
Abstract
Disease arises during the storage of potatoes as a result of pathogens introduced during growing and harvest of the tubers. In this study, the causative fungi of domestic potato dry rot disease were identified, and their pathogenicity was confirmed. A total of 76 species were isolated from 93 potato necropods collected from samples inoculated with dry rot disease. These 76 isolates were identified as Fusarium boothii, F. circinatum, F. citricola, F. foetens, F. iranicum, F. longifundum, F. oxysporum, F. pseudoanthophilum, F. solani, Botryotinia ranunculi, Clonostachys rosea, and Humicola nigrescens. The average size of the inoculation site was ≥ 4.6 mm in F. oxysporum and F. solani, which were pathogenic to dry rot in potatoes but were not pathogenic compared to other strains up to 5 weeks in a 15 to 20 °C and 99% RH environment. The pathogenicity of F. foetens and F. pseudoanthophilum was related to a strong relationship by forming a single system with F. oxysporum. However, except for F. oxysporum and F. solani, these strains have not yet been reported to be associated with dry rot disease. Additionally, the length of the cross-section and longitudinal section of the potato damage symptom inoculated with C. rosea increased the most among all strains. This suggests that C. rosea is the dominant species involved in domestic potato dry rot disease. By contrast, there are no reports of the involvement of B. ranunculi and H. nigrescens in dry rot disease. Therefore, these strains can be seen as parasitic using potato sclerosis as nutrients in in vivo experiments through wounds and are not directly related to dry rot disease.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Potatoes belong to the Solanaceae family, with approximately 230 species found in natural ecosystems, among which approximately 160 species form tubers (Kwon et al. 2005). Today, Solanum tuberosum is grown worldwide, with approximately 388 million tons of tubers produced annually across 130 countries (Kim 2020), 690,419 MT (metric ton) of which were produced in Korea in 2019 (KOSIS 2021). As the highest source of energy and of production per unit area among the world’s major cultivated crops, potato production is expected to continue to increase (RDA 2020). However, potatoes are vulnerable to a variety of diseases when stored for long periods of time. For example, losses caused by dry rot disease are estimated to range from 6 to 25% worldwide, with losses of up to 60% during long-term storage periods (Desjardins 2006; Secor and Salas 2001). The estimated economic damage of crops lost in the United States, the largest potato producer, ranges from $100 million to $250 million per year (Slininger and Schisler 2002), and accounts for up to 88% of the total post-harvest losses of potatoes in Gansu, China (He et al. 2004). Despite a lack of accurate statistics or reports, most potatoes with pathological disorders during potato storage in Korea have been identified as having dry rot disease (data not shown).
Dry rot disease is caused by the intrusion of various Fusarium species through wounds on the surface of potatoes during their harvest (Nelson et al. 1981). This pathogen invades potatoes lodging in the soil and seeds and causes crop loss during storage (Tiwari et al. 2020). Starting as a small brown lesion, potato dry rot disease gradually expands, developing into a concentric dry region around dead tissue (RDA 2020). Subsequently, the wrinkled tissue changes from brown to black and can be distinguished with the naked eye. The lesion of the wrinkled tissue produces creamy white, pink, or orange spores and mycelium during long-term storage at temperatures ranging from 5 to 30 °C (Bojanowski et al. 2013; Elsherbiny et al. 2016).
Dry rot caused by Fusarium occurs in all potato cultivation areas, and the distribution of Fusarium species varies depending on the cultivation time and geographical location (Tiwari et al. 2020). More than 13 species, including F. acuminatum, F. avenaceum, F. crookwellense, F. culmorum, F. equiseti, F. graminearum, F. oxysporum, F. sambucinum, F. scirpi, F. semitectum, F. solani, F. sporotrichioides, and F. tricinctum, are involved in the dry rot disease of potatoes worldwide (Cullen et al. 2005). The dominant species that cause dry rot in potatoes are F. sambucinum and F. graminearum in North America, F. sambucinum in Europe, F. solani in the United Kingdom, F. solani and F. oxysporum in South Africa, and F. sambucinum in China (Tiwari et al. 2020). Until 2000, F. solani was suggested as a fungus that caused potato dry rot disease in Korea, and F. oxysporum was reported as a pathogen that caused withering diseases in various crops; however, this was not explained in connection with potato dry rot disease (Kim 2000; Park et al. 1988). Currently, dry rot disease pathogens in potatoes are registered as Fusarium spp. in the National Agricultural Products Disease and Pest Management System, and various Fusarium spp. have been reported to cause the disease (NCPMS 2023). In addition, one study showed that Clonostachys rosea, also known as Gliocladium roseum, is involved in dry rot disease and simultaneously acts as an antagonist to F. solani and F. oxysporum (Theron and Holz 1991).
Research has been conducted in North America, Europe, the UK, South Africa, and China to identify the dominant species of pathogens causing potato dry rot. However, few studies have identified the causative fungi of dry rot disease during storage or reported the dominant species in Korea. Therefore, this study was conducted to identify and confirm the pathogenicity of the causative pathogens of domestic potato dry rot disease in Korea.
Materials and Methods
Collection of Pathogens for Dry Rot of Potatoes
Tubers were collected from potatoes stored in traditional potato warehouses located on highland areas, including Wangsan-myeon and Daegwallyeong-myeon, and located on plain area, including Gangneung Wonju University’s farm and Sacheon-myeon in Gangwon Province. The potato varieties included Atlantic, Chubaek, Doobak, and Superior. The collected pathogens of potato were stored in a 20 °C incubator at Gangneung Wonju University’s Plant Pathology Laboratory for 3 days to induce growth.
Separation and Identification of Pathogens
Dry rot pathogens were isolated from the plant pathology laboratory of Gangneung Wonju University using Solannum tuberosum damage symptoms with signs of disease. To separate pathogens, fragments, including the surface of the potato damage symptom containing the boundary of the disease, were prepared by cutting into 1-cm diameter slices. The potato slices were sterilized in 70% ethanol for 30 s and 1% sodium hypochlorite (NaOCl) for 2 min on a clean bench, washed once with sterilized water, and dried on a sterilized filter paper for 30 min. One dried fragment from the infected damage symptom was placed in 10 WA (water agar medium) and cultured for 3 days in a 25 ℃ incubator (Fig. 1). Mycelia of pathogens grown in potato fragments were cultured in a 20 °C incubator for 7 days by cutting the tip of the pathogen for pure separation and putting on potato dextrose agar (PDA) medium. Each fungus was separated from the potato agar medium after confirming the shape of mycelial growth, and the separated samples were analyzed using the ITS1/ITS4 primer pair. The phylogenetic tree was constructed using the Neighbor-joining method with MEGA7.

Colony morphologies of the isolated fungal species on water agar medium incubated at 25 ℃ for 3 days from potatoes showing symptoms of dry rot disease on tubers stored in cold storages. A Beckjak; B Dano; C Doobak; D Jayoung; E Superior; F Wangsan
Pathogenicity Test of Dried Potato Rot Strain
The potatoes used in the pathogenicity test were superior varieties grown in Wangsan-myeon, Gangneung-si, and Gangwon-do. The 12 identified strains were used as inoculators. The potato samples were immersed in 1% NaOCl for 30 min, washed three times with sterile water after sterilization, and dried for 30 min. The inoculator prepared 12 identified pathogens by implanting them in PDA and culturing them in an incubator at 20 °C for 7 days. For the treatment of the inoculum, a 2-mm-deep hole was made in the base of the potato necrosis using a ø8mm cork borer, and the inoculum was cut and inserted with a ø10mm cork borer. Potatoes inoculated with the identified fungi were separated and cultured in a sealed plastic container to prevent air contamination by pathogens during culturing. The invasion and pathogenicity of pathogens through wounds were investigated by placing the fungally inoculated sclera in a humidified translucent plastic box and maintaining a relative humidity of 99% at room temperature of 15–20 °C (Table 1).
The inoculation site and pathogenicity were visually checked at intervals of 7 days after inoculation, and the length of disease spread was measured in the cross-section (Fig. 2) and longitudinal section (Fig. 3) of the potato damage symptom at 5 weeks after inoculation (Table 2). Each treatment port, including the control port, was performed five times based on 13 unusual diameters. Statistical analysis was performed using SPSS (IBM SPSS Statistics for Windows version 26.0 (IBM Corp., Armonk, NY, USA) to analyze ANOVA, and the significance of each treatment was verified at P < 0.05 using Duncan’s multiple range test (DMRT).

Disease symptoms of infected areas on potato tubers 7 weeks after the artificial inoculation of 12 fungal species incubated at 15 ~ 20 ℃ and 99% RH. A Control; B Fusarium boothii; C Fusarium circinatum; D Fusarium citricola; E Fusarium foetens; F Fusarium iranicum; G Fusarium longifundum; H Fusarium oxysporum; I Fusarium pseudoanthophilum; J Fusarium solani; K Botryotinia ranunculi; L Clonostachys rosea; M Humicola nigrescens

Appearance of the longitudinal sections of potato tubers 7 weeks after the artificial inoculation of 12 fungal species incubated at 15 ~ 20 ℃ and 99% RH. A Control; B Fusarium boothii; C Fusarium circinatum; D Fusarium citricola; E Fusarium foetens; F Fusarium iranicum; G Fusarium longifundum; H Fusarium oxysporum; I Fusarium pseudoanthophilum; J Fusarium solani; K Botryotinia ranunculi; L Clonostachys rosea; M Humicola nigrescens
Results and Discussion
Collection of Pathogens for Dry Rot in Potatoes
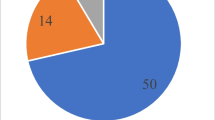
The total number of potatoes with dry rot disease after storage was 93, including 76 in Gangneung plain, 3 in Pyeongchang, 11 in Gangneung highland. To collect individuals with dry rot disease from potatoes, potato-only storage was investigated, and most potato objects with external diseases were infected with dry rot disease. There were a few cases of infection during storage, and it is believed that the spread of the disease was suppressed by screening and curing after harvest. Dry rot disease that occurred in certain varieties, such as Atlantic, Beckjak, Chubaek, Dano, Doobak, Jayoung, Superior, and Wangsan, showed typical symptoms of disease regardless of the variety (Fig. 4).

Potato tubers with symptoms of dry rot disease under common storage conditions (2021–2022). A Atlantic, B Beckjak, C Chubaek, D Dano, E Doobak, F Jayoung, G Superior, and H Wangsan
Separation and Identification of Pathogens
Of the 93 potatoes harbouring dry rot disease, some had duplicate or separate fungal strains. As such, a total of 76 were ultimately isolated (Table 1). After analyzing the sequence of the ITS region using the ITS1/ITS4 primer pair in the 76 separate fungal strains, a total of 12 strains were analyzed, including nine from Fusarium, one from Botryotinia, one from Clonostachys, and one from Humicola.
The nine strains of the Fusarium spp. were F. boothii, F. circinatum, F. citricola, F. foetens, F. iranicum, F. longifundum, F. oxysporum, F. pseudoanthophilum, and F. solani. Among the Fusarium, the most frequently isolated strains were F. foetens (16), F. citricola (14), and F. solani. Other Fusarium strains were separated in the following order: F. boothii, F. circinatum, F. iranicum, F. pseudoanthophilum, F. longifundum, and F. oxysporum. The Fusarium strains involved in dry rot disease, which are most commonly found in domestic potato stores, are believed to be F. foetens, F. citricola, and F. solani. The remaining strains were identified as Botryotinia ranunculi, Clonostachys rosea, and Humicola nigrescens (Fig. 5). Among these, F. solani, F. oxysporum, and C. rosea have been reported to cause dry rot disease (Bojanowski et al. 2013; Theron and Holz 1991). C. rosea is involved in dry rot disease and simultaneously acts as an antagonist of F. solani and F. oxysporum (Theron and Holz 1991), although there have been no reports of any other identified pathogens associated with dry rot disease. The following have been reported to be representative of diseases caused by each pathogen: F. boothii has been reported to be associated with corn ear rot, pecan malady (Gryzenhout et al. 2016) and wheat red mold disease (Wegulo et al. 2018); F. circinatum causes pine branch disease (Woo et al. 2011) and pine pitch canker (Maphosa et al. 2016); F. citricola causes tangerine branch ulcer disease (Sandoval-Denis et al. 2018); F. foetens is pathogenic to begonia (Schroers et al. 2004) and has been associated with the occurrence of damping-off of rooibos seedlings (Lamprecht and Tewoldemedhin 2017); F. iranicum has been identified in mushroom (Torbati et al. 2019) and wheat head light (Senatore et al. 2021); F. longifundum causes brown spots on the leaves of wild leafy vegetables (Matić et al. 2020); F. pseudoanthophilum is highly pathogenic in corn stalks in western Iran (Chehri et al. 2010) and is associated with Fusarium crown rot (Kazan and Gardiner 2018); B. ranunculi is a complete generation of Botrytis species, a plant pathogen that causes damage to economically important crops worldwide and is pathogenic to Ranunculaceae, which are distributed in warm and cold regions (Staats et al. 2005); H. nigrescens is a soil by-product of potato packaging and is known to be pathogenic to plants (UAMH 2023).

Colony morphology of the twelve isolated fungal species on potato dextrose agar medium incubated at 20 ℃ for 7 days from potato tubers infected dry rot in this study. A Fusarium boothii; B Fusarium circinatum; C Fusarium citricola; D Fusarium foetens; E Fusarium iranicum; F Fusarium longifundum; G Fusarium oxysporum; H Fusarium pseudoanthophilum; I Fusarium solani; J Botryotinia ranunculi; K Clonostachys rosea; L Humicola nigrescens
In the phylogenic tree, which showed a close relationship between 12 strains as a result of identification, F. solani, generally known as the main cause of dry rot disease, was found to be far from related among other Fusarium spp. (Fig. 6). F. oxysporum (Kwon et al. 2005) is considered another major cause of dry rot disease in Korea and exhibits a high relationship through the formation of a single system with F. foetens, F. pseudoanthophilum, and F. circinatum. In this study, F. acuminatum, F. avenaceum, F. crookwellense, F. culmorum, F. equiseti, F. sambucinum, F. scirpi, F. semitectum, F. sporotrichioides, and F. tricinctum were not investigated in Fusarium (Cullen et al. 2005). F. sambucinum (Tiwari et al. 2020), known as a common or dominant species in North America, Europe, and China, was not identified among the isolated strains. These results suggest that F. sambucinum is not common in Korea among the various types of Fusarium strains that do not cause dry rot disease.

Phylogenetic relationship between Fusarium spp. and fungi isolated from potatoes from 76 fungi of 93 tubers showing symptoms of dry rot disease based on ITS region gene sequence analysis
Pathogenicity Test of Dried Potato Rot Strain
A pathogenic test of the 12 strains identified separately showed no signs of disease in the control potatoes, while all potatoes inoculated with the 12 strains in vivo into the sclera of the Sumi variety showed dry rot disease with a cross-sectional length of ≥ 3.5 mm and a longitudinal length of ≥ 2.9 mm. The average size of the inoculation site was ≥ 4.6 mm in cross-sectional length in the F. oxysporum and F. solani treatment zones, which are known to be involved in potato dried rot in Korea. When comparing the cross-sectional length of the two strains known as the main fungi of dry rot disease with 3.8–5.4 mm, the cross-sectional length of F. citricola, F. foetens, F. longifundum, and F. pseudoanthophilum, pathogenicity was found to have similar diseases with no statistically significant difference. However, F. oxysporum and F. solani had pathogenic properties of dry rot disease in potatoes, but were not as pathogenic as other strains until 5 weeks in a suitable environment (15–20 ℃, 99% RH). It is possible that the F. oxysporum and F. solani strains were used after more than three series of cultures, which reduced their vitality. In addition, the optimal mycelium growth temperature of F. oxysporum and F. solani is 24–25 ℃ (Ezrari et al. 2021; Hibar et al. 2006; Jeon et al. 2013), and it is believed that mycelium growth was relatively inactive at 15–20 ℃, which is the condition used in this experiment. Among Fusarium-based pathogens, F. foetens and F. pseudoanthophilum are known to exhibit high relationship through their formation of a single system with F. oxysporum. However, strains other than F. oxysporum and F. solani have not yet been reported to be associated with dry rot disease, and there is a need to determine whether these strains develop dry rot disease. The cross-sectional length of the potato sclera inoculated with C. rosea was 11.48 mm and the longitudinal length was 6.9 mm, which represents the greatest disease growth among all strains. The length of the longitudinal section of all strains used in the experiment was approximately 2.9–4.8 mm in other treatment areas except for potatoes inoculated with C. rosea, and there was no statistically significant difference. The results of this study are similar to those of Theron and Holz (1991), who reported that C. rosea is also involved in dry rot disease, and it is thought that C. rosea may be a dominant species involved in domestic potato dry rot disease.
In the pathogenicity test, traces of strain invasion were observed in B. ranunculi and H. nigrescens. There are no reports on the involvement of B. ranunculi and H. nigrescens in dry rot disease. In particular, H. nigrescens is known to be non-pathogenic due to soil by-products, and in this experiment, it appeared to have been invaded through potato sclerosis. This suggests that B. ranunculi and H. nigrescens exhibited parasitic behaviour, using potato sclera as nutrients in through wounds in vivo, and indicates that they are not directly related to the development of dry rot disease.
References
Bojanowski A, Avis TJ, Pelletier S, Tweddell RJ (2013) Management of potato dry rot. Postharvest Biol Technol 84:99–109
Chehri K, Zafari D, Nurhazrati M, Salleh B, Reddy KRN, Karami E (2010) Natural occurrence of Fusarium species associated with root and stalk rot of maize in Kermanshah Province, Iran. J Biol Sci 10:795–799
Cullen DW, Toth IK, Pitkin Y, Boonham N, Walsh K, Barker I, Lees AK (2005) Use of quantitative molecular diagnostic assays to investigate Fusarium dry rot in potato stocks and soil. Phytopathology 95:1462–1471. https://doi.org/10.1094/PHYTO-95-1462
Desjardins AE (2006) Fusarium mycotoxins: chemistry, genetics, and biology. American Phytopathological Society, St Paul, MN
Elsherbiny EA, Amin BH, Baka ZA (2016) Efficiency of pomegranate (Punica granatum L.) peels extract as a high potential natural tool towards Fusarium dry rot on potato tubers. Postharvest Biol Technol 111:256–263
Ezrari S, Radouane N, Tahiri A, Amiri S, Lazraq A, Lahlali R (2021) Environmental effects of temperature and water potential on mycelial growth of Neocosmospora solani and Fusarium spp. causing dry root rot of citrus. Curr Microbiol 78:3092–3103. https://doi.org/10.1007/s00284-021-02570-1
Gryzenhout M, Khooa B, Landman L (2016) First report of Fusarium boothii from pecan (Carya illinoinensis) and camel thorn (Vachellia erioloba trees in South Africa. S Afr J Bot 105:158–162. https://doi.org/10.1016/j.sajb.2016.03.003
He SQ, Jin XL, Wei ZQ, Zhang TY, Du X, Luo DG (2004) Isolates and identification of pathogens causing dry rot of potato tubers in Ding Xi prefecture of Gansu province. J Yunnan Agric Univ (Nat Sci) 19:550–552
Hibar K, Daami-Remadi M, Jabnoun-Khiareddine H, El Mahjoub M (2006) Temperature effect on mycelial growth and on disease incidence of Fusarium oxysporum f.sp. radicis-lycopersici. Plant Pathol J 5:233–238. https://doi.org/10.3923/ppj.2006.233.238
Jeon CS, Kim GH, Son KI, Hur JS, Jeon KS, Yoon JH, Koh YJ (2013) Root rot of balloon flower (Platycodon grandiflorum) caused by Fusarium solani and Fusarium oxysporum. Plant Pathol J 29:440–445. https://doi.org/10.5423/PPJ.NT.07.2013.0073
Kazan K, Gardiner DM (2018) Fusarium crown rot caused by Fusarium pseudograminearum in cereal crops: recent progress and future prospects. Mole Plant Pathol 19:1547–1562. https://doi.org/10.1111/mpp.12639
Kim JW (2000) Plant pathology. Daegu University Press, Daegu
Kim GW (2020) Evaluation of 45 potato genetic resources for application of new potato varieties with excellent processing characteristics. Thesis of Master Degree, Graduate School of Kangwon National University, Korea
Korean Statistical Information Service (KOSIS) (2021) Agricultural production (potatoes and sweet potatoes). Korea. https://kosis.kr/statHtml/statHtml.do?orgId=101&tblId=DT_2KAA406_OECD. Accessed 20 Mar 2022
Kwon M, Kim SY, Kim CG, Kim JS, Kim HJ, Ryu KY, Park YE, Park, CS et al (2005) Potato book. National Institute of Highland Agriculture, Rural Development Administration (RDA), Korea
Lamprecht SC, Tewoldemedhin YT (2017) Fusarium species associated with damping-off of rooibos seedlings and the potential of compost as soil amendment for disease suppression. S Afr J Bot 110:110–117
Maphosa MN, Steenkamp ET, Wingfeld BD (2016) Genome-based selection and characterization of Fusarium circinatum-specifc sequences. G3 6:631–639. https://doi.org/10.1534/g3.115.025817
Matić S, Tabone G, Guarnaccia V, Gullino ML, Garibaldi A (2020) Emerging leafy vegetable crop diseases caused by the Fusarium incarnatum-equiseti species complex. Phytopathol Medit 59:303–317. https://doi.org/10.14601/Phyto-10883
National Crop Pest Management System (NCPMS) (2023) Dry rot of potato. RURAL Development Administration, Korea. https://ncpms.rda.go.kr/npms/ImageSearchInfoR1.np?detailKey=D00000075. Accessed 5.1.2023
Nelson PE, Toussoun TA, Cook RJ (1981) Fusarium: diseases, biology and taxonomy. The Pennsylvania State University Press, University Park, PA
Park JS, Kwon OK, Kim KC, Kim MS, Kim MH, Kim JS, Park KH, Seo IS et al (1988) Plant pathology. Hyangmoonsa, Korea
Rural Development Administration (RDA) (2020) Agricultural technology guide (Potatoes). Korea. http://www.nongsaro.go.kr/portal/ps/psb/psbx/cropEbookFileViewPop.ps?indexPage=26&indexBasePage=0&cropsEbookFileNo=00001&ebookCode=20. Accessed 20 Mar 2022
Sandoval-Denis M, Guarnaccia V, Polizzi G, Crous PW (2018) Symptomatic citrus trees reveal a new pathogenic lineage in Fusarium and two new Neocosmospora species. Persoonia 40:1–25. https://doi.org/10.3767/persoonia.2018.40.01
Schroers HJ, Baayen RP, Meffert JP, de Gruyter J, Hooftman M, O’Donnell K (2004) Fusarium foetens, a new species pathogenic to begonia elatior hybrids (Begonia x hiemalis) and the sister taxon of the Fusarium oxysporum species complex. Mycologia 96:393–406
Secor GA, Salas B (2001) Fusarium dry rot and Fusarium wilt. In WR Stevenson, R Loria, GD Franc, DP Weingartner, eds, Compendium of potato diseases. Ed 2. American Phytopathological Society. St Paul, MN, USA, pp 23–25
Senatore MT, Ward TJ, Cappelletti E, Beccari G, McCormick SP, Busman M, Laraba I, O’Donnell K, Prodi A (2021) Species diversity and mycotoxin production by members of the Fusarium tricinctum species complex associated with Fusarium head blight of wheat and barley in Italy. Intl J Food Microbiol 358:109298. https://doi.org/10.1016/j.ijfoodmicro.2021.109298
Slininger PJ, Schisler DA (2002) Spray-on fungi stop potato rot fungus. United States Department of Agriculture (USDA). AgResearch Magazine, USA. https://agresearchmag.ars.usda.gov/2002/jun/fungus. Accessed 5.1.2023
Staats M, van Baarlen P, van Kan JAL (2005) Molecular phylogeny of the plant pathogenic genus Botrytis and the evolution of host specificity. Mole Biol Evol 22:333–346. https://doi.org/10.1093/molbev/msi020
Theron DJ, Holz G (1991) Dry rot of potatoes caused by Gliocladium roseum. Plant Pathol 40:302–305
Tiwari RK, Kumar R, Sharma S, Sagar V, Aggarwal R, Naga KC, Lal MK, Chourasia KN, Kumar D, Kumar M (2020) Potato dry rot disease: current status, pathogenomics and management. Biotech 10:503. https://doi.org/10.1007/s13205-020-02496-8
Torbati M, Arzanlou M, Sandoval-Denis M, Crous PW (2019) Multigene phylogeny reveals new fungicolous species in the Fusarium tricinctum species complex and novel hosts in the genus Fusarium from Iran. Mycol Prog 18:119–133. https://doi.org/10.1007/s11557-018-1422-5
University of Alberta Microfungus Herbarium (UAMH) (2023) Centre for Global Microfungal Biodiversity, Dalla Lana School of Public Health, University of Toronto, Canada. https://www.uamh.ca/details.php?id=3078. Accessed 5.1.2023
Wegulo SN, Valverde-Bogantes E, Bolanos-Carriel C, Hallen-Adams H, Bianchini A, McMaster N, Schmale DG III (2018) First report of Fusarium boothii causing head blight of wheat in the United States. Plant Dis 102:2646–2646
Woo KS, Yoon JH, Han SU, Woo SY (2011) Effects of Fusarium circinatum on disease development and gas exchange in the seedlings of Pinus spp. Res Plant Dis 17:177–183. https://doi.org/10.5423/RPD.2011.17.2.177
Funding
This work was carried out with the support of “Cooperative Research Program for Agriculture Science and Technology Development (Project No. RS-2021-RD009975)” Rural Development Administration, Republic of Korea.
Author information
Authors and Affiliations
Contributions
NS Kim: investigation and data curation. SJ Hong: supervision, validation, writing, review, and editing. HS Won: review. BS Kim: conceptualization and methodology. SH Gwon: investigation, data curation, writing, and review.
Corresponding authors
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Communicated by Se Hwi Gwon
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kim, N.S., Hong, S.J., Won, H.S. et al. Identification and Pathogenicity of Species Isolated from Stored Potato Tubers Showing Symptoms of Dry Rot Disease. Potato Res. (2024). https://doi.org/10.1007/s11540-024-09709-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11540-024-09709-0