
Abstract
The in vitro gastric digestibility of the quinoa variety Riobamba was investigated, especially the influence of the quinoa matrix. Dry-fractionated quinoa protein concentrate, which is just milled and sieved, was much better digestible than the same concentrate that was reconstituted from wet fractionated quinoa protein isolate, quinoa starch isolate, and quinoa fibre isolate. In the reconstituted concentrate, the presence of quinoa starch and fibre next to quinoa protein reduces its in vitro gastric digestibility significantly. However, the effect of starch is partially counteracted if the fibre is also present. While the effects of starch and fibre separately can be understood from the decrease of the accessibility for pepsin to hydrolyse proteins, due to the hydrated starch and fibre, we suspect that the synergistic effect of starch and fibre may be due to a relative reduction of the hydration of starch due to the presence of the also strongly hydrating fibre. We concluded that the presence of starch and fibre decreases the protein in vitro gastric digestibility. Therefore, the presence of fibre partially countered the decreased of the protein digestibility of starch. Heating of the matrices to 120 °C generally resulted in much lower digestion rates, due to extensive aggregation of the protein.
Similar content being viewed by others



Avoid common mistakes on your manuscript.
Introduction
Quinoa (Chenopodium quinoa Willd.), a pseudocereal that is native to South America, has attracted much research interest lately, not only for its nutritional profile but also for its adaptability to different growing conditions [1, 2]. Quinoa has a relatively high protein content (14–16%) [3], and presents a wider amino acid spectrum than cereals and legumes [4], with higher lysine (5.1–6.4%) and methionine (0.4–1.0%) contents [5]. This makes quinoa complementary to cereals and legumes [6].
The major fraction of proteins in quinoa are 11S globulin and 2S albumin, representing around 72–77% of the total protein [7, 8]. The molar mass of 11S globulin is 22–23 kDa for the basic subunit and 32–39 kDa for the acid subunit; the molar mass of 2S albumin is 8–9 kDa [9]. The remainder of the proteins is composed mainly of prolamines [10].
Starch is the major component of quinoa seeds, making up more than 50% of the dry weight [11, 12]. The starch granules are rather small (1–3 μm) and are usually located in the perisperm of the seed [2, 13, 14]. The starch is mainly constituted of amylose and amylopectin; amylose being a linear glucosyl chain connected by an α-1,4 linkage, while amylopectin is highly branched by α-1,6 linkages in a clustered manner [15]. Starch may influence the digestion of proteins, by taking up gastric fluid and physically hindering the ingression of acid and pepsin into the protein.
The fibre content of quinoa is known to be in the between 1.3 and 6.1 wt% [16]. Dietary fibre comes from the carbohydrate parts of the plant cells that are resistant to enzymatic human digestion [17]. By forming a viscous gel-like substance in the digestive system, fibre can slow the transit time of nutrients through the intestines and shield these nutrients from digestion. Besides, evidence suggests that fibre can inhibit the absorption of sugar, cholesterol and various minerals, which may also affect the absorption of the protein [18].
Generally, quinoa is used as food in the same way as most cereals. Quinoa can be served plain, as a side dish or incorporated into other recipes. Quinoa seeds can be milled into flour. Alternatively a flour with a high protein content can be obtained via dry fractionation (coarse grinding and subsequently sieving) [19, 20]. However, saponins located in the pericarp (seed coats) of quinoa seeds impede its utilization as a practical and commercially attractive food source [21]. Saponins are triterpenoid glucoside compounds found in many plant genera that possess pharmacological properties [22], but most saponins have an intensely bitter flavour and all are potentially toxic if ingested in large quantities [19]. The levels of the saponins are highly variable among different quinoa varieties and, in accordance with the saponin concentration, quinoa varieties can be classified into sweet quinoa containing <0.11 wt% of saponins and bitter quinoa containing >0.11 wt% of saponins [23]. Saponins have to be removed by rinsing out with much water, which makes wet processing unattractive and is incompatible with dry processing because it needs to be dried beforehand. Dry processing represents a more sustainable alternative since it does not use water, requires much less energy and utilises more of the raw material in high-value fractions [24], in contrast to the wet fractionation which involves large amounts of water, chemicals and energy.
From 1999 three sweet varieties have been registered: Atlas (1999), Pasto (2005) and Riobamba (2005). All these varieties have the potential to be dry fractionated. Dry fractionation to concentrate proteins is more resource efficient, but yields fractions which still contain significant levels of components such as oil, starch and fibres. These components may influence the digestibility of the protein.
A protein with high digestibility has potentially better nutritional value than one with low digestibility because it provides more amino acids for absorption on proteolysis [25]. It is known that exogenous (interaction of proteins with non-protein components like polyphenols, non-starch polysaccharides, starch, tannins, dietary fibre, phytates and lipids) and endogenous factors (changes within the proteins themselves) contribute to poor digestibility of proteins [25]. During the process of milling and cooking, proteins may interact with non-protein components and the proteins themselves, thereby affecting their digestibility [26].
We have previously estimated the effect of starch on protein digestibility of a bitter quinoa variety [20] and characterized the protein-enriched fractions of two dry fractionated sweet quinoa varieties [27]. However, the effects of fibre and the combined effect of starch and fibre on protein digestibility of quinoa are not yet known. Thus, the aim of this work is to assess the effects of starch and fibre together and separately on the in vitro gastric digestion of quinoa proteins suspensions. of the quinoa sweet variety Riobamba. These protein suspensions are unheated or preheated at different temperatures (60 and 120 °C).
Materials and Methods
Materials
The quinoa seeds (variety Riobamba) were acquired from GreenFood50 (The Netherlands). Pepsin form porcine gastric mucosa and all other chemicals were purchased from Sigma-Aldrich, Inc. (St. Louis, MO, U.S.A.).
Material Preparation
Quinoa seeds were dry fractionated to concentrate protein and starch by milling and subsequently sieving according to the procedure described by Opazo-Navarrete et al. [27]. A protein-enriched flour was obtained by separation of the cotyledons from the seed and subsequently sieving using a mesh size of 630 and 315 μm, while starch was concentrate using a mesh size of 800 μm. In turn, quinoa protein was isolated from whole seeds according to the procedure described by Opazo-Navarrete et al. [27].
Fibre and starch were isolated from whole quinoa flour. After defatting, the defatted quinoa flour was suspended in Milli-Q water (1:3 w/w) and stirred for 4 h at 20 °C. The mix was sieved using three consecutive sieves with a mesh size of 212, 90 and 53 μm, respectively. The residues within the mesh size 90–53 μm were separated and oven dried at 80 °C for 4 h in order to obtain a fibre concentrate [28]. The remaining suspension after sieving was centrifuged at 500 g for 1 min at 20 °C in order to sediment the hulls. The resulting supernatant was centrifuged at 3000 g for 8 min at 20 °C obtaining a crude starch as a product. The crude starch was suspended in 0.05 M NaOH, stirred for 48 h at 20 °C and centrifuged at 3000 g for 8 min at 20 °C. After a white layer was obtained and removed with a spoon, which was suspended in water and centrifuged again (3000 g, 8 min, 20 °C). This was repeated four times in order to obtain high purity. The resulting starch was dried at 40 °C and 40 mbar in a vacuum oven (Binder VD53, Tuttlingen, Germany).
Compositional Analysis
The protein content was measured by Dumas analysis (Nitrogen analyser, FlashEA 1112 series, Thermo Scientific, Interscience, Breda, The Netherlands) in triplicate. A conversion factor of 5.85 was used to convert nitrogen values to protein. The oil content was determined with a fully automated Büchi extraction system B-811 (Büchi Labortechnik AG, Flawil, Switzerland). The oil extraction was performed with petroleum ether (boiling range 40–60 °C) in Standard Soxhlet mode for 3 h with a sample-to-solvent ratio 1:6. The extracted oil was determined by the difference in weight of the oil beakers, before and after the extraction. The total dietary fibre content was determined according to AACC method 32–05.01 using the Megazyme assay kit K-TDFR (Megazyme International, Bray, Ireland). The ash content was determined according to the AACC official method 08–01 [29].
Heat Treatment of Quinoa Protein Suspensions
The protein, starch and fibre from quinoa were subsequently used to reconstitute the quinoa protein-enriched fraction that was also obtained directly via dry fractionation. Therefore, the same amount of starch, fibre or starch/fibre was added to the protein isolate to investigate the effect of every component on the protein digestibility. Later, suspensions of 5% of protein (% w/v, in Milli-Q water) were prepared at room temperature in Eppendorf tubes of 2 ml. The suspensions were stirred with a Multi Reax shaker (Heidolph Instruments, Schwabach, Germany) for 30 min at 1800 rpm. Subsequently, the suspensions were subjected to heat treatment at 60 °C for 30 min and 1400 rpm of shaking in a preheated Eppendorf Thermomixer (Eppendorf AG, Hamburg, Germany). Heating at 120 °C for 30 min was carried out in a heating block (Grant GBT4, Cambridge, UK). After heating, the suspensions were immediately cooled and kept at room temperature until measurement the same day.
In Vitro Gastric Digestion of Quinoa Suspensions
The unheated and preheated suspensions of 5% protein (w/v, in Milli-Q water) were incubated in simulated gastric juice (SGJ) at 37 °C for 3 h. The simulated gastric juice was prepared according to Opazo-Navarrete et al. [20] with minor modifications. For this, pepsin from porcine gastric mucosa (1 g/L) and NaCl (8.775 g/L) were dissolved in Milli-Q water (18.2 MΩ cm at 25 °C, Millipore Corp., Molsheim, France) and the pH was adjusted to 2 using 2 M HCl. The enzyme:substrate ratio was kept constant at 1:2 (weight/weight) during all experiments. The vessels containing the SGJ were continuously stirred at 100 rpm and sealed with parafilm (Pechiney Plastic Packaging, Inc., IL, U.S.A) to avoid evaporation. Samples of 1 ml were taken at 20, 60, 120 and 180 min and immediately heated in a Thermomixer at 90 °C and 1400 rpm of shaking for 5 min in order to inactivate the pepsin. The pH of these samples was approximately 6. All digestion experiments were performed in triplicate.
High-Performance Size Exclusion Chromatography (HPSEC)
The SGJ, undigested and in vitro digested samples were analysed via high-performance size exclusion chromatography using an Ultimate 3000 UHPLC system (Thermo Scientific, MA) equipped with a TSKgel G3000SWXl column (7.8 mm X 300 mm) (Tosoh Bioscience LLC, PA) and TSKgel G2000SWXl column (7.8 mm X 300 mm) (Tosoh Bioscience LLC, PA) connected in line. For analysis, 1 mL of undiluted sample was centrifuged at 1000 rpm for 30 s to separate the large particles. 10 μL of the supernatant was injected into the system each time. The mobile phase was acetonitrile (30%) in Milli-Q water (70%) containing trifluoroacetic acid (0.1%). The flow rate was 1.5 mL/min and the UV detector was set at 214 nm. Calibration was done with: thyroglobulin (670 kDa), g-globulin (158 kDa), ovalbumin (44.3 kDa), α-lactalbumin (14.1 kDa), aprotinin (6.51 kDa), insulin (5.7 kDa), bacitracin (1.42 kDa) and phenylalanine (165 Da) (Sigma-Aldrich, Inc., St. Louis, MO, U.S.A.). The molecular mass was estimated against the elution time of molecular weights markers. All measurements were done in duplicate.
Degree of Hydrolysis (DH)
The OPA method was used to determine the degree of hydrolysis (DH) attained. The OPA reagent and standard L-serine solution were prepared according to Opazo-Navarrete et al. [20]. A standard curve using an L-serine solution was measured in the range of 50–200 mg/L. The OPA assay was carried out by the addition of 200 μL of sample (or standard) to 1.5 mL of OPA reagent. The samples with the OPA reagent added were measured after 3 min at 340 nm with a spectrophotometer DU 720 (Beckman Coulter Inc. Pasadena, CA, U.S.A). The absorbance values were converted to free amino groups (mmol/l) from a standard curve. Free amino group levels from the digestion samples were corrected by subtracting the contribution of free amino groups that were already present in the SGJ. The free amino groups were expressed as serine amino equivalents (Serine NH2). From this, the DH values were calculated according to Opazo-Navarrete et al. [20].
SDS-Page
The quinoa flour suspensions were analysed under non-reducing SDS-PAGE conditions. The samples were diluted with buffer (0.5 M Tris–HCl, pH 6.8; 2 wt% SDS; 2.5 wt% glycerol; 0.2 wt% bromophenol blue). The weight ratio of sample-to-buffer was 1:1. Each sample was heated to 95 °C for 4 min in an Eppendorf thermomixer (Eppendorf AG, Hamburg, Germany) and mixing at 800 rpm. An amount of 10 μL of the molecular weight marker Precision Plus Protein All Blue Standards (Bio-Rad Laboratories Inc., Hercules, USA) and each sample were loaded on a 12% Tris–HCl Mini-PROTEAN TGX Precast Gel (Bio-Rad Laboratories Inc., USA). The electrophoresis was carried out at 200 V. Afterwards, the gel was stained with Bio-safe Coomassie Stain (Bio-Rad Laboratories Inc., USA) and gel images were taken using a GS-900 Calibrated Densitometry System (Bio-Rad Laboratories, Inc., USA).
Statistical Analysis
Significance testing was performed using the IBM SPSS Statistic 21 for Windows computerized statistical analysis package. DH values were examined using one-way analysis of variance (ANOVA) to compare means between different samples. The differences were taken to be statistically significant when p < 0.05. When the F-values were found significant (p < 0.05), Tukey’s multiple comparisons procedure was used to determine any significant differences within the groups (Post Hoc-LSD). Results are expressed as a mean ± its standard deviation.
Results and Discussion
Protein-enriched flour was obtained via dry fractionation. The compositional analysis of every fraction used during the digestion experiments is presented in Table 1. Protein-enriched flour had high fibre and oil contents, while the starch content was lower than the whole quinoa flour. Starch was successfully isolated using the same dry fractionation method, while fibre could only be concentrated using a wet fractionation method.
Effect of Protein Concentration Method and Temperature on Protein Digestibility
Suspensions of quinoa protein isolate (QPI) and a protein-enriched fraction, unheated and preheated at 60 and 120 °C were used during in vitro gastric digestion analyses. The pH was registered throughout the digestion and a generally minor change in pH was observed over the time of digestion; however, this change was larger with the unheated samples with a pH change from 2 to 2.3. The protein digestibility is assessed by degrees of hydrolysis (DH), which is shown in Fig. 1. Protein-enriched flour obtained by dry fractionation, whether unheated or preheated, showed a higher in vitro gastric protein digestibility (p < 0.05) than quinoa protein that was isolated using a conventional wet fractionation method. QPI unheated and digested for 180 min gave a DH value of 10.5%, while a DH value of 8.5% is found for QPI preheated at 120 °C. On the other hand, protein-enriched flour unheated and digested for 180 min gave a DH value of 16%, while after pre-heating at 120 °C the DH value is 13.1%. This shows that the protein digestibility is affected by the isolation method of the proteins. Neucere & Ory [30] indicated that organic solvents may lead to a decrease in the protein solubility, which could affect the protein digestibility. This already was found by Opazo-Navarrete et al. [20] on a bitter quinoa variety, who found that quinoa protein concentrated via a dry fractionation method presents higher in vitro gastric digestibility than QPI obtained via a wet fractionation method. Therefore, the concentration method of proteins affects the protein gastric digestibility of quinoa.

In vitro gastric digestibility of unheated and preheated (a) protein-enriched fraction and (b) quinoa protein isolate
Regardless of the protein concentration method, solutions preheated at 120 °C present a significantly lower (p < 0.05) degree of hydrolysis (DH) than the solutions that were unheated or heated at 60 °C. The methods to purify proteins as wet fractionation are known to cause denaturation of proteins, which results in aggregation of the proteins. The quinoa preheated at 120 °C may have become chemically modified, resulting in crosslinks. Similarly, denaturation of the protein could have resulted in aggregation. Both would affect the accessibility of the protein for pepsin. The aggregation was corroborated by microscopy: after heating at 120 °C QPI shows big aggregates (Fig. 2). The dry fractionated, protein-enriched flour showed clearly smaller aggregates in comparison to the QPI. This could explain the lower protein digestibility obtained with QPI. This effect by aggregation was previously suggested by Opazo-Navarrete et al. [20] and Avila et al. [31] who found that quinoa protein preheated at 120 °C had lower gastric digestibility as a consequence of protein aggregation.

Light microscopy images of unheated and preheated at 60 and 120 °C quinoa protein isolate (QPI) and protein-enriched flour (PEF) dispersions
The preheated and unheated protein-enriched flour suspensions obtained from the dry fractionation method were analysed by HPSEC (Fig. 3). After digesting unheated and preheated quinoa suspensions for 180 min, higher levels of small peptides, ranging from 0.2 to 2 kDa, had been released. Quinoa protein suspensions preheated at 120 °C showed less released peptides than quinoa solutions that were unheated or preheated at 60 °C, but gave larger peptides (>2 kDa); even after 180 min of gastric digestion time. These results matched the values obtained for the degree of hydrolysis (DH).

HPSEC profiles of gastric digestion of quinoa suspensions prepared with a dry fractionated quinoa fraction and digested by pepsin at 37 °C for (a) 20 min, (b) 60 min, (c) 120 min and (d) 180 min
The chromatograms of the digested QPI suspensions (unheated and preheated) are presented in the first column of Fig. 4. The level of larger peptides (>2 kDa) from the unheated and preheated QPI suspensions is higher than that of the protein-enriched flour suspensions, while less individual amino acids are formed in the QPI suspensions, in comparison with the protein-enriched flour suspensions, both unheated and preheated. This higher amount of amino acids formed can be seen in the greater area generated between the elution times of 15 and 15.7 min. This range corresponds to a MW of 75–150 Da, 75 Da being the MW of lysine, the smallest amino acid.

HPSEC profiles of gastric digestion of unheated and preheated quinoa protein isolate (QPI) alone and mixed with starch and fibre suspensions digested by pepsin at 37 °C
Both the level of larger peptides, as well as that of the amino acids agree with the DH values that were obtained.
SDS-PAGE results under non-reducing conditions are shown in the Fig. 5. Proteins with a molecular weight (MW) ranging from 11 to 66 kDa were found in the unheated and preheated at 60 °C samples, while in the samples preheated at 120 °C, no bands were found. The two bands found between 49 and 66 kDa correspond to globulins called chenopodin subunits (11S), while around 10 kDa a band was found corresponding to the albumin subunit (2S). Defatting of quinoa flour seems to mainly affect the globulins (11S): these bands are less intense than those of dry fractionated and whole quinoa flour. The disappearance of bands of samples preheated at 120 °C indicate extensive aggregation of the quinoa proteins into aggregates larger than 250 kDa, which are not visible in the gel. This is clearly visible in the chromatograms of the dry fractionated quinoa flour (Fig. 3) and QPI (Fig. 4) digested suspensions: at retention times lower than 12.5 min (>50 kDa) the suspensions that were preheated at 120 °C showed a larger integrated peak area than the other samples, implying protein aggregation. Therefore, the aggregation might have reduced the accessibility of the proteins to the pepsin, leading to a decrease in the protein digestibility.

SDS-PAGE under non-reducing conditions patterns. M: marker; P-U: protein-enriched flour unheated; P-60: protein-enriched flour preheated at 60 °C; P-120: protein-enriched flour preheated at 120 °C; D-U: defatted flour unheated; D-60: defatted flour preheated at 60 °C; D-120: defatted flour preheated at 120 °C; F-U: flour unheated; F-60 flour preheated at 60 °C; F-120: flour preheated at 120 °C
Effect of Starch on Protein Digestibility
To assess the effect of starch on the protein digestibility (Fig. 6), the starch that was isolated via wet fractionation was added to the QPI to obtain the same starch concentration as in the dry fractionated protein-enriched fraction (Table 1). The oil was omitted from this study. Heating was applied to the suspensions to study the combined effect of starch and temperature on the protein digestibility.

The degree of hydrolysis (DH) of quinoa protein-starch suspensions during in vitro gastric digestion
The presence of starch reduced significantly (p < 0.05) the digestion rates of unheated and preheated QPI at 60 °C, but after 180 min of digestion time almost the same level of hydrolysis (8.5 and 8.6%) was obtained as without starch (Fig. 1b). Preheating at 120 °C gave a strong decrease in the digestion rate and even after 180 min (4.5%), the degree of hydrolysis was still only half of the value obtained without starch. These results show that starch strongly affects the protein digestibility, the effect being most pronounced at 120 °C. Wong et al. [32] found that the protein digestibility increased considerably when starch was removed from sorghum flour. López-Barón et al. [33] indicated that heat-induced protein denaturation or protease hydrolysis promote the enhancement of the protein-starch interactions. In their study, these protein-starch interactions reduced the enzymatic starch hydrolysis. The same protein-starch interaction could be responsible for the reduced digestibility of protein after heat-treatment at 120 °C in our study.
The chromatograms of quinoa protein-starch suspensions are shown in the second column of Fig. 4. The chromatograms of unheated and preheated at 60 °C suspensions had a similar integrated peak area after 180 min of gastric digestion, which implies similar peptides and amino acids formation (> 2 kDa). However, the digestion rate of the suspensions preheated at 60 °C again is lower in the first 120 min, in agreement to the DH values obtained earlier. The quinoa protein-starch suspensions preheated at 120 °C showed a much lower integrated peak area than the suspensions that were unheated or preheated at 60 °C, even after 180 min of gastric digestion. These results are therefore in agreement with the DH values obtained earlier.
Effect of Fibre on Protein Digestibility
The protein-enriched flour was reconstituted according to the protein, starch and fibre contents (Table 1). The oil was omitted from this study. Quinoa starch isolate and fibre concentrate were mixed with QPI to obtain the same concentration as in the dry fractionated protein-enriched flour. The in vitro gastric digestibility values of reconstituted quinoa protein-enriched flour shown in Fig. 7a. The profile of this figure is qualitative similar as was obtained with pure protein and protein with starch added (Figs. 1 and 6), however, the initial digestion rate (20 min of digestion) of the unheated suspension and the suspension preheated at 60 °C is slightly higher (5.6 and 4.7%) in comparison to that obtained with only starch.

The degree of hydrolysis (DH) of unheated and preheated (a) reconstituted dry fractionated quinoa and (b) dry fractionated suspensions quinoa during in vitro gastric digestion
It is interesting that the digestion rate and the DH after 180 min are clearly higher for the suspension preheated at 120 °C (7.3%), compared to what was obtained with only protein and starch (4.5%). This indicates that the effect of starch on the protein digestibility significantly decreases (p < 0.05) when the fibre is present as well. We suggest that the fibre may partly prevent the interactions between protein and starch. Besides, is important to consider that fibre prevents the starch to be fully hydrated, which will increase the gelatinisation temperature of the starch too much higher temperatures. Thus the inhibiting effect of starch on the protein digestibility is partly counteracted. However, the suspensions preheated at 120 °C still present lower DH values than the QPI digested for 180 min (Fig. 1b), which indicates that the effect of starch is counteracted only partially. Numerous studies have explored the effects of fibre on protein digestion by measuring the degree of nitrogen loss in human excretion [34]. Likewise, some studies in pigs have shown that fibre reduces the protein digestibility [35, 36]. According to the FAO, the reduction in the apparent digestibility of protein is typically less than 10%. While of course, many more effects are important over the whole digestive tract, the effect that we found may be one of the effects that could explain this observation.
Kritchevsky [37] indicated that fibre modifies and usually decreases the digestibility of proteins, along with lipids and certain minerals. The decrease in the digestibility might be caused by pectin and other gel-forming polysaccharides by retention of amino acids and peptides [38]. Other causes which may affect (decrease) the protein digestibility could be that the fibre inhibits access of enzymes to the protein. The presence of fibre in a system with a limited amount of water will also limit the hydration of starch, thereby increasing the temperature of starch gelatinization, which may result in a lower degree of gelatinisation and therefore a reduced effect of starch on the impediment of acid and enzyme ingression into the protein matrix.
A comparison of Fig. 7a to Fig. 7b reveals that the reconstitution of the dry fractionated protein concentrate from the individual fraction gives very different digestion dynamics. In the reconstituted concentrate, the digestion rate and the DH after 180 min are much lower, although the overall composition of both systems is the same. Apparently, the food matrix does have an effect on the digestibility; in this case, the original quinoa matrix that is present in the dry fractionated concentrate is much better digestible that would be expected based on its composition.
The chromatograms of quinoa protein-starch-fibre suspensions, which are reconstituted based on the concentration of protein-enriched flour, are shown in third column of Fig. 4. The protein-starch-fibre suspensions showed a greater amount of single amino acids formed in all treatments in comparison to the protein-starch suspensions (Fig. 4), which is in agreement with the higher DH values obtained for these suspensions. The chromatograms of unheated suspensions showed a faster initial increase of peptides of different sizes (0.2–2 kDa), but after 180 min of digestion, similar levels are seen as with the suspensions that were preheated at 60 °C. The suspensions preheated at 120 °C, however, yield lower levels in the MW range of 0.2–2 kDa, which indicates less peptide formation in comparison with the unheated and mildly heated (60 °C) suspensions. At the same time, the suspensions preheated at 120 °C give lower levels of single amino acids than the unheated and mildly preheated suspensions, which is in line with the DH values. While the initial hydrolysis rate was higher for the unheated suspensions, after 180 min of gastric digestion, both unheated and mildly preheated give similar DH values.
Surprisingly, the effect of the amount of fibre does not seem very important (Fig. 8). Two different concentrations of fibre were added to a dispersion of 0.1 g of protein and 0.01 g of starch: 10% or 0.01 g fibre, and 20% or 0.02 g fibre. Subsequently, the dispersions were preheated and then cooled to room temperature, and then subjected to in vitro gastric digestion.

The degree of hydrolysis (DH) of (a) unheated, (b) preheated at 60 °C and (c) preheated at 120 °C protein isolate, protein-starch, protein-starch-fibre (reconstituted dry fractionated quinoa) suspensions and with 10 and 20% of additional fibre added during in vitro gastric digestion
The presence of fibre reduces significantly (p < 0.05) the detrimental effect of starch on the protein digestibility when suspensions are preheated, mainly at high temperature. Apparently, this effect is not dependent on fibre concentration present in the dispersion. Therefore, a small amount of fibre could be enough to partly counteract the effect of starch on the in vitro protein digestibility.
In Fig. 9 we propose a mechanism of the effect of protein state and interaction with starch and fibre on the protein digestibility based on our results.
-
1.
Having native protein, starch and fibre in a (dry fractionated) concentrate imply that both fibre and starch do not absorb much water. The protein, which is well soluble in this state, is well accessible to digestion. Heating this system will denature the protein, but at the same time gelatinise the starch and hydrate the fibre, and therefore the protein aggregates will remain small, which keeps it still relatively accessible to digestion after dispersion in gastric juice.
-
2.
A protein that was isolated using a wet process (which implies pH changes and a thermal load during drying), is already partly denatured, less soluble and less accessible for digestion. Heating this protein will result in extensive aggregation and strongly lower accessibility for acid and pepsin, resulting in lower digestibility.
-
3.
Combining the wet isolated protein with starch will result in moderate digestibility: the protein is already partly aggregated, while the starch will remove some of the water and therefore will hinder the ingression of pepsin. Heating this suspension will exacerbate this, due to extensive starch gelatinization.
-
4.
Combining all three isolates (protein, starch and fibre) into a reconstituted concentrate will combine the moderate digestibility of the partially denatured protein, with the hindrance of the hydrating starch and fibre. Heating this system will result in a dense matrix that does not allow much ingression of pepsin, while the protein is also aggregated: slow overall digestion is the result.
Fig. 9 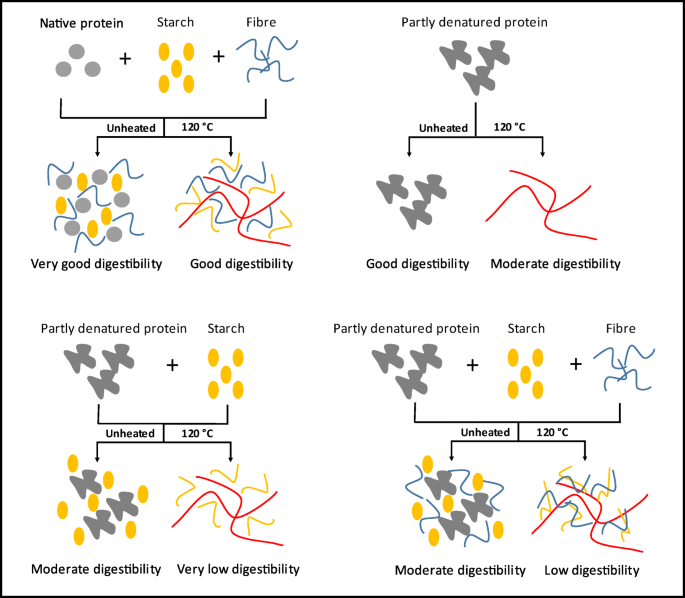
Schematic diagram of protein digestibility according to the protein state and interaction with other components after protein denaturation

This interpretation predicts, that all effects are kinetic: in the end, all protein will still be digested, but the present of partially or completely hydrated starch and fibre, plus the partially or fully aggregated state of the protein, will slow the hydrolysis. It should be noted, that even after 180 min of digestion time, we still found significant differences. Longer time scales are not relevant to gastric digestion. Thus, for all practical purposes, our interpretation implies that dry fractionated foods and foods with more or less starch and fibre will enter the duodenum in very different states.
The results present in this article could be relevant for future food formulations. The effect of fibre on the digestion of a protein-starch matrix is an important finding, because proteins, starch and fibre are ingredients commonly used for the manufacturing of foods. This study could be of help to design food for various target groups, both for a fast protein digestibility as well as a slow protein digestibility.
Conclusions
While the extraction method used to isolate the protein reduces the protein digestibility, the presence of starch and fibre reduce even more the digestibility of quinoa protein. This could be explained by hindrance by starch and fibre to the ingression of pepsin, reducing the rate of hydrolysis of the proteins. Starch reduces the quinoa protein digestibility more strongly than fibre. The most important finding is that the effect of starch is partially counteracted by the presence of fibre, which would be really interesting for future quinoa based food formulations. This phenomenon is not dependent on the concentration of the fibre. Heating at 120 °C does affect the protein digestibility, which we expect is due to the formation of larger protein aggregates which are also less accessible by pepsin.
References
L.E. Abugoch, Adv. Food Nutr. Res. 58, 1–31 (2009)
G. Li, F. Zhu, Food Chem. 221, 1560–1568 (2017)
R. Navarro-Lisboa, C. Herrera, R.N. Zúñiga, J. Enrione, F. Guzmán, S. Matiacevich, C. Astudillo-Castro, Food Bioprod. Process. 102, 20–30 (2017)
J. Ruales, B.M. Nair, Plants Foods Hum. Nutr. 42(1), 1–11 (1992)
A. Bhargava, S. Shukla, D. Ohri, Genet. Pl. Br. 63, 359–360 (2003)
S.A. Elsohaimy, T.M. Refaay, M.A.M. Zaytoum, Annals Agric. Sci. 60(2), 297–305 (2015)
E. Kaspchak, M.A. Schüler, S. Fogagnoli, C.R. Cavicchiolo, J.L. Meira, M.R. Mafra, L. Igarashi-Mafra, Food Chem. 232, 263–271 (2017)
S. Fischer, R. Wilckens, J. Jara, M. Aranda, W. Valdivia, L. Bustamante, F. Graf, I. Obal, Ind. Crop. Prod. 107, 558–564 (2017)
C. Brinegar, S. Goundan, J. Agric. Food Chem. 41(2), 182–185 (1993)
M. Koziol, J. Food Compos. Anal. 5(1), 35–68 (1992)
N. Lindeboom, P.R. Chang, K.C. Falk, R.T. Tyler, Cereal Chem. 82(2), 216–222 (2005)
M.E. Steffolani, A.E. León, G.T. Pérez, Starch–Stärke 65(11-12), 976–983 (2013)
K. Lorenz, Starch–Stärke 42(3), 81–86 (1990)
J. Ruales, B.M. Nair, Plant Foods Hum. Nutr. 45(3), 223–246 (1994)
E. Bertoft, Cereal Chem. 90(4), 294–311 (2013)
S. Navruz-Varli, N. Sanlier, J. Cereal Sci. 69, 371–376 (2016)
D. Dhingra, M. Michael, H. Rajput, R.T. Patil, J. Food Sci. Technol. 49(3), 255–266 (2012)
J.M. Lattimer, M.D. Haub, Nutrients 2(12), 1266–1289 (2010)
A.M. Gómez-Caravaca, G. Iafelice, V. Verardo, E. Marconi, M.F. Caboni, Food Chem. 157, 174–178 (2014)
M. Opazo-Navarrete, M.A.I. Schutyser, R.M. Boom, A.E.M. Janssen, Int. J. Food Sci. Nutr. 31, 1–11 (2017)
S. Chauhan, N.A.M. Eskin, R. Tkachuk, Cereal Chem. 69(1), 85–88 (1992)
I. Dini, G.C. Tenore, A. Dini, LWT – Food Sci. Technol. 43(3), 447–451 (2010)
A. Bacigalupo, M.E. Tapia, in Cultivos Andinos Subexplotados y Su Aporte a la Alimentacion, ed. by C. Andinos. Agroindustria (Oficina Regional de La FAO para América Latina y el Caribe, Santiago, 2000)
M.A.I. Schutyser, A.J. van der Goot, Trends Food Sci. Technol. 22, 154–164 (2011)
F.S. Pushparaj, A. Urooj, Food Nutr. Sci. 2(08), 895–900 (2011)
K.G. Duodu, J.R.N. Taylora, P.S. Beltonb, B.R. Hamaker, J. Cereal Sci. 38(2), 117–131 (2003)
M. Opazo-Navarrete, D. Tagle, R.M. Boom, A.E.M. Janssen, M.A.I. Schutyser, J. Food Compos. Anal. 74, 95–101 (2018)
D.D. Dalgetty, B.-K. Baik, Cereal Chem. 80(3), 310–315 (2003)
AACC International, Method 08-01: Ash-Basic Method, 10th edn. (AACC International, St. Paul, 1983)
N.J. Neucere, R.L. Ory, J. Agric, Food Chem. 16(2), 364–365 (1968)
G. Avila Ruiz, M. Opazo-Navarrete, M. Meurs, M. Minor, G. Sala, M. van Boekel, A.E.M. Janssen, Food Biophys. 11(2), 184–197 (2016)
J.H. Wong, T. Lau, N. Cai, J. Singh, J.F. Pedersen, W.H. Vensel, W.J. Hurkman, J.D. Wilson, P.G. Lemaux, B.B. Buchanan, J. Cereal Sci. 49(1), 73–82 (2009)
N. López-Barón, Y. Gu, T. Vasanthan, R. Hoover, Food Hydrocoll. 69, 19–27 (2017)
FAO, Energy and Protein Requirements (Switzerland, Geneva, 1985)
G. Le Goff, J. van Milgen, J. Noblet, Anim. Sci. J. 74(03), 503–515 (2002)
L. Buraczewska, J. Anim. Feed Sci. 10(1), 139–152 (2001)
D. Kritchevsky, Annu. Rev. Nutr. 8(1), 301–328 (1988)
R. Mosenthin, W.C. Sauer, F. Ahrens, J. Nutr. 124(8), 1222–1229 (1994)
Acknowledgements
Mauricio Opazo Navarrete contributed to this research thanks to a PhD scholarship from CONICYT (Formation of Advanced Human Capital Program), Chile.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no conflict of interest.
Informed Consent
Informed consent was obtained from all individual participants included in the study. Additional informed consent was obtained from all individual participants for whom identifying information is included in this article.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Opazo-Navarrete, M., Tagle Freire, D., Boom, R.M. et al. The Influence of Starch and Fibre on In Vitro Protein Digestibility of Dry Fractionated Quinoa Seed (Riobamba Variety). Food Biophysics 14, 49–59 (2019). https://doi.org/10.1007/s11483-018-9556-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11483-018-9556-1