft genotype, and weather conditions may be possible factors shaping female flowering output.
In subset I, female flowers appeared one year after grafting, however the number of flowers and genotypes that flowered were very low. This suggests that, even if it were possible to stimulate the scion to flower earlier, a substantial degree of individual variation in female flowering will still be present, thus the least productive genotypes might benefit from additional flowering induction techniques and treatments (Bonnet-Masimbert 1987; Wheeler and Bramlett 1991; Almqvist 2018; Li et al 2021) to carry out the entire crossing scheme.
Properties of interstock genotypes (such as latitudinal and perhaps altitudinal origin, see Table S2) in this study may have had an impact on the outcome of grafting and flowering induction. Previous observations of the flowering behavior of interstocks and their capacity to induce flowering in the topgrafts did not find any correlation between the two (Almqvist and Ekberg 2001). Regular flowering of the interstocks and some fluctuations due to year were detected and, possibly, the interstock genotype. The practical advice is to not rely on interstock flowering as a sole criterion of choice.
Latitudinal origin of northern interstocks may have been advantageous in terms of survival, but flowering behavior is usually more complex (Crain and Cregg 2018), especially for female flower production. Experience from grafting in Scots pine shows that the seed set in ramets that originated from northern Sweden and were grafted in different parts of the country was highly variable and prediction of cone set was limited (Tellalov 2006). Pollen production in Scots pine also decrease with higher latitudes (Pessi and Pulkkinen 1994), suggesting that populations originating from the northern part of their range typically have fewer years of successful reproduction and experience heavier impacts of weather fluctuations. A study of Aleppo pine grafts showed that vegetative growth and reproductive behavior are very plastic, and individual variation in the studied traits is high (Pardos et al 2003), and it seems to be the tendency for pines (Bilir et al 2008) that further complicates predictions for seed crop production.
Crown architecture and position of scions within the crown are important for survival and define the type of flowers, regularity of flowering and bud initiation. Medina Perez et al. (Medina Perez et al 2007) also showed that grafting in the upper crown promoted female flowering, which is in agreement with our results and with other reports on conifer species (Bilir et al 2008). A study on crown architecture of grafted Stone pine (Pinus pinea L.) found only one significant correlation between the mother shoot formed the previous year and the number of cones formed three years earlier (Mutke et al 2005b). Swedish researchers found relationships between scion length and flowering in Scots pine (Almqvist and Ekberg 2001). This suggests that vigorous first-order branches with good growth create more opportunities for the development of female cones.
In conclusion, survival of scions is higher in the upper part of the crown and repeated grafting on the best interstock genotypes has a potential for improving survival sufficiently. Topgrafting can induce production of single female flowers one year after grafting and ample male flowering three years after grafting, but the extent of flowering is greatly impacted by genotypes, their interactions, and by weather. Large discrepancies between male and female flower production, great variability in flowering production among genotypes, and potential interactions may hinder execution of full crossing schemes. Supplementary flowering induction techniques can secure crop production, and further research should provide more conclusive results on the extent of interactions and the effects of orientation within the crown on survival and flowering.
References
Ahuja MR, Libby WJ (1993) Genetics, biotechnology and clonal forestry. In: Ahuja MR, Libby WJ (eds) Clonal forestry I. Springer Berlin Heidelberg, pp 1–4. https://doi.org/10.1007/978-3-642-84175-0_1
Almqvist C (2001) Improvement of flowering competence and capacity with reference to Swedish conifer breeding. Doctoral thesis, Swedish University of Agricultural Sciences
Almqvist C (2013a) Metoder för tidig blomning hos tall och gran. Skogforsk
Almqvist C (2013b) Interstock effects on topgraft vitality and strobili production after topgrafting in Pinus sylvestris. Can J for Res 43(6):584–588. https://doi.org/10.1139/cjfr-2012-0507
Almqvist C (2013c) Survival and strobili production in topgrafted scions from young Pinus sylvestrisseedlings. Scand J for Res 28(6):533–539. https://doi.org/10.1080/02827581.2013.803598
Almqvist C (2018) Improving floral initiation in potted Picea abies by supplemental light treatment. Silva Fenn. https://doi.org/10.14214/sf.7772
Almqvist C, Ekberg I (2001) Interstock and GA4/7 effects on flowering after topgrafting in Pinus sylvestris. For Genet 8(4):279–284
Andersson Gull B, Persson T, Fedorkov A, Mullin TJ (2018) Longitudinal differences in Scots pine shoot elongation. Silva Fennica. https://doi.org/10.14214/sf.10040
Barrera-Ramírez R, Vargas-Hernández JJ, López-Aguillón R, Muñoz-Flores HJ, Treviño-Garza EJ, Aguirre-Calderón OA (2021) Impact of external and internal factors on successful grafting of Pinus pseudostrobus var. oaxacana (Mirov) Harrison. Revista Chapingo Serie Ciencias Forestales y Del Ambiente 27(2):243–256
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models Using lme4. J Stat Soft 67(1):1–48
Bilir N, Prescher F, Ayan S, Lindgren D (2006) Growth characters and number of strobili in clonal seed orchards of Pinus sylvestris. Euphytica 152(2):1–9. https://doi.org/10.1007/s10681-006-9216-2
Bilir N, Prescher F, Lindgren D, Kroon J (2008) Variation in cone and seed characters in clonal seed orchards of Pinus sylvestris. New for 36(2):187–199. https://doi.org/10.1007/s11056-008-9092-9
Bonnet-Masimbert M (1987) Floral induction in conifers: a review of available techniques. For Ecol Manag 19(1–4):135–146. https://doi.org/10.1016/0378-1127(87)90019-3
Bramlett DL (1997) Southeastern conifers: genetic gain from mass controlled pollination and topworking. J for 95(3):15–19. https://doi.org/10.1093/jof/95.3.15
Bramlett D, Burris L (1995) Topworking young scions into reproductively-mature loblolly pine. In: Proceedings of the 23rd Southern Forest Tree Improvement Conference. pp 20–22
Castro-Garibay SL, Terrazas T, López-Upton J, Arévalo-Galarza L, Sandoval-Villa M, Villegas-Monter Á (2023) Anatomical traits in pine stems, the first step to define grafting combinations? Braz J Bot 46(1):153–161. https://doi.org/10.1007/s40415-022-00851-y
Copes DL (1973) Inheritance of graft compatibility in Douglas-fir. Bot Gaz 134(1):49–52. https://doi.org/10.1086/336679
Crain BA, Cregg BM (2018) Regulation and management of cone induction in temperate conifers. For Sci 64(1):82–101. https://doi.org/10.5849/FS-20l6-131
Danusevičius D, Lindgren D (2004) Progeny testing preceded by phenotypic pre-selection–timing considerations. Silvae Genet 53(1–6):20–26. https://doi.org/10.1515/sg-2004-0004
Darikova JA, Savva YV, Vaganov EA, Grachev AM, Kuznetsova GV (2011) Grafts of woody plants and the problem of incompatibility between scion and rootstock (a review). J Sib Fed Univ Biol 1:54–63
Development Core Team R (2024) R: A language and environment for statisical computing
Dormling I (1962) Ympningsmetoder för tall och gran. Medelanden från Statens Skogsforskningsinstitut 51. https://pub.epsilon.slu.se/10048/1/medd_statens_skogsforskningsinst_051_02.pdf
Feng M, Augstein F, Kareem A, Melnyk CW (2024) Plant grafting: molecular mechanisms and applications. Mol Plant 17(1):75–91. https://doi.org/10.1016/j.molp.2023.12.006
Fernández de Simón B, Aranda I, López-Hinojosa M, Miguel L, Cervera MT (2021) Scion-rootstock interaction and drought systemic effect modulate the organ-specific terpene profiles in grafted Pinus pinaster Ait. Environ Exp Bot 186:104437. https://doi.org/10.1016/j.envexpbot.2021.104437
Fober H (1976) Relation between climatic factors and scots pine (Pinus silvestris L.) cone crops in Poland. Arboretum Kórnickie 21:367–374
Gooding G, Bridgwater FE, Bramlett DL, Lowe WJ (1999) Top grafting loblolly pine in the western gulf region. In: Proceedings of the 25th Biennial Southern Forest Tree Improvement Conference. New Orleans, Louisiana, USA. 60–66
Greenwood MS, Gladstone WT (1978) Topworking loblolly pine for precocious flowering. Forest Res Tech Rep 42(3004):78
Greenwood MS, Day ME, Schatz J (2010) Separating the effects of tree size and meristem maturation on shoot development of grafted scions of red spruce (Picea rubens Sarg.). Tree Physiol 30(4):459–468
Harfouche A, Meilan R, Kirst M, Morgante M, Boerjan W, Sabatti M, Scarascia Mugnozza G (2012) Accelerating the domestication of forest trees in a changing world. Trends Plant Sci 17(2):64–72. https://doi.org/10.1016/j.tplants.2011.11.005
He P, Fontana S, Sui X, Gessler A, Schaub M, Rigling A, Jiang Y, Li MH (2018) Scale dependent responses of pine reproductive traits to experimental and natural precipitation gradients. Environ Exp Bot 156:62–73. https://doi.org/10.1016/j.envexpbot.2018.08.028
Ho RH (1991) Promotion of cone production in potted black-spruce grafts, using gibberellins, heat-treatment and root-pruning. For Ecol Manag 40(3–4):261–269. https://doi.org/10.1016/0378-1127(91)90044-V
Humphrey BE (2019) The bench grafter’s handbook: principles and practice. CRC Press. https://doi.org/10.1201/9781315171463
Isik F, McKeand SE (2019) Fourth cycle breeding and testing strategy for Pinus taeda in the NC state university cooperative tree improvement program. Tree Genet Genomes 15(5):70. https://doi.org/10.1007/s11295-019-1377-y
Jayawickrama KJ, McKeand SE, Jett JB (1997) Rootstock effects on scion growth and reproduction in 8-year-old grafted loblolly pine. Can J for Res 27(11):1781–1787. https://doi.org/10.1139/x97-152
Jinks RL (1999) Propagation of conifers. pp 303–307
Kang K, Bilir N (2021) Seed orchards (establishment, management and genetics). CRN Promotion and Press, Ankara
Karlsson I, Woods JH (1992) Effects of rootstock source on the size, cone production, and compatibility of Douglas-fir grafts. West J Appl for 7(3):73–77. https://doi.org/10.1093/wjaf/7.3.73
Kroon J, Wennström U, Prescher F, Lindgren D, Mullin TJ (2009) Estimation of clonal variation in seed cone production over time in a Scots pine (Pinus sylvestris L.) seed orchard. Silvae Genet 58(1–6):53–62
Kroon J, Ericsson T, Jansson G, Andersson B (2011) Patterns of genetic parameters for height in field genetic tests of Picea abies and Pinus sylvestris in Sweden. Tree Genet Genomes 7(6):1099–1111. https://doi.org/10.1007/s11295-011-0398-y
Larson RA (2006) Grafting: a review of basics as well as special problems associated with conifer grafting. In: Combin Proc Int Plant Propagat Soc 56:318–322
Lawson KM (2020) Selection and propagation of pinyon pine. Doctoral thesis, Utah State University
Li YJ, Dungey HS (2018) Expected benefit of genomic selection over forward selection in conifer breeding and deployment. PLoS ONE 13(12):e0208232. https://doi.org/10.1371/journal.pone.0208232
Li Y, Li X, Zhao MH, Pang ZY, Wei JT, Tigabu M, Chiang VL, Sederoff H, Sederoff R, Zhao XY (2021) An overview of the practices and management methods for enhancing seed production in conifer plantations for commercial use. Horticulturae 7(8):252. https://doi.org/10.3390/horticulturae7080252
Loewe-Muñoz V, Del Río R, Delard C, Balzarini M (2022) Enhancing Pinus pinea cone production by grafting in a non-native habitat. New for 53(1):37–55. https://doi.org/10.1007/s11056-021-09842-5
Lott L, Lott L, Stine M, Kubisiak TL, Nelson CD (2003) Top grafting longleaf×slash pine F1 hybrids on mature longleaf and slash pine interstocks. In: Proceedings of the 27th Southern Forest Tree Improvement Conference
Ma JJ, Chen X, Han FX, Song YT, Zhou B, Nie YM, Li Y, Niu SH (2022) The long road to bloom in conifers. For Res 2(1). https://doi.org/10.48130/fr-2022-0016
McKeand S, Jett J (2000) Grafting loblolly pine. Bulletin of the American Conifer Society 22–30
McKeand S, Raley EM (2000) Interstock effects on strobilus initiation in topgrafted loblolly pine. For Genet 7:179–182
Medina Perez AM, White TL, Huber DA, Martin TA (2007) Graft survival and promotion of female and male strobili by topgrafting in a third-cycle slash pine (Pinus elliottii var. elliottii) breeding program. Can J for Res 37(7):1244–1252
Melchior G (1984) The influence of defined rootstocks on grafts of Norway spruce (Picea abies L. Karst). Silvae Genet 33:28–32
Miller LK, DeBell J (2013) Current seed orchard techniques and innovations. National proceedings: forest and conservation nursery associations—2012 USDA Forest Service, Rocky Mountain Research Station, Fort Collins, Colorado, Proceedings RMRS-P-69 80–86
Mudge K, Janick J, Scofield S, Goldschmidt EE (2009) A history of grafting. Hortic Rev 35:437–493
Mutke S, Gordo J, Gil L (2005a) Variability of Mediterranean Stone pine cone production: yield loss as response to climate change. Agric for Meteor 132(3–4):263–272. https://doi.org/10.1016/j.agrformet.2005.08.002
Mutke S, Sievänen R, Nikinmaa E, Perttunen J, Gil L (2005b) Crown architecture of grafted Stone pine (Pinus pinea L.): shoot growth and bud differentiation. Trees 19(1):15–25
Nikkanen T, Ruotsalainen S (2000) Variation in flowering abundance and its impact on the genetic diversity of the seed crop in a Norway spruce seed orchard. Silva Fenn 34(3):205–222
Nikkanen T, Velling P (1987) Correlations between flowering and some vegetative characteristics of grafts of Pinus sylvestris. For Ecol Manag 19(1–4):35–40. https://doi.org/10.1016/0378-1127(87)90009-0
Oleksyn J, Reich PB, Chalupka W, Tjoelker MG (1999) Differential above- and below-ground biomass accumulation of European Pinus sylvestris populations in a 12-year-old provenance experiment. Scand J for Res 14(1):7–17. https://doi.org/10.1080/02827589950152241
Pardos M, Climent J, Gil L, Pardos JA (2003) Shoot growth components and flowering phenology in grafted Pinus halepensis Mill. Trees 17(5):442–450. https://doi.org/10.1007/s00468-003-0259-x
Pérez-Luna A, Prieto-Ruíz JÁ, López-Upton J, Carrillo-Parra A, Wehenkel C, Chávez-Simental JA, Hernández-Díaz JC (2019) Some factors involved in the success of side veneer grafting of Pinus engelmannii carr. Forests 10(2):112. https://doi.org/10.3390/f10020112
Pérez-Luna A, Wehenkel C, Prieto-Ruíz JÁ, López-Upton J, Hernández-Díaz JC (2020) Survival of side grafts with scions from pure species Pinus engelmannii Carr. and the P. engelmannii×P. arizonica Engelm var. arizonica hybrid. PeerJ 8:e8468
Pessi AM, Pulkkinen P (1994) Temporal and spatial variation of airborne Scots pine (Pinus sylvestris) pollen. Grana 33(3):151–157. https://doi.org/10.1080/00173139409428992
Philipson JJ (1987) A review of coning and seed production in Picea sitchensis. Proc, Sect B Biol Sci 93(1–2):183–195. https://doi.org/10.1017/s0269727000006382
Protz CG, Silins U, Lieffers VJ (2000) Reduction in branch sapwood hydraulic permeability as a factor limiting survival of lower branches of lodgepole pine. Can J for Res 30(7):1088–1095. https://doi.org/10.1139/x00-054
Santos-del-Blanco L, Climent J (2014) Costs of female reproduction in a conifer tree: a whole-tree level assessment. J Ecol 102(5):1310–1317. https://doi.org/10.1111/1365-2745.12283
Schmidtling RC (1983) Rootstock influences flowering, growth, and survival of loblolly pine grafts. For Sci 29(1):117–124. https://doi.org/10.1093/forestscience/29.1.117
Schmidtling R (1991) Effect of rootstock and scion on flowering, growth and foliar nutrients of loblolly pine (Pinus taeda) grafts. Silva Fenn 25(4):215–227
Simak M (1978) Precocious flower induction in Pinus sylvestris by grafting. In: Proceedings, Flowering and Seed Development in Trees: A Symposium. Mississippi State University, pp 15–18
Susilowati A, Iswanto AH, Wahyudi I, Supriyanto S, Siregar IZ (2016) Morphological and anatomical evaluation of grafted Pinus merkusii. J Korean Wood Sci Technol 44(6):903–912. https://doi.org/10.5658/wood.2016.44.6.903
Świerczyński S, Kolasiński M, Stachowiak A, Rybus-Zając M (2020) Influence of rootstocks and the time of grafting procedure on the efficiency of propagation by grafting two cultivars of mountain pine (Pinus mugo Turra) and estimation of chloroplast pigments level in the needles. Acta Sci Polon Hortorum Cultus 19(2):75–85
Tellalov Y (2006) Relative female reproductive output of grafted Scots pine (Pinus sylvestris) clones planted on different places. Doctoral thesis, Swedish University of Agricultural Sciences
Velisevich SN, Bender OG, Goroshkevich SN (2021) The influence of scion donor tree age on the growth and morphogenesis of Siberian stone pine grafts. New for 52(3):473–491. https://doi.org/10.1007/s11056-020-09805-2
Weatherhead I, Barnett JR (1986) Development and structure of unusual xylem elements during graft union formation in Picea sitchensis L. Ann Bot 57(4):593–598. https://doi.org/10.1093/oxfordjournals.aob.a087139
Wheeler NC, Bramlett DL (1991) Flower stimulation treatments in a loblolly pine seed orchard. South J Appl for 15(1):44–50. https://doi.org/10.1093/sjaf/15.1.44
Ying CC, Murphy JC, Andersen S (1985) Cone production and seed yield of lodgepole pine grafts. For Chron 61(3):223–228. https://doi.org/10.5558/tfc61223-3
Zobel B, Talbert J (1984) Applied forest tree improvement. John Wiley and Sons, New Jersey
Acknowledgements
AH would like to thank Dr. Shuntaro Koizumi and Prof. Richard Bindler (EMG, Umeå University) for their help with language editing and Dr. Jade Bruxaux (EMG, Umeå University) for her valuable comments. All authors thank the technical staff at Skogforsk station in Sävar for performing grafting and post-grafting activities.
Author information
Authors and Affiliations
Contributions
TP, CA designed the experiment; TP coordinated grafting activities; AH collected the field observations and performed data analysis; AH, DH planned and discussed the statistical analysis; AH wrote the manuscript; AH, CA, UW, TP, DH discussed the results and revised manuscript.
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: This study was supported by Formas (2021–02155) and T4F programs, Sweden, and Graduate Research School in Forest Genetics, Biotechnology and Breeding, UPSC, Sweden.
The online version is available at https://link.springer.com/.
Corresponding editor: Tao Xu.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Heuchel, A., Hall, D., Almqvist, C. et al. Topgrafting as a tool in operational Scots pine breeding. J. For. Res. 35, 111 (2024). https://doi.org/10.1007/s11676-024-01766-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11676-024-01766-y