
Abstract
Purpose
The studies aimed to reconstruct major phases of litho-morpho-pedogenic processes and past environmental changes in the Baltic coastal zone near Poddąbie (54°37,070′ N, 16°57,897′ E) based on physical, chemical, pollen, and fossil Cladocera analysis of the polycyclic pedocomplex.
Materials and methods
The polycyclic pedocomplex under study included seven fossil soils developed from the Early Holocene until modern times from various materials. The soils were sampled and analyzed using standard procedures in soil science to determine their physical and chemical properties. Pollen and fossil Cladocera analysis was also carried out in the examined profile. Radiocarbon dates of selected samples were also specified.
Results and discussion
The studied polycyclic pedocomplex has been developed from materials of varied origin, including glaciofluvial, fluvial, aeolian, and biogenic deposits of the lake. According to the results of pollen and fossil Cladocera analysis and radiocarbon dating, this body of water has evolved from the beginning of Preboreal period to the half of Boreal. The studied soils represent various reference groups and advancement of development. They constitute relics of former geomorphological and soil-forming processes, which have taken place over the last 9500 years, including periglacial processes, wildfires, Baltic Sea shoreline changes, and aeolian activity. The last stage of the coastal relief dynamics was conditioned by the fire which took place about 1472–1637 AD. The lack of plant cover and intensive abrasion due to direct vicinity of the coastline has caused the deposition of aeolian material, which is the parent material of nutrient-poor initial soils.
Conclusions
The analysis of fossil soil development with the use of specialized research methods allowed for reconstructing the dynamics of past geomorphological processes and natural environment changes in the study area. The studied polycyclic pedocomplex constitutes a record of environmental changes taking place in the central part of the Polish Baltic coastal zone from the Early Holocene to the present day. The studied soil sequence is located within a land depression developed in place of the former water reservoir created as a result of permafrost degradation, and filled with Holocene deposits.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Landscape evolution is a continuous process, which depends on various natural and anthropogenic factors and processes, e.g., erosion related to runoff, climate change, soil, and vegetation cover and impact of human activity (Tsara et al. 2001; Bogaart et al. 2003; de Alba et al. 2004; Yetemen et al. 2010). Contemporary landscape of the Polish Baltic coastal zone is modified mainly by abrasion and aeolian processes. Fossil soils are preserved within the cliff walls and are exposed during ongoing abrasion processes. This instability of cliffs is a threat to fossil soils due to the risk of soil profile destruction. Under natural conditions, relief dynamics, especially on undulated landscapes, are common and they depend on slope reduction, grain size of superficial material, plant cover, rainfall, etc. (Holliday 1992). Coastal zones are an example of young-glacial landscape with specific features conditioned by the influence of the sea and morphogenetic processes dependent on the cliff structure (e.g., geological structure, lithology, and plant cover). The relationships between plant cover, erosional processes, and pedogenesis dynamics are directly determined by climatic conditions. Favorable climatic conditions influenced the formation of a dense plant coverage, which promoted the development of soils. However, in periods of climate cooling, the surface instability was intensified due to depletion of the plant cover and intensification of aeolian processes, which significantly impeded or inhibited soil formation (e.g., Engstrom and Wright 1984; Hošek et al. 2017). These relationships have already been studied in the soils of the Baltic coastal zone near Orzechowo (Kruczkowska et al. 2017).
The investigated cliff section is an integral part of the coastal zone with contemporary borders direct to the sea; however, primarily, it has developed under different environmental conditions. The witness of changes of the Baltic Sea range are, e.g., the trunks of pine trees (radiocarbon age estimated on 10,900–10,700 uncal. year BP) located 23 m b.s.l., about 15 km north of the central part of the contemporary Polish Baltic coastal zone and more than 3000 years old oak and beech forest, which were revealed on a beach near Słowiński National Park (Uścinowicz 2003, 2006). At the early beginning of the Preboreal period, the rising of sea level took place as an effect of the ice sheet melting and increased the area of Scandinavia. The southern Baltic coast has moved southward; however, they could still be located, from several to dozen kilometers, from the north of today’s coast. The Baltic Sea transformations have relatively stabilized during the last 6000 years, through slow changes of sea level and the course of the shoreline. The erosion processes on the cliffs and the coastal zone peneplanation, initiated already in the Atlantic period, began to gradually dominate (Uścinowicz 2003, 2006). Despite the intensive processes and evolution of the coast, there were periods of relative stabilization that allowed the formation of soils.
Dynamics of coastal zone, characteristic features of soil-forming environment of cliffs, and the geochemical feedback between the cliff landscape and the impact of the sea determines the development of soils with unique features (Bednarek 1979). Soils located nearby the coastlines are extremely exposed to the degradation, and as a result of the impact of external factors, the soil system can be disturbed, often irreversibly. Soils as creations developed as a result of soil-forming processes with the participation of natural environment components are repositories of knowledge about the past and contemporary processes, in view of their persistent pedogenic solid-phase characteristics (Targulian and Sokolova 1996; Targulian and Goryachkin 2004; Targulian and Krasilnikov 2007). Depending on the strength, violence, and the range of these processes, former soil characteristics can be preserved to varying degrees. Soil morphology changes in combination with the interference of physical and chemical properties of soil that allows making environmental reconstructions. Fossil soils are common in the coastal zone near the study area (e.g., Krzyszkowski et al. 1999; Florek et al. 2008; Olszak et al. 2008, 2011; Florek et al. 2010; Wróblewski et al. 2013; Kruczkowska et al. 2017), and they are represented by both mineral and organic units. History of Young Glacial landscape development is also related to the existence of transitory water reservoirs of different genesis. The traces of existence of a palaeolake were recorded in the studied polycyclic pedocomplex. Layers associated with the direct impact of the lake required application of additional research methods, such as Cladocera and pollen analysis, which are supplementary to the development reconstruction of the analyzed polycyclic pedocomplex and are the basis for determining the biotic conditions prevailing during the development of these soils.
The study aimed to reconstruct the dynamics of Early Holocene litho-morpho-pedogenic processes in the southern Baltic Coastal zone near Poddąbie based on various properties of fossil soils and Cladocera, and palynological data.
2 Materials and methods
2.1 Site characteristics
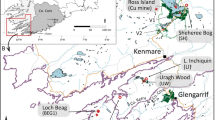
The study was performed near Poddąbie (54°37,070′ N, 16°57,897′ E) within the coastal cliff located in the central part of the Słowińskie Coast, North Poland (Fig. 1). The study was based on the polycyclic pedocomplex covering contemporary soil and a sequence of six fossil soils of varied age beneath it. The studied soil sequence is located within a fossil land depression filled with Holocene deposits over Late Pleistocene deposits. Young-glacial landscape of the study area has been developed 18–16 ka BP as a result of deglaciation of the Gardno Phase of the Vistula Glaciation (Kozarski 1995). The height of the cliff varies along the Słowińskie Coast. In the examined section, it was about 13 m. Gray glacial till occurs in the bottom of the studied cliff section. It is covered by a fluvioglacial sand. Over the sand was noted a thin layer of lacustrine sediments and then a sequence of aeolian deposits.

The study area. a Location. b Soil profiles
2.2 Soil sampling and analysis
The soils were described according to the WRB classification system (IUSS Working Group WRB 2015) and sampled. One disturbed sample and two samples of undisturbed structure (using 100-cm3 steel rings) were taken from each horizon. Disturbed samples were processed for further analysis using standard procedures, including drying in 40 °C and sieving through a 2.0-mm sieve to remove skeleton fraction. Particle-size distribution was determined by the mixed pipette and sieve method, and textural fractions and groups were divided according to the Polish Soil Science Society 2008 (PSSS 2009). Bulk density and total porosity have been analyzed by the drying-weight method, and pH was measured potentiometrically in a suspension with water and 1 mol−1 KCl in a soil/water/KCl ratio of 1:2.5 (pH-meter Elmetron CPC-401). Total organic carbon (TOC) was determined by Tyurin method (Dziadowiec and Gonet 1999), and total nitrogen (TN) with the Kjeldahl method (van Reeuvijk 2002). Total content of elements was analyzed in a solution after sample digestion in a mixture of 40% HF and 60% HClO4 (ETHOS PLUS microwave digestion system). The content of phosphorus was determined using molybdenum blue method (van Reeuvijk 2002), whereas contents of K, Ca, Mg, Fe, and Mn by flame atomic absorption spectrometry (FAAS, Perkin Elmer AAS 2100). “Free” iron oxides (Fed) were determined by FAAS after sample extraction by Mehra and Jackson (1960), and amorphous iron oxides (Feo) were also determined using the same technique, after extraction of samples according to the procedure of Schwertmann (van Reeuvijk 2002). Based on the results of the above analyses, we also calculated geometric graphical measures after Folk and Ward (1957), including mean diameter (MG), sorting (σG), skewness (SkG), and kurtosis (KG), using GRADISTAT 5.11 software (Blott and Pye 2001), weathering ratio (Fed/Fet) of soil mineral phase, and Feo/Fed ratio.
2.3 Radiocarbon dating
Four samples of organic materials (peat, charcoal, humus) were collected for the purposes of radiocarbon dating. Analysis was performed in the Laboratory of Absolute Dating, Cianowice, Poland. OxCal v 4.2 software (Bronk Ramsey, 2013) and IntCal13 calibration curve (Reimer et al. 2013) were used for calibration of the obtained dates. Detailed information concerning dating is included in the Table 1.
2.4 Pollen analysis
Nine organic soil samples of 1 cm3 were collected from Dystric Fibric Histosol in 5-cm intervals. The samples were boiled in 10% KOH, treated with 10% HCl or soaked for several days in HF (depending on the sediment composition) and then acetolyzed (Faegri and Iversen 1989; Dybova-Jachowicz and Sadowska 2003). Sums of total pollen grains were counted up to a minimum of 200 grains of tree and shrub pollen (AP). The non-pollen palynomorphs (NPPs, remains of algae, Cyanobacteria, fungi, and some faunal remains) enable the reconstruction of the history of local aquatic and wetland ecosystems by reflecting palaeohydrological changes.
Identification of pollen and spores of aquatic and wetland species was performed by following Beug (2004). The identification of microremains of fungi and some faunal organisms (NPPs) was based on van Geel (1978, van Geel et al. 1981) and Barthelmes et al. (2012). Zonation was conducted visually based on changes in dominant pollen, spores, and NPP taxa. The diagram was constructed using the POLPAL software (Nalepka and Walanus 2003).
2.5 Cladocera analysis
The Cladocera analysis was performed in samples taken from Dystric Fibric Histosol at 5-cm intervals, from the depth of 372.5–412.5 cm. Samples with a volume of 1 cm3 were prepared using the standard procedure (Frey 1986; Korhola and Rautio 2001). After removal of carbonates using 10% HCl, each sample was boiled in 10% KOH for 30 min, washed with distilled water, and sieved through a 40-μm mesh sieve. The fine material was transferred into a polycarbonate test tube. Prior to counting, the remains were colored with safranine T. The analysis was performed with a Nikon model ECLIPSE Ci-L microscope with magnifications of ×10, ×40, and ×60. A minimum of 200 remains of Cladocera (three to eight slides) were examined in each sample. First, all remains from each slide were counted (headshields, shells, postabdomens, postabdominal claws, and antennules) and then converted to one Cladocera specimen, and all ephippia were grouped together. Identification and ecological interpretation of the Cladocera remains were based on Goulden (1964), Szeroczyńska (1985, 1998), Hofmann (1986, 2000), Korhola (1990), Duigan (1992), Flössner (2000), and Szeroczyńska and Sarmaja-Korjonen (2007). The results of Cladocera analyses are shown in the Fig. 3, and they include relative abundance diagram of subfossil cladoceran taxa, the total number of Cladocera individuals, and the number of species. The numerical analysis was performed with the POLPAL software (Nalepka and Walanus 2003).
3 Results and discussion
3.1 Chronology of the litho-morpho-pedogenic processes, morphology, and textural parameters of the soils
The studied polycyclic pedocomplex developed from the Early Holocene until modern times from various materials, including glaciofluvial, fluvial, aeolian, and biogenic deposits. The soils represent different reference groups and advancement of development. Detailed information concerning parent material origin and taxonomy of the soils under study is included in Table 2.
Mineral soils were developed from fine and medium aeolian sands, usually moderately and poorly sorted with positively skewed, strongly positively skewed, or near symmetrical particle size distribution curves (Table 3). Dystric Arenosol (Protospodic) at the depth of 210–233 cm is a border between apparently aeolian origin of surface sediments and the fluvial character of Dystric Fluvisol (Relictiturbic). Fine and medium aeolian sands to the depth of 210 cm represent a genetically similar group of sediments with mainly platykurtic distribution, while deeper soil horizons are very leptokurtic or mesocurtic. Soils to a depth of 210 cm are also better sorted.
Between 370 and 415 cm, a fossil Dystric Fibric Histosol occurs over aeolian sandy material. This soil has developed in different climate conditions as compared to the overlying soils. It took place during the Preboreal (radiocarbon age of bottom 8772–8551 cal. year BC) and Boreal periods (radiocarbon age of topsoil 7748–7590 cal. year BC). Low admixture of mineral material may suggest formation of basal peat in a wide and a shallow hollow with dead ice in the bottom. According to Błaszkiewicz (2005) and van Loon et al. (2012), in the studied area, permafrost was present the time mentioned. It is also confirmed by the studies of Kruczkowska et al. (2017) on permafrost-affected soil in a cliff near Orzechowo (Słowińskie Coast). Local morphology of depressions and the possibility of permanent drainage were particularly important factors in a longer conservation of dead ice (Błaszkiewicz 2008). Permafrost degradation has caused land surface lowering and reservoir backfilling. Charcoals were found in the roof of Dystric Fibric Histosol, which indicates the fire that may additionally have led to the erosion intensification, filling up of the reservoir and the start of a new phase of the coast development. Directly on the surface of Dystric Fibric Histosol, Dystric Fluvisol (Relictiturbic) profile was located, which may be the consequence of environmental changes that may have led to the permafrost degradation and slope erosion. The signs of cryoturbations are visible in the whole profile of this soil, especially in its bottom part. Fluctuations of melt-water level are marked in rusty discolorations of 6C@ horizon. This profile can be treated as a boundary between periglacial conditions and environmental stabilization represented by the occurrence of Dystric Arenosol (Protospodic). These soils located at the depths of 233–124 cm constitute sequence providing for the ongoing temperate or boreal climate conditions interspersed with episodes of aeolian activity of varying intensity during their development. Radiocarbon age of charcoals found in topsoil of Dystric Arenosol (Protospodic) at the depth of 124 cm was estimated at 1472–1637 cal. year AD, while 5ABs horizon (210–220 cm) at 543–158 cal. year BC. The presence of charcoals should be interpreted as evidence of human activity. Two weakly developed soils (Dystric Arenosols) have developed within the roof part of this profile. During the periods of terrain stability, due to the higher vegetation density, the enhancement of sandy material in humic substances occurred, which is visible in soil profiles as thin humus horizons. Parent materials of these soils constitute sandy aeolian deposits. To the depth of 370 cm, soil profiles were developed from the aeolian sandy material. However, at the depth of 233–370 cm, this material also has features of fluvial remodeling. In soils located at greater depths was observed increasing content of silt and loam, especially at the depth of 210–233 cm within Dystric Arenosol (Protospodic) profile and 6A@, 6Cg@ horizons, and with medium content of silt: 9.3 and 10.5%, respectively.
3.2 Reconstruction of environmental changes based on pollen and fossil Cladocera analysis
Fossil Cladocera and pollen analysis were performed in samples of lacustrine origin (the profile of Dystric Fibric Histosol). Based on the pollen analysis of selected tree and herbaceous plant species, the taxa of aquatic, mire plants and non-pollen palynomorphs, aquatic and mire vegetation, and other macrofossils (NPPs; Table 4; Fig. 2), we distinguished five zones (LPAZ) illustrating the evolution of the studied aquatic-mire ecosystem and the history of forest in the neighborhood of the study site (Table 4).

Pollen diagram of samples collected from Dystric Fibric Histosol
Subfossil Cladocera fauna of sediments from the studied cliff section are represented by nine species from two families: Chydoridae and Daphniidae (Table 5). Most of the remains belong to the families of Chydoridae (8). The predominant Cladocera remains were identified as species, which are component of the group that inhabited a littoral zone, depending on the presence of aquatic plants (Flössner 1972) and as living in the bottom. The most numerous in this group are Alona quadrangularis, Alona affinis, Alona rectangula, and Chydorus sphaericus. The spread of horsetail (Equisetum) in the littoral zone about 8660 cal year BC (at the depth of 412.5 cm) confirms that the accumulation area was at least temporarily flooded. At the same time, a relatively high water level, probably associated with humidified climate, was registered in other parts of Poland (Ralska-Jasiewiczowa et al. 1998). Primarily, it was the lack of Cladocera in the reservoir. The first remains of Cladocera were found at the depth of 407.5 cm. After a short period of high water level, it has gradually began to decline. The presence of the open areas with features of steppe-tundra illustrates mainly grass pollen (Poaceae). Different frequency and concentration enables to distinguish five Cladocera assemblage zones in the studied profile (Table 5, Fig. 3).

Relative abundance diagram of subfossil Cladocera taxa and the diagram of total number of Cladocera individuals, and the number of species of the Poddąbie palaeolake (Cliff 3) CAZ-Cladocera Assemblage Zones (det. M. Niska)
Pollen diagram records two distinct decreases of water level in the reservoir with a clear increase of trophism. First, at the depth of 410 cm, it was dominated by birch and pine forests, which represent the Preboreal period. The decrease of the water level has initiated the development of the lowland bog with sward features and dominance of marsh ferns, which documents the high share of spores of the Thelypteris palustris species and sedge pollen (Cyperaceae family). During this period, the reservoir was inhabited by Cladocera species such as Alona quadrangularis—a species dwelling in the sediments that are rich in decaying plants, indicating the occurrence of higher water level; however, its gradual decline suggests a progressive lowering of the water level in the reservoir (Błędzki and Rybak 2016), and the dwelling of Camptocercus rectirostris species among aquatic plants could indicate a slight warming of the water in the reservoir.
The appearance of remains of the fungus HdV 200 at the depth of about 395 cm (van Geel et al. 1989; Kuhry 1997) and the high turnout of spores of other saprophytic fungi, including Sordaria, Coniochaeta ligniaria, Gaeumannomyces sp., and Pleospora, indicates the presence of relatively dry microhabitats on helophyte stems and plant remains in a shallow, temporarily drying water reservoir. The recorded lowering of the water level was caused by climate warming. It was the main factor of the hazel (Corylus) expansion dated to about 10,300 cal. year BP on the Sławno Plain (West Pomerania) by Gadziszewska (2015), and it was dated to 10,500 cal. year BP in large areas of Europe (Finsinger et al. 2006; Giesecke et al., 2011). In the rush communities of the water reservoir was found Typha latifolia, a species preferring eutrophic water and requiring the average temperature in July at a minimum of 13 °C (Isarin and Bohncke 1999; Ralska-Jasiewiczowa et al. 2004). The decrease of water level was marked by a decrease in the attendance of Alona quadrangularis species and a general low turnout of species and individuals of Cladocera in the reservoir. A slight increase in the turnout was recorded only in species that require a higher level of trophy: Alona rectangula and Chydorus sphaericus (Duigan 1992).
Phases of water level increase were recorded at about 400 to 375 cm. The first raise of water level illustrates the reduction of the area occupied by the marsh fern and documents the decline of T. palustris spores, which is accompanied by increased frequency of macrophytes. At the same time, there was increase in the growth in the reservoir due to the supply of nutrients as well as the supply of mineral materials from the basin (Phase II).
The increase of water level was also recorded by an increase in the attendance of Cladocera. At the depth of 402.5 cm, highest turnouts of the Cladocera individuals in the reservoir (1425 specimens per 1 cm3 of sediments) indicated favorable conditions for the development in this period. The highest turnout of Alona quadrangularis species indicated a higher water level and the increase of growth in the reservoir (Błędzki and Rybak 2016). There were also noted species associated with macrophytes zone such as Alona affinis, Alona rectangula, and Sida crystallina (Flössner 2000). A slight increase of water level at the depth of 387.5 cm may also be indicated by a small proportion of the Daphnia longispina species from the open water zone. The increase of the water level at a depth of 375 cm was documented by the increase in the attendance of Alona affinis species associated with the macrophyte zone (Nevalainen 2012).
Above a depth of 372.5 cm, the significant drop in the attendance of Cladocera may indicate the deterioration of living conditions in the reservoir and its gradual terrestrialization. During this period of approximately 9530 cal. year BP (7660 cal. year BC), the presence of spores of the fungus Glomus in sediments confirms strongly influenced erosion processes leading to enhanced accumulation of sediments.
3.3 Physical properties of the soils
Physical properties of the studied soils are typical for sandy and peaty soils (Fig. 4). Bulk density ranged from 0.23–0.75 g cm−3 in organic horizons to 1.00–1.59 g cm−3 in mineral ones. The low content of water is characteristic of sandy aeolian materials poor in fine granulometric fractions. In deeper zones of the studied profile, increases at several times within were correlated with the enhanced content of silt and clay granulometric fractions. Total porosity of mineral horizons varies between 34.8 and 58.4%, while in organic, these values are averagely about twice as high. Very high porosity is also observed in 3Ab horizon, which is much more abundant in TOC than remaining A-horizons. The lack of indigenous mineral sediments within the fossil peat soil profile shows that during the peat formation, there were no intensive geomorphological processes. Organic material of the Dystric Fibric Histosol was compacted, which was reflected in relatively high bulk density.

Profile variability of selected physical properties in the soil sequence under study
3.4 Chemical properties of the soils
Contemporary soils represented by two profiles of Dystric Arenosols are much poorer in nutrients than the fossil soils (Table 6). Depending on the nutrient content in the whole, pedocomplex may be divided into three zones: (1) with low nutrients content (mineral material to the depth of 124 cm), (2) with increased nutrients content (mineral material to the depth of 370 cm), and (3) with very high content of nutrients (organic material at the depth of 370–415 cm). Among the presented elements, the largest difference in the content was concerned with the TOC, TN, and partially P. The content of TOC in the buried A-horizons of Dystric Arenosols (Protospodic) reaches values from 9.71 to 46.91 g kg−1, TN 1.28–2.17 g kg−1, and P 0.28–0.55 g kg−1, which forms the evidence of these soil formation during periods of relatively stable and warm climate conditions and continuous supply of organic material (Sifeddine et al. 1996; Macaire et al. 2006). Profile distribution of K, Ca, and Mg also refers to the presented schema but is characterized by less variation in the content of these elements between particular profiles and gradual increase along with the depth (especially in the case of K). Soil biological activity index expressed by TOC/TN ratio also has increased in deeper located fossil soils, achieving the highest values in A-horizons and 7Oi horizon (Table 6). The reaction of the soils ranged from extremely acidic to moderately acidic. The increase of soil pH is observed from the top of the polycyclic pedocomplex to the 5Bhs horizon. In Dystric Fluvisol (Relictiturbic) and Dystric Fibric Histosol profiles, soil pH decreases to 3.81 (pHH2O) and 3.59 (pHKCl) at the depth of 405–410 cm.
3.5 Advancement of soil-forming processes
The role of iron in the course of soil forming processes is inalienable and depends on many factors, mainly, parent material properties, climatic conditions, and plant cover. Most of the studies were focused on iron content and profile distribution in mineral soils. Meanwhile, there are still only few studies on this issue with reference to organic soils (Norrstrom 1995). The content of Fet is relatively low in two profiles of Dystric Arenosols and ranges from 4.50 to 6.18 g kg−1 (Table 7). For these young soils, the high content of Fegk in comparison with Fed, which is also characteristic, was determined by the maritime microclimate and intensive weathering processes. After soil substrate weathering, forms of Fegk do not move into the deeper parts of profile (Karltun et al. 2000; Degórski 2002). In older soils, especially in most layers of Dystric Fibric Histosol, this dependency is reciprocal and the value of Fed is much higher than the content of Fegk (Konecka-Betley 1968; Kuźnicki and Skłodowski 1970). Within this organic soil profile, it is possible to recognize advancement of different phases of soil forming processes with the predominance of Fed on Fegk content. Increased Feo content in humus horizons is the evidence of formation of iron–humic complexes. Simultaneously, the content of Feo decreases with depth. An exception is the organic soil profile in which the content of Feo ranges from 4.84 g kg−1 at the primary surface to 18.61 g kg−1 at the depth of 405–420 cm. In deeper parts of Dystric Fibric Histosol, the content of Fet is low (12.33 g kg−1) and values of Fed, Feo, and Fegk are relatively aligned, which is characteristic of young glacial areas. From the depth of 410 cm to the top of organic soil profile, increased content of Fet is observed, which can be associated with the influx of iron-rich water. The degree of soil material weathering is high in A and B horizons of Dystric Arenosols (Protospodic) (at the depths of 124–155 and 210–233 cm). However, the highest values are observed within the Dystric Fibric Histosol.
4 Conclusions
The investigated soils constitute relicts of the former geomorphological and soil forming processes, which has been taking place almost over the last 9500 years. The polycyclic pedocomplex of interest consists of seven soils showing different degrees of development. Additionally, traces of a palaeolake were found in the profile. The palaeolake was directly related to the gradual disappearance of the ice sheet from the study area and melting of the dead ice. The genetic similarity of the areas of the southern coast of the Baltic Sea allows for concluding that there are more such palaeolakes in this zone. The bottom part of the polycyclic pedocomplex, occupied by a Dystric Fibric Histosol profile, represents a period directly related to the existence of the palaeolake with the main phase of organic material accumulation as a basal peat. Intensification of erosion processes related to climatic conditions changes in this region in the Boreal period is clearly visible in the pedocomplex of interest as inhibition of the peat-forming phase and the presence of spores of Glomus. The radiocarbon dates of the top of peat soil coincide with the literature data confirming that the significant reduction in average temperatures and humidity characteristic for the Boreal period had an impact on soil formation of the Baltic coastal zone. The finding of a predominance of Betula and Pinus sylvestris pollen in samples collected from peat soil confirms the thesis about the prevalence of these species in the Preboreal period. The presence of Thelypteris palustris in the Dystric Fibric Histosol can be a sign of succession in conditions of strong flooding and inflow of moderately fertile water, which is confirmed by the properties of the peat soil investigated. According to the results of pollen and fossil Cladocera analysis and radiocarbon dating, it can be concluded that the studied reservoir has evolved from the beginning of the Preboreal period to the half of Boreal period and has been shaping as an effect of the impact of permafrost degradation. Ultimately, it was buried by the fluvial material due to the effect of fire, whose occurrence was confirmed by charcoals found at the top of the Dystric Fibric Histosol. The top part of these sediments were subjected to the influence of periglacial conditions, which has been marked in the Dystric Fluvisol (Relictiturbic) as layers disrupted by freezing and thawing processes. The next stages of the relief development of the studied area were closely associated with changes of Baltic Sea shoreline, lowering of the ground water level and increased intensity of aeolian processes, resulted in the formation of dunes and indirectly led to Dystric Arenosols (Protospodic) profile development. The sea water has been encroaching deeper in the land, covering mainly terrestrial ecosystems. After covering the organic material by mineral colluvial deposits, humidity conditions were still better than present days and promoted the development of complete soil profiles with organic matter-rich horizons. The fire which took place 1472–1637 cal. year AD began the last stage of the coastal relief dynamics. The lack of plant cover and intensive abrasion due to direct vicinity of the coastline caused the accumulation of aeolian deposits that were the parent material of initial soils with very low nutrient content. The direct result of the aeolian processes, whose intensity depends additionally on the research area location in the direct vicinity of Baltic Sea, was deposition of a poor substrate to soil formation.
Our research has shown a unique nature of pedogenesis in sea coastal zones. It is due to the influence of many external factors including inter alia the sea vicinity, which has a direct impact on the continual soil rejuvenation through the influence of the aeolian processes and storms. Considering the similarity of lithological and environmental permit with geological history of this part of Baltic coastal zone, it can be concluded that our findings can be successfully used as a basis for the reconstruction of former soil cover of other parts of the Baltic coast.
References
de Alba S, Lindstrom M, Schumacher TE, Malo DD (2004) Soil landscape evolution due to soil redistribution by tillage: a new conceptual model of soil catena evolution in agricultural landscapes. Catena 58:77–100
Barthelmes A, de Klerk P, Prager A, Theuerkauf M, Unterseher M, Joosten H (2012) Expanding NPP analysis to eutrophic and forested sites: significance of NPPs in Holocene wood peat section (NE Germany). Rev Palaeobot Palyn 186:22–37
Bednarek R (1979) Gleby wybranych odcinków klifowych polskiego wybrzeża Bałtyku, Seria Sectio D (Botanica), vol. X(6) (in Polish)
Beug HJ (2004) Leitfagen der Pollenbestimmung für Mitteleuropa und angrenzende Gebiete. Verlag Dr. Friedrich Pfeil. München
Błaszkiewicz M (2005) Późnoglacjalna i wczesnoholoceńska ewolucja obniżeń jeziornych na Pojezierzu Kociewskim (wschodnia część Pomorza). Prace Geograficzne 201:11–92 (in Polish)
Błaszkiewicz M (2008) Wytapianie się pogrzebanych brył martwego lodu w późnym glacjale i wczesnym holocenie a zdarzenia ekstermalne. Landform Analysis 8:9–12 (in Polish)
Błędzki LA, Rybak JI (2016) Freshwater crustacean zooplankton of Europe Cladocera & Copepoda (Calanoida, Cyclopoida) key to species identification, with notes on ecology, distribution, methods and introduction to data analysis. Springer International Publishing, AG, p 933
Blott SJ, Pye K (2001) Gradistat: a grain size distribution and statistics package for the analysis of unconsolidated sediments. Earth Surf Process Land 26(11):1237–1248
Bogaart PW, Tucker GE, de Vries JJ (2003) Channel network morphology and sediment dynamics under alternating periglacial and temperate regimes: a numerical simulation study. Geomorphology 54:257–277
Bronk Ramsey C (2013) OxCal. Version 4.2.3, Oxford Radiocarbon Accelerator Unit Research Laboratory for Archaeology, Oxford
Degórski M (2002) Przestrzenna zmienność właściwości gleb bielicoziemnych środkowej i północnej Europy a geograficzne zróżnicowanie czynników pedogenicznych, Prace Geograficzne, IGiPZ PAN, P 182 (summary in English)
Duigan CA (1992) The ecology and distribution of the littoral freshwater Chydoridae (Branchiopoda, Anomopoda) of Ireland with taxonomic comments on some species. Hydrobiol 241:1–70
Dybova-Jachowicz S, Sadowska A (2003) Palinologia. Wyd. Instytut Botaniki PAN, Kraków, p 411
Dziadowiec H, Gonet S (1999) Przewodnik metodyczny do badań materii organicznej gleb. Warszawa: Prace Komisji Nauk PTG 120:66
Engstrom DR, Wright HE Jr (1984) Chemical stratigraphy of lake sediments as a record of environmental change. In: Haworth EY, Lund JWG (eds) Lake sediments and environmental history. Leicester University Press
Faegri K, Iversen J (1989) Textbook of pollen analysis, IV edn. J Wiley & Sons Ltd., Chichester-Singapore
Finsinger W, Tinner W, van der Knaap WO, Ammann B (2006) The expansion of hazel (Corylus avellana L.) in the southern Alps: a key for understanding its early Holocene history in Europe? Quat Sci Rev 25:612–631
Florek W, Kaczmarzyk J, Majewski M, Olszak IJ (2008) Zmiany rzeźby klifu w rejonie Ustki jako efekt warunków litologicznych oraz procesów ekstremalnych i przeciętnych, Landform analysis 7:53–68 (abstract in English)
Florek W, Kaczmarzyk J, Majewski M (2010) Dynamics of the polish coast east of Ustka. Geogr Pol 83(1):51–60
Flössner D (1972) Krebstiere, Crustacea. Kiemen- und Blattfüsser, Branchiopoda, Fischläuse, Branhiura. Tierwelt Deutschlands 60:1–501
Flössner D (2000) Die Haplopoda und Cladocera (ohne Bosminidae) Mitteleuropas. Backhuys Publishers, Leiden, p 428
Folk RL, Ward W (1957) Brazos River bar: a study in the significance of grain size parameters. J Sediment Petrol 27:3–26
Frey DG (1986) Cladocera analysis. In: Berglund BE (ed) Handbook of Holocene Palaeoecology and Palaeohydrology, pp 667–692
Gadziszewska J (2015) The role of natural and anthropogenic factors in postglacial environmental changes in Wrześnica Area PhD thesis, Univeristy of Gdańsk
van Geel B (1978) A palaeoecological study of Holocene peat bog sections in Germany and the Netherlands. Rev Palaeobot Palynol 25:1–120
van Geel B, Bohncke SJP, Dee H (1981) A palaeoecological study of an upper late glacial and Holocene sequence from “De Borchert”, the Netherlands. Rev Palaeobot Palynol 31:367–448
van Geel B, Coope GR, van der Hammen T (1989) Palaeoecology and stratigraphy of the Lateglacial type section at Usselo (the Netherlands). Rev Palaeobot Palynol 60:25–129
Giesecke T, Bennett KD, Birks HJB, Bjune AE, Bozilova E, Feurdean A, Finsinger W, Froyd C, Pokorný P, Rösch M, Seppä H, Tonkov S, Valsecchi V, Wolters S (2011) The pace of Holocene vegetation change—testing for synchronous developments. Quat Sci Rev 30:2805–2814
Goulden CE (1964) The history of the cladoceran fauna of Esthwaite water (England) and its limnological significance. Arch Hydrobiol 60:1–53
Hofmann W (1986) Developmental history of the Grosser Plöner See and the Schöhsee (North Germany): cladoceran analysis with special reference to eutrophication. Arch Hydrobiol Suppl 74:259–287
Hofmann W (2000) Response of the chydorid faunas to rapid climatic changes in four alpine lakes at different altitudes. Palaeogeogr Palaeoclimatol Palaeoecol 159(3–4):281–292
Holliday VT (1992) Soil formation, time and archaeology. In: Holliday VT (ed) Soils in Archaeology: Landscape Evolution and Human Occupation. Smithsonian Institution Press, Washington, DC, pp 107–117
Hošek J, Pokorný P, Prach J, Lisá L, Grygar TM, Knésl I, Trubač J (2017) Late glacial erosion and pedogenesis dynamics: evidence from high-resolution lacustrine archives and paleosols in south Bohemia (Czech Republic). Catena 150:261–278
Isarin RFB, Bohncke SJP (1999) Mean July temperatures during the younger Dryas in northwestern and Central Europe as inferred from climate Indicator plant species. Quat Res 51(2):158–173
IUSS Working Group WRB (2015) World Reference Base for soil resources 2014, update 2015. International soil classification system for naming soils and creating legends for soil maps. World soil resources report no. 106 FAO, Rome
Karltun E, Bain DC, Gustafsson JP, Mannerkoski H, Murad E, Wagner U, Fraser AR, McHardy WJ, Starr M (2000) Surface reactivity of poorly-ordered minerals in podzol B horizons. Geoderma 94:265–288
Konecka-Betley K (1968) Zagadnienie żelaza w procesie glebotwórczym. Rocz Glebozn 19(1):51–97 (summary in English)
Korhola A (1990) Paleolimnology and hydroseral development of the Kotasuo bog, southern Finland, with special reference to the Cladocera. Ann Acad Sci Fenn 155:5–40
Korhola A, Rautio M (2001) Cladocera and other branchiopod crustaceans. In: Smol JP, Birks HJB, Last WM (eds) Tracking environmental change using Lake sediments, zoological indicators 4. Kluwer Academic Publishers, Dordrecht, pp 5–41
Kozarski S (1995) Deglacjacja północno-zachodniej Polski: warunki środowiska i transformacja geosystemu (∼20 ka → ∼10 ka BP). Dokumentacja Geograficzna 1, IGiPZ PAN, Warszawa (in Polish)
Kruczkowska B, Jonczak J, Gadziszewska J (2017) Characteristics of permafrost-affected soil in a cliff in the central part of the polish Baltic coastal zone. J Soils Sediments 17(4):960–973
Krzyszkowski D, Dobracka E, Dobracki R, Czerwonka J, Kuszell T (1999) Stratigraphy of Weichselian deposits in the cliff sections between Łukęcin and Niechorze, Baltic Coast, Northwestern Poland. Quatern Stud Pol 16:27–45
Kuhry P (1997) The palaeoecology of a treed bog in western boreal Canada: a study based on microfossils, macrofossils and physico-chemical properties. Rev Palaeobot Palynol 96:183–224
Kuźnicki F, Skłodowski P (1970) Zawartość w glebie wolnego żelaza, wolnego glinu i wolnej krzemionki jako kryterium typologiczne. Rocz Glebozn 21(1):13–19 (summary in English)
Macaire JJ, Bernard J, Di-Giovanni C, Hinschberger F, Limondin-Lozouet N, Visset L (2006) Quantification and regulation of organic and mineral sedimentation in a late Holocene floodplain as a results of climatic and human impacts. The Holocene 16(5):647–660
Mehra O, Jackson J (1960) Iron oxide removal from soils and clays by a dithionite-citrate system buffered with sodium bicarbonate. Clay Clay Miner 5:317–327
Nalepka D, Walanus A (2003) Data processing in pollen analysis. Acta Palaeobot 43(1):125–134
Nevalainen L (2012) Distribution of benthic microcrustaceans along a water depth gradient in an Austrian Alpine lake—sedimentary evidence for niche separation. Limnologica 42:65–71
Norrstrom AC (1995) Concentration and chemical species of iron in soils from groundwater surface water ecotones. Hydrol Sci 40(3):319–329
Olszak IJ, Florek W, Seul C, Majewski M (2008) Stratigraphy and lithology of minerogenic deposits in coastal cliffs, middle section of the Polish Baltic coast. Landform Analysis 7:113–118 (abstract in English)
Olszak IJ, Florek W, Seul C, Majewski M (2011) Lithology of sediments and stratigraphy of some cliff sections of central and western Polish Coast. Geologija 53(1(73)):1–9
Polish Soil Science Society (PSSS) (2009) Klasyfikacja uziarnienia gleb i utworów mineralnych, PTG 2008. Rocz Glebozn 60(2):5–17 (in Polish)
Ralska-Jasiewiczowa M, Goslar T, Madeyska T, Starkel L (eds) (1998) Lake Gościąż, Central Poland. A Monographic Study. W. Szafer Institute of Botany, Kraków (Part I)
Ralska-Jasiewiczowa M, Tobolski K, Nalepka D (2004) Typha latifolia L. –Bulrush, cat-tail. In: Ralska-Jasiewiczowa M, Latałowa M, Wasylikowa K, Tobolski K, Madeyska T, Wright HE Jr, Turner Ch (eds) Late Glacial and Holocene history of vegetation in Poland based on isopollen maps. W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków, pp 359–369
Reimer PJ, Bard E, Bayliss A (2013) IntCal13 and marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 55:1869–1887
Sifeddine A, Bertrand P, Laillier-Vergcs E, Patience AJ (1996) Lacustrine organic fluxes and paleoclimatic variations during the last 15 ka: Lac du Bouchet (Massif Central, France). Quat Sci Rev 15:203–211
Szeroczyńska K (1985) Cladocera jako wskaźnik ekologiczny w późnoczwartorzędowych osadach jeziornych Polski Północnej (Cladocera as ecologic indicator in late quaternary lacustrine sediments in northern Poland). Acta Palaeontol Polon 30(1–2):3–69
Szeroczyńska K (1998) Wioślarki (Cladocera, Crustacea) jako źródło informacji w badaniach osadów jeziornych. [Cladocerans (Cladocera, Crustacea) as a reference in lake sediment studies]. Studia Geologica Polonica 112:9–28 (in Polish)
Szeroczyńska K, Sarmaja-Korjonen K (2007) Atlas of subfossil Cladocera from central and northern Europe. Friends of Lower Vistula Society Świecie, pp 87
Targulian VO, Goryachkin SV (2004) Soil memory: types of record, carriers, hierarchy and diversity. Rev Mex Cienc Geol 21:1–8
Targulian VO, Sokolova TA (1996) Soil as a bio-abiotic natural system; a reactor, memory and regulator of biospheric interactions. Eurasian Soil Sci 29(1):34–47
Targulian VO, Krasilnikov PV (2007) Soil system and pedogenic processes: self-organization, time scales and environmental significance, Catena 71: 373–381
Tsara M, Gerontidis S, Marathianou M, Kosmas C (2001) The long-term effect of tillage on soil displacement of hilly areas used forgrowing wheat in Greece. Soil Use Manag 17:113–120
Uścinowicz SZ (2003) The southern Baltic relative sea level changes, glacio-isostatic rebound and shoreline displacement. Polish Geological Institute Special Papers 10:1–79
Uścinowicz SZ (2006) A relative sea-level curve for the polish southern Baltic Sea. Quaternary Int 145/146:86–105
Van Loon A, Błaszkiewicz M, Degórski M (2012) The role of permafrost in shaping the late glacial relief of northern Poland. Neth J Geosci Geol Mijnb 91:223–231
Van Reeuvijk LP (2002) Procedures for soil analysis. Sixth edition. ISRiC, FAO, Wageningen
Wróblewski R, Fedorowicz S, Kamieńska K (2013) Holocene sediments of the Ustka cliff (northern Poland) in view of radiometric dating. Geochronometria 40(3):187–194
Yetemen O, Istanbulluoglu E, Vivoni ER (2010) The implications of geology, soils and vegetation on landscape morphology: inferences from semi-arid basins with complex vegetation patterns in Central New Mexico, USA. Geomorphology 116:246–263
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Arnaud Temme
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Kruczkowska, B., Jonczak, J., Gadziszewska, J. et al. The record of postglacial environmental changes of the southern Baltic coastal zone in the sequence of fossil soils. J Soils Sediments 19, 848–861 (2019). https://doi.org/10.1007/s11368-018-2058-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-018-2058-4