
Abstract
Purpose
Although past studies have found well-constrained soil carbon (C)/nutrient ratios, the effects of increased nitrogen (N) and water inputs on these ratios across soil depths have rarely been assessed in semi-arid grasslands.
Methods
In this study, we evaluated the contents of total C, N, phosphorus (P), sulfur (S), and their stoichiometric ratios in a 0–80 cm soil profile following 13 years of successive N (at rates of 5 and 15 g m−2 yearr−1) and water addition (180 mm per growing season) in a semi-arid grassland of the Mongolian Plateau.
Results
In the 0–10 cm soil layer, long-term N addition tended to increase total C and N contents but decreased soil total P and S contents compared to the control. The effects of N addition, as observed in 0–10 cm soil, however, were not consistent with that in the deep 10–80 cm soil layers. Water addition increased the total C, N, and P contents across the entire soil profile but increased total S content only in 0–40 cm soil. Moreover, the combined addition of N and water generally had stronger effects on the four elements across the whole soil profile. For the stoichiometry of the four elements, a low rate of N addition (5 g m−2 year−1) increased soil C:N ratios and decreased soil P:S ratios in the 0–80 cm soils, but a high rate of N addition (15 g m−2 year−1) produced the opposite effect. Both N addition rates resulted in an increase in the soil C:P, C:S, N:P, and N:S ratios. Similarly, in plots that received water, water addition alone decreased the soil C:N ratios, while N addition caused higher fluctuations in these six elemental ratios. However, there was no consistent pattern of change in any one ratio, independent of the addition of water, when taking into account N addition rates and soil depths.
Conclusion
Our findings showed that the effects of N addition on soil total C, N, P, and S contents and their stoichiometric ratios were highly influenced by the rate of N addition and the depth of soil, and that these effects could be modulated by increasing precipitation. These results need to be carefully considered while managing the ecological environment in semi-arid steppes.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
The biogeochemical cycling of mineral elements, including nitrogen (N), phosphorus (P), sulfur (S), and their equilibrium and interactions in terrestrial ecosystems, are crucial for the preservation of ecosystem services (Zechmeister-Boltenstern et al. 2015; Sardans et al. 2021). Because soil is typically a “black box” with high spatial heterogeneity and structural complexity, it is difficult to analyze and predict the availability of these nutrients in soils (Bünemann et al. 2018). The majority of earlier investigations discovered well-constrained relationships among soil C, N, and P, suggesting that alterations in the stoichiometry of these nutrients may better reflect variations in their availability in soils under environmental changes (Cleveland and Liptzin 2007; Elser et al. 2010; Yue et al. 2017). Apart from C, N, and P in soils, the element S is also a fundamental component of proteins and critical to the growth of organisms (Blum et al. 2013; Li et al. 2019). It is crucial to recognize changes in S availability and its stoichiometric relationships with C, N, and P in order to comprehend how these elements interact in the dynamics of ecosystem services and to evaluate the degree of nutrient restriction in ecosystems (Chen et al. 2016). However, compared to the elements C, N, and P, soil S cycling is poorly understood due to the paucity of research that has measured soil S content and investigated the interactions of soil C, N, P, and S at the same time (Kirkby et al. 2011; Khan et al. 2016; Tipping et al. 2016). In addition, more than 55% of the C and nutrients reside in deep soils below 20 cm, but few studies have directly compared the responses of these elements and their stoichiometry to environmental changes between topsoil versus subsoil, particularly in field experiments (Doetterl et al. 2018; Spohn 2020; Xu et al. 2021).
Around 20% of the Earth’s land area is covered by grasslands, which serve crucial ecosystem services but are currently severely threatened by anthropogenic activities such as increasing N deposition (Bengtsson et al. 2019). The mean annual N deposition in temperate grasslands worldwide has already exceeded 1 g N m−2 and is expected to rise 5–15-fold by 2050 if no mitigation measures are taken (Heyburn et al. 2017; Lü and Tian 2007; Wen et al. 2020).
Experimental findings show that N addition altered the soil stoichiometry in grassland ecosystems by boosting soil C and N contents while decreasing soil P and S levels (Li et al. 2019; Ning et al. 2021; Xu et al. 2021). However, related findings are not entirely consistent among different experiments because the effects of N addition would vary with several factors like the rate of N addition, the depth of soil, and the availability of water (Chen et al. 2016; Gao et al. 2018; Coonan et al. 2019). It is worth noting that soil C:N:P:S ratios are largely stable, and their equilibrium can be recovered even when soils have been severely disturbed (Cleveland and Liptzin 2007; Kirkby et al. 2011; Tian et al. 2010; Yang et al. 2019). For instance, in N-limited grasslands, short-term N addition was deemed to reduce the C:N ratio by alleviating microbial N starvation; however, when N addition treatment is sustained, C:N ratio may not change any further after declining to a supposedly saturated point (Manzoni et al. 2010; Schleuss et al. 2019; Zhu et al. 2021).
Most temperate grasslands are found in semi-arid regions, where both abiotic and biotic processes are highly constrained by the availability of water (Cregger et al. 2014; Nielsen and Ball 2015; Bengtsson et al. 2019). In the future, severe precipitation events are anticipated to occur more frequently in Northern China, resulting in increased annual precipitation (Wu et al. 2011; Ren et al. 2017; Pendergrass and Knutti 2018). Previous studies have indicated that N addition and altered precipitation could have either an independent or a combined effect on the cycling of N and other nutrients in semi-arid and arid ecosystems (Wu et al. 2011; Ochoa-Hueso et al. 2014; Nielsen and Ball 2015). Additionally, higher precipitation not only stimulated the above- and belowground biomass, but also enhanced the flux of C and nutrients (such as N, P, and S) by influencing soil microbial activities (Nielsen and Ball 2015; Greaver et al. 2016; Engelhardt et al. 2019). Furthermore, increased precipitation, particularly during heavy precipitation events, may alter the distribution of nutrients along the soil profile by accelerating leaching and weathering processes (Wu et al. 2011; Doetterl et al. 2018). Overall, it is unclear how increased precipitation regulates the impacts of N addition on soil C, N, P, and S contents along the soil profile in semi-arid ecosystems.
Currently, knowledge of how N addition affects the stoichiometric relationships among these elements in the soil is still lacking, for at least three reasons. (1) The dynamics of the topsoil C:N:P:S ratio following N addition may not be applicable to the entire profile (i.e., deep soils). In contrast to topsoil, the dynamics of soil nutrients in deep soil layers are closely tied to the rates of soil nutrient diffusion and infiltration (Tian et al. 2010). Furthermore, P and S supplies are mainly derived from the parent materials, and their contents would decline with soil depth more slowly than C and N (Gao et al. 2018; Spohn 2020). (2) The treatment duration of most previous studies was relatively short (≤ 6 years), and their results might not accurately reflect the long-term responses of soil nutrients. For example, in a global meta-analysis, Xu et al. (2021) observed that long-term N addition had a stronger impact on soil C content than short-term N addition. (3) The majority of past investigations did not differentiate between the effects of low-N addition and high-N addition rates. Empirical evidence demonstrated that the positive effects of N deposition would disappear or even turn negative when the rate of N addition was high, or the duration of the treatment was prolonged (Bai et al. 2010; Stevens et al. 2015).
Northern China is a relatively populous region with a range of farming-pastoral ecotones. The temperate grassland of this region is an essential component of the Eurasian steppe and is susceptible to changes in N and water availability (Ren et al. 2008, 2017). We conducted a controlled field experiment in this temperate steppe to investigate how long-term N and water addition affect soil contents of the four key elements C, N, P, and S, as well as their stoichiometric ratios. Our objectives were to (i) distinguish the effects of low- and high-N additions on the total amounts of C, N, P, and S in soil and their stoichiometric ratios; (ii) understand how water addition regulates the effects of N addition on these elemental ratios; and (iii) illustrate the stratified properties of elemental ratios in different soil depths under N and water addition. We also attempted to explore the relationship between these ratios and soil physicochemical properties in various soil layers.
2 Materials and methods
2.1 Experimental sites and treatments
This study was conducted at the Inner Mongolia Duolun Ecosystem Research Station (IMDERS; 116° 170″ E, 42° 02″ N; elevation 1324 m). The long-term mean annual temperature is 2.2 ℃, with monthly mean temperatures ranging from − 17.5 °C (January) to 18.9 °C (July), and the mean annual precipitation (MAP) is 382 mm, with 85% of the precipitation from May to September (IMDERS data, 1953–2016). The soil type is chestnut (Chinese classification) or Haplic Calcisols (FAO classification), with 5.2% clay and 44.2% silt in the top 0–10 cm layer (Niu et al. 2021a). Vegetation in the study area is dominated by Stipa krylovii, Agropyron cristatum, and Artemisia frigida, and three species together account for 65% of the community’s total aboveground biomass (Ren et al. 2017).
In 2005, seven (107 m × 8 m) blocks of naturally assembled vegetation communities were set up in a flat field using a split-plot experimental design. Identical treatments were placed into all blocks, and each block was split into two main plots that randomly received water addition and ambient precipitation, respectively. Each main plot was further split into six (8 m × 8 m) subplots, which randomly received either nutrient addition or ambient nutrient deposition. In this study, we only selected a few levels of N addition treatments in both plots with and without the addition of water, including control and two levels of N addition (5 and 15 g N m−2 year−1) treatments. These rates, while still higher than the combined rate of atmospheric N deposition and N fertilization (2 ~ 5 g N m−2 year−1) in our study area, are useful for evaluating the accumulative impacts of N addition on soil nutrients. 1-m walkways separated all subplots. Compared to plots that received ambient precipitation, the water addition plots received an additional 180 mm of precipitation (approximately 50% of the long-term MAP in the study area), with 15 mm of water sprinkled weekly during the primary growing seasons (June to August). Even though the 50% increase in precipitation is greater than the projected precipitation increases in the near future, it is comparable to the variation in annual precipitation between wet and dry years over the past 30 years, given that increases in precipitation during the growing season are more frequent in our study region (IPCC 2021; Xu et al. 2015; Ren et al. 2017). In our experiment, N was applied in the form of urea, with each application of N accounting for 50% of the annual amount (May and July). The study site had only been used for grazing until it was fenced in 2000, and no fertilizer had been administered to any of the plots prior to our study.
2.2 Field sampling and laboratory analysis
Soil samples were collected from the surface layer to 80 cm in depth, with intervals of 10 cm or 20 cm (0–10, 10–20, 20–30, 30–40, 40–60, and 60–80 cm) in September 2017 (i.e., after 13 consecutive years of treatment). Three randomly chosen cores were gathered and combined from each plot to create a composite sample. In this study, there were five replicates for each treatment, resulting in 180 composite samples. Each fresh soil sample was sieved, air-dried, and further ground to determine the contents of total C, N, P, and S in the soil, as well as other physicochemical parameters.
The water content of the soil was quantified using the gravimetric method, and the pH was measured using a pH meter in soil supernatant at a soil-to-water ratio (w:v) of 1:2.5. Soil available nitrogen (AVN) was estimated using alkali hydrolysis and Conway’s diffusion technique. The soil’s total C, N, and S contents were determined using the combustion method in an N/C/S analyzer (CHNOS Elemental Analyzer, Germany). The total P content of the soil was measured using an inductive coupled plasma emission spectrometer (ICP-ES) after acid extraction. In a nutshell, 6 mL of HNO3, 2 mL of HClO3, and 2 mL of HF were added to 1 g of the ground soil sample and extracted in a microwave oven at 180 °C for 4 h. The extracted solutions were diluted to 25 mL volume, filtered, and quantified by ICP-ES. To ensure the accuracy of the obtained data acquired, we used standard certified soils for C, N, P, and S during the measurement process, which were purchased from the Institute of Geographical and Geochemical Prospecting of China.
2.3 Data analysis
All data analyses were performed in R 4.1.3 (https://cran.r-project.org/src/base/R-4/). Before data analysis, we verified that the data set meets the requirements of normality and homogeneity using the “Q-Q plot” and “bartlett. test” functions in the “Car” package, respectively. Two-way analysis of variance (ANOVA) was used to assess the effects of N and water addition on the contents of the four elements and their stoichiometric ratios. Next, one-way ANOVA with Tukey HSD test was used to examine the effects of N addition rates on these parameters in plots with or without water addition. One-way ANOVA was also used to assess the effects of soil depth on these soil variables. Additionally, a t-test was used to evaluate the effects of adding water at the same rate of N addition on these soil variables. To better illustrate changes in C:N:P:S stoichiometry in each soil layer, we also calculated the response ratio (RR) of various elemental stoichiometric compositions (i.e., C:N, C:P, C:S, N:P, N:S, and P:S ratios) using the following formula:
where RT is the mean value of soil nutrient ratio (e.g., the C:N ratio) from treatment plots and RC is the control mean value from control plots. If the nutrient ratio increased under long-term treatments compared to the control, then RR > 0 and vice versa. The closer the RR value approximates to zero, the weaker the treatment effects are. Additionally, the shifts in soil total C, N, P, and S contents and their ratios among different treatments are demonstrated using principal component analysis (PCA) and permutational multivariate analysis of variance (PERMANOVA) in “FactoMineR,” “Factoextra,” and “Vegan” packages. Pearson correlation analysis further highlighted the relationships between soil C, N, P, and S parameters and principal component axes. Pearson correlation was also used to indicate the relationships of soil stoichiometric ratios with soil C, N, P, and S contents and soil properties.
3 Results
3.1 Changes in soil total C, N, P, and S contents
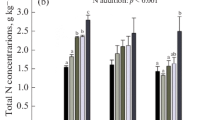
The total C, N, P, and S contents all decreased significantly with soil depth regardless of the addition of N and water (Figs. 1 and 2). The overall effects of N addition on total C and N contents were positive, whereas the impacts on total P and S contents were negative; these effects depend on water addition and soil depth (Figs. 1 and 2, Table S1). Specifically, when compared to the corresponding layers in control plots, low-N addition significantly increased the total C content in 0–10, 20–30, and 40–60 cm soils, while high-N addition increased the contents of total C, N, and P in 10–20 cm soil (Figs. 1 and 2). The values of total S content in the 0–10, 30–40, 40–60, and 60–80 cm soil layers were lower under both N addition rates, and the negative effects were pronounced under high-N addition (Fig. 2b).

Content of total carbon (TC) and total nitrogen (TN) across the 0–80 cm soil profile following 13 years of N and water additions in a semi-arid steppe. Notes: colored bars and error bars indicate the mean values and standard errors (n = 5). Vertical comparisons (i.e., between the six layers of soil) of mean changes in soil elements are denoted by different capital letters (p ≤ 0.05). For each soil layer, significant differences among different treatments are indicated by different lowercase letters (p ≤ 0.05). Furthermore, the effects of water addition at the same N additions were detected using a t-test; the symbols + and – indicate increased and decreased effects, respectively

Content of total phosphorus (TP) and total sulfur (TS) across the 0–80 cm soil profile following 13 years of N and water addition in a semi-arid steppe. For notes, see Fig. 1
In comparison to the control plots, water addition alone increased total C content in the 0–10 cm soil and increased total S content in the 10–20 cm soil and decreased total S content in the 30–40, 40–60, and 60–80 cm soil layers (p ≤ 0.05, Figs. 1 and 2). Nitrogen plus water addition resulted in overall higher contents of soil total C, N, and P compared with plots with water addition alone, while low-N plus water addition significantly increased total S content in the 10–20, 20–30, 30–40, and 60–80 cm soil layers (Figs. 1 and 2). When comparing C, N, P, and S contents at the same N addition rates, N plus water additions generally increased soil total C and P contents in most soil layers. It also increased total S content in top 0–30 cm soils but decreased that in 30–80 cm soils except for 40–60 cm soil, where high-N addition increased soil total S content (Figs. 1 and 2).
3.2 Shifts in soil total C: N: P: S stoichiometry
Regardless of N and water addition, C:N and C:P rations decreased, and N:P, N:S, and P:S increased with increasing soil depth (p ≤ 0.05, Fig. S1). Compared to control plots, low-N addition considerably raised C:N ratios across the entire soil profile except for the 10–20 cm soil (p ≤ 0.05), whereas high-N addition significantly lowered this ratio except for the 60–80 cm soil (Fig. 3a). In most soil layers, both N addition rates generally increased ratios of C:P, C:S, N:P, and N:S (p ≤ 0.1; Figs. 3 and 4). Low-N addition increased total P:S in 30–40 and 40–60 cm soil layers, whereas high-N addition marginally increased this ratio in 0–10,10–20, 40–60, and 60–80 cm soils (p ≤ 0.1, Fig. 4c).

Changes in the TC: TN, TC: TP, and TC: TS ratios across the 0–80 cm soil profile following 13 years of N and water addition in a semi-arid steppe. Notes: points and error bars indicate the mean values and standard errors (n = 5). The effects were quantified as changes in the ratios of total C (TC): total N (TN), TC: total P (TP), and TC: total S (TS) under N and water addition compared to the control plots. The values below zero indicate decreased effects and those above zero indicate increased effects. Unless otherwise stated, if the error bars have not crossed the zero line, there are significant differences between the N addition treatments and control. Furthermore, the effects of water addition at the same N additions were detected using a t-test; the symbols + and – indicate increased and decreased effects, respectively

Changes in the ratios of TN:TP, TN:TS, and TP: TS across the 0–80 cm soil profile following 13 years N and water addition in a semi-arid steppe. For notes, see notes in Fig. 3
Water addition alone typically increased C:P and N:P ratios in 0–60 cm soil layers but decreased the ratios in 60–80 cm soil (p ≤ 0.1, Figs. 3e and 4d). Further, water addition alone enhanced the C:N, C:S, and N:S ratios in soil with depths of 0–10 and 40–60 cm, as well as the C:S and N:S ratios in soils with a depth of 30–40 cm (p ≤ 0.05, Figs. 3 and 4). In plots receiving water addition, both N addition rates generally increased the C:N ratio across the entire soil profile, while low-N plus water addition typically decreased the N:P ratio except for 10–20 cm soil (Figs. 3 and 4). Likewise, when comparing the values of these ratios at the same N addition rates, we only observed consistent positive effects of water addition on the C:N ratio throughout the entire soil profile (Fig. 4d).
3.3 Covariations and correlation of soil C:N:P:S stoichiometry with their contents
Ten variables, including soil C, N, P, and S contents and six related ratios, were subjected to PCA, which revealed two main mixed axes that, together, accounted for 77.0%, 72.6%, 74.1%, 73.2%, 73.7%, and 66.2% of total variances in the soil layers 0–10, 10–20, 20–30, 30–40, 40–60, and 60–80 cm, respectively (Fig. 5). The N and water additions impacted covariations of soil C:N:P:S stoichiometry in distinct ways by altering different characteristics, and these impacts varied with soil depth (Fig. 5). For instance, in the 0–10 cm soil layer, the PC1 axis explained 52.3% of the total variance and was associated with the majority of soil parameters, with the exception of C:N and P:S ratios (Fig. 5a, Table S3); however, in the 60–80 cm soil layer, the PC1 axis explained 41.1% of the variations and was represented by contents of total C, N, and P, as well as six ratios, signaling that water addition alone had the greatest impact on these ten variables (Fig. 5f, Table S3). When all soil samples were considered, changes in soil C:N:P:S stoichiometry were, in fact, more often connected to their contents than soil properties (Table S3). In particular, the ratios of C:P, C:S, N:P, N:S, and P:S were negatively correlated to soil pH and positively correlated with C and N contents in the 0–10 cm soil, whereas C:N ratio was significantly positively correlated to total C content across all soil depths (p ≤ 0.05, Table S4). All soil nutrient ratios were typically positively correlated with total C and N contents and negatively correlated with total P and S contents, albeit these associations were only significant in specific soil layers (Table S4).

Effects of N and water addition on variations in soil total C (TC), total N (TN), total P (TP), and total S (TS) contents and their six stoichiometric ratios in a semi-arid steppe. The ten parameters were used to assess the effects among different treatments, and each treatment was summarized as a single point by PCA (n = 50). The results of the PERMANOVA test are attached to the corner of the figure
4 Discussion
4.1 Effect of N addition on topsoil C:N:P:S stoichiometry depends on N addition rates and water availability
The status of the nutrient limitation in grasslands at the regional scale can be evaluated effectively using elemental ratios, and the majority of research shows that variations in the N:P ratio can be regarded as an indicator of both N saturation and limitation (Schade et al. 2005; Tipping et al. 2016; Sardans et al. 2021). In the present study, we used different elemental ratios related to soil C, N, P, and S in an effort to deepen our understanding of the ecological stoichiometry theory. Despite the fact that both rates of N addition tended to increase soil total C and N contents and decrease soil total P and S contents, we found no consistent responses between the low and high rates of N addition. Moreover, the mean response ratios of C:S (0.18), N:S (0.21), and P:S (0.19) were higher than those of C:N (0.02), C:P (0.07), and N:P (0.08), suggesting that changes in these nutrients were asynchronous and that C, N, P, and S had distinct sensitivity to N addition (Khan et al. 2016). Furthermore, water addition alone did not appear to have any apparent effects on the contents of soil total C, N, P, and S in most soil layers. Still, there were notable changes in elemental ratios in water addition plots, implying that increased precipitation altered interactions among these elements in soils. Additionally, we observed that the sensitivities of total amounts of the four elements to water addition gradually decreased from C, N, and P to S in the 0–10 cm soil, which may be related to the elements’ different affinities for water (Kramer and Chadwick 2018).
The effects of N addition on elemental ratios in water addition plots were also incongruent with those in plots with ambient precipitation. In contrast to ambient precipitation plots, low N addition significantly decreased soil N:P ratios in the water addition plots, indicating that water addition regulated, or perhaps altered the effects of N addition on soil elemental ratios.
The effects of N and water addition on these four soil elements and their stoichiometry varied with N addition rates in the semi-arid steppe, which may be explained by a number of distinct mechanisms. First, in semi-arid grassland ecosystems, water is the key limiting resource for the growth of plants and microorganisms, while N is a close second. Adding water alone has a major impact on the soil microbial communities and biogeochemical processes, particularly during dry years (Cregger et al. 2014; Nielsen and Ball 2015). Plant and microbial populations can interact to uptake additional N only under wet soil conditions; otherwise, most N could be lost by leaching into the underground or as gas to air (Nielsen and ball 2015; Niu et al. 2021b). As a result, we observed significant interaction effects of N and water addition in most soil layers (Table S2). Second, soil N availability for plants and microbial communities would be altered with increasing intensity and duration of N addition (Elser et al. 2010; Niu et al. 2022a). In the long run, plant growth processes and plant residue inputs into the soil varied significantly under low- and high-N addition rates (Bai et al. 2010), which may further affect soil C and nutrient cycling. High-N addition typically inhibits plant growth and the subsequent return of plant residues to soil due to low soil pH and the toxic effects of ammonium on plants rather than having a favorable impact on these processes (Tian and Niu 2015; Li et al. 2019; Niu et al. 2022b). Third, alterations in plant and microbial traits, especially their competition for soil nutrients, may result in an unsynchronized response of these soil elements (Moreau et al. 2015; Craig and Fraterrigo 2017). The nutritional needs of different plants and microbial species vary, and long-term N addition may further accentuate these nutritional differences concurrently while also causing changes in the structure of the plant and microbe community (Zhang et al. 2014; Ma et al. 2016; Niu et al. 2021b).
4.2 Effect of N and water addition on soil C: N: P: S stoichiometry varied with soil depth
An increasing number of research demonstrated that soil total C, N, P, and S contents declined as soil depth increased, but fewer studies have examined changes in their ratios along the soil profile (Manzoni et al. 2010; Khan et al. 2016; Chen et al. 2016; Niu et al. 2022a). Based on the decreasing trends of soil C:N and C:P ratios with soil depth and the increasing trends of soil N:P, N:S, and P:S ratios regardless of N and water addition, it may be inferred that C and N are the key factors limiting the microbial activity in deep soil. In our investigated grassland, soil total C and N contents in the 0–10 cm soil were elevated after 13 years of N addition, but total P and S contents were reduced (Figs. 1 and 2). The findings in the 0–10 cm soil layer were in accordance with those of earlier research on grassland ecosystems, probably because short-term N input promotes plant growth, which in turn increases C and N input to the soil as plant residuals (Stevens et al. 2015; Yue et al. 2017; Tang et al. 2018; Niu et al. 2022b). It may also be caused by the gradual depletion of other nutrients (such as P, S, and K) as a result of further N addition, which raises the readily available amount of N. This could alleviate the N limitation experienced by plants and microorganisms (Fujita et al. 2010; Hill and Jones 2019; Dong et al. 2021). Furthermore, we confirmed that the responses of soil nutrient contents and stoichiometric ratios in 0–10 cm soil were distinct from those in 10–80 cm subsoil (Figs. 1, 2, 3, and 4, S1). These findings are consistent with earlier research, such as that by Han et al. (2019), who found that N addition increased available N only in 0–10 cm soil and had no significant effects on the total P content of top- and sub-soil in three different grassland types.
The singular or combined effect of water and N addition varied with soil depth. For instance, the soil C:S ratio increased with low-N addition in the 0–10, 30–40, and 40–60 cm soil layers and decreased in other soil layers (Fig. 3e). The varied responses at different soil depths may be closely related to a leaching process (Cameron et al. 2013; Kramer and Chadwick 2018). Deep soils (10–80 cm), as opposed to topsoil (0–10 cm), primarily receive leaching materials from the topsoil, which may have a differential impact on soil C and nutrient cycling in various soil layers (Cameron et al. 2013; Mnich and Houlton 2016). However, the amount and composition of these leaching materials present in each layer of soil remain unknown, thus warranting further studies. In our study, no significant correlations between soil properties and these six elemental ratios were found across the soil profile when all soil samples were taken into account, with the exception of soil pH in the 0–10 cm soil layer. Importantly, we found significant positive correlations between these six elemental ratios and the total C and N contents, but negative correlations with the total P and S contents, which may be strongly related to the processes of formation and leaching (Mnich and Houlton 2016; Doetterl et al. 2018).
4.3 Implications for ecosystem management
To the best of our knowledge, this is the first comprehensive analysis of the responses of six soil elemental ratios to long-term N and water addition across the entire 0–80 cm soil profile of a semi-arid grassland ecosystem. Our findings highlight the effects of N addition rates and enhanced water availability on soil C and nutrient balance in semi-arid ecosystems, which are crucial for maintaining productivity and conserving grassland biodiversity. Our results further confirmed that soil responses of elemental ratios rather than elemental contents were sensitive to long-term N addition, partly contrary to prior studies that suggested that plants rather than soils were sensitive to N deposition (Bowman et al. 2008; Bai et al. 2010). Nevertheless, the interpretation of our findings is subject to several restrictions. First, there were only two N addition rates (5 versus 15 g m−2 year−1), which may not adequately represent the distinction between low- and high-N addition rates in affecting soil C and nutrient dynamics. Second, although potential explanations for variations in the contents of these soil nutrients and their ratios were proposed, the underlying mechanisms are still poorly understood. For instance, our current understanding of the interplay between weathering and N addition and their effects on soil nutrients is limited. Therefore, further research is required to understand the broad trends and mechanisms underlying the dynamic changes in soil nutrient ratios caused by the addition of N and alternative precipitation conditions.
5 Conclusions
After 13 years of N and water addition, we found that soil total C, N, P, and S contents and their elemental ratios significantly changed, although there were no consistent patterns when specific N addition rate and soil depth were taken into account. N addition generally had positive effects on total C and N contents only in 0–10 cm of soil, but had negative effects on total P and S, while water addition raised the contents of these four elements. The elemental ratios responded differently to low- and high-N addition rates across the soil profile, and water addition further regulated N addition effects on these elemental ratios. Our findings demonstrated that N addition rates and soil depths in semi-arid steppe influence the impact of N and water addition on soil C, N, P, and S contents and their stoichiometric ratios. Our research offers valuable insights for assessing the cycling of essential soil elements, particularly in semi-arid grasslands where nutrient inputs and changes in precipitation are co-occurring.
Data Availability
Supplementary data to this article can be found online at https://figshare.com/s/01b1d638a77fa9d84137 (https://doi.org/10.6084/m9.figshare.19663995).
References
Bai Y, Wu J, Clark CM, Naeem S, Pan Q, Huang J, Zhang L, Han X (2010) Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from inner Mongolia grasslands. Glob Chang Biol 16:358–372
Bengtsson J, Bullock JM, Egoh B, Everson C, Everson T, O’Connor T, O’Farrell PJ, Smith HG, Lindborg R (2019) Grasslands—more important for ecosystem services than you might think. Ecosphere 10:e02582
Blum SC, Lehmann J, Solomon D, Caires EF, Alleoni LR (2013) Sulfur forms in organic substrates affecting S mineralization in soil. Geoderma 200:156–164
Bowman WD, Cleveland CC, Halada L, Hresko J, Baron J (2008) Negative impact of nitrogen deposition on soil buffering capacity. Nat Geosci 1:767–770
Bünemann EK, Bünemann G, Bai Z, Creamer RE, Deyn GD, Goede RD, Fleskens L, Geissen V, Kuyper TW, Mäder P, Pulleman M, Sukkel W, Groenigen JW, Brussaard L (2018) Soil quality – a critical review. Soil Biol and Biochem 120:105–125
Cameron KC, Di HJ, Moir JL (2013) Nitrogen losses from the soil/plant system: a review. Ann Appl Biol 162:145–173
Chen H, Yang L, Wen L, Luo P, Liu L, Yang Y, Wang K, Li D (2016) Effects of nitrogen deposition on soil sulfur cycling. Global Biogeochem Cy 30:1568–1577
Cleveland CC, Liptzin DC (2007) N: P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 85:235–252
Coonan EC, Richardson AE, Kirkby CA, Kirkegaard JA, Amidy MR, Simpson RJ, Strong CL (2019) Soil carbon sequestration to depth in response to long-term phosphorus fertilization of grazed pasture. Geoderma 338:226–235
Craig ME, Fraterrigo JM (2017) Plant–microbial competition for nitrogen increases microbial activities and carbon loss in invaded soils. Oecologia 184:583–596
Cregger MA, McDowell NG, Pangle RE, Pockman WT, Classen AT (2014) The impact of precipitation change on nitrogen cycling in a semi-arid ecosystem. Funct Ecol 28:1534–1544
Doetterl S, Berhe A, Arnold C, Bodé S, Fiener P, Finke P, Fuchslueger L, Griepentrog M, Harden J, Nadeu E, Schnecker J, Six J, Trumbore S, Oost K, Vogel C, Boeckx P (2018) Links among warming, carbon and microbial dynamics mediated by soil mineral weathering. Nat Geosci 11:589–593
Dong C, Wang W, Liu H, Xu X, Chen X, Zeng H (2021) Comparison of soil microbial responses to nitrogen addition between ex-arable grassland and natural grassland. J Soils Sediments 21:1371–1384
Elser JJ, Fagan WF, Kerkhoff AJ, Swenson NG, Enquist BJ (2010) Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change. New Phytol 186:593–608
Engelhardt IC, Blazewicz SJ, Barnard RL (2019) Effects of contrasting precipitation patterns on the trajectory of actively growing and inactive microbial communities after rewetting. Soil Biol and Biochem 134:172–174
Fujita Y, Robroek BJ, de Ruiter PC, Heil GW, Wassen MJ (2010) Increased N affects P uptake of eight grassland species: the role of root surface phosphatase activity. Oikos 119:1665–1673
Gao Y, Ma MZ, Yang T, Chen WL, Yang TT (2018) Global atmospheric sulfur deposition and associated impaction on nitrogen cycling in ecosystems. J Clean Prod 195:1–9
Greaver TL, Clark CM, Compton JE, Vallano D, Talhelm AF, Weaver CP, Band LE, Baron JS, Davidson EA, Tague CL, Felker-Quinn E, Lynch JA, Herrick JD, Liu L, Goodale CL, Novak KJ, Haeuber RA (2016) Key ecological responses to nitrogen are altered by climate change. Nat Clim Change 6:836–843
Han Y, Dong S, Zhao Z, Sha W, Li S, Shen H, Xiao J, Zhang J, Wu X, Jiang X, Zhao J, Liu S, Dong Q, Zhou H, Jane C (2019) Response of soil nutrients and stoichiometry to elevated nitrogen deposition in alpine grassland on the Qinghai-Tibetan Plateau. Geoderma 43:263–268
Heyburn JP, McKenzie M, Crawley J, Fornara DA (2017) Effects of grassland management on plant C: N: P stoichiometry: implications for soil element cycling and storage. Ecosphere 8:e01963
Hill PW, Jones DL (2019) Plant–microbe competition: does injection of isotopes of C and N into the rhizosphere effectively characterise plant use of soil N? New Phytol 221:796–806
IPCC (2021) Climate change 2021: the physical science basis: contribution of working group I to the sixth assessment report of the intergovernmental panel on climate change. Cambridge University Press
Khan KS, Mck R, Castillo X, Kaiser M, Joergensen R (2016) Microbial biomass, fungal and bacterial residues, and their relationships to the soil organic matter C/N/P/S ratios. Geoderma 271:115–123
Kirkby CA, Kirkegaard JA, Richardson AE, Wade LJ, Blanchard C, Batten G (2011) Stable soil organic matter: a comparison of C: N: P: S ratios in Australian and other world soils. Geoderma 163:197–208
Kramer M, Chadwick O (2018) Climate-driven thresholds in reactive mineral retention of soil carbon at the global scale. Nat Clim Chang 8:1104–1108
Li T, Liu H, Wang R, Lü X, Yang J, Zhang Y, He P, Wang Z, Han X, Jiang Y (2019) Frequency and intensity of nitrogen addition alter soil inorganic sulfur fractions, but the effects vary with mowing management in a temperate steppe. Biogeosciences 16:2891–2904
Lü C, Tian H (2007) Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data. J Geophys Res-Atmos 112:D22
Ma H, Bai G, Sun Y, Kostenko O, Zhu X, Lin S, Ruan W, Zhao N, Bezemer T (2016) Opposing effects of nitrogen and water addition on soil bacterial and fungal communities in the Inner Mongolia steppe: A field experiment. Appl Soil Ecol 108:128–135
Manzoni S, Trofymow JA, Jackson RB, Porporato A (2010) Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter. Ecol Monogr 80:89–106
Mnich ME, Houlton BZ (2016) Evidence for a uniformly small isotope effect of nitrogen leaching loss: results from disturbed ecosystems in seasonally dry climates. Oecologia 181:323–333
Moreau D, Pivato B, Bru D, Bussset H, Deau F, Faivre C, Matejicek A, Strbik F, Philippot L, Mougel C (2015) Plant traits related to nitrogen uptake influence plant-microbe competition. Ecology 96:2300–2310
Nielsen UN, Ball BA (2015) Impacts of altered precipitation regimes on soil communities and biogeochemistry in arid and semi-arid ecosystem. Global Change Biolo 21:1407–1421
Ning Z, Zhao X, Li Y, Wang L, Lian J, Yang H, Li Y (2021) Plant community C: N: P stoichiometry is mediated by soil nutrients and plant functional groups during grassland desertification. Ecol Eng 162:106179
Niu G, Wang R, Hasi M, Wang Y, Geng Q, Wang C, Jiang Y, Huang J (2021a) Availability of soil base cations and micronutrients along soil profile after 13-year nitrogen and water addition in a semi-arid grassland. Biogeochemistry 152:223–236
Niu G, Hasi M, Wang R, Wang Y, Gen Q, Hu S, Xu X, Yang J, Wang C, Han X, Huang J (2021b) Soil microbial community responses to long-term nitrogen addition at different soil depths in a typical steppe. Appl Soil Ecol 167:104054
Niu G, Wang Y, Wang R, Ning Q, Guan H, Yang J, Lu X, Wang C, Han X, Huang J (2022a) Intensity and duration of nitrogen addition jointly alter soil nutrient availability in a temperate grassland. J Geophys Res Biogeosci 127:e2021JG006698
Niu G, Liu L, Wang Y, Guan H, Ning Q, Liu T, Ning Q, Liu T, Rousk K, Zhong B, Yang J, Lu X, Han X, Huang J (2022b) Effects of decadal nitrogen addition on carbon and nitrogen stocks in different organic matter fractions of typical steppe soils. Ecol Indic 144:109471
Ochoa-Hueso R, Bell MD, Manrique E (2014) Impacts of increased nitrogen deposition and altered precipitation regimes on soil fertility and functioning in semiarid Mediterranean shrublands. J Arid Environ 104:106–115
Pendergrass AG, Knutti R (2018) The uneven nature of daily precipitation and its change. Geophys Res Lett 45:11980–11988
Ren J, Hu Z, Zhao J, Zhang D, Hou F, Lin H, Mu X (2008) A grassland classification system and its application in China. Rangel J 30:199–209
Ren H, Xu Z, Isbell F, Huang J, Han X, Wan S, Chen S, Wang R, Zeng D, Jiang Y, Fang Y (2017) Exacerbated nitrogen limitation ends transient stimulation of grassland productivity by increased precipitation. Ecol Monogr 87:457–469
Sardans J, Janssens IA, Ciais P, Obersteiner M, Penuelas J (2021) Recent advances and future research in ecological stoichiometry. Perspectives Plant Ecol 50:125611
Schade J, Espeleta J, Klausmeier C, McGroddy M, Thomas S, Zhang L (2005) A conceptual framework for ecosystem stoichiometry: balancing resource supply and demand. Oikos 109:40–51
Schleuss P, Widdig M, Heintz-Buschart A, Guhr A, Martin S, Martina K, Spohn M (2019) Stoichiometric controls of soil carbon and nitrogen cycling after long-term nitrogen and phosphorus addition in a mesic grassland in South Africa. Soil Biol and Biochem 135:294–303
Spohn M (2020) Phosphorus and carbon in soil particle size fractions: a synthesis. Biogeochemistry 147:225–242
Stevens CJ, Lind RM, Hautier Y, Harpole WS, Borer ET, Hobbie S, Seabloom EW, Ladwig L, Bakker JD, Chu C, Collins S, Davies KF, Firn J, Hillebrand H, Pierre KJ, Macdougall A, Melbourne B, Mcculley RL, Morgan J, Orrock JL, Prober SM, Risch AC, Schuetz M, Wragg P (2015) Anthropogenic nitrogen deposition predicts local grassland primary production worldwide. Ecology 96:1459–1465
Tang Z, Xu W, Zhou G, Bai Y, Li J, Tang X, Chen D, Liu Q, Ma W, Xiong G, He H, He N, Guo Y, Guo Q, Zhu J, Han W, Hu H, Fang J, Xie Z (2018) Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems. Proc Natl Acad Sci USA 115:4033–4038
Tian D, Niu S (2015) A global analysis of soil acidification caused by nitrogen addition. Environ Res Lett 10:024019
Tian H, Chen G, Zhang C, Melillo J, Hall CA (2010) Pattern and variation of C: N: P ratios in China’s soils: a synthesis of observational data. Biogeochemistry 98:139–151
Tipping E, Somerville CJ, Luster J (2016) The C: N: P: S stoichiometry of soil organic matter. Biogeochemistry 130:117–131
Wen Z, Xu W, Li Q, Han MJ, Tang AH, Zhang Y, Luo XS, Shen JL, Wang W, Li KH, Pan YP, Zhang L, Li W, Collett J, Zhong B, Wang X, Gouldin K (2020) Changes of nitrogen deposition in China from 1980 to 2018. Environ Int 144:106022
Wu Z, Dijkstra P, Koch G, Penuelas J, Hungate B (2011) Responses of terrestrial ecosystems to temperature and precipitation change: a meta-analysis of experimental manipulation. Global Change Biol 17:927–942
Xu Z, Ren H, Li M-H, van Ruijven J, Han X, Wan S, Li H, Yu Q, Jiang Y, Jiang L (2015) Environmental changes drive the temporal stability of semi-arid natural grasslands through altering species asynchrony. J Ecol 103:1308–1316
Xu C, Xu X, Ju C, Chen-Han Y, Wilsey B, Luo Y, Fan W (2021) Long-term, amplified responses of soil organic carbon to nitrogen addition worldwide. Glob Change Biol 27:1170–1180
Yang D, Song L, Jin GZ (2019) The soil C: N: P stoichiometry is more sensitive than the leaf C:N: P stoichiometry to nitrogen addition: a four-year nitrogen addition experiment in a Pinus koraiensis plantation. Plant Soil 442:183–198
Yue K, Fornara DA, Yang W, Peng Y, Li Z, Wu FZ, Peng C (2017) Effects of three global change drivers on terrestrial C: N: P stoichiometry: a global synthesis. Glob Chang Biol 23:2450–2463
Zechmeister-Boltenstern S, Keiblinger KM, Mooshammer M, Penuelas J, Richter A, Sardans J, Wanek W (2015) The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecol Monogr 85:133–155
Zhang Y, Lü XT, Isbell F, Stevens C, Han X, He N, Zhang GM, Yu Q, Huang JH, Han XG (2014) Rapid plant species loss at high rates and at low frequency of N addition in temperate steppe. Glob Change Biol 20:3520–3529
Zhu Z, Zhou J, Shahbaz M, Tang H, Liu S, Zhang W, Yuan H, Zhou P, Alharbi H, Wu J, Kuzyakov Y, Ge T (2021) Microorganisms maintain C: N stoichiometric balance by regulating the priming effect in long-term fertilized soils. Appl Soil Ecol 167:104033
Acknowledgements
We are very grateful to the anonymous reviewers for their insightful and constructive comments, which have helped us improve our manuscript. We greatly thank Lettice Hicks and Johannes Rousk at Lund University, and Dr. Danillo O. Alvarenga at the University of Copenhagen for language polishing and constructive suggestions. We thank Ms. Xiaoping Pan for her excellent technical assistance in laboratory work. This study was supported by the China Postdoctoral Science Foundation (2022M713196) and the National Natural Science Foundation of China (31870440).
Funding
Open access funding provided by Lund University.
Author information
Authors and Affiliations
Contributions
G.X.N., J.H.H., and Y.J. designed this filed experiment, while G.X.N., Y.Q.J., and Y.L.W. executed the field and laboratory work. G.X.N. analyzed data and wrote the manuscript, and R.Z.W., B.Q.Z., J.H.H., and X.T. L. also contributed to the data analysis and revision of the manuscript.
Corresponding author
Ethics declarations
Competing interest
The authors declare no competing interests.
Additional information
Responsible editor: Chengrong Chen
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Niu, G., Zhong, B., Wang, R. et al. Effects of nitrogen and water addition on soil carbon, nitrogen, phosphorus, sulfur, and their stoichiometry along soil profile in a semi-arid steppe. J Soils Sediments 23, 3298–3309 (2023). https://doi.org/10.1007/s11368-023-03558-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-023-03558-z