
Abstract
Urban parks are an important part of urban ecosystems and play an important role in biodiversity conservation. However, it is still unclear how park characteristics affect plant diversity which could hinder the conservation of urban biodiversity due to the ineffective design of urban parks. The diversity of woody plants in 33 parks of Changchun, China, was measured with plot measurements and linked with urban park characteristics (e.g., size and age of the park) to uncover the relationship between them. The results show that urban woody plant species were abundant, with 98 species belonging to 51 genera and 26 families in the snow climate city of Changchun. The variation in woody plant diversity was largely explained by internal patch characteristics (e.g., size, age, shape), and external factors surrounding the park (e.g., land use type and socioeconomic level) accounted for only 16.0% in our study. For internal patch characteristics, older urban parks with larger areas demonstrated a richer level of plant diversity and increased nonlinearly with increasing park area. The threshold size significantly affecting plant diversity variation was approximately 30 ha. Plant diversity had positive linear relationships with the ages of urban parks, supporting the legacy effect. In addition, woody plant diversity nonlinearly decreased with increasing park shape index, which suggested that the plant diversity could also be increased by optimizing the park shape. Regarding the external factors surrounding parks, the spatial distribution of woody plant diversity varied greatly from the urban center to the suburbs. House prices around the park had positive linear relationships with woody plant diversity in parks, supporting the luxury effect. However, building and road proportions, and green space proportion had no relationship with plant diversity in parks. This study can provide a robust reference for enriching plant diversity in urban parks, thus improving the development of urban sustainable cities.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Human development has led to the extinction of species dozens many times faster than the average level over the past 10 million years, with more than a million species currently at risk of extinction (Bongaarts 2019). Global biodiversity is decreasing at an unprecedented rate as a complex response to several human-induced changes in the global environment (Shin et al. 2022). Cities are areas where human beings make use of nature and transform nature most thoroughly (Rigolon et al. 2018). Cities gradually occupy natural habitats suitable for wildlife, and the pressure on urban biodiversity is increasing sharply (Aronson et al. 2017; McDonald et al. 2018). Therefore, it is necessary for urban planners to understand how to protect and promote biodiversity in cities under rapid urbanization.
Urban green space, as the basis for improving the urban environment and shaping urban landscapes, has not only maintained resident well-being and eased urban heat islands but also plays an important role in conserving urban biodiversity (Ren et al. 2019; Dong et al. 2022; Liu et al. 2022). As the main component of the urban green space, urban parks are the most concentrated and representative areas of urban biodiversity because of their large plant area, rich plant species, complex community structure, and diverse biological functions. Understanding the potential drivers of plant diversity in urban parks is of considerable significance to the conservation of urban biodiversity.
Previous studies have focused on the assessment of plant diversity (Huang et al. 2017; Figueroa et al. 2018; Yeshitela et al. 2020) and the planning and design of plant diversity protection (Li et al. 2019; Chang et al. 2021). Plant diversity in urban parks is related to the characteristics of the park itself, such as park type, area, age (the age of the park as the quadrat survey year (2018) minus the year when the park was first established.), and the external environment of the park (Nielsen et al. 2014; Dong et al. 2019; Liu et al. 2019; Wang and Zhao 2020). Studies have reported that larger parks have higher plant diversity and are more beneficial for protecting rare species (Tjørve et al. 2008; Chaiyarat et al. 2019; Zajdel et al. 2019; Yang et al. 2020). Older parks may preserve relatively complete native vegetation (Carpintero et al. 2014). In addition, recent studies show that some landscape characteristics surrounding the park (e.g., land use type and socioeconomic levels) and anthropogenic factors (e.g., urban areas and population) might also influence plant diversity (Miller et al. 2018; Roman et al. 2018; Gonçalves et al. 2021; Dasgupta et al. 2022). High tree diversity was found in old, high-income locations (Clarke et al. 2013). However, the potential drivers of plant diversity in urban parks are not fully understood. The relative contributions of the external landscape characteristics surrounding the park and the internal patch factors of urban parks to biodiversity are unclear. In addition, many in-depth studies on the species diversity of urban biodiversity have been carried out in subtropical and temperate regions such as Australia, Caledonia, the Caribbean, the Philippines, and in Chinese cities such as Wuhan and Haikou (Kendal et al. 2012; Fisher et al. 2021; Liu et al. 2021; Nizamani et al. 2021; Pasion et al. 2021). Related research in the northern cold temperate zone (snow climate) of Northeast China has received little attention, which could hinder developing a better understanding of plant diversity in the urban parks. Changchun, as a typical snow climate city in northeast China, has experienced accelerated urbanization since 1979, which may have caused changes in plant diversity in urban parks.
Based on a field-based survey with GF-2 images from Changchun, this study investigated the patterns and potential drivers of woody plant diversity in urban parks. The specific objectives were to: (1) identify the driving factors affecting plant diversity in urban parks; (2) explore the contribution of the park’s internal patch factors and external landscape characteristics to species biodiversity; and (3) explore the quantitative relationships between these driving factors and species diversity. The goal is to provide useful information for urban planners and designers on the design of urban parks to protect, maintain or enhance biodiversity.
Materials and methods
Study area
Changchun city (125°11′ − 125°25 E′, 43°43′ − 43°59′ N), is the political and socioeconomic center of Jilin Province located in the hinterland of Northeast China. It has a continental climate characterized by long, cold, dry winters and short, hot summers. The city receives an average annual rainfall of 567 mm and an annual average temperature of 4.8 °C (Zhang et al. 2016); the highest temperature in summer is 38.5 °C, and the lowest in winter is − 36.5 °C.
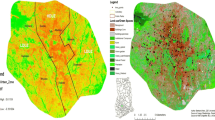
Changchun was named a national forest city by 2016, with almost 45% of its area covered by vegetation (Ren et al. 2018). The forest types are mainly composed of evergreen and deciduous, represented by willows (e.g., Salix babylonica L., oaks such as Quercus mongolica Fisch. ex Ledeb.), pines (e.g., Pinus tabulaeformis var. mukdensis Uyeki). Cypress (e.g., Juniperus procumbens Miq.) is common in the shrub layer. There were 14 waterfront parks, 25 typical urban parks, 4 large-scale orchards, more than 700 street and road forest belts, and more than 500 garden-style units. An urban greening system was developed combining points, lines, and surfaces. The goal of greening, beautification and landscaping was achieved, and it is moving toward a higher quality forest city. Obviously, Changchun is ideal for research on forest structure and species diversity of urban parks (Fig. 1).

Study area within the fifth-ring road of Changchun
Park biodiversity
A stratified random sampling method was used (Margules and Pressey 2000; Yang et al. 2021). Plot surveys on plant diversity were carried out during the summer of 2018 − 2019. An overall survey of the park was first carried out before the plot survey to ensure that the quadrat selection was scientific and reasonable. The number of quadrats was determined according to the different plant communities and park sizes (Cheng et al. 2015) to represent the overall biodiversity of the park. A total of 230 sample plots were set up in 33 typical urban parks, in which the sampling quadrats were 30 m × 30 m (900 m2). To avoid any deviation caused by artificial selection during sampling, the random quadrats were selected for each park on the Baidu map by the methods used in the UFORE (the urban forest effects) Model (Nowak et al. 2003). The UFORE computer model was developed by U.S. Forest Service in 2000 to help managers and researchers quantify UF structure and function.
All woody plants in each quadrat were recorded. Some urban forest studies divide woody plants into trees and shrubs according to their height (Yang et al. 2017c; Li et al. 2020). Although the concepts “tree” and “shrub” do have distinct definitions from a perspective of common sense, they are not rigorous in an academic context. Instead, the terminologies of forest vertical structure research were used for this study (Mestre et al. 2017; Pregitzer et al. 2019).
Within different plots, the species was recorded and the diameter at breast height (DBH), basal area, height, leaf area index and growth status of each species were determined. For the availability, typicality, and the ability to reflect the biodiversity of plant communities (Chao et al. 2014; Yang et al. 2017b; Muvengwi et al. 2019), the species richness, Margalet index, Shannon–Wiener index and Simpson index were used for 33 urban parks (Table 1).
Driving factors of urban biodiversity
Ecological research shows that some varieties of patch characteristics could determine the dynamics of urban biodiversity (Liu et al. 2019; Wang and Zhao 2020). Consequently, four parameters were selected as patch characteristics: the park area and perimeter, the shape index of the park, and the age of the park. GF-2 remote sensing with high spatial resolution was acquired in this study at 10:25 am, 14 September 2018 to extract area and perimeter of the selected parks. The calculation of areas and perimeters was completed in ArcGIS 10.0. The shape index was calculated from the ratio of perimeter to area. Park age data were collected from the statistical yearbook and the Gardens and Forestry Bureau in Changchun.
To explore possible changes in plant diversity across different parks, the parks were placed into three classes (Cheng et al. 2015): large (≥ 30 ha), medium-size (10 ha ≤ area < 30 ha), and small (< 10 ha). In addition, the parks also divided into four types according to years since establishment (I: ≥ 70 years; II: 41 years ≤ age < 70 years; III: 19 years ≤ age < 41 years; and IV: age < 19 years). In addition, to confirm the impact of geographical locations on park plant diversity, the ring road map was used to classify parks into five categories, i.e., parks located on ring roads 1 − 5 (R1 − R5). Moreover, to determine the impact of economic factors on park diversity index, the housing prices in the surrounding area were divided into three levels (high: ≥ 12,000 yuan RMB; middle: 5000 yuan RMB ≤ price < 12,000 yuan RMB; low: < 5000 yuan RMB).
Since urban biodiversity may be affected by anthropogenic activities around the park (McKinney 2002; Yan et al. 2019), four parameters that might affect woody plant diversity were selected as environmental variables: housing prices, proportion of buildings, roads, and green space within a 300-m buffer zone (Yang et al. 2017a). Data on housing prices were extracted from https://www.58.com/fangjiawang/shi-2018/. Before analysis, the two GF-2 images were radiometrically calibrated and geometrically corrected with an accuracy ˂ 0.5 pixels (Dong et al. 2020). All image preprocessing was performed in ENVI 5.3 software. Visual interpretation was used to extract building, road, and green space information from high spatial resolution images. The multispectral and panchromatic images were fused to produce a four-band multispectral image at a 2-m resolution. The edge of each land use classification was identified based on its color, size, and location (Dong et al. 2022; Xue et al. 2019).
Data analysis
To compare differences in the diversity indices of the urban parks under different sizes, time since establishment, and locations, multiple comparison methods were used. Redundancy analysis (RDA) was used to calculate the contribution of internal patch characteristics and external landscape factors to the park biodiversity level. Variation partitioning analysis was employed to distinguish the individual and mixed effects of driving factors. A linear regression model and correlation coefficient were then applied to detect possible impacts of factors on park biodiversity. In all statistical analyses, the internal patch characteristics (area, perimeter, shape, and age) and external landscape factors (housing prices, and percentages of buildings, roads, and green space within the 300-m buffer zone) were used as independent variables, whereas the biodiversity indices (Richness, Margalet index, Shannon–Wiener index, and Simpson index) were dependent variables.
Results
Changes in woody plant diversity
Urban woody plant species in Changchun were abundant, with 98 species belonging to 51 genera and 26 families, and diversity changed significantly across different parks. Overall, plant richness had relatively large ranges from 3 to 22, with an average of 10. The averaged Margalet index was 1.12, ranging from 0.57 to 2.80, and the Shannon index ranged from 0.70 to 2.69, with an average of 1.13. The Simpson index ranged from 0.42 to 0.79, with an average of 0.58. The results showed that the plant diversity indices (Richness, Margalet, Shannon–Weiner and Simpson) in large parks were significantly higher (p < 0.05) than in small parks (Fig. 2a). This suggests that large parks could have more woody plant species than small and medium-sized parks in cities (Fig. 2b, 2c, and 2d). In parks with large areas, the diversity indices were significantly higher. In addition, older parks had more abundant plant species, and the Margalet index, Shannon index and Simpson index were significantly higher in parks ≥ 70 years than in the other parks (Fig. 3), which can support the legacy effect. In addition, the spatial distribution of diversity varied greatly across ring-road areas with different urbanization intensities. The species diversity levels of the parks in the first ring-road area in the urban center and in the fifth ring-road areas in the suburbs were significantly higher than those of parks in other ring-road areas (Fig. 4). Moreover, the parks located in areas with higher housing prices had more woody plant species, and the Shannon and Simpson indices were significantly where housing prices were ≥ 12,000 yuan RMB (Fig. 5), which supports the luxury effect. Furthermore, according to the proportion of roads, buildings, and green space in the buffer zone around the park, the woody plant diversity indices of the different classes of parks did not change significantly (Fig. S1, S2 and S3).

Species diversity index of urban parks with different park area; different letters show significant differences, P < 0.05

Species diversity index of urban parks with different ages (I: ≥ 70, II: 41 ≤ < 70, III: 19 ≤ < 41, IV: < 19); different letters show significant differences, P < 0.05

Species diversity index of urban parks under different ring road.(R1: the first ring; R2: the second ring; R3: the third ring; R4: the fourth ring; and R5: the fifth ring; different letters show significant differences, P < 0.05.)

Species diversity index of urban parks surrounded by different housing price.( Different letters show significant differences, P < 0.05.)
The main driving factors of woody plant diversity in urban parks
Redundancy analysis (RDA) was conducted between the diversity indices and the internal and external factors of the parks (Fig. 6). The results show that the overall impact of the internal and external factors on woody plant diversity was 43.9%, among which the internal factors were approximately twice that of external factors. Internal patch characteristics of area, perimeter, shape, and age accounted for 27.9% of the variation in woody plant diversity, and external landscape features (housing prices, proportion of buildings, roads, and green space in the 300-m buffer zone) contributed 16.0%. The mixed contribution of patch characteristics and landscape features of the parks contributed 7.3% of the variation. For the internal factors, park area, perimeter, and age contributed 23.3%, 21.9% and 18.6% of the variation, respectively. For the external factors, the proportion of buildings, roads, and green space in the 300-m buffer zone contributed 7.2% to the changes in woody plant diversity.

RDA analysis and variation partitioning of internal patch factors and external factors for species diversity changes in urban parks. (G300, B300 and R300 represent the proportions of green space, buildings, and roads, respectively, in the 300-m buffer zone around the park)
Statistical models between the main drivers and woody plant diversity
Spearman correlation coefficient (Fig. 7) showed that the urban park area (R2 = 0.58, P < 0.01) and perimeter (R2 = 0.53, P < 0.05) had a significant positive relationship with the richness index. Park age also had a significant positive relationship with the Margalet, Shannon and Simpson indices. However, there was a negative relationship between park shape (R2 = 0.53, P < 0.05) and the richness index. For exterior factors around parks, housing price was positively correlated with Shannon (R2 = 0.47, P < 0.05) and Simpson (R2 = 0.64, P < 0.01) indices.

Heatmaps of the correlation coefficients between plant diversity of urban parks and their drivers
Additional quantitative regression models were further established to reveal the relationship between diversity indices and their driving factors (Figs. S4 and S5). There is a logarithmic function relationship between urban park area and richness (Fig. S4a). Further analysis showed that the threshold value of urban park area for significantly affecting the variation in woody plant diversity was approximately 30 ha. As park area increased but remained < 30 ha, the diversity sharply increased with increasing area. However, when the area was > 30 ha, plant richness increased slowly according to the model curves (Fig. S4a). The linear positive model was established between park perimeter and richness (Fig. S5b), explaining 31% of the variation in species diversity. This result suggests that the number of species in the park increased linearly with the increase in park perimeter. In addition, it was also found that park shape (perimeter/area) had a negative nonlinear relationship with plant richness (Fig. S4c), indicating that diversity decreased with increasing shape. Twenty-eight percent of the diversity variance could be explained by the nonlinear model with urban park shape as an independent variable. Therefore, park shape can also play a more important role in improving biodiversity.
There were linear positive relationships between age of parks and the Margalet and Shannon indices (Figs. S5a and S5b). This suggests that woody plant diversity increased linearly with park age, and those older parks can have greater plant diversity, which supports the legacy effect. The Shannon and Simpson indices had a linear positive correlation with housing prices around urban parks (Figs. S5c and S5d). The higher the housing price, the higher the level of plant diversity, which supports the luxury effect. In general, the results show that the park patch factor has a great influence on the diversity of urban parks, and patch factors should be considered in the renewal and development of parks to promote biodiversity conservation.
Discussion
Urban plant diversity in parks
In recent years, the development of an ecological civilization has been elevated to a national strategic level, and the development of a “park city” has become an important goal for residents for a better life (An et al. 2019). A city park usually has a wide variety of species and is the most representative and biodiversity-rich area in the city, which has direct correlation with urban ecological function. With the continuous enhancement and improvement of people’s awareness and subsequent demand for ecological services, a good urban environment with high biodiversity will become the key advantage and competitive aspect of sustainable urban development in the future. Besides, in the context of the development of a national forest city, the concept has also gradually changed from afforestation/reforestation to the development of urban parks with great biodiversity and high ecological functions (Ren et al. 2021). Therefore, the study of plant species diversity in urban parks is of great significance for protecting biodiversity, improving ecological quality, and maintaining urban sustainable ecological development.
Effects of patch characteristics on species diversity in parks
According to the species-area relationship, diversity increases with patch area (He and Legendre 1996), and habitat size is one of the main factors affecting species survival. In the urban ecosystem, a park, as an independent landscape unit, has unique patch properties and can be regarded as a “green island” (Ren et al. 2013) with a fixed area and shape, and provides an excellent location for studying the relationship between urban biodiversity and patch characteristics. Our results show that the plant diversity index in large parks was significantly higher (P < 0.05) than that in small parks, which supports the observation that area determines the number of habitats and plant species (Carpintero et al. 2014; Sing et al. 2016; Chaiyarat et al. 2019; Liu et al. 2019). To some extent, the larger the park area, the higher the plant species richness, the more plants tend to be tall shade trees, and the higher the diversity and heterogeneity of internal habitats are, so the park has a greater chance to maintain and protect biodiversity and have a higher conservation value (Bräuniger et al. 2010; Peng et al. 2020). Research has defined minimum patch size as the area of the smallest patch that allows native plants to survive, and in many ecosystems, the smallest patch controls the entire ecosystem. Therefore, determining the minimum park area has an important role in maintaining biodiversity (Figueroa et al. 2018). This research shows that when the area of the park reaches 30 ha, the diversity level of the park peaks. In addition, it also found that the richness index of parks located on the periphery of the city showed an upward trend, which may be due to blending in with the increased natural vegetation found in suburbs (compared to city centers) to form diverse habitats, thereby increasing the diversity of plant species. However, our research also found that the richness index of large parks located in the middle of the city showed a downward trend, which may be due to their geographical location to effectively allocate sufficient diversity of material, resulting in unbalanced species allocation. Urban planners need to strengthen the development of large parks on the periphery of cities.
In addition to park size, shape is also an important factor affecting biodiversity. Studies of tropical forest biodiversity have shown that the species diversity of trees, shrubs and vines is mainly affected by patch shape (Yan et al. 2021). In Mexico, research has shown a negative correlation between species richness and boundary shapes (Nava-Díaz et al. 2022). Our research also showed that park diversity levels tend to decrease as the shape index increases, an indication of the complexity of the park. The higher the shape index, the more complex the shape of the park. A complex park shape reduces the seed dispersal and pollinator services in green spaces, making the flow of organisms between patches more difficult (Yan et al. 2021). In addition, complex shapes can alter biotic and abiotic conditions at park edges, leading to the loss of sensitive species (Omar et al. 2018). The combined effect of these two processes may explain why the species richness of saplings/seedlings was negatively associated with shape index.
In addition, there are certain differences in the species diversity of trees in different parks, which is also related to the cultural background, economic level, and aesthetic level of people in different time periods. A legacy hypothesis suggests that activities by prior land managers persist throughout urban development and influence current vegetation cover and diversity (Pouget et al. 2016). The composition and growth of plant species in parks have distinct historical and regional characteristics (Talal and Santelmann 2019). All these factors indirectly redistribute species within a park, thereby affecting diversity levels. Our results found that species richness was positively correlated with park age, and the earlier the urban park was established, the higher the species diversity (Figs. S5a and S5b). It may be that tree species cultivated in the early years of the park were at normal stocking levels, and newly planted species in the later years increased the number of species (Figueroa et al. 2018). Our results also showed that habitat homogeneity of a new park was higher than that of an older one, with fewer habitats and less species richness. Years of park development and species diversity not only had the effect of early configuration, but also the involvement of different park managers, resulting in greater habitat diversity that could support more plant growth (Chan et al. 2018). Our findings agreed with Figueroa et al. (2018) that parks created earlier have more stable environments, more species, and richer diversity than newly established parks. Many of the urban parks in Changchun formed in the early years were based on the original green space of the area and have specific native species. After long-term growth and development, the tree species are stable and in abundant. Urban parks created relatively more recently are mostly formed by one or several main tree species and the variety of tree species is relatively limited. In general, the older the park, the higher the level of diversity, which also supports the legacy effect (Clarke et al. 2013).
Effects of exterior characteristics on species diversity
The urban buffer division gradient can also be regarded as a habitat loss gradient (McKinney et al. 2002). In this gradient, parks, and other green spaces have the characteristics of isolated green islands in urban environments. When the urban matrix cannot penetrate and diffuse, an isolation effect occurs (Garden et al. 2010). Landscape or habitat fragmentation is the result of a gradual reduction in the natural environmental surface as well as its progressive isolation (Ziv and Davidowitz 2019). This process is one of the main threats to biodiversity (Hanski 2015; Fletcher et al. 2018; Ntshanga et al. 2021). This study showed that, on a small scale, the plant diversity level of a park is less affected by the landscape surrounding the park, i.e., roads, buildings and green space within the buffer zone have weaker correlations with species richness, and Margalet, Shannon and Simpson indices. The reason is that in the urban environment, the park belongs to an artificial ecosystem; the internal plants have the attributes of urban plants and their spatial arrangement is influenced by planners. However, roads and buildings in the buffer zone, as a corridor in the landscape, increase the fragmentation of the landscape (Koren and Butler 2006; Peng et al. 2022). This can threaten the environment within the park itself, as the connections of matter circulation and energy flow are broken. This observation is not significant in our research, but it can provide suggestions. For example, in many cases in the immediate zone bordering a park, buffer zones are created to maintain ecological processes and the stability of ecosystems to preserve biodiversity (Kubacka et al. 2022).
Moreover, socioeconomic differences may also influence urban biodiversity and the luxury effect may be dominant at the biodiversity level. The relationship between wealth and plant diversity is termed the “luxury effect” (Hope et al. 2003), whereby as human economic conditions improve, urban landscapes with higher plant diversity are occupied. Overall, our research shows that the closer a house is to the city center, the higher the house price, and the more significant the luxury effect (Figs. S5c and S5d). High-priced housing reflects the social status and quality of life of residents in wealthier areas. People in the high-income class generally have higher demands on the environment around them; they require a higher level of biodiversity to meet their requirements. At the same time, higher species diversity requires more resources and space to be maintained. The mutual promotion of residents’ wealth and biodiversity shows the linear relationship: the level of biodiversity increases with the increase in housing prices, supporting the luxury effect (Huang et al. 2019).
From this study on an urban scale, species diversity indices of a park are affected by internal natural factors but are not strongly correlated with external factors of human activities. However, with increasing urbanization, the influence of humans on the structural characteristics and biodiversity of park forests may gradually become stronger. Further research needs to integrate the five driving factors of nature, society, politics, technology, and culture to conduct comprehensive explorations from multiple perspectives.
Management implications
It is of considerable management significance for urban planners to study the driving factors of urban species diversity. This suggests that the biological attributes may be optimized by changing the characteristics of the park and its surroundings.
Because of the contradiction between social development and biodiversity conservation, large areas of land cannot be protected as biodiversity habitats. It is necessary to explore the landscape pattern that can maintain high biodiversity and optimize the land area. This has important implications for protecting biodiversity. Our research shows that park area is positively correlated with species diversity, and after a certain area threshold has been reached (30 ha), species diversity shows a slow growth. The shape index of a park is also an important factor affecting biodiversity. The edges of natural landscapes, such as parks near suburban areas, many natural edges and community intersections, are often irregular and have relatively rich biodiversity, while man-made barriers such as roads often reduce biodiversity. Therefore, during the development of a park, urban planners should set up a green buffer zone to avoid negative effects of building roads and other isolated edges. This study highlighted that the luxury effect around an urban park may also be an important factor affecting biodiversity. The diversity of tree species in parks in areas with low housing prices is low, increasing the park’s inability to protect the stability of the ecosystem. Our findings suggest that cities should increase human and financial investment in the green management of low-income parks to improve biodiversity levels.
The limitation of this study is that Changchun is a single city in the cold temperate zone of China, and the universality of the conclusions to other cities needs to be verified. After this, research should be carried out in cities of other climate zones.
Conclusion
From the field surveys and remote sensing satellite images of Changchun city, we identified the relationship between urban park patch characteristics and external landscape characteristics and species diversity index. The following conclusions can be made:
-
Variation in woody plant diversity was largely explained by internal patch characteristics (size, ages, shape), which accounted for 27.9% of diversity, and external factors surrounding the park (e.g., land use type and socioeconomic levels) accounted for 16.0%.
-
Larger urban parks areas have a rich level of plant diversity. The threshold level for significantly affecting plant diversity is approximately 30 ha. Plant diversity had a linear, positive trend with park age and house prices around the park supporting legacy and luxury effects. It had a nonlinear decreasing trend with the increasing park shape, suggesting that plant diversity could be increased by optimizing the urban park shape.
-
Our findings indicate that city managers should increase human and financial investment in the green infrastructure of parks and set up a green buffer zone to improve biodiversity levels.
Our study highlights the importance of urban parks as fundamental areas for biodiversity conservation. The results may help researchers overcome the randomness of park allocation and to conduct in-depth exploration the perspective of enriching urban plants and protecting urban biodiversity.
References
An BY, Wang D, Liu XJ, Guan HM, Wei HX, Ren ZB (2019) The effect of environmental factors in urban forests on blood pressure and heart rate in university students. J for Res 24(1):27–34
Aronson MF, Lepczyk CA, Evans KL, Goddard MA, Lerman SB, MacIvor JS, Nilon CH, Vargo T (2017) Biodiversity in the city: key challenges for urban green space management. Front Ecol Environ 15(4):189–196
Bongaarts J (2019) Summary for policymakers of the global assessment report on biodiversity and ecosystem services of the Intergovernmental science-policy platform on biodiversity and ecosystem services. Popul Dev Rev 45(3):680–681
Bräuniger C, Knapp S, Kühn I, Klotz S (2010) Testing taxonomic and landscape surrogates for biodiversity in an urban setting. Landsc Urban Plan 97(4):283–295
Carpintero S, Reyes-López J (2014) Effect of park age, size, shape, and isolation on ant assemblages in two cities of Southern Spain. Entomolog Sci 17(1):41–51
Chaiyarat R, Wutthithai O, Punwong P, Taksintam W (2019) Relationships between urban parks and bird diversity in the Bangkok metropolitan area. Thail Urban Ecosyst 22(1):201–212
Chan CS, Si FH, Marafa LM (2018) Indicator development for sustainable urban park management in Hong Kong. Urban For Urban Green 31:1–14
Chang CR, Chen MC, Su MH (2021) Natural versus human drivers of plant diversity in urban parks and the anthropogenic species-area hypotheses. Landsc Urban Plan 208:104023
Chao A, Gotelli NJ, Hsieh TC, Sander EL, Ma KH, Colwell RK, Ellison AM (2014) Rarefaction and extrapolation with hill numbers: a framework for sampling and estimation in species diversity studies. Ecol Monogr 84(1):45–67
Cheng XY, Wei BS, Chen GJ, Li JX, Song CH (2015) Influence of park size and its surrounding urban landscape patterns on the park cooling effect. J Urban Plan Dev 141(3):A4014002
Clarke LW, Jenerette GD, Davila A (2013) The luxury of vegetation and the legacy of tree biodiversity in los angeles, CA. Landsc Urban Plan 116:48–59
Dasgupta S, Bhattacharya T, Bhamburkar P, Kaul R (2022) Role of anthropogenic drivers in altering the forest community structure in a prime tiger habitat in central India. Can J for Res 52(3):406–415
Dong D, Qin ZQ, Li CG, Jiang DB, Li TT (2019) Comparison of landscape and species diversity of woody plants in new and old urban park in Huainan city. J Northwest Fore Univ 34(5):247–254 ((in Chinese))
Dong YL, Ren ZB, Fu Y, Miao ZX, Yang R, Sun YH, He XY (2020) Recording urban land dynamic and its effects during 2000–2019 at 15-m resolution by cloud computing with landsat series. Remote Sens 12(15):2451
Dong YL, Ren ZB, Fu Y, Hu NL, Guo YJ, Jia GL, He XY (2022) Decrease in the residents’ accessibility of summer cooling services due to green space loss in Chinese cities. Environ Int 158:107002
Figueroa JA, Castro SA, Reyes M, Teillier S (2018) Urban park area and age determine the richness of native and exotic plants in parks of a latin American city: santiago as a case study. Urban Ecosyst 21(4):645–655
Fisher JC, Bicknell JE, Irvine KN, Hayes WM, Fernandes D, Mistry J, Davies ZG (2021) Bird diversity and psychological wellbeing: A comparison of green and coastal blue space in a neotropical city. Sci Total Environ 793:148653
Fletcher RJ Jr, Didham RK, Banks-Leite C, Barlow J, Ewers RM, Rosindell J, Holt RD, Gonzalez A, Pardini R, Damschen EI, Melo FPL, Ries L, Prevedello JA, Tscharntke T, Laurance WF, Lovejoy T, Haddad NM (2018) Is habitat fragmentation good for biodiversity? Biol Cons 226:9–15
Garden JG, McAlpine CA, Possingham HP (2010) Multi-scaled habitat considerations for conserving urban biodiversity: native reptiles and small mammals in Brisbane. Aust Landsc Ecol 25(7):1013–1028
Gonçalves P, Grilo F, Mendes RC, Vierikko K, Elands B, Marques TA, Santos-Reis M (2021) What’s biodiversity got to do with it? Perceptions of biodiversity and restorativeness in urban parks. Ecol Soc 26(3):25
Hanski I (2015) Habitat fragmentation and species richness. J Biogeogr 42(5):989–993
He FL, Legendre P (1996) On species-area relations. Am Nat 148(4):719–737
Hope D, Gries C, Zhu W, Fagan WF, Redman CL, Grimm NB, Nelson AL, Martin C, Kinzig A (2003) Socioeconomics drive urban plant diversity. Proc Natl Acad Sci 100(15):8788–8792
Huang LJ, Wang Q, Lin LL, Zhang ZK, Liu XZ, Su ZM, Huang HD (2017) The effect of urbanization on distribution of woody plant diversity in parks. J Anhui Agric Univ 44(6):1052–1059 ((in Chinese))
Huang Y, Chen Y, He YJ, Wang M, Kuang SJ, Chen GJ, Li JX (2019) Effects of socioeconomic factors on urban plant diversity of residential areas in Shanghai. China J Appl Ecol 30(10):3403–3410 ((in Chinese))
Kendal D, Williams NS, Williams KJ (2012) Drivers of diversity and tree cover in gardens, parks and streetscapes in an Australian city. Urban For Urban Green 11(3):257–265
Koren HS, Butler CD (2006) The interconnection between the built environment ecology and health. Environ Secur Environ Manag: Role Risk Assess 5:111–125
Kubacka M, Żywica P, Subirós JV, Bródka S, Macias A (2022) How do the surrounding areas of national parks work in the context of landscape fragmentation? A case study of 159 protected areas selected in 11 EU countries. Land Use Policy 113:105910
Li XP, Fan SX, Kühn N, Dong L, Hao PY (2019) Residents’ ecological and aesthetical perceptions toward spontaneous vegetation in urban parks in China. Urban For Urban Green 44:126397
Li XT, Jia BQ, Zhang W, Ma J, Liu XP (2020) Woody plant diversity spatial patterns and the effects of urbanization in Beijing. China Urban For Urban Green 56:126873
Liu KL, Peng MH, Hung YC, Neoh KB (2019) Effects of park size, peri-urban forest spillover, and environmental filtering on diversity, structure, and morphology of ant assemblages in urban park. Urban Ecosyst 22(4):643–656
Liu J, Mao WP, Ding H, Li HG, Chen L, Chen XM, Tian JH (2021) Composition and diversity of mosquito community in Wuhan from 2017 to 2019. J Cent China Norm Univ 55:416–423
Liu K, Li XK, Wang SD, Gao XJ (2022) Assessing the effects of urban green landscape on urban thermal environment dynamic in a semiarid city by integrated use of airborne data, satellite imagery and land surface model. Int J Appl Earth Obs Geoinf 107:102674
Margules CR, Pressey RL (2000) Systematic conservation planning. Nature 405(6783):243–253
McDonald RI, Colbert M, Hamann M, Simkin R, Sharp R (2018) Nature in the urban century: a global assessment of where and how to conserve nature for biodiversity and human wellbeing.
McKinney ML (2002) Urbanization, biodiversity, and conservation. Bioscience 52(10):883
Mestre L, Toro-Manríquez M, Soler R, Huertas-Herrera A, Martínez-Pastur G, Lencinas MV (2017) The influence of canopy-layer composition on understory plant diversity in southern temperate forests. For Ecosyst 4(1):1–13
Miller KM, McGill BJ, Mitchell BR, Comiskey J, Dieffenbach FW, Matthews ER, Perles SJ, Schmit JP, Weed AS (2018) Eastern national parks protect greater tree species diversity than unprotected matrix forests. For Ecol Manag 414:74–84
Muvengwi J, Kwenda A, Mbiba M, Mpindu T (2019) The role of urban schools in biodiversity conservation across an urban landscape. Urb Fore Urb Green 43:126370
Nava-Díaz R, Zuria I, Pineda-López R (2022) Taxonomic, phylogenetic and functional diversity of bird assemblages in urban green spaces: null model analyses, temporal variation and ecological drivers. Front Ecol Evol 9:1037
Nielsen AB, Van Den Bosch M, Maruthaveeran S, van den Bosch CK (2014) Species richness in urban parks and its drivers: A review of empirical evidence. Urb Ecosyst 17(1):305–327
Nizamani MM, Harris AJ, Cheng XL, Zhu ZX, Jim CY, Wang HF (2021) Positive relationships among aboveground biomass, tree species diversity, and urban greening management in tropical coastal city of Haikou. Ecol Evol 11(17):12204–12219
Nowak DJ, Crane DE, Stevens JC, Hoehn RE (2003) The urban forest effects (UFORE) model: Field data collection manual. US Department of Agriculture Forest Service Northeastern Research Station: Syracuse, NY, USA, 4-11.
Ntshanga NK, Procheş S, Slingsby JA (2021) Assessing the threat of landscape transformation and habitat fragmentation in a global biodiversity hotspot. Aust Ecol 46(7):1052–1069
Omar M, Al Sayed N, Barré K, Halwani J, Machon N (2018) Drivers of the distribution of spontaneous plant communities and species within urban tree bases. Urb For Urb Green 35:174–191
Pasion BO, Barrias CD, Asuncion MP, Angadol AH, Pabiling RR, Pasion A Jr, Braulio AA, Baysa AM Jr (2021) Assessing tree diversity and carbon density of a riparian zone within a protected area in southern Philippines. J Asia-Pacific Biodivers 14(1):78–86
Peng MH, Hung YC, Liu KL, Neoh KB (2020) Landscape configuration and habitat complexity shape arthropod assemblage in urban parks. Sci Rep 10(1):1–12
Peng SJ, Zhang J, Zhang XL, Li YQ, Liu YP, Wang ZH (2022) Conservation of woody species in China under future climate and land-cover changes. J Appl Ecol 59(1):141–152
Pouget M, Youssef S, Dumas PJ, Baumberger T, San Roman A, Torre F, Médail LAF, Baumel A (2016) Spatial mismatches between plant biodiversity facets and evolutionary legacy in the vicinity of a major Mediterranean city. Ecol Ind 60:736–745
Pregitzer CC, Charlop-Powers S, Bibbo S, Forgione HM, Gunther B, Hallett RA, Bradford MA (2019) A city-scale assessment reveals that native forest types and overstory species dominate New York City forests. Ecol Appl 29(1):e01819
Ren ZB, He XY, Zheng HF, Zhang D, Yu XY, Shen GQ, Guo RC (2013) Estimation of the relationship between urban park characteristics and park cool island intensity by remote sensing data and field measurement. Forests 4(4):868–886
Ren ZB, He XY, Pu RL, Zheng HF (2018) The impact of urban forest structure and its spatial location on urban cool island intensity. Urb Ecosyst 21(5):863–874
Ren ZB, Zheng HF, He XY, Zhang D, Shen GQ, Zhai C (2019) Changes in spatio-temporal patterns of urban forest and its above-ground carbon storage: Implication for urban CO2 emissions mitigation under China’s rapid urban expansion and greening. Environ Int 129:438–450
Ren ZB, Zhao HB, Fu Y, Xiao L, Dong YL (2021) Effects of urban street trees on human thermal comfort and physiological indices: a case study in Changchun city. China J For Res 33(3):12
Rigolon A, Browning M, Jennings V (2018) Inequities in the quality of urban park systems: an environmental justice investigation of cities in the United States. Landsc Urb Plan 178:156–169
Roman LA, Pearsall H, Eisenman TS, Conway TM, Fahey RT, Landry S, Vogt J, Doorn NS, Grove JM, Locke DH, Bardekjian AC, Battles JJ, Cadenasso ML, Konijnendijk van den Bosch CC, Avolio M, Berland A, Jenerette GD, Mincey SK, Pataki DE, Staudhammer C (2018) Human and biophysical legacies shape contemporary urban forests: a literature synthesis. Urb For Urb Green 31:157–168
Shin YJ, Midgley GF, Archer ER, Arneth A, Barnes DK, Chan L, Hashimoto S, Hoegh-Guldberg O, Insarov G, Leadley P, Levin LA, Ngo HT, Pandit R, Pires APF, Pörtner H, Rogers AD, Scholes RJ, Settele J, Smith P, Smith P (2022) Actions to halt biodiversity loss generally benefit the climate. Glob Change Biol 28(9):2846–2874
Sing KW, Jusoh WF, Hashim NR, Wilson JJ (2016) Urban parks: refuges for tropical butterflies in Southeast Asia? Urb Ecosyst 19(3):1131–1147
Talal ML, Santelmann MV (2019) Plant community composition and biodiversity patterns in urban parks of Portland. Or Front Ecol Evolut 7:201
Tjørve E, Kunin WE, Polce C, Calf Tjørve KM (2008) Species–area relationship: separating the effects of species abundance and spatial distribution. J Ecol 96(6):1141–1151
Wang Y, Zhao L (2020) A comparative study on diversity and composition of greening tree species in parks of different ages in Chongqing. Guihaia 40(4):546–556
Xue Z, Hou G, Zhang Z, Lyu X, Jiang M, Zou Y, Shen XJ, Wang J, Liu X (2019) Quantifying the cooling-effects of urban and peri-urban wetlands using remote sensing data: case study of cities of Northeast China. Landsc Urb Plan 182:92–100
Yan ZG, Teng MJ, He W, Liu AQ, Li YR, Wang PC (2019) Impervious surface area is a key predictor for urban plant diversity in a city undergone rapid urbanization. Sci Total Environ 650:335–342
Yan Y, Jarvie S, Zhang Q, Zhang S, Han P, Liu Q, Liu P (2021) Small patches are hotspots for biodiversity conservation in fragmented landscapes. Ecol Ind 130:108086
Yang CB, He XY, Yu LX, Yang JC, Yan FQ, Bu K, Chang LP, Zhang SW (2017a) The cooling effect of urban parks and its monthly variations in a snow climate city. Remote Sens 9(10):1066
Yang CY, Li EG, Chen HY, Zhang JH, Huang YM (2017b) Biodiversity of natural vegetation and influencing factors in western Inner Mongolia. Biodivers Sci 25(12):1303
Yang J, Yan PB, He RX, Song XQ (2017c) Exploring land-use legacy effects on taxonomic and functional diversity of woody plants in a rapidly urbanizing landscape. Landsc Urb Plan 162:92–103
Yang XR, Tan XW, Chen CW, Wang YP (2020) The influence of urban park characteristics on bird diversity in Nanjing. China Avian Res 11(1):1–9
Yang MY, Pickering CM, Xu L, Lin X (2021) Tourist vehicle as a selective mechanism for plant dispersal: Evidence from a national park in the eastern Himalaya. J Environ Manag 285:112109
Yeshitela K (2020) Attitude and perception of residents towards the benefits, challenges and quality of neighborhood parks in a sub-saharan Africa city. Land 9(11):450
Zajdel B, Borański M, Kucharska K, Jojczyk A, Brzezińska K (2019) Bumblebee communities (apidae, bombini) in urban parks in relation to park area and other characteristics. Pol J Ecol 67(1):84–93
Zhang D, Zheng HF, He XY, Ren ZB, Zhai C, Yu XY, Mao ZX, Wang PJ (2016) Effects of forest type and urbanization on species composition and diversity of urban forest in Changchun. North China Urb Ecosyst 19(1):455–473
Ziv Y, Davidowitz G (2019) When landscape ecology meets physiology: effects of habitat fragmentation on resource allocation trade-offs. Front Ecol Evol 7:137
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: This work was supported by the Youth Innovation Promotion Association of Chinese Academy of Sciences (Grant No. 2020237 and 2019234), and the National Natural Science Foundation of China (Grant No. 42171109 and 32130068).
The online version is available at http://www.springerlink.com.
Corresponding editor: Yu Lei.
Supplementary Information
Below is the link to the electronic supplementary material.





Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Ma, Z., Zhang, P., Hu, N. et al. Understanding the drivers of woody plant diversity in urban parks in a snow climate city of China. J. For. Res. 34, 1021–1032 (2023). https://doi.org/10.1007/s11676-022-01535-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-022-01535-9