
Abstract
Modeling under the framework of ecological stoichiometric allows the investigation of the effects of food quality on food web population dynamics. Recent discoveries in ecological stoichiometry suggest that grazer dynamics are affected by insufficient food nutrient content (low phosphorus (P)/carbon (C) ratio) as well as excess food nutrient content (high P:C). This phenomenon is known as the “stoichiometric knife edge.” While previous models have captured this phenomenon, they do not explicitly track P in the producer or in the media that supports the producer, which brings questions to the validity of their predictions. Here, we extend a Lotka–Volterra-type stoichiometric model by mechanistically deriving and tracking P in the producer and free P in the environment in order to investigate the growth response of Daphnia to algae of varying P:C ratios. Bifurcation analysis and numerical simulations of the full model, that explicitly tracks phosphorus, lead to quantitative different predictions than previous models that neglect to track free nutrients. The full model shows that the fate of the grazer population can be very sensitive to excess nutrient concentrations. Dynamical free nutrient pool seems to induce extreme grazer population density changes when total nutrient is in an intermediate range.






Similar content being viewed by others
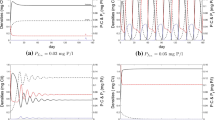
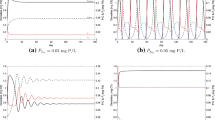

References
Andersen T (1997) Pelagic nutrient cycles: herbivores as sources and sinks. Springer, New York
Bennett EM, Carpenter SR, Caraco NF (2001) Human impact on erodable phosphorus and eutrophication: a global perspective. Bioscience 51(3):227–234
Boersma M, Elser J (2006) Too much of a good thing: on stoichiometrically balanced diets and maximal growth. Ecology 87:1325–1330
Cease A, Elser J, Ford C, Hao S, Kang L, JF H (2012) Heavy livestock grazing promotes locust outbreaks by lowering plant nitrogen content. Science 335:467–469
Demott W, Gulati R, Siewertsen K (1998) Effects of phosphorus-deficient diets on the carbon and phosphorus balance of Daphnia magna. Limnol Oceanogr 43:1147–1161
Diehl S (2007) Paradoxes of enrichment: effects of increased light versus nutrient supply on pelagic producer–grazer systems. Am Nat 169(6):E173–E191
Elser J, Bennett E (2011) Phosphorus cycle: a broken biogeochemical cycle. Nature 478:29–31
Elser J, Hayakawa K, Urabe J (2001) Nutrient limitation reduces food quality for zooplankton: Daphnia response to seston phosphorus enrichment. Ecology 82:898–903
Elser J, Watts J, Schampell J, Farmer J (2006) Early Cambrian food webs on a trophic knife-edge? A hypothesis and preliminary data from a modern stromatolite-based ecosystem. Ecol Lett 9:295–303
Elser JJ, Loladze I, Peace AL, Kuang Y (2012) Lotka re-loaded: modeling trophic interactions under stoichiometric constraints. Ecol Model 245:3–11
Frost P, Benstead J, Cross W, Hillebrand H, Larson J, Xenopoulos M, Yoshida T (2006) Threshold elemental ratios of carbon and phosphorus in aquatic consumers. Ecol Lett 9:774–779
Gaxiola RA, Edwards M, Elser JJ (2011) A transgenic approach to enhance phosphorus use efficiency in crops as part of a comprehensive strategy for sustainable agriculture. Chemosphere 84(6):840–845
Hessen DO, Elser JJ, Sterner RW, Urabe J (2013) Ecological stoichiometry: an elementary approach using basic principles. Limnol Oceanogr 58(6):2219–2236
Li X, Wang H, Kuang Y (2011) Global analysis of a stoichiometric producer–grazer model with holling type functional responses. J Math Biol 63(5):901–932
Loladze I, Kuang Y, Elser J (2000) Stoichiometry in producer–grazer systems: linking energy flow with element cycling. Bull Math Biol 62:1137–1162
Markus L (1956) Asymptotically autonomous differential systems. In: Contributions to the theory of nonlinear oscillations III, vol 36, pp 17–29
Peace A, Zhao Y, Loladze I, Elser JJ, Kuang Y (2013) A stoichiometric producer–grazer model incorporating the effects of excess food-nutrient content on consumer dynamics. Math Biosci 244(2):107–115
Rosenzweig ML (1971) Paradox of enrichment: destabilization of exploitation ecosystems in ecological time. Science 171(3969):385–387
Rosenzweig ML, MacArthur RH (1963) Graphical representation and stability conditions of predator–prey interactions. Am Nat 97(895):209
Smith V, Schindler D (2009) Eutrophication science: where do we go from here? Trends Ecol Evol 24:201–207
Sterner RW, Andersen T, Elser JJ, Hessen DO, Hood JM, McCauley E, Urabe J (2008) Scale-dependent carbon: nitrogen: phosphorus seston stoichiometry in marine and freshwaters. Limnol Oceanogr 53(3):1169
Stiefs D, Van Voorn GA, Kooi BW, Feudel U, Gross T (2010) Food quality in producer–grazer models: a generalized analysis. Am Nat 176(3):367–380
Thieme HR (1992) Convergence results and a poincaré-bendixson trichotomy for asymptotically autonomous differential equations. J Math Biol 30(7):755–763
Urabe J, Sterner RW (1996) Regulation of herbivore growth by the balance of light and nutrients. Proc Nat Acad Sci 93(16):8465–8469
Wang H, Kuang Y, Loladze I (2008) Dynamics of a mechanistically derived stoichiometric producer–grazer model. J Biol Dyn 2(3):286–296
Wang H, Sterner RW, Elser JJ (2012) On the strict homeostasis assumption in ecological stoichiometry. Ecol Model 243:81–88
Acknowledgments
This work was supported in part by NSF DMS-0920744 and NSERC Discovery Grant RES0001528. The authors thank two anonymous reviewers for their comments that improved this manuscript.
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix 1: Remaining Cases for Proof of Theorem 3.1
Proof
Case 2: \(x(t_1)=0\)
Let \(\overline{f}=f'(0)=\underset{x\rightarrow 0}{\lim }\frac{f(x)}{x}\) and \(\overline{y}=\underset{t\in [0,t_1]}{\max }y(t) <\frac{P}{\theta }\). Then for every \(t\in [0,t_1]\),
This implies that \(x(t_1)\ge x(0)e^{\alpha t_1} > 0\), where \(\alpha \) is a constant. This contradicts \(x(t_1)=0\) and proves that \(S(t_1)\) does not reach this boundary.
Case 3: \(y(t_1)=0\)
Then, for every \(t\in [0,t_1]\),
This implies that \(y(t_1)\ge y(0)e^{-\hbox {d}t_1}>0\). This contradicts \(y(t_1)=0\) and proves that \(S(t_1)\) does not reach this boundary.
Case 4: \(Qx+\theta y =P\)
Since \(v(P-Q(t_1)x(t_1)-\theta y(t_1))=0\)
Thus, \(S(t_1)\) can not cross this boundary.
Case 5: \(x(t_1)=K\)
Then, for every \(t\in [0,t_1]\),
Then, \(x(t_1)\le K\) by a standard comparison argument, thus \(S(t_1)\) can not cross this boundary.
Case 6: \(Q(t_1)=\hat{Q}\)
Since \(v(P-Q(t_1)x(t_1)-\theta y(t_1),Q(t_1))=0\)
Thus, \(S(t_1)\) can not cross this boundary.\(\square \)
Appendix 2: Proof of Theorem 3.2
Proof
To prove that \(E_0\) is unstable, it is sufficient to show the system linearized at this equilibrium has an eigenvalue whose real part is positive. This is seen in the following Jacobian,
where \(b\left( 1-\frac{q}{Q_0}\right) >0\).\(\square \)
Appendix 3: Proof of Lemma 3.1
Proof
We consider two cases (\(1-\frac{x}{k}<1-\frac{q}{Q}\) and \(1-\frac{x}{k}>1-\frac{q}{Q}\)).
Case 1: \(1-\frac{x}{k}<1-\frac{q}{Q}\)
In this case, (Eq. 15a) becomes
and \(x_1=k\). (Eq. 15c) becomes
therefore, \(v(P-Q_1k, Q_1)=0\). There are two cases to consider here (\(\frac{P}{k}>\hat{Q}\) and \(\frac{P}{k}<\hat{Q}\) ). If \(\frac{P}{k}>\hat{Q}\), then \(Q_1=\hat{Q}\) to remain in \(\Omega \). Since \(P\ge Q_1x_1=Q_1k\), the case when \(\frac{P}{k}<\hat{Q}\) results in \(\hat{Q}>\frac{P}{k}\ge Q_1\), thus \(Q_1=\frac{P}{k}\). The two cases are summarized below
Case 2: \(1-\frac{x}{k}>1-\frac{q}{Q}\) In this case, (Eq. 15a) becomes
and \(Q_1=q\). (Eq. 15c) becomes
therefore \(v(P-qx_1, q)=0\) and thus \(x_1=\frac{P}{q}\).
To show that the two equilibrium forms cannot coexist, we need to show that they satisfy two opposite conditions.
In case 1: \(1-\frac{x}{k}<1-\frac{q}{Q}\) and \(E_1=\left( k,0,\min \left\{ \frac{P}{k},\hat{Q}\right\} \right) \), therefore
Here
In case 2: \(1-\frac{x}{k}>1-\frac{q}{Q}\) and \(E_1=\left( \frac{P}{q},0,q\right) \), therefore
The two cases follow opposite conditions. Actually, when \(\frac{P}{k}=q\), the two forms of \(E_1\) collide to \((k,0,q)\).\(\square \)
Appendix 4: Proof of Theorem 3.3
Proof
Assume that \(\min \left\{ \hat{e}f(x_1),\frac{Q_1}{\theta }f(x_1),\hat{e}\hat{f}\frac{\theta }{Q_1}\right\} <d\). To prove that \(E_1\) is stable, we consider two cases (\(1-\frac{x}{k}<1-\frac{q}{Q}\) and \(1-\frac{x}{k}>1-\frac{q}{Q}\)) where we look at the linearized system and use the Routh–Hurwitz criterion.
Case 1: \(1-\frac{x}{k}<1-\frac{q}{Q}\)
Here, \(E_1=\left( k,0,\min \left\{ \frac{P}{k},\hat{Q}\right\} \right) \) by Lemma 3.1 and the Jacobian takes the following form,
Let \(\alpha _1 = \min \left\{ \hat{e}f(k),\frac{\min \left\{ \frac{P}{k},\hat{Q}\right\} }{\theta }f(k),\hat{e}\hat{f}\frac{\theta }{\min \left\{ \frac{P}{k},\hat{Q}\right\} }\right\} -d<0\) and \(\alpha _2 = \frac{\hbox {d}v}{\hbox {d}Q} \big |_{E_1}<0\). Then, the Jacobian simplifies to
The characteristic equation may be written
The eigenvalues of \(J(E_1)\) are \(-b, \alpha _1, \alpha _2\), which are all negative.
Case 2: \(1-\frac{x}{k}>1-\frac{q}{Q}\)
Here, \(E_1=(\frac{P}{q},0,q)\) by Lemma 3.1 and the Jacobian takes the following form,
Let \(\alpha _1=\min \left\{ \hat{e}f(\frac{P}{q}),\frac{\hat{q}}{\theta }f(\frac{P}{q}),\hat{e}\hat{f}\frac{\theta }{q}\right\} -d<0\), \(\alpha _2=\frac{\hbox {d}v}{\hbox {d}Q}\big |_{E_1}-b<0\), \(\alpha _3=\frac{\hbox {d}v}{\hbox {d}x}\big |_{E_1}<0\). Then the Jacobian simplifies down to
The characteristic equation may be written
Since \(\alpha _1, \alpha _2,\alpha _3<0\) we find that \(-\alpha _1-\alpha _2>0\), \(\frac{Pb}{q^2}\alpha _1\alpha _3>0\), and \((-\alpha _1-\alpha _2)(\alpha _1\alpha _2-\frac{Pb}{q^2}\alpha _3)>\frac{Pb}{q^2}\alpha _1\alpha _3\). These are the conditions of the Routh–Hurwitz criterion that guarantee all the eigenvalues of \(J(E_1)\) have strictly negative real parts. Thus, \(E_1\) is locally asymptotically stable for both cases.\(\square \)
Appendix 5: Proof of Theorem 3.4
Proof
The set \(\Omega \) is positively invariant under System (15) by Lemma 3.1. Let
For all \((x,y,Q)\in \Omega \), the expression for \(y'\) may be expressed as
This implies that \(\underset{t \rightarrow \infty }{\lim }y(t)=0\). In autonomous System (15), \(y(t)\) converges to 0. We may consider the behavior of System (15) on the plane \(y=0\) with the limit system
defined on the domain
System (27) is the limiting system of the asymptotically autonomous System (15) under the constraint \(\min \left\{ \hat{e}f(K),\frac{\hat{Q}}{\theta }f(K),\hat{e}\hat{f}\frac{\theta }{q}\right\} -d\). Results from Markus (1956) and Thieme (1992) allow us to compare solutions of an autonomous system with those of the asymptotically autonomous limit system. System (27) has one equilibrium \(\bar{E}_1 = (\bar{x}_1, \bar{Q}_1)\), and this equilibrium is globally asymptotically stable. To show this global stability, we consider two cases (\(1-\frac{x}{k}<1-\frac{q}{Q}\) and \(1-\frac{x}{k}>1-\frac{q}{Q}\)) where we look at the linearized system and then consider the existence of periodic orbits.
Case 1: \(1-\frac{x}{k}<1-\frac{q}{Q}\)
Here, \(\bar{E_1}=\left( k,\min \left\{ \frac{P}{k},\hat{Q}\right\} \right) \) and the Jacobian takes the form,
The eigenvalues are \(-b<0\) and \(\frac{\hbox {d}v}{\hbox {d}Q}\big |_{\bar{E_1}}<0\).
Case 2: \(1-\frac{x}{k}>1-\frac{q}{Q}\)
Here, \(\bar{E_1}=(\frac{P}{q},q)\) and the Jacobian takes the form,
Here, trace\((J(\bar{E}_1))=\frac{\hbox {d}v}{\hbox {d}Q}\big |_{\bar{E_1}}-b<0\) and det\((J(\bar{E}_1))= -b\frac{P^2}{q^2}\frac{\hbox {d}v}{\hbox {d}x}\big |_{\bar{E_1}}>0\). In both cases, \(\bar{E}_1\) is locally asymptotically stable. To show that no periodic orbits exist in \(\bar{\Omega }\), consider
Therefore, there is no \(t_1\) such that \(Q'(t_1)<0\) and \(x'(t_1)<0\); hence, there can not be any periodic solutions. Since \(\bar{\Omega }\) is simply connected and is positively invariant under System (27) and contains no periodic orbits, by the Poincaré–Bendixson Theorem, all solutions of System (27) starting in \(\bar{\Omega }\) will converge to \(\bar{E}_1\). Thus, \(\bar{E}_1\) is globally asymptotically stable. The \(\omega -\)limit set of a forward bounded solution of the autonomous System (15) consists of the equilibrium of its limit autonomous System (27) (Thieme 1992). Thus, the \(\omega -\)limit set of System (15) is \(\{{E}_1\}\). The grazer only extinction equilibrium \(E_1\) is globally asymptotically stable,
\(\square \)
Appendix 6: Supplementary Material Notes
An animation of the three dimensional phase space is provided in the supplementary material. The animation was created using MATLAB. Here, we used the Holling-type II function \(f(x)=\frac{\hat{f}x}{a+x}\) for the grazer ingestion rate and the parameter values listed in Table 1 and P varying from 0.001 to 0.14 mg P/L. The surfaces are the producer (blue), grazer (yellow), and producer P:C (red) nullsurfaces. The intersections of these surfaces depict equilibria. The equilibria that simulations suggest are stable are labeled with black dots; equilibria that simulations suggest are unstable are labeled with white dots.
Rights and permissions
About this article

Cite this article
Peace, A., Wang, H. & Kuang, Y. Dynamics of a Producer–Grazer Model Incorporating the Effects of Excess Food Nutrient Content on Grazer’s Growth. Bull Math Biol 76, 2175–2197 (2014). https://doi.org/10.1007/s11538-014-0006-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11538-014-0006-z